Transcriptome Analysis Reveals Cotton (Gossypium hirsutum) Genes That Are Differentially Expressed in Cadmium Stress Tolerance


Abstract
:1. Introduction
2. Results
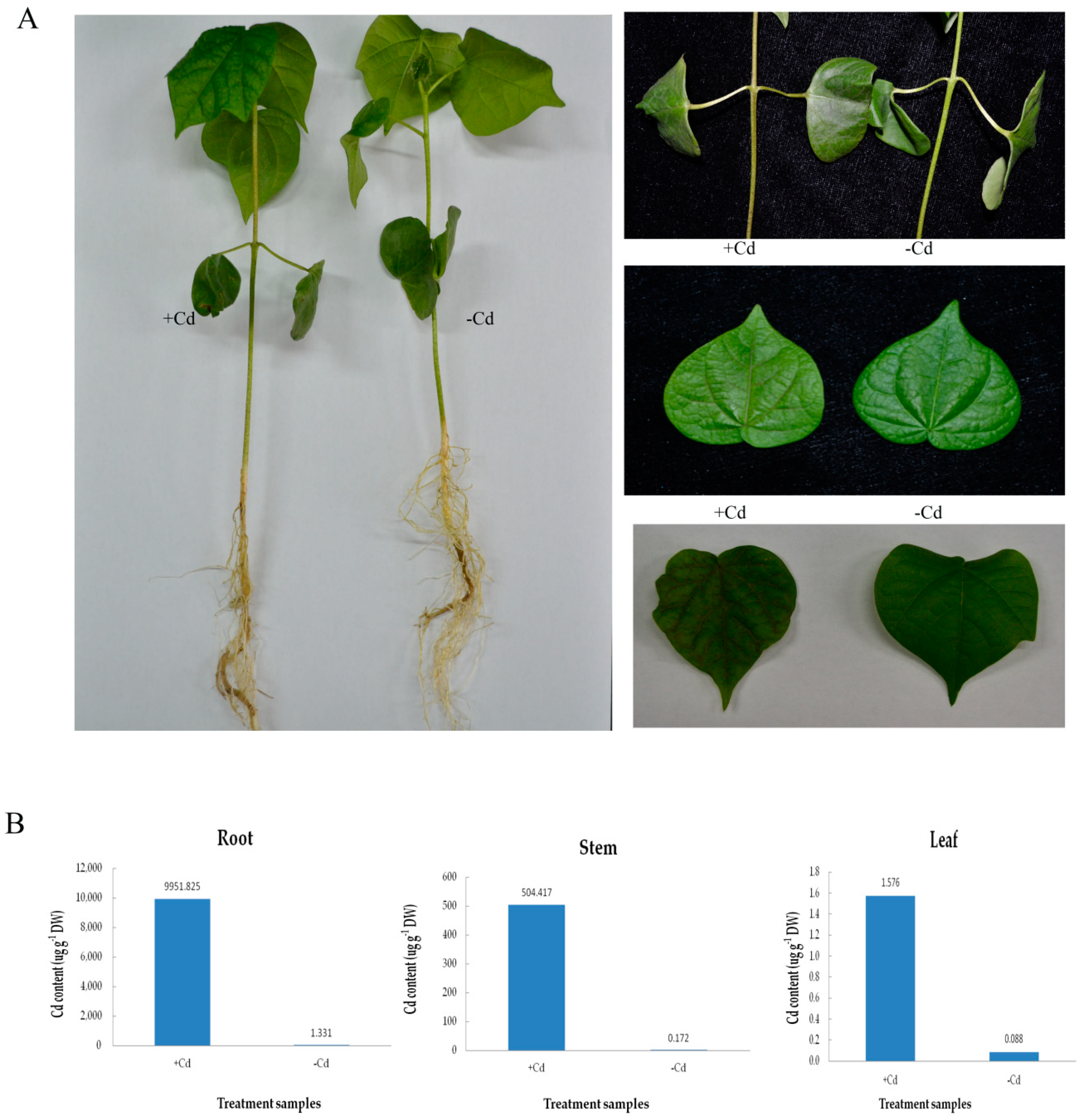
2.1. Phenotypic Analysis of Cotton under Cd Stress
2.2. Quality Analysis of the Transcriptome Sequence
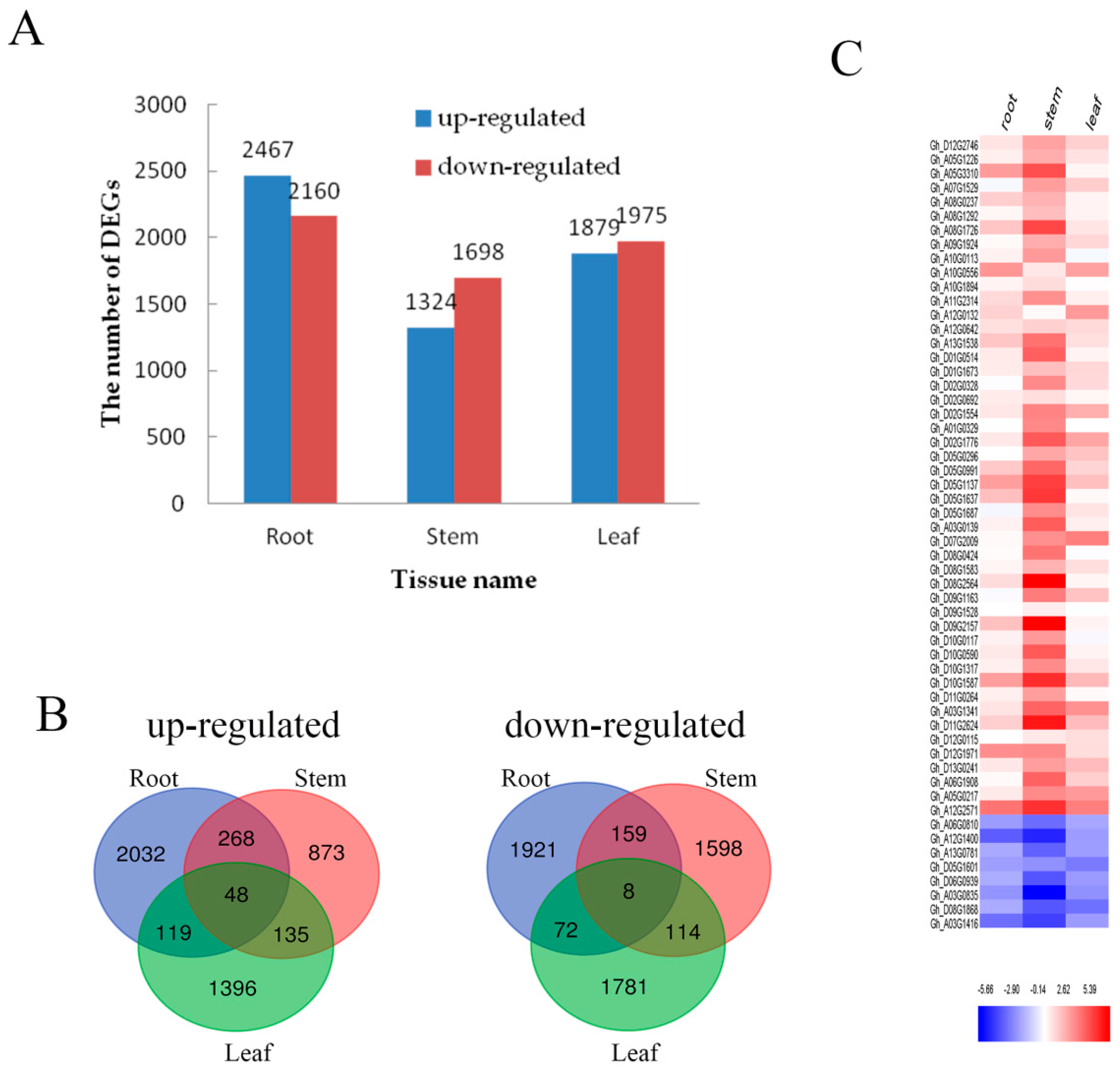
2.3. Analysis of Differential Expression Genes under Cd Stress in Cotton
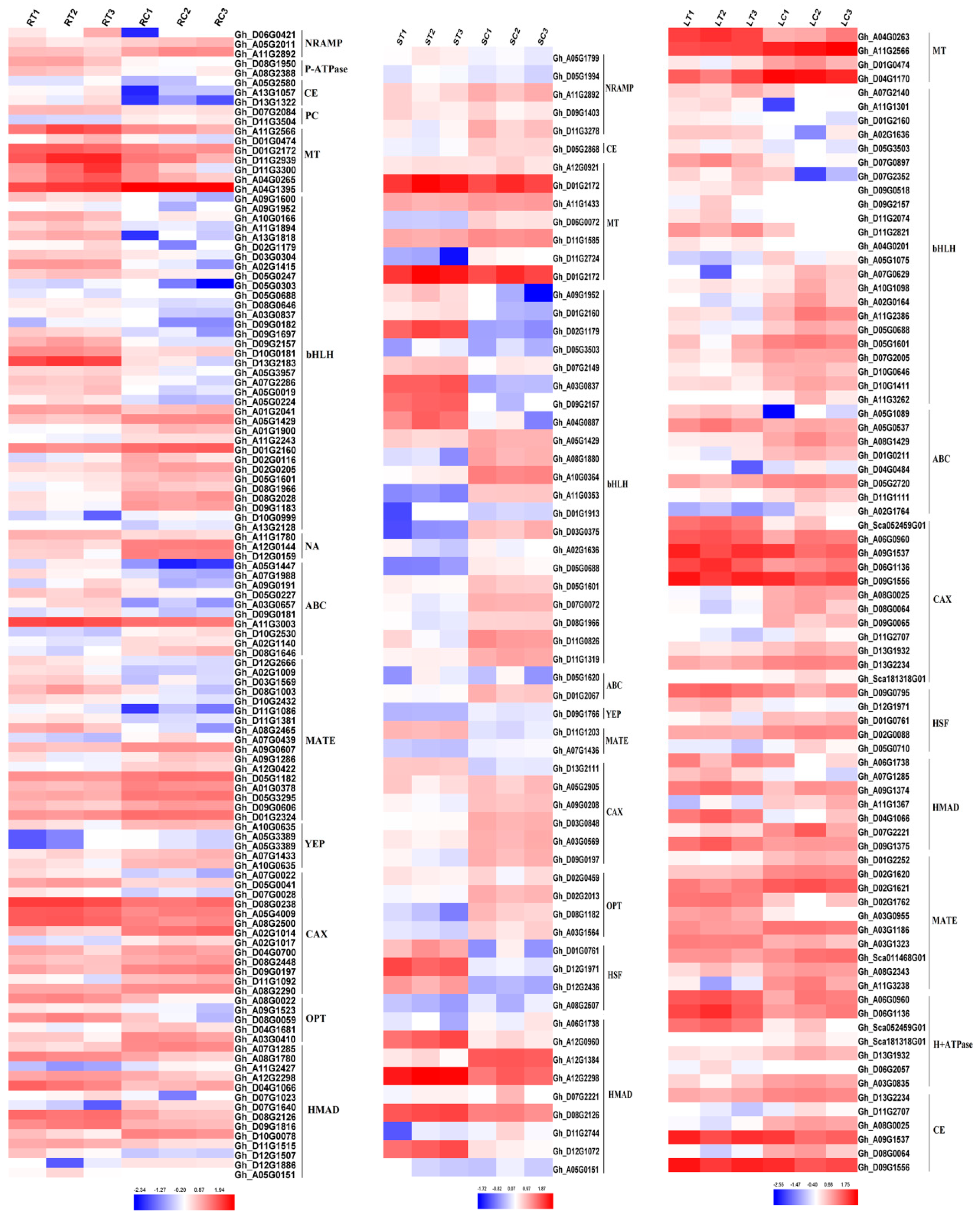
2.4. Expression of Heavy Metal Transport Proteins in Cotton under Cd Stress
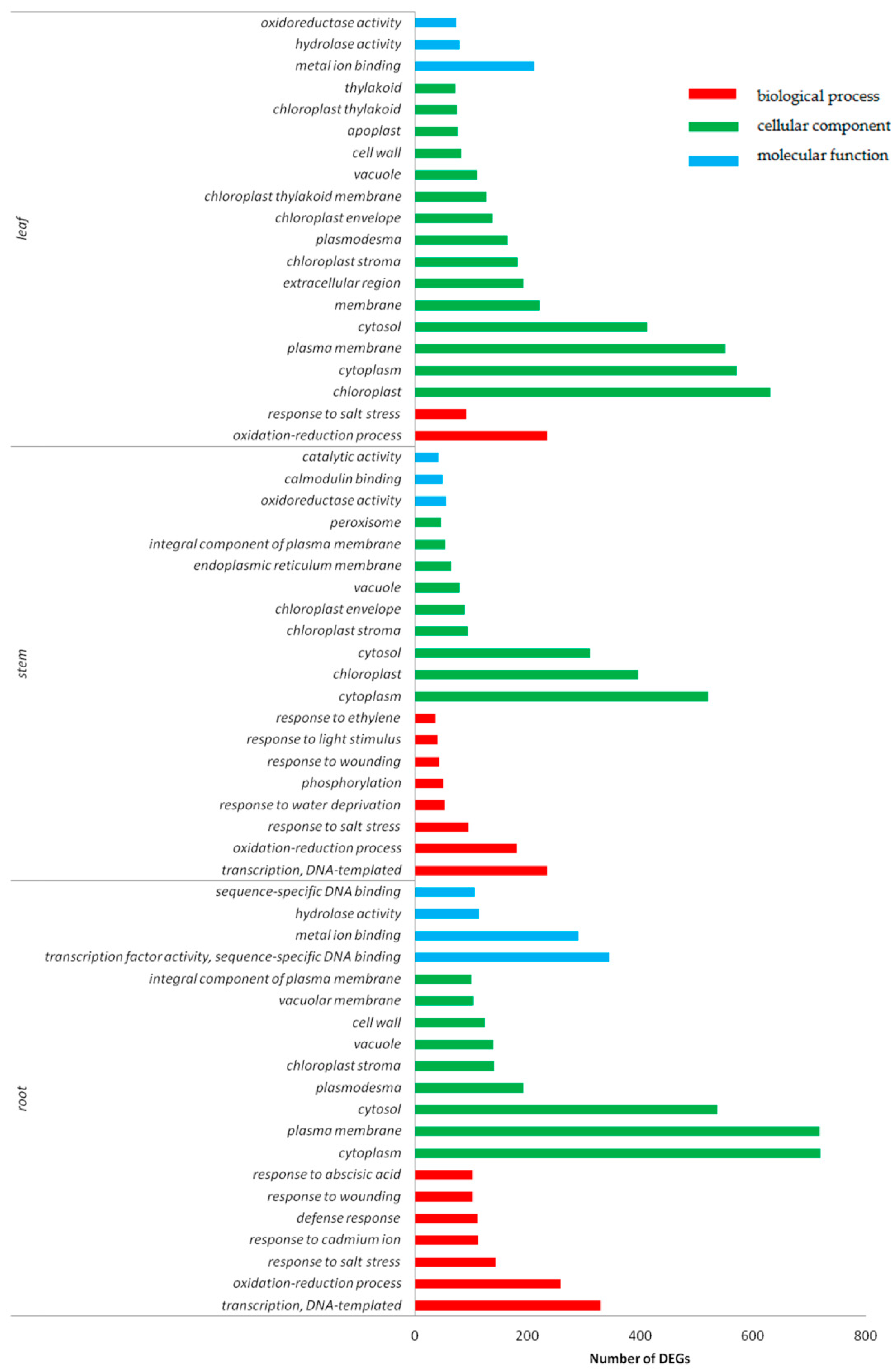
2.5. Functional Classification of DEGs
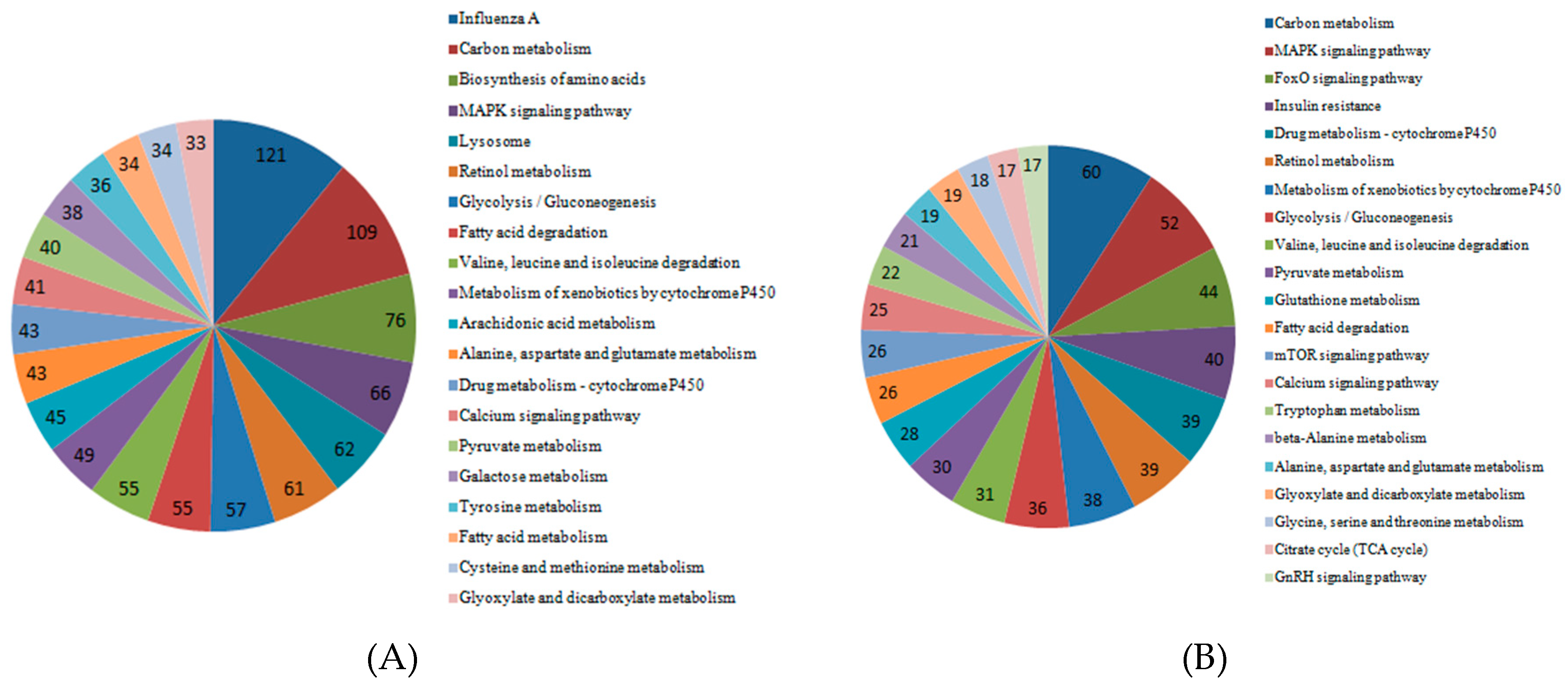
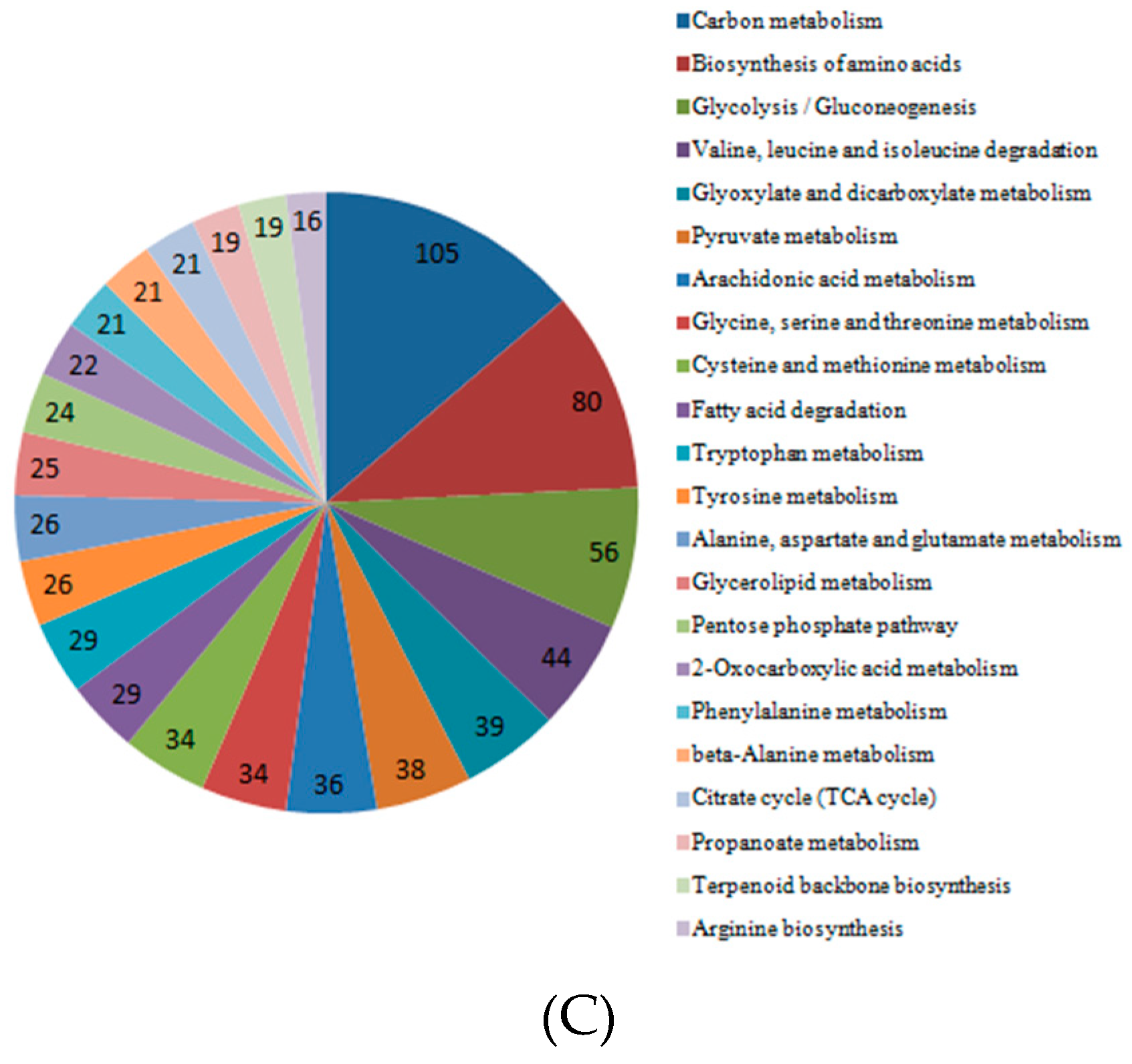
2.6. KEGG Analysis of DEGs
2.7. Expression of Transcription Factors under Cd Stress of Cotton
2.8. Verification of Sequence Data
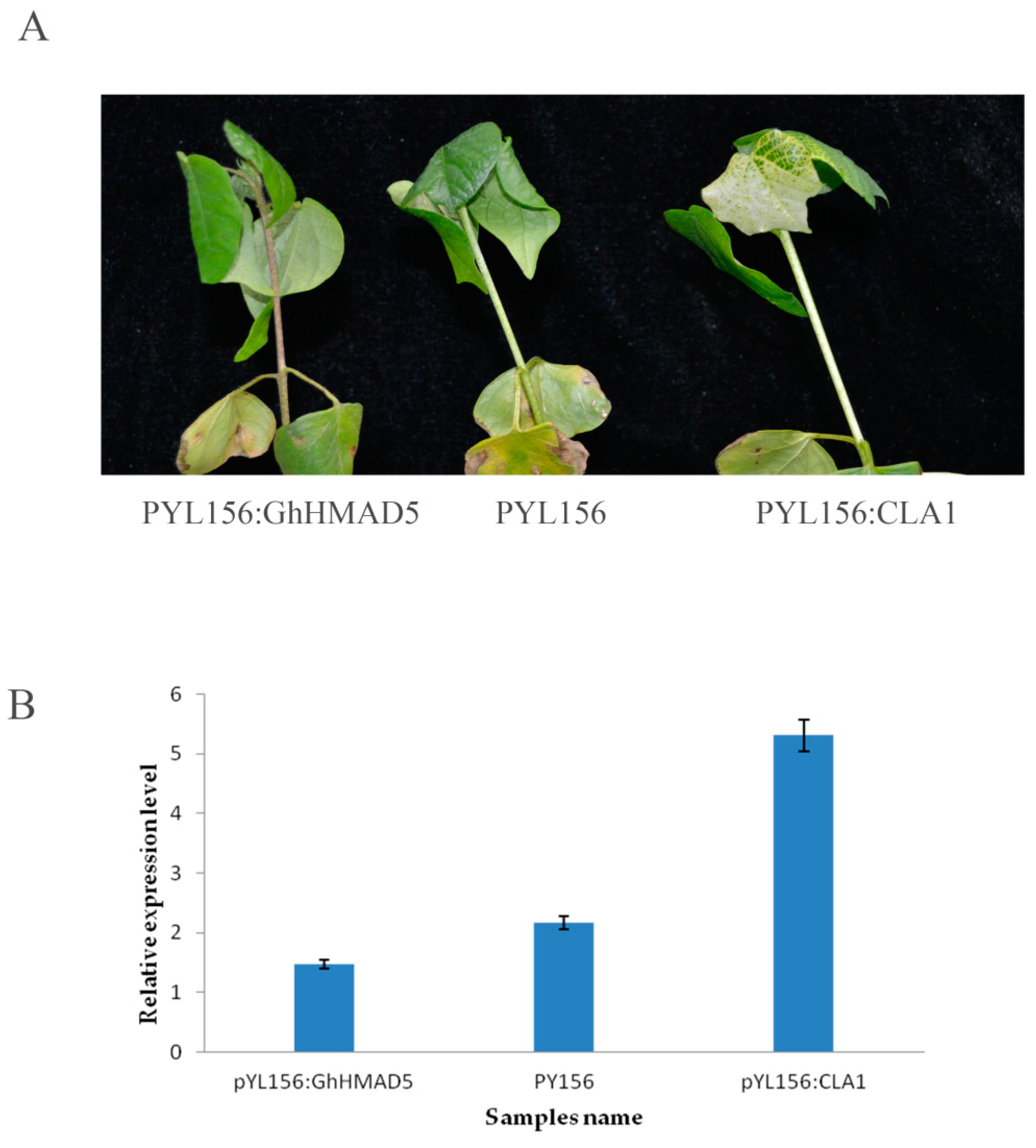
2.9. Silencing of GhHMAD5: Phenotype and Expression
2.10. Overexpression of GhHMAD5 Could Enhance the Cd Tolerance of Arabidopsis Thaliana
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Cd Stress Treatment
4.2. Measurement of Relative Cd Content
4.3. RNA Extraction, cDNA Library Construction, and RNA-Seq
4.4. Quality Control, and DEG Analysis
4.5. Gene Ontology and Gene Pathway Enrichment Analysis
4.6. Quantitative Real-Time PCR Analysis
4.7. Overexpression and Virus Induced Gene Silence (VIGS) Construction of GhHMAD5
4.8. Analysis of Cd Tolerance in Transgenic Arabidopsis Thaliana
4.9. The VIGS Analysis of GhHMAD5
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kaplan, O.; Ince, M.; Yaman, M. Sequential extraction of cadmium in different soil phases and plant parts from a former industrialized area. Environ. Chem. Lett. 2011, 9, 397–404. [Google Scholar] [CrossRef]
- Groppa, M.D.; Ianuzzo, M.P.; Rosales, E.P.; Vázquez, S.C.; Benavides, M.P. Cadmium modulates NADPH oxidase activity and expression in sunflower leaves. Biol. Plant. 2012, 56, 167–171. [Google Scholar] [CrossRef]
- Satarug, S.; Baker, J.R.; Urbenjapol, S.; Haswell-Elkins, M.; Reilly, P.E.B.; Williams, D.J.; Moore, M.R. A global perspective on cadmium pollution and toxicity in non-occupationally exposed population. Toxicol. Lett. 2003, 137, 65–83. [Google Scholar] [CrossRef]
