Cryopreservation Differentially Alters the Proteome of Epididymal and Ejaculated Pig Spermatozoa

 ,
, 
 ,
,  and
and
Abstract
:
1. Introduction
2. Results
2.1. Fresh and Post-Thaw Sperm Parameters
2.2. Protein Repertory
2.3. Differentially Abundant Proteins
2.4. Bioinformatics
2.4.1. Gene Ontology Enrichment Analysis
2.4.2. Protein–Protein Interaction Networks
3. Discussion
3.1. Post-Thaw Sperm Quality and Functionality
3.2. Quantitative Changes in Sperm Proteins
4. Materials and Methods
4.1. Boars, Epididymis, and Ejaculates
4.2. Experimental Workflow
4.3. Sperm Cryopreservation
4.4. Quality and Sperm Functionality Assessments
4.5. Sperm Proteomics
4.5.1. Protein Extraction
4.5.2. SDS-PAGE and In-Gel Digestion
4.5.3. Sequential Window Acquisition of All Theoretical Spectra (SWATH) Analysis
4.5.4. Protein Quantification
4.5.5. Gene Ontology and Bioinformatics Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abram McBride, J.; Lipshultz, L.I. Male Fertility Preservation. Curr. Urol. Rep. 2018, 19, 49. [Google Scholar] [CrossRef]
- Kawai, K.; Nishiyama, H. Preservation of fertility of adult male cancer patients treated with chemotherapy. Int. J. Clin. Oncol. 2018, in press. [Google Scholar] [CrossRef]
- Morrell, J.M.; Mayer, I. Reproduction biotechnologies in germplasm banking of livestock species: A review. Zygote 2017, 25, 545–557. [Google Scholar] [CrossRef]
- Comizzoli, P. Biobanking efforts and new advances in male fertility preservation for rare and endangered species. Asian J. Androl. 2015, 17, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, M.A.; Papa, F.O.; Ramires Neto, C. Advances in Stallion Semen Cryopreservation. Vet. Clin. N. Am. Equine Pract. 2016, 32, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M.; Rodríguez-Gil, J.E.; Bonet, S. Artificial insemination with frozen-thawed boar sperm. Mol. Reprod. Dev. 2017, 84, 802–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozati, H.; Handley, T.; Jayasena, C.N. Process and Pitfalls of Sperm Cryopreservation. J. Clin. Med. 2017, 6, 89. [Google Scholar] [CrossRef]
- Rosenbusch, B. To What Extent Are Cryopreserved Sperm and Testicular Biopsy Samples Used in Assisted Reproduction? J. Reprod. Infertil. 2018, 19, 115–118. [Google Scholar]
- Sharma, R.; Kattoor, A.J.; Ghulmiyyah, J.; Agarwal, A. Effect of sperm storage and selection techniques on sperm parameters. Syst. Biol. Reprod. Med. 2015, 61, 1–12. [Google Scholar] [CrossRef]
- Knox, R.V. The Fertility of Frozen Boar Sperm When used for Artificial Insemination. Reprod. Domest. Anim. 2015, 50, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Pini, T.; Leahy, T.; de Graaf, S.P. Sublethal sperm freezing damage: Manifestations and solutions. Theriogenology 2018, 118, 172–181. [Google Scholar] [CrossRef]
- Morris, G.J.; Acton, E.; Murray, B.J.; Fonseca, F. Freezing injury: The special case of the sperm cell. Cryobiology 2012, 64, 71–80. [Google Scholar] [CrossRef]
- Li, C.J.; Wang, D.; Zhou, X. Sperm proteome and reproductive technologies in mammals. Anim. Reprod. Sci. 2016, 173, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hezavehei, M.; Sharafi, M.; Kouchesfahani, H.M.; Henkel, R.; Agarwal, A.; Esmaeili, V.; Shahverdi, A. Sperm cryopreservation: A review on current molecular cryobiology and advanced approaches. Reprod. Biomed. Online 2018, 37, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.Y.; Kuo, Y.H.; Lee, W.C.; Tsou, H.L.; Lee, Y.P.; Chang, H.L.; Wu, J.J.; Yang, P.C. Substantial decrease of heat-shock protein 90 precedes the decline of sperm motility during cooling of boar spermatozoa. Theriogenology 1999, 51, 1007–1016. [Google Scholar] [CrossRef]
- Lessard, C.; Parent, S.; Leclerc, P.; Bailey, J.L.; Sullivan, R. Cryopreservation alters the levels of the bull sperm surface protein P25b. J. Androl. 2000, 21, 700–707. [Google Scholar] [PubMed]
- Chatterjee, S.; de Lamirande, E.; Gagnon, C. Cryopreservation alters membrane sulfhydryl status of bull spermatozoa: Protection by oxidized glutathione. Mol. Reprod. Dev. 2001, 60, 498–506. [Google Scholar] [CrossRef]
