Developmental Stage- and Genotype-Dependent Regulation of Specialized Metabolite Accumulation in Fruit Tissues of Different Citrus Varieties


Abstract
:
1. Introduction
2. Results and Discussion
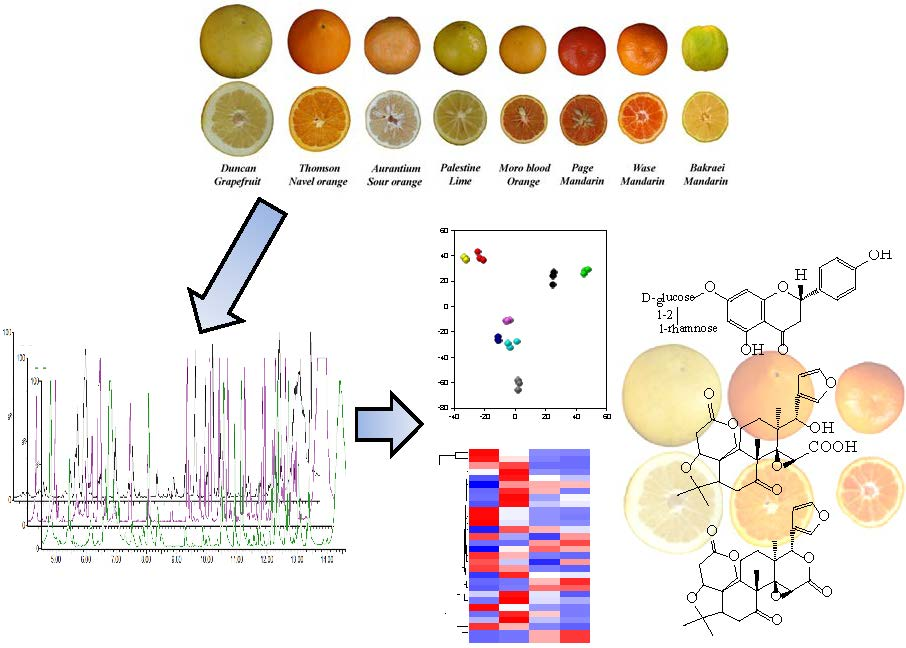
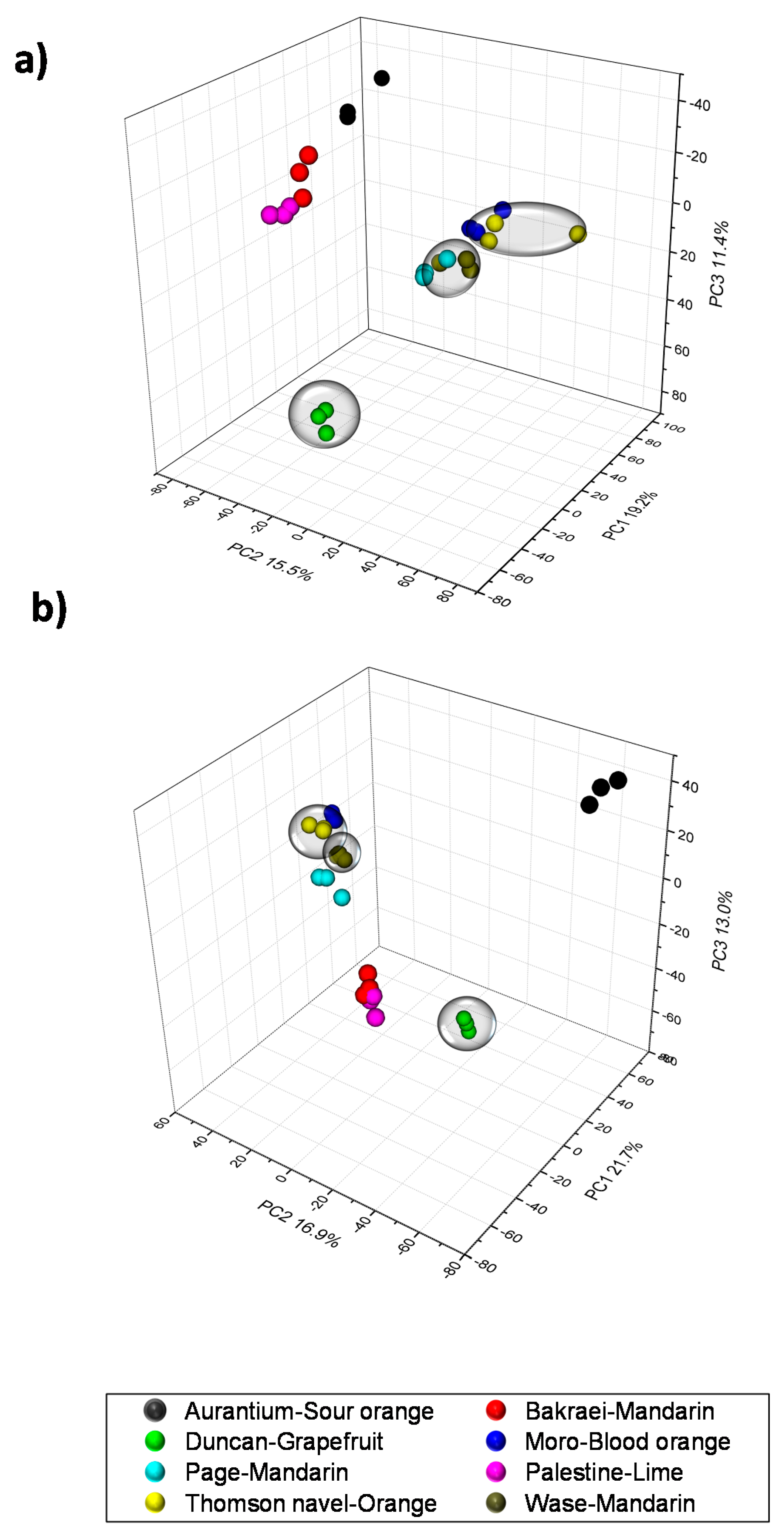
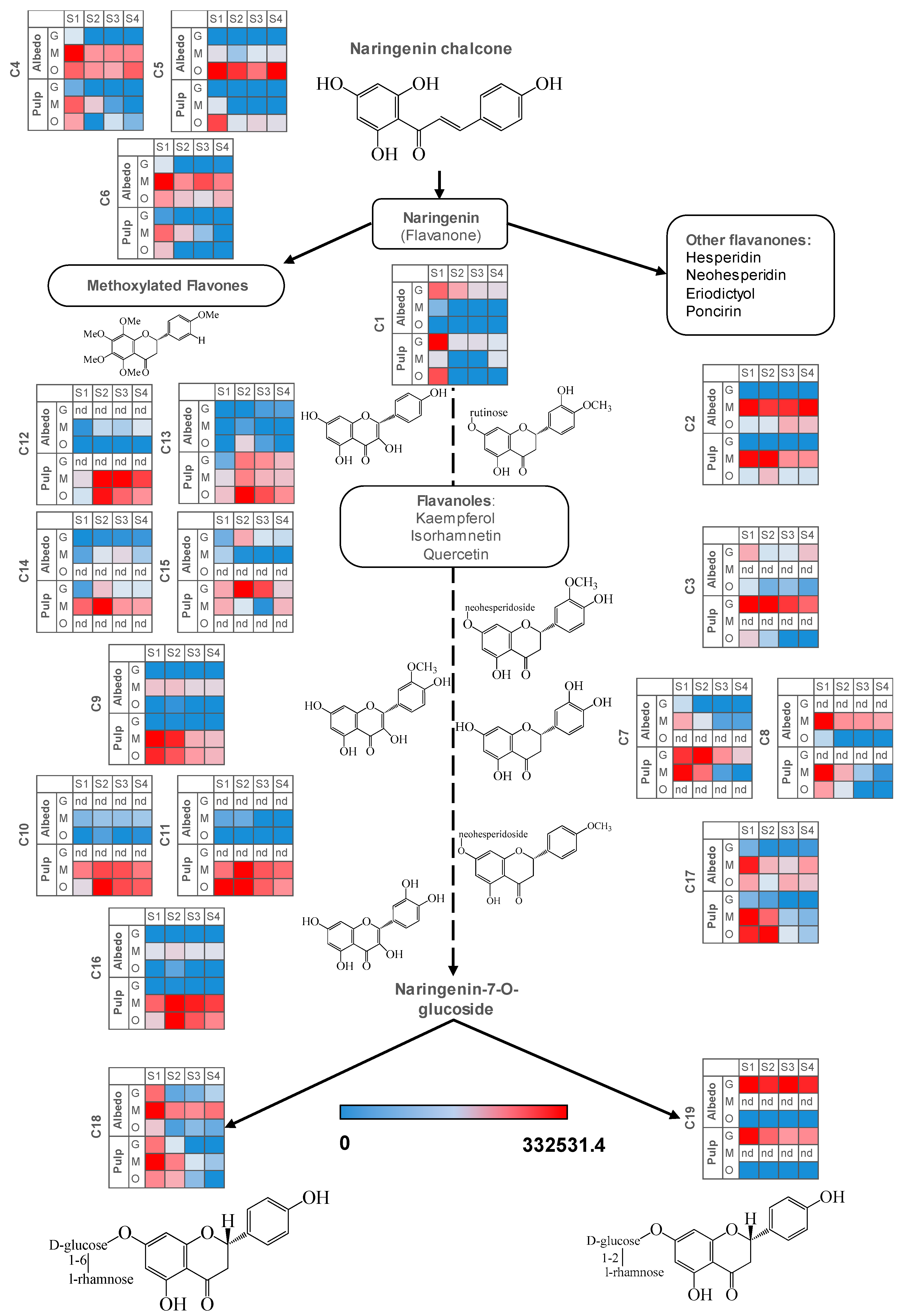
2.1. Validation of Results, Variable Selection and Annotation of Compounds
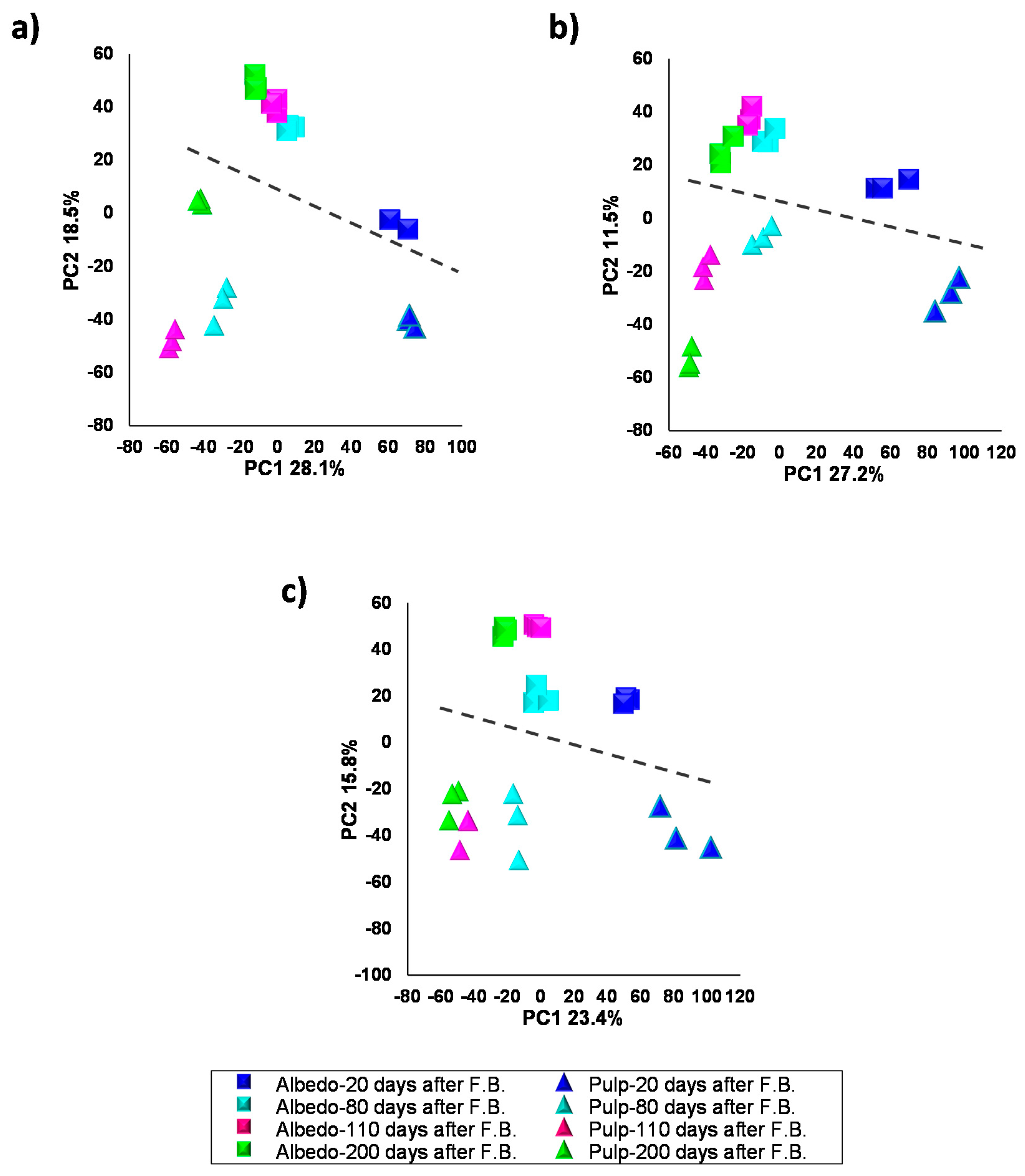
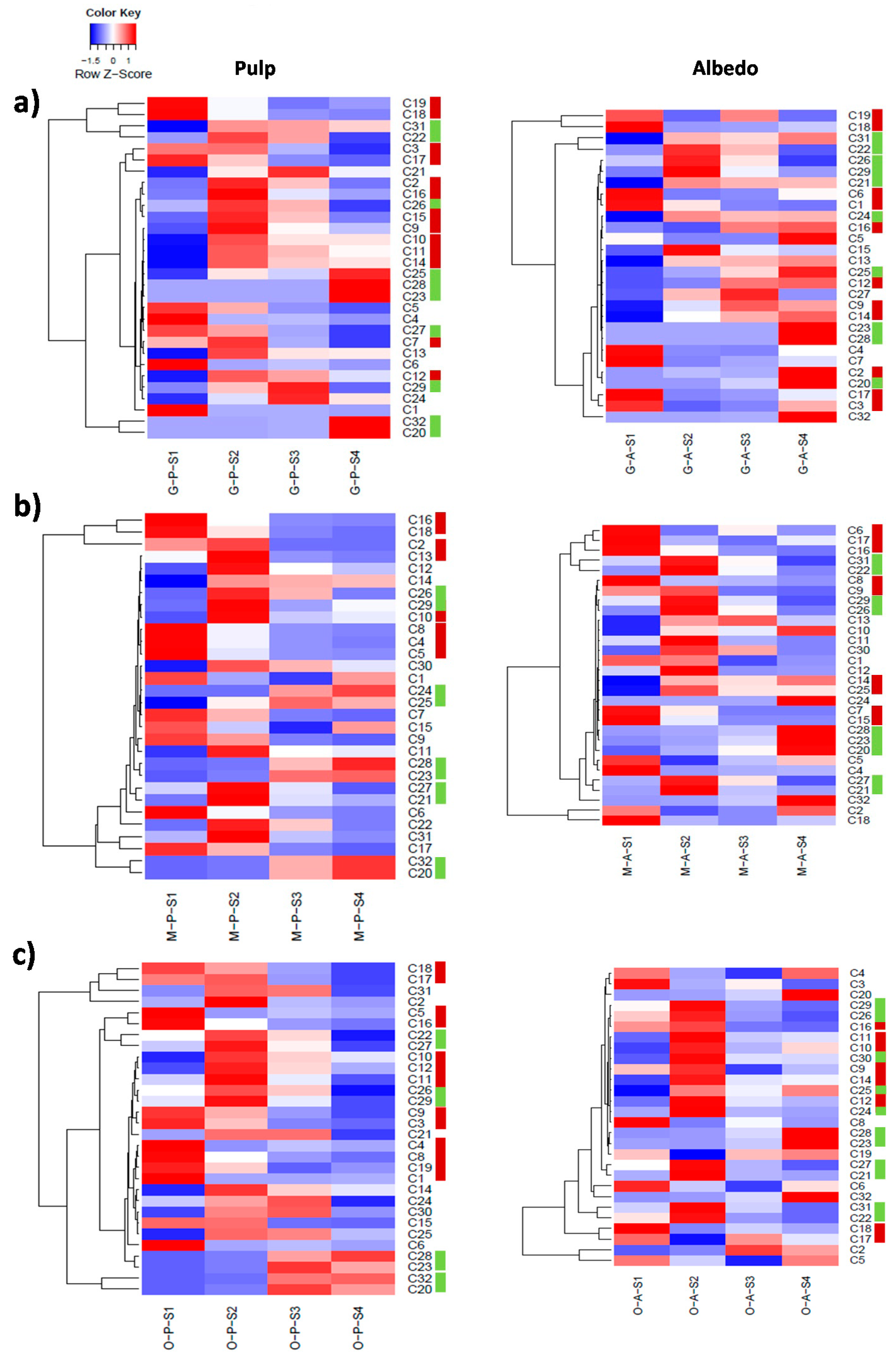
2.2. Characterization of Metabolite Profiles in Albedo and Pulp Tissues
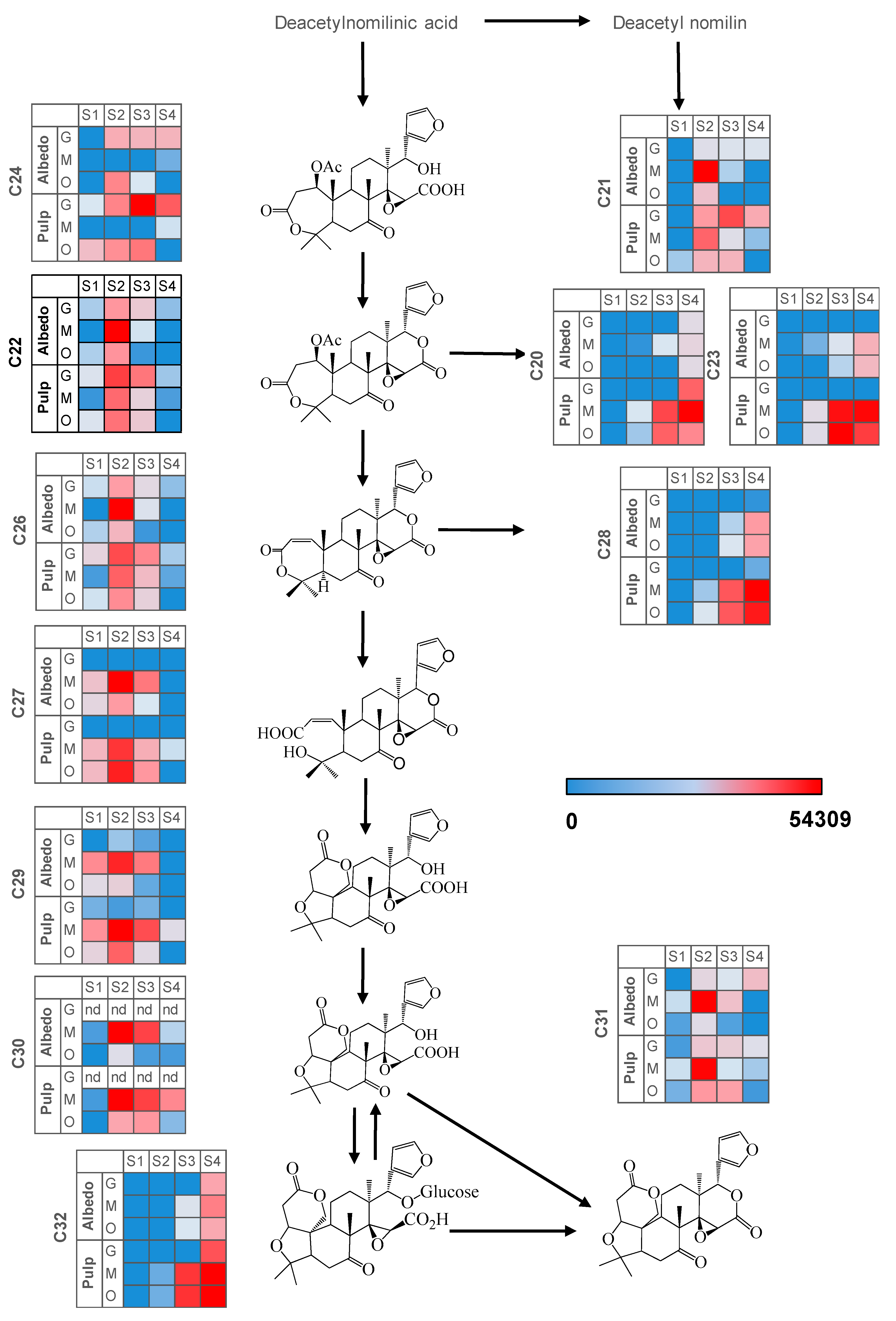
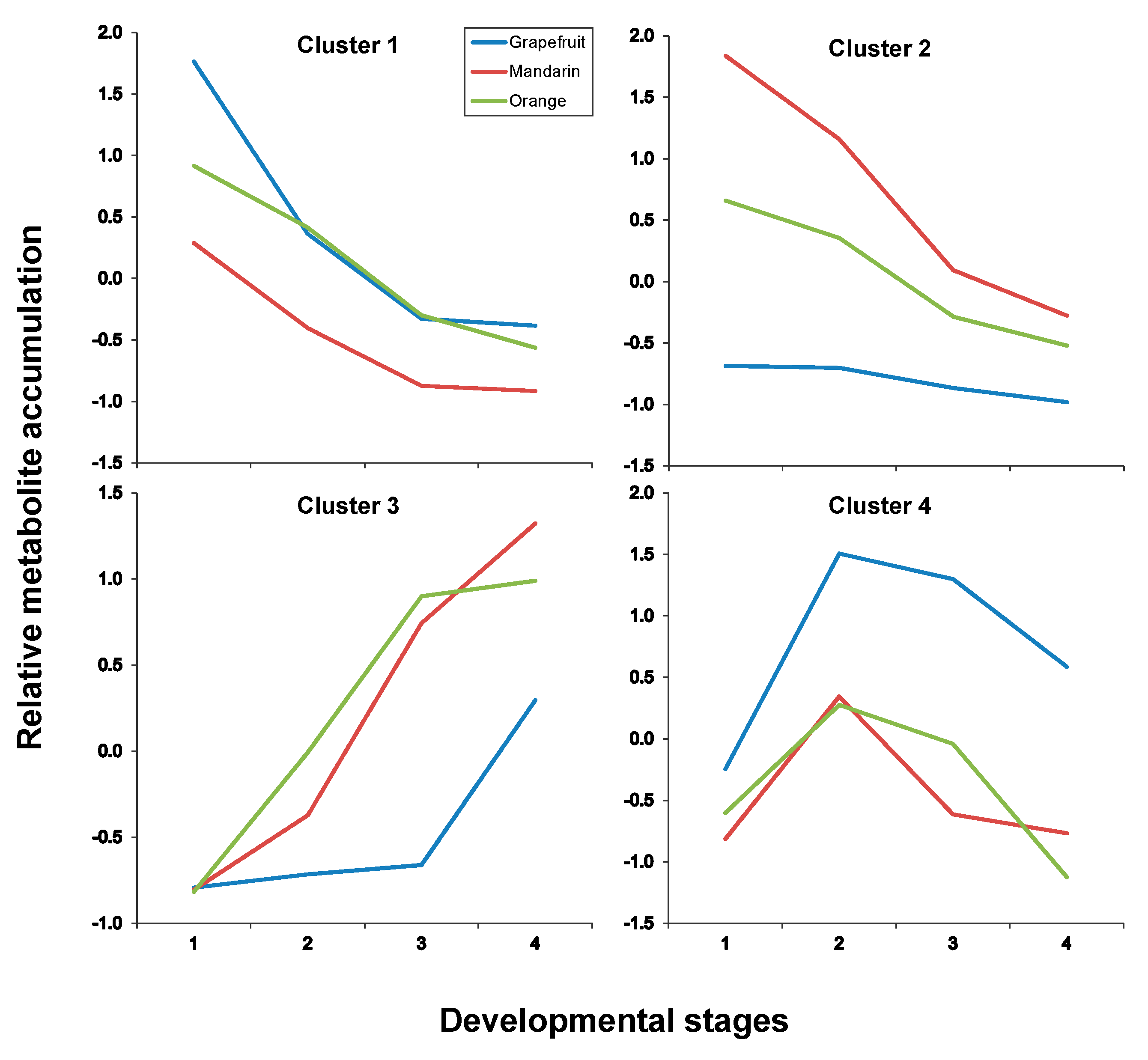
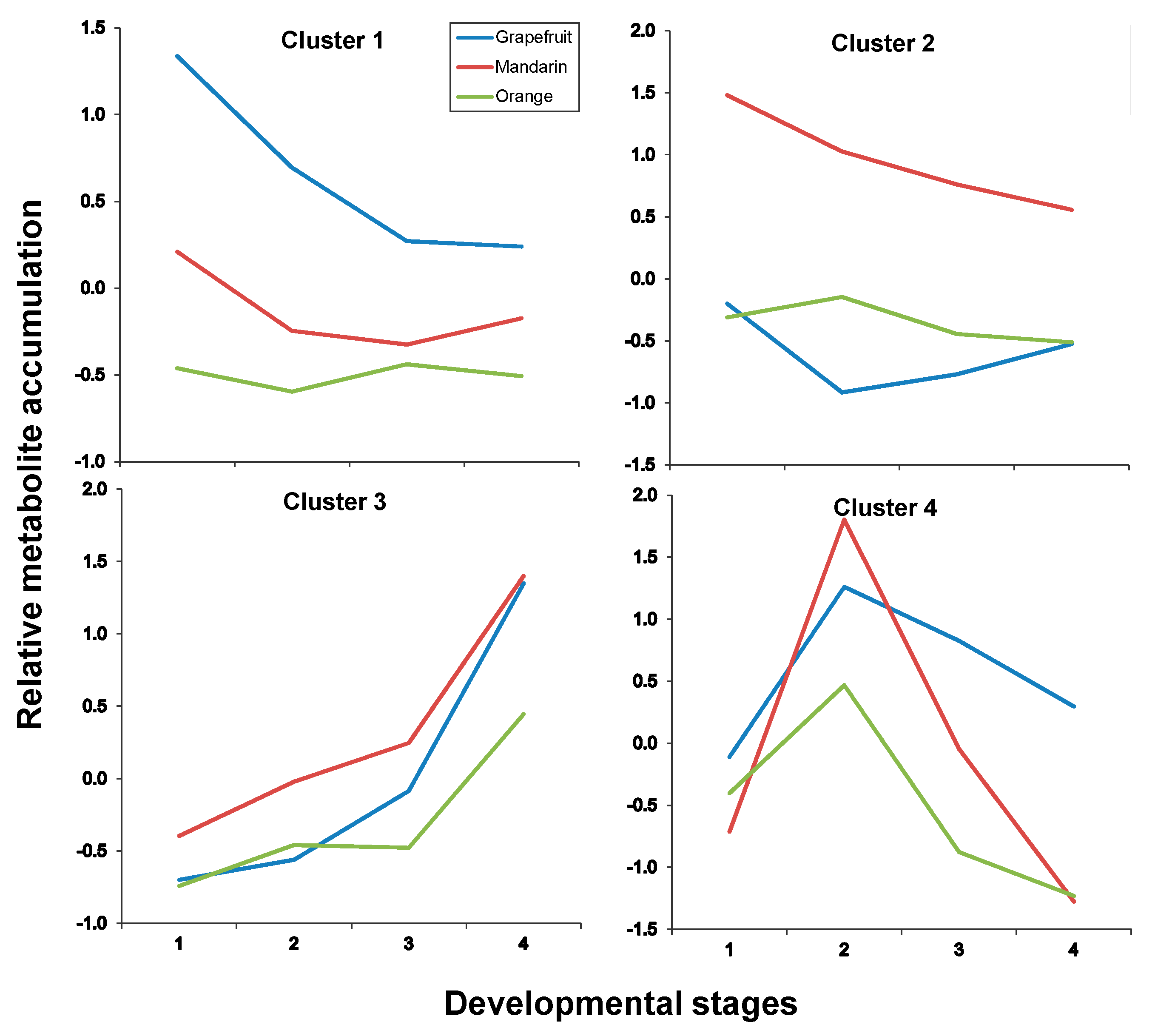
2.3. Fruit Ripening: Co-Regulation of Specialized Metabolites in Citrus Fruit Tissues
3. Materials and Methods
3.1. Plant Material and Sample Preparation
3.2. Extraction of Samples for Chromatographic Analyses
3.3. Chromatographic and QqTOF-MS Conditions
3.4. Mass Spectrometry (MS) Data Processing, Statistical Analyses and Compound Identification
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Terol, J.; Soler, G.; Talon, M.; Cercos, M. The aconitate hydratase family from Citrus. BMC Plant Biol. 2010, 10, 222. [Google Scholar] [CrossRef]
- Dugrand-Judek, A.; Olry, A.; Hehn, A.; Costantino, G.; Ollitrault, P.; Froelicher, Y.; Bourgaud, F. The distribution of coumarins and furanocoumarins in Citrus species closely matches Citrus phylogeny and reflects the organization of biosynthetic pathways. PLoS ONE 2015, 10, e0142757. [Google Scholar] [CrossRef]
- Patil, B.S.; Jayaprakasha, G.K.; Chidambara Murthy, K.N.; Vikram, A. Bioactive compounds: Historical perspectives, opportunities and challenges. J. Agric. Food Chem. 2009, 57, 8142–8160. [Google Scholar] [CrossRef]
- Manners, G.D. Citrus limonoids: Analysis, bioactivity, and biomedical prospects. J. Agric. Food Chem. 2007, 55, 8285–8294. [Google Scholar] [CrossRef]