- Toppi, L.S.D.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Brooks, R.R.; Lee, J.; Reeves, R.D.; Jaffre, T. Detection of nickeliferous rocks by analysis of herbarium specimens of indicator plants. J. Geochem. Explor 1977, 7, 49–57. [Google Scholar] [CrossRef]
- Zhao, Y.; Hu, H.X.; Chen, Z.F.; Hu, L.J.; Xu, D.Y. The potential for phytoextraction of hexachloro-cyclohexanes contaminated calcareous soils in eastern Suburb of Beijing by cotton. Fresenius Environ. Bull. 2012, 21, 1948–1955. [Google Scholar]
- Ling, L.I.; Chen, J.H.; Qiu-Ling, H.E.; Zhu, S.J. Accumulation, transportation, and bioconcentration of cadmium in three upland cotton plants under cadmium stress. Cotton Sci. 2012, 24, 535–540. [Google Scholar]
- Kolevavalkova, L.; Vasilev, A. Physiological parameters of young cotton plants, grown on heavy metal contaminated soils. Agrarni Nauki 2015, VII, 61–66. [Google Scholar]
- Daud, M.K.; Sun, Y.; Dawood, M.; Hayat, Y.; Variath, M.T.; Wu, Y.X.; Raziuddin; Mishkat, U.; Salahuddin; Najeeb, U. Cadmium-induced functional and ultrastructural alterations in roots of two transgenic cotton cultivars. J. Hazard. Mater. 2008, 161, 463–473. [Google Scholar] [CrossRef]
- Liu, L.T.; Sun, H.C.; Chen, J.; Zhang, Y.J.; Wang, X.D.; Li, D.X.; Li, C.D. Cotton seedling plants adapted to cadmium stress by enhanced activities of protective enzymes. Plant Soil Environ. 2016, 62, 80–85. [Google Scholar] [Green Version]
- Ibrahim, W.; Ahmed, I.M.; Chen, X.; Cao, F.; Zhu, S.; Wu, F. Genotypic differences in photosynthetic performance, antioxidant capacity, ultrastructure and nutrients in response to combined stress of salinity and Cd in cotton. Biometals 2015, 28, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Angelova, V.; Ivanova, R.; Delibaltova, V.; Ivanov, K. Bio-accumulation and distribution of heavy metals in fibre crops (flax, cotton and hemp). Ind. Crops Prod. 2004, 19, 197–205. [Google Scholar] [CrossRef]
- Wu, F.; Wu, H.; Zhang, G.; Dml, B. Differences in growth and yield in response to cadmium toxicity in cotton genotypes. J. Plant Nutr. Soil Sci. 2004, 167, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Hartwell, L.H.; Culotti, J.; Reid, B. Genome-wide expression profiling of maize in response to individual and combined water and nitrogen stresses. BMC Genomics. 2013, 14, 3. [Google Scholar] [CrossRef]