- Casas, I.; Sancho, S.; Ballester, J.; Briz, M.; Pinart, E.; Bussalleu, E.; Yeste, M.; Fàbrega, A.; Rodríguez-Gil, J.E.; Bonet, S. The HSP90AA1 sperm content and the prediction of the boar ejaculate freezability. Theriogenology 2010, 74, 940–950. [Google Scholar] [CrossRef]
- Flores, E.; Fernández-Novell, J.M.; Peña, A.; Rigau, T.; Rodríguez-Gil, J.E. Cryopreservation-induced alterations in boar spermatozoa mitochondrial function are related to changes in the expression and location of midpiece mitofusin-2 and actin network. Theriogenology 2010, 74, 354–363. [Google Scholar] [CrossRef]
- Wang, S.; Wang, W.; Xu, Y.; Tang, M.; Fang, J.; Sun, H.; Sun, Y.; Gu, M.; Liu, Z.; Zhang, Z.; et al. Proteomic characteristics of human sperm cryopreservation. Proteomics 2014, 14, 298–310. [Google Scholar] [CrossRef]
- Bogle, O.A.; Kumar, K.; Attardo-Parrinello, C.; Lewis, S.E.M.; Estanyol, J.M.; Ballesca, J.L.; Oliva, R. Identification of protein changes in human spermatozoa throughout the cryopreservation process. Andrology 2017, 5, 10–22. [Google Scholar] [CrossRef]
- He, Y.; Wang, K.; Zhao, X.; Zhang, Y.; Ma, Y.; Hu, J. Differential proteome association study of freeze-thaw damage in ram sperm. Cryobiology 2016, 72, 60–68. [Google Scholar] [CrossRef]
- Pini, T.; Rickard, J.P.; Leahy, T.; Crossett, B.; Druart, X.; de Graaf, S.P. Cryopreservation and egg yolk medium alter the proteome of ram spermatozoa. J. Proteom. 2018, 181, 73–83. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, H.; Hu, C.; Hao, H.; Zhang, J.; Li, K.; Zhao, X.; Qin, T.; Zhao, K.; Zhu, H.; et al. Identification of differentially expressed proteins in fresh and frozenethawed boar spermatozoa by iTRAQcoupled 2D LCeMS/MS. Reproduction 2014, 147, 321–330. [Google Scholar] [CrossRef]
- Kuzmuk, K.N.; Schook, L.B. Pigs as a model of biochemical science. In The Genetics of the Pig, 2nd ed.; Rothschild, M.F., Ruvinsky, A., Eds.; CAB International: Oxfordshire, UK, 2011; pp. 426–444. [Google Scholar]
- Roca, J.; Martinez-Alborcia, M.J.; Gil, M.A.; Parrilla, I.; Martinez, E.A. Dead spermatozoa in raw semen samples impair in vitro fertilization outcomes of frozen-thawed spermatozoa. Fertil. Steril. 2013, 100, 875–881. [Google Scholar] [CrossRef]
- Alkmin, D.V.; Perez-Patiño, C.; Barranco, I.; Parrilla, I.; Vazquez, J.M.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. Boar sperm cryosurvival is better after exposure to seminal plasma from selected fractions than to those from entire ejaculate. Cryobiology 2014, 69, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Rath, D.; Niemann, H. In vitro fertilization of porcine oocytes with fresh and frozen-thawed ejaculated or frozen-thawed epididymal semen obtained from identical boars. Theriogenology 1997, 47, 785–793. [Google Scholar] [CrossRef]
- Holt, W.V.; Medrano, A.; Thurston, L.M.; Watson, P.F. The significance of cooling rates and animal variability for boar sperm cryopreservation: Insights from the cryomicroscope. Theriogenology 2005, 63, 370–382. [Google Scholar] [CrossRef]
- Roca, J.; Hernández, M.; Carvajal, G.; Vázquez, J.M.; Martínez, E.A. Factors influencing boar sperm cryosurvival. J. Anim. Sci. 2006, 84, 2692–2699. [Google Scholar] [CrossRef]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef]
- Vilagran, I.; Castillo, J.; Bonet, S.; Sancho, S.; Yeste, M.; Estanyol, J.M.; Oliva, R. Acrosin-binding protein (ACRBP) and triosephosphate isomerase (TPI) are good markers to predict boar sperm freezing capacity. Theriogenology 2013, 80, 443–450. [Google Scholar] [CrossRef]
- Vilagran, I.; Yeste, M.; Sancho, S.; Casas, I.; Rivera del Álamo, M.M.; Bonet, S. Relationship of sperm small heat-shock protein 10 and voltage-dependent anion channel 2 with semen freezability in boars. Theriogenology 2014, 82, 418–426. [Google Scholar] [CrossRef]
- Guimarães, D.B.; Barros, T.B.; van Tilburg, M.F.; Martins, J.A.M.; Moura, A.A.; Moreno, F.B.; Monteiro-Moreira, A.C.; Moreira, R.A.; Toniolli, R. Sperm membrane proteins associated with the boar semen cryopreservation. Anim. Reprod. Sci. 2017, 183, 27–38. [Google Scholar] [CrossRef]