- Frydman, A.; Liberman, R.; Huhman, D.V.; Carmeli-Weissberg, M.; Sapir-Mir, M.; Ophir, R.; W Sumner, L.; Eyal, Y. The molecular and enzymatic basis of bitter/non-bitter flavor of citrus fruit: Evolution of branch-forming rhamnosyltransferases under domestication. Plant J. 2012, 73, 166–178. [Google Scholar] [CrossRef]
- Wang, F.; Wang, M.; Liu, X.; Xu, Y.; Zhu, S.; Shen, W.; Zhao, X. Identification of putative genes involved in limonoids biosynthesis in citrus by comparative transcriptomic analysis. Front. Plant Sci. 2017, 8, 782. [Google Scholar] [CrossRef]
- Akyildiz, A.; Erdal, A. Food Processing: Strategies for Quality Assessment; Malik, A., Erginkaya, Z., Ahmad, S., Erten, H., Eds.; Food Engineering Series; Springer: New York, NY, USA, 2014; ISBN 978-1-4939-1377-0. [Google Scholar]
- Dala Paula, B.M.; Raithore, S.; Manthey, J.A.; Baldwin, E.A.; Bai, J.; Zhao, W.; Glória, M.B.A.; Plotto, A. Active taste compounds in juice from oranges symptomatic for Huanglongbing (HLB) citrus greening disease. LWT Food Sci. Technol. 2018, 91, 518–525. [Google Scholar] [CrossRef]
- Zaare-Nahandi, F.; Hosseinkhani, S.; Zamani, Z.; Asadi-Abkenar, A.; Omidbaigi, R. Delay expression of limonoid UDP-glucosyltransferase makes delayed bitterness in citrus. Biochem. Biophys. Res. Commun. 2008, 371, 59–62. [Google Scholar] [CrossRef]
- Ververidis, F.; Trantas, E.; Douglas, C.; Vollmer, G.; Kretzschmar, G.; Panopoulos, N. Biotechnology of flavonoids and other phenylpropanoid-derived natural products. Part I: Chemical diversity, impacts on plant biology and human health. Biotechnol. J. 2007, 2, 1214–1234. [Google Scholar] [CrossRef]