- Yu, S.; Zhang, F.; Yu, Y.; Zhang, D.; Zhao, X.; Wang, W. Transcriptome profiling of dehydration stress in the Chinese Cabbage (Brassica rapa L. ssp. pekinensis) by tag sequencing. Plant Mol. Biol. Rep. 2012, 30, 17–28. [Google Scholar] [CrossRef]
- Liu, T.; Zhu, S.; Tang, Q.; Tang, S. Genome-wide transcriptomic profiling of ramie (Boehmeria nivea L. Gaud) in response to cadmium stress. Gene 2015, 558, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Yue, R.; Lu, C.; Qi, J.; Han, X.; Yan, S.; Guo, S.; Liu, L.; Fu, X.; Chen, N.; Yin, H. Transcriptome analysis of cadmium-treated roots in maize (Zea mays L.). Front. Plant Sci. 2016, 7, 1298. [Google Scholar] [CrossRef] [PubMed]
- Oono, Y.; Yazawa, T.; Kanamori, H.; Sasaki, H.; Mori, S.; Handa, H.; Matsumoto, T. Genome-wide transcriptome analysis of cadmium stress in rice. BioMed Res. Int. 2016, 2016, 1–9. [Google Scholar] [CrossRef]
- Liu, Y.J.; Yu, X.F.; Feng, Y.M.; Zhang, C.; Wang, C.; Zeng, J.; Huang, Z.; Kang, H.Y.; Fan, X.; Sha, L.N.; et al. Physiological and transcriptome response to cadmium in cosmos (Cosmos bipinnatus Cav.) seedlings. Sci. Rep. 2017, 7, 14691. [Google Scholar] [CrossRef] [Green Version]
- Aprile, A.; Sabella, E.; Vergine, M.; Genga, A.; Siciliano, M.; Nutricati, E.; Rampino, P.; De Pascali, M.; Luvisi, A.; Miceli, A.; et al. Activation of a gene network in durum wheat roots exposed to cadmium. BMC Plant Biol. 2018, 18, 238. [Google Scholar] [CrossRef] [PubMed]
- Shukla, D.; Huda, K.M.; Banu, M.S.; Gill, S.S.; Tuteja, R.; Tuteja, N. OsACA6, a P-type 2B Ca 2+ ATPase functions in cadmium stress tolerance in tobacco by reducing the oxidative stress load. Planta 2014, 240, 809–824. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Saier, M.H.A., Jr. Novel family of ubiquitous heavy metal ion transport proteins. J. Membr. Biol. 1997, 156, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Rivetta, A.; Negrini, N.; Cocucci, M. Involvement of Ca2+-calmodulin in Cd2+ toxicity during the early phases of radish (Raphanus sativus L.) seed germination. Plant Cell Environ. 1997, 20, 600–608. [Google Scholar] [CrossRef]
- Hegedüs, A.; Erdei, S.; Horváth, G. Comparative studies of H(2)O(2) detoxifying enzymes in green and greening barley seedlings under cadmium stress. Plant Sci. 2001, 160, 1085–1093. [Google Scholar] [CrossRef]