- Wang, S.; Tu, H.; Wan, J.; Chen, W.; Liu, X.; Luo, J.; Xu, J.; Zhang, H. Spatio-temporal distribution and natural variation of metabolites in citrus fruits. Food Chem. 2016, 199, 8–17. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Menichini, F. An overview on chemical aspects and potential health benefits of limonoids and their derivatives. Crit. Rev. Food Sci. Nutr. 2014, 54, 225–250. [Google Scholar] [CrossRef]
- Drewnowski, A.; Gomez-Carneros, C. Bitter taste, phytonutrients, and the consumer: A review 1–3. Am. J. Clin. Nutr. 2000, 72, 1424–1435. [Google Scholar] [CrossRef]
- Arbona, V.; Iglesias, D.J.; Talón, M.; Gómez-Cadenas, A. Plant phenotype demarcation using nontargeted LC-MS and GC-MS metabolite profiling. J. Agric. Food Chem. 2009, 57, 7338–7347. [Google Scholar] [CrossRef]
- Arbona, V.; Iglesias, D.J.; Gómez-Cadenas, A. Non-targeted metabolite profiling of citrus juices as a tool for variety discrimination and metabolite flow analysis. BMC Plant Biol. 2015, 15, 38. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Sales, C.; Beltrán, J.; Gómez-Cadenas, A.; Arbona, V. Activation of secondary metabolism in citrus plants is associated to sensitivity to combined drought and high temperatures. Front. Plant Sci. 2016, 7, 1954. [Google Scholar] [CrossRef]
- Durand-Hulak, M.; Dugrand, A.; Duval, T.; Bidel, L.P.R.; Jay-Allemand, C.; Froelicher, Y.; Bourgaud, F.; Fanciullino, A.L. Mapping the genetic and tissular diversity of 64 phenolic compounds in Citrus species using a UPLC-MS approach. Ann. Bot. 2015, 115, 861–877. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Xia, R. Expression of chalcone synthase and chalcone isomerase genes and accumulation of corresponding flavonoids during fruit maturation of Guoqing No. 4 satsuma mandarin (Citrus unshiu Marcow). Sci. Hortic. (Amsterdam) 2010, 125, 110–116. [Google Scholar] [CrossRef]
- Barreca, D.; Bellocco, E.; Caristi, C.; Leuzzi, U.; Gattuso, G. Flavonoid composition and antioxidant activity of juices from Chinotto (Citrus x myrtifolia Raf.) fruits at different ripening stages. J. Agric. Food Chem. 2010, 58, 3031–3036. [Google Scholar] [CrossRef]