- Chen, B.C.; Wang, P.J.; Ho, P.C.; Juang, K.W. Nonlinear biotic ligand model for assessing alleviation effects of Ca, Mg, and K on Cd toxicity to soybean roots. Ecotoxicology 2017, 26, 1–14. [Google Scholar] [CrossRef]
- Gong, B.; Nie, W.; Yan, Y.; Gao, Z.; Shi, Q. Unravelling cadmium toxicity and nitric oxide induced tolerance in Cucumis sativus: Insight into regulatory mechanisms using proteomics. J. Hazard. Mater. 2017, 336, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, H.; Wu, L.; Liu, A.; Zhao, F.J.; Xu, W. Heavy metal ATPase 3 (HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytol. 2017, 215, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; An, T.; Zhu, X.; Ji, J.; Wang, G.; Guan, C.; Jin, C.; Yi, L. GR1-like gene expression in Lycium chinense was regulated by cadmium-induced endogenous jasmonic acids accumulation. Plant Cell Rep. 2017, 36, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Shan, C.; Zhao, H.; Jia, G.; Chen, D. Lanthanum improves the cadmium tolerance of Zea mays seedlings by the regulation of ascorbate and glutathione metabolism. Biol. Plant. 2016, 61, 1–6. [Google Scholar] [CrossRef]
- Huilan, W.; Chunlin, C.; Juan, D.; Hongfei, L.; Yan, C.; Yue, Z.; Yujing, H.; Yiqing, W.; Chengcai, C.; Zongyun, F. Co-overexpression FIT with AtbHLH38 or AtbHLH39 in Arabidopsis-enhanced cadmium tolerance via increased cadmium sequestration in roots and improved iron homeostasis of shoots. Plant Physiol. 2012, 158, 790–800. [Google Scholar]
- Tiwari, M.; Sharma, D.; Dwivedi, S.; Singh, M.; Tripathi, R.D.; Trivedi, P.K. Expression in Arabidopsis and cellular localization reveal involvement of rice NRAMP, OsNRAMP1, in arsenic transport and tolerance. Plant Cell Environ. 2014, 37, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Sekhar, K.; Priyanka, B.; Reddy, V.D.; Rao, K.V. Metallothionein 1 (CcMT1) of pigeonpea (Cajanus cajan, L.) confers enhanced tolerance to copper and cadmium in Escherichia coli and Arabidopsis thaliana. Environ. Exp. Bot. 2011, 72, 131–139. [Google Scholar] [CrossRef]
- Shukla, D.; Kesari, R.; Tiwari, M.; Dwivedi, S.; Tripathi, R.D.; Nath, P.; Trivedi, P.K. Expression of Ceratophyllum demersum phytochelatin synthase, CdPCS1, in Escherichia coli and Arabidopsis enhances heavy metal(loid)s accumulation. Protoplasma 2013, 250, 1263–1272. [Google Scholar] [CrossRef]