- Chaudhary, P.R.; Bang, H.; Jayaprakasha, G.K.; Patil, B.S. Variation in key flavonoid biosynthetic enzymes and phytochemicals in “Rio Red” grapefruit (Citrus paradisi Macf.) during fruit development. J. Agric. Food Chem. 2016, 64, 9022–9032. [Google Scholar] [CrossRef]
- Asikin, Y.; Taira, I.; Inafuku, S.; Sumi, H.; Sawamura, M.; Takara, K.; Wada, K. Volatile aroma components and antioxidant activities of the flavedo peel extract of unripe shiikuwasha (Citrus depressa Hayata). J. Food Sci. 2012, 77, 469–475. [Google Scholar] [CrossRef]
- Liu, C.; Yan, F.; Gao, H.; He, M.; Wang, Z.; Cheng, Y.; Deng, X.; Xu, J. Features of citrus terpenoid production as revealed by carotenoid, limonoid and aroma profiles of two pummelos (Citrus maxima) with different flesh color. J. Sci. Food Agric. 2015, 95, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Ballester, A.R.; Lafuente, M.T.; González-Candelas, L. Spatial study of antioxidant enzymes, peroxidase and phenylalanine ammonia-lyase in the citrus fruit-Penicillium digitatum interaction. Postharvest Biol. Technol. 2006, 39, 115–124. [Google Scholar] [CrossRef]
- Ben-Yehoshua, S.; Rodov, V.; Nafussi, B.; Feng, X.; Yen, J.; Koltai, T.; Nelkenbaum, U. Involvement of limonene hydroperoxides formed after oil gland injury in the induction of defense response against Penicillium digitatum in lemon fruit. J. Agric. Food Chem. 2008, 56, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.Q.; Azrina, A. Antioxidant content and activity in different parts of pomelo [Citrus grandis (L.) Osbeck] by-products. Acta Hortic. 2017, 27–34. [Google Scholar] [CrossRef]
- Cronjé, P.J.R.; Zacarías, L.; Alférez, F. Susceptibility to postharvest peel pitting in Citrus fruits as related to albedo thickness, water loss and phospholipase activity. Postharvest Biol. Technol. 2017, 123, 77–82. [Google Scholar] [CrossRef]
- Chen, J.; Li, S.; Xu, J.; Ding, F.; Wang, Z.; Cheng, Y.; Deng, X. Concentration and distribution of main bitter compounds in fruit tissues of “Oroblanco” (Citrus grandis L.× Citrus paradisi Macf.). Sci. Hortic. (Amsterdam) 2015, 193, 84–89. [Google Scholar] [CrossRef]
- Breksa, A.P.; Manners, G.D. Determination of limonin D-ring lactone hydrolase activity by solid phase extraction with indirect fluorescence detection. J. Agric. Food Chem. 2004, 52, 3772–3775. [Google Scholar] [CrossRef]
- Hamdan, D.; El-Readi, M.Z.; Tahrani, A.; Herrmann, F.; Kaufmann, D.; Farrag, N.; El-Shazly, A.; Wink, M. Chemical composition and biological activity of Citrus jambhiri Lush. Food Chem. 2011, 127, 394–403. [Google Scholar] [CrossRef]
- Pan, Z.; Li, Y.; Deng, X.; Xiao, S. Non-targeted metabolomic analysis of orange (Citrus sinensis [L.] Osbeck) wild type and bud mutant fruits by direct analysis in real-time and HPLC-electrospray mass spectrometry. Metabolomics 2014, 10, 508–523. [Google Scholar] [CrossRef]
- Zhang, A.; Zhou, X.; Zhao, H.; Guan, Y.; Zou, S.; Yan, G.; Ma, C.W.; Liu, Q.; Wang, X. Correction: Rapidly improved determination of metabolites from biological data sets using the high-efficient TransOmics tool. Mol. BioSyst. 2015, 11, 317. [Google Scholar] [CrossRef]
- De Castro, W.V.; Mertens-Talcott, S.; Rubner, A.; Butterweck, V.; Derendorf, H. Variation of flavonoids and furanocoumarins in grapefruit juices: A potential source of variability in grapefruit juice-drug interaction studies. J. Agric. Food Chem. 2006, 54, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, C.; Tautenhahn, R.; Böttcher, C.; Larson, T.R.; Neumann, S. CAMERA: An integrated strategy for compound spectra extraction