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchishinozaki, K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, C.; Liang, Y.; Wang, C.; Yang, C.; Liu, G. A novel bZIP gene from Tamarix hispida mediates physiological responses to salt stress in tobacco plants. J. Plant Physiol. 2010, 167, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Chmielowska-Bąk, J.; Lefèvre, I.; Lutts, S.; Deckert, J. Short term signaling responses in roots of young soybean seedlings exposed to cadmium stress. J. Plant Physiol. 2013, 170, 1585–1594. [Google Scholar] [CrossRef]
- Wei, W.; Zhang, Y.; Han, L.; Guan, Z.; Chai, T. A novel WRKY transcriptional factor from Thlaspi caerulescens negatively regulates the osmotic stress tolerance of transgenic tobacco. Plant Cell Rep. 2008, 27, 795–803. [Google Scholar] [CrossRef]
- Mao, J.; Li, W.; Mi, B.; Dawuda, M.M.; Calderón-Urrea, A.; Ma, Z.; Zhang, Y.; Chen, B. Different exogenous sugars affect the hormone signal pathway and sugar metabolism in “Red Globe” (Vitis vinifera L.) plantlets grown in vitro as shown by transcriptomic analysis. Planta 2017, 246, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gupta, O.P.; Sharma, P.; Gupta, R.K.; Sharma, I. MicroRNA mediated regulation of metal toxicity in plants: Present status and future perspectives. Plant Mol. Biol. 2014, 84, 1–18. [Google Scholar] [CrossRef]
- Sghayar, S.; Ferri, A.; Lancilli, C.; Lucchini, G.; Abruzzese, A.; Porrini, M.; Ghnaya, T.; Nocito, F.F.; Abdelly, C.; Sacchi, G.A. Analysis of cadmium translocation, partitioning and tolerance in six barley (Hordeum vulgare L.) cultivars as a function of thiol metabolism. Biol. Fertil. Soils 2015, 51, 311–320. [Google Scholar] [CrossRef]
- Salim, R.; Al-Subu, M.M.; Douleh, A.; Khalaf, S. Effects on growth and uptake of broad beans (Vicia fabae L.) by root and foliar treatments of plant with lead and cadmium. Environ. Lett. 2012, 27, 1619–1642. [Google Scholar] [CrossRef]
- Moussa, H.R. Effect of cadmium on growth and oxidative metabolism of faba bean plants. Acta Agron. Hungarica 2004, 52, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Bull, P.C.; Cox, D.W. Wilson disease and Menkes disease: New handles on heavy-metal transport. Trends Genet. 1994, 10, 246–252. [Google Scholar] [CrossRef]