and annotation of liquid chromatography/mass spectrometry data sets. Anal. Chem. 2012, 84, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.A.; Terol, J.; Ibanez, V.; López-García, A.; Pérez-Román, E.; Borredá, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Van den Berg, R.A.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Nueda, M.J.; Ferrer, A.; Talón, M. maSigPro: a method to identify significantly differential expression profiles in time-course microarray experiments. Bioinformatics 2006, 22, 1096–1102. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Class | Compound Name | Taste Trait |
|---|---|---|
| Flavanones | Naringin (O-neohesperidoside) | Bitter |
| Narirutin (O-rutinoside) | Tasteless | |
| Diosmin (O-rutinoside) | Tasteless | |
| Neodiosmin (O-neohesperidoside) | Bitter | |
| Flavones | Tangeretin | Bitter |
| Nobiletin | Bitter | |
| Sinensetin | Bitter | |
| Flavonols | Rutin | Tasteless |
| Limonoid aglycones | Limonin D-ring lactone | Tasteless |
| Nomilin | Bitter | |
| Limonin A-ring lactone | Bitter | |
| Limonoid glycosides | Limonin D-ring glycoside | Tasteless |
| Compound | Chemical Formula | Quantifier Ion ESI+ | Annotation Positive | Quantifier Ion ESI− | Annotation Negative | Retention Time (min) | |
|---|---|---|---|---|---|---|---|
| Flavonoids | |||||||
| C1 | Naringenin | C15H12O5 | 273.07 | [M+H]+ | 271.06 | [M−H]− | 9.05 |
| C2 | Hesperidin | C28H34O15 | 303.09 | [M-hesperidoside]+ | 301.07 | [M-hesperidoside]− | 6.92 |
| C3 | Neohesperidin | C28H34O15 | 303.09 | [M-neohesperidoside]+ | 301.07 | [M-hesperidoside]− | 7.7 |
| C4 | Isosinensetin | C20H20O7 | 373.13 | [M+H]+ | nd | nd | 9.75 |
| C5 | Sinensetin | C20H20O7 | 373.13 | [M+H]+ | nd | nd | 10.4 |
| C6 | Tangeretin | C20H20O7 | 373.13 | [M+H]+ | nd | nd | 11.6 |
| C7 | Eriodictyol rutinoside #1 | C27H32O15 | 597.17 | [M+H]+ | 595.17 | [M−H]− | 5.31 |