- Gitschier, J.; Moffat, B.; Reilly, D.; Wood, W.I.; Fairbrother, W.J. Solution structure of the fourth metal-binding domain from the Menkes copper-transporting ATPase. Nat. Struct. Biol. 1998, 5, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Lebherz, H.G.; Leadbetter, M.M.; Bradshaw, R.A. Isolation and characterization of the cytosolic and chloroplast forms of spinach leaf fructose diphosphate aldolase. J. Biol. Chem. 1984, 259, 1011–1017. [Google Scholar] [PubMed]
- Yamada, S.; Komori, T.; Hashimoto, A.; Kuwata, S.; Imaseki, H.; Kubo, T. Differential expression of plastidic aldolase genes in Nicotiana plants under salt stress. Plant Sci. 2000, 154, 61–69. [Google Scholar] [CrossRef]
- Chen, H.; Li, Y.; Ma, X.; Guo, L.; He, Y.; Ren, Z.; Kuang, Z.; Zhang, X.; Zhang, Z. Analysis of potential strategies for cadmium stress tolerance revealed by transcriptome analysis of upland cotton. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Lin, Z.; Cui, H.M.; Wang, J.J.; Hou, X.L.; Ying, L.I. Effects of Cadmium stress on genes expression in L-Galactose pathway involved in VC biosynthesis and antioxidant system of Brassica campestris ssp. chinensis. Plant Physiol. J. 2015, 51, 1099–1108. [Google Scholar]
- Shao, X.J.; Yang, H.Q.; Ran, K.; Jiang, Q.Q.; Sun, X.L. Effects of Salicylic acid on plasma membrane ATPase and free radical of grape root under cadmium stress. Sci. Agric. Sin. 2010, 43, 1441–1447. [Google Scholar]
- Schellingen, K.; Straeten, D.V.D.; Remans, T.; Vangronsveld, J.; Keunen, E.; Cuypers, A. Ethylene signalling is mediating the early cadmium-induced oxidative challenge in Arabidopsis thaliana. Plant Sci. 2015, 239, 137–146. [Google Scholar] [CrossRef]
- Tougane, K.; Komatsu, K.; Bhyan, S.B.; Sakata, Y.; Ishizaki, K.; Yamato, K.T.; Kohchi, T.; Takezawa, D. Evolutionarily Conserved regulatory mechanisms of abscisic acid signaling in land plants: characterization of -like type 2C protein phosphatase in the liverwort. Plant Physiol. 2010, 152, 1529–1543. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.J.; Liu, X.S.; Tao, H.; Tan, S.K.; Chu, S.S.; Oono, Y.; Zhang, X.D.; Chen, J.; Yang, Z.M. Variation of DNA methylation patterns associated with gene expression in rice (Oryza sativa) exposed to cadmium. Plant Cell Environ. 2016, 39, 2629–2649. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Cheadle, C.; Vawter, M.P.; Freed, W.J.; Becker, K.G. Analysis of microarray data using Z score transformation. J. Mol. Diagn. JMD 2003, 5, 73–81. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Bi, R.; Liu, P. Sample size calculation while controlling false discovery rate for differential expression analysis with RNA-sequencing experiments. BMC Bioinform. 2016, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucl. Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucl. Acids Res. 2011, 39, 316–322. [Google Scholar] [CrossRef]
- Qian, W.; Tao, T.; Zhang, Y.; Wu, W.; Li, D.; Yu, J.; Han, C. Rice black-streaked dwarf virus P6 self-interacts to form punctate, viroplasm-like structures in the cytoplasm and recruits viroplasm-associated protein P9-1. Virol. J. 2011, 8, 1–15. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Number of Raw Reads | Number of Valid Reads | Q20 Percentage (%) | Q30 Percentage (%) | GC Content (%) |
|---|---|---|---|---|---|
| R_T1 | 62,264,174 | 61,471,556 | 98.04 | 91.00 | 45 |
| R_T2 | 55,526,318 | 54,811,464 | 97.93 | 91.32 | 45 |
| R_T3 | 57,133,152 | 56,462,772 | 98.27 | 92.14 | 45 |
| R_C1 | 55,124,734 | 54,473,502 | 97.93 | 90.83 | 45 |
| R_C2 | 65,102,420 | 64,390,908 | 98.45 | 92.15 | 44 |
| R_C3 | 52,774,982 | 52,144,004 | 97.78 | 90.97 | 45 |
| S_T1 | 54,098,272 | 53,474,704 | 99.35 | 95.05 | 44 |
| S_T2 | 43,663,718 | 43,152,570 | 99.19 | 95.13 | 45 |
| S_T3 | 43,207,572 | 42,631,528 | 99.43 | 96.20 | 44 |
| S_C1 | 57,188,916 | 56,544,630 | 98.99 | 94.40 | 45 |
| S_C2 | 42,417,384 | 41,057,536 | 98.01 | 89.91 | 44 |
| S_C3 | 50,332,770 | 48,940,728 | 98.28 | 90.06 | 45 |
| L_T1 | 42,697,218 | 41,184,508 | 98.22 | 89.03 | 44 |
| L_T2 | 45,204,982 | 44,593,254 | 98.25 | 89.42 | 45 |
| L_T3 | 48,167,202 | 47,153,586 | 98.30 | 90.14 | 44 |
| L_C1 | 63,540,928 | 62,794,552 | 99.67 | 96.60 | 44 |
| L_C2 | 39,136,086 | 38,791,254 | 99.66 | 95.22 | 44 |
| L_C3 | 46,343,822 | 45,925,388 | 99.74 | 95.38 | 44 |
| Gene ID | RNA-Seq (Log2FC) | qRT-PCR (FC) | Description |
|---|---|---|---|
| Gh_A01G1234 | 3.24 | 2.89 | PLANT CADMIUM RESISTANCE 2 |
| Gh_D11G2939 | 3.12 | 2.42 | metallothionein 3 |
| Gh_D06G0421 | 2.97 | 5.86 | NRAMP metal ion transporter 6 |
| Gh_D04G1180 | 2.68 | 2.24 | Fes1A |