| C8 | Eriodictyol rutinoside #2 | C27H32O15 | 597.17 | [M+H]+ | 595.175 | [M−H]− | 6.14 |
| C9 | Isorhamnetin-3-O-rutinoside | C28H32O16 | 625.19 | [M+H]+ | 623.18 | [M−H]− | 6.71 |
| C10 | Isorhamnetin rutinoside hexoside | C34H42O21 | 787.22 | [M+H]+ | 785.21 | [M−H]− | 5.36 |
| C11 | Isorhamnetin rutinoside deoxyhexoside | C34H42O20 | 771.23 | [M+H]+ | 769.22 | [M−H]− | 6.07 |
| C12 | Kaempferol diDeoxyhexoside hexoside | C33H40O19 | 595.16 | [M-Hexose]+ | 739.21 | [M−H]− | 6.03 |
| C13 | Kaempferol Deoxyhexoside hexoside | C27H30O15 | 595.17 | [M+H]+ | nd | nd | 6.62 |
| C14 | Kaempferol Caffeoyl Hexoside Deoxyhexoside | C36H36O18 | 757.23 | [M+H]+ | 755.21 | [M−H]− | 5.28 |
| C15 | Kaempferol hidoxymethyl glutaryl (HMG)-glucoside tentative | C27H28O15 | 593.15 | [M+H]+ | 591.13 | [M−H]− | 7.12 |
| C16 | Quercetin hexoside rutinoside | C33H40O21 | 773.21 | [M+H]+ | 771.26 | [M−H]− | 5.01 |
| C17 | Poncirin | C28H34O14 | 595.20 | [M+H]+ | 593.19 | [M−H]− | 7.94 |
| C18 | Narirutin | C27H32O14 | 581.18 | [M+H]+ | 579.17 | [M−H]− | 5.53 |
| C19 | Naringin | C27H32O14 | 581.18 | [M+H]+ | 579.17 | [M−H]− | 6.65 |
| Limonoids | |||||||
| C20 | Deacetyl Nomilinic acid glycoside tentative | C32H46O15 | nd | nd | 669.27 | [M−H]− | 6.55 |
| C21 | Deacetyl Nomilin glycoside | C32H44O14 | nd | nd | 651.27 | [M−H]− | 6.96 |
| C22 | Nomilin | C28H34O9 | 515.23 | [M+H]+ | 513.21 | [M−H]− | 11.14 |
| C23 | Nomilinic acid glycoside | C34H48O16 | 735.29 | [M+Na]+ | 711.29 | [M−H]− | 7.37 |
| C24 | Nomilin A-ring lactone | C28H36O10 | 533.24 | [M+H]+ | 531.23 | [M−H]− | 10.53 |
| C25 | Nomilin glycoside tentative | C34H44O14 | nd | nd | 675.27 | [M−H]− | 9.51 |
| C26 | Obacunone | C26H30O7 | 455.23 | [M+H]+ | nd | nd | 11.16 |
| C27 | Obacunoic acid | C26H32O8 | 473.21 | [M+H]+ | 471.20 | [M−H]− | 10.20 |
| C28 | Obacunone glycoside | C32H42O13 | nd | nd | 633.25 | [M−H]− | 7.71 |
| C29 | Ichangin | C26H32O9 | 489.21 | [M+H]+ | 487.19 | [M−H]− | 9.63 |
| C30 | Limonoate A-ring lactone | C26H32O9 | 489.21 | [M+H]+ | 487.19 | [M−H]− | 9.81 |
| C31 | Limonin | C26H30O8 | 471.20 | [M+H]+ | 469.18 | [M−H]− | 10.57 |
| C32 | Limonin 17-β-d-glucopyranoside | C32H42O14 | 471.20 | [M-Hexose]+ | 649.25 | [M−H]− | 6.43 |
| Pulp | |
|---|---|
| Cluster # | Metabolites * |
| Cluster 1 | C1, C7, C8, C13, C17, C19 |
| Cluster 2 | C2, C3, C4, C5, C6, C9, C27, C29, C30 |
| Cluster 3 | C14, C20, C23, C28, C32 |
| Cluster 4 | C15, C21, C22, C24, C25, C26, C31 |
| Albedo | |
| Cluster # | Metabolites |
| Cluster 1 | C1, C15, C17, C19 |
| Cluster 2 | C2, C3, C6, C7, C8, C12, C30 |
| Cluster 3 | C14, C18, C23, C25, C28, C32 |
| Cluster 4 | C21, C22, C26, C31 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadi, R.; Golein, B.; Gómez-Cadenas, A.; Arbona, V. Developmental Stage- and Genotype-Dependent Regulation of Specialized Metabolite Accumulation in Fruit Tissues of Different Citrus Varieties. Int. J. Mol. Sci. 2019, 20, 1245. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051245
Nadi R, Golein B, Gómez-Cadenas A, Arbona V. Developmental Stage- and Genotype-Dependent Regulation of Specialized Metabolite Accumulation in Fruit Tissues of Different Citrus Varieties. International Journal of Molecular Sciences. 2019; 20(5):1245. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051245
Chicago/Turabian StyleNadi, Roya, Behrouz Golein, Aurelio Gómez-Cadenas, and Vicent Arbona. 2019. "Developmental Stage- and Genotype-Dependent Regulation of Specialized Metabolite Accumulation in Fruit Tissues of Different Citrus Varieties" International Journal of Molecular Sciences 20, no. 5: 1245. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051245