| Gh_A11G2566 | 2.37 | 3.90 | metallothionein 3 |
| Gh_A04G0265 | 2.33 | 5.37 | metallothionein 3 |
| Gh_D08G1950 | 1.25 | 2.37 | heavy metal atpase 5 |
| Gh_A04G0713 | 2.12 | 4.38 | Fes1A |
| Gh_A08G2485 | 5.03 | 5.67 | AUX/IAA transcriptional regulator family protein |
| Gh_D07G2124 | 2.28 | 7.62 | AUX/IAA transcriptional regulator family protein |
| Gh_D08G2126 | 2.45 | 7.96 | Heavy metal transport/detoxification superfamily protein |
| Gh_D04G0262 | 3.29 | 3.84 | Auxin-responsive GH3 family protein |
| Gh_D04G0260 | 3.29 | 7.16 | Auxin-responsive GH3 family protein |
| Gh_A01G2049 | 3.13 | 3.80 | Arabidopsis thaliana gibberellin 2-oxidase 1 |
| Gh_A07G1285 | −2.49 | 0.13 | Heavy metal transport/detoxification superfamily protein |
| Gh_D07G1640 | 2.34 | 1.80 | Heavy metal transport/detoxification superfamily protein |
| Gh_D10G0078 | −2.61 | 0.49 | Heavy metal transport/detoxification superfamily protein |
| Gh_D13G1609 | 2.74 | 3.26 | Arabidopsis thaliana gibberellin 2-oxidase 1 |
| Gh_D09G1816 | 2.01 | 1.08 | Heavy metal transport/detoxification superfamily protein |
| Gh_A05G0151 | 2.08 | 2.43 | Heavy metal transport/detoxification superfamily protein |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, M.; Lu, X.; Yu, J.; Chen, X.; Wang, X.; Malik, W.A.; Wang, J.; Wang, D.; Wang, S.; Guo, L.; et al. Transcriptome Analysis Reveals Cotton (Gossypium hirsutum) Genes That Are Differentially Expressed in Cadmium Stress Tolerance. Int. J. Mol. Sci. 2019, 20, 1479. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061479
Han M, Lu X, Yu J, Chen X, Wang X, Malik WA, Wang J, Wang D, Wang S, Guo L, et al. Transcriptome Analysis Reveals Cotton (Gossypium hirsutum) Genes That Are Differentially Expressed in Cadmium Stress Tolerance. International Journal of Molecular Sciences. 2019; 20(6):1479. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061479
Chicago/Turabian StyleHan, Mingge, Xuke Lu, John Yu, Xiugui Chen, Xiaoge Wang, Waqar Afzal Malik, Junjuan Wang, Delong Wang, Shuai Wang, Lixue Guo, and et al. 2019. "Transcriptome Analysis Reveals Cotton (Gossypium hirsutum) Genes That Are Differentially Expressed in Cadmium Stress Tolerance" International Journal of Molecular Sciences 20, no. 6: 1479. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061479