- Perez-Patiño, C.; Parrilla, I.; Li, J.; Barranco, I.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. The proteome of pig spermatozoa is remodeled during ejaculation. Mol. Cell. Proteom. 2019, 18, 41–50. [Google Scholar] [CrossRef]
- Rodriguez-Martinez, H.; Kvist, U.; Ernerudh, J.; Sanz, L.; Calvete, J.J. Seminal plasma proteins: What role do they play? Am. J. Reprod. Immunol. 2011, 66, 11–22. [Google Scholar] [CrossRef]
- Pini, T.; Leahy, T.; Soleilhavoup, C.; Tsikis, G.; Labas, V.; Combes-Soia, L.; Harichaux, G.; Rickard, J.P.; Druart, X.; de Graaf, S.P. Proteomic Investigation of Ram Spermatozoa and the Proteins Conferred by Seminal Plasma. J. Proteome Res. 2016, 15, 3700–3711. [Google Scholar] [CrossRef]
- Perez-Patiño, C.; Barranco, I.; Parrilla, I.; Valero, M.L.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. Characterization of the porcine seminal plasma proteome comparing ejaculate portions. J. Proteom. 2016, 142, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Westfalewicz, B.; Dietrich, M.A.; Ciereszko, A. Impact of cryopreservation on bull () semen proteome. J. Anim. Sci. 2015, 93, 5240–5253. [Google Scholar] [CrossRef]
- Yoon, S.J.; Rahman, M.S.; Kwon, W.S.; Ryu, D.Y.; Park, Y.J.; Pang, M.G. Proteomic identification of cryostress in epididymal spermatozoa. J. Anim. Sci. Biotechnol. 2016, 7, 67. [Google Scholar] [CrossRef]
- Brewis, I.A.; Gadella, B.M. Sperm surface proteomics: From protein lists to biological function. Mol. Hum. Reprod. 2010, 16, 68–79. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, M.A. The role of proteomics in understanding sperm cell biology. Int. J. Androl. 2008, 31, 295–302. [Google Scholar] [CrossRef]
- Druart, X.; de Graaf, S. Seminal plasma proteomes and sperm fertility. Anim. Reprod. Sci. 2018, 194, 33–40. [Google Scholar] [CrossRef]
- Gurupriya, V.S.; Divyashree, B.C.; Roy, S.C. Cryogenic changes in proteases and antiprotease activities of buffalo (Bubalus bubalis) and cattle (Bos taurus) semen. Theriogenology 2014, 81, 396–402. [Google Scholar] [CrossRef]
- Rich, P.R.; Maréchal, A. The mitochondrial respiratory chain. Essays Biochem. 2010, 47, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, T.; Kaji, Y.; Agatsuma, T.; Niki, T.; Arisawa, M.; Shuto, S.; Ariga, H.; Iguchi-Ariga, S.M. DJ-1 cooperates with PYCR1 in cell protection against oxidative stress. Biochem. Biophys. Res. Commun. 2013, 436, 289–294. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, Y.; Zhao, X.; Yuan, R.; Jiang, H.; Pu, X. Downregulation of DJ-1 Fails to Protect Mitochondrial Complex I Subunit NDUFS3 in the Testes and Contributes to the Asthenozoospermia. Mediat. Inflamm. 2018, 2018, 6136075. [Google Scholar] [CrossRef]
- Moraes, C.R.; Meyers, S. The sperm mitochondrion: Organelle of many functions. Anim. Reprod. Sci. 2018, 194, 71–80. [Google Scholar] [CrossRef]
- Alkmin, D.V.; Parrilla, I.; Tarantini, T.; Del Olmo, D.; Vazquez, J.M.; Martinez, E.A.; Roca, J. Seminal plasma affects sperm sorting in boars. Reprod. Fertil. Dev. 2016, 28, 556–564. [Google Scholar] [CrossRef]
- Pérez-Patiño, C.; Li, J.; Barranco, I.; Martínez, E.A.; Rodriguez-Martínez, H.; Roca, J.; Parrilla, I. The proteome of frozen-thawed pig spermatozoa is dependent on the ejaculate fraction source. Sci. Rep. 2019, 9, 705. [Google Scholar] [CrossRef]
- Shevchenko, A.; Jensen, O.N.; Podtelejnikov, A.V.; Sagliocco, F.; Wilm, M.; Vorm, O.; Mortensen, P.; Shevchenko, A.; Boucherie, H.; Mann, M. Linking genome and proteome by mass spectrometry: Large-scale identification of yeast proteins from two dimensional gels. Proc. Natl. Acad. Sci. USA 1996, 93, 14440–14445. [Google Scholar] [CrossRef] [Green Version]
- Gillet, L.C.; Navarro, P.; Tate, S.; Röst, H.; Selevsek, N.; Reiter, L.; Bonner, R.; Aebersold, R. Targeted data extraction of the MS/MS spectra generated by data-independent acquisition: A new concept for consistent and accurate proteome analysis. Mol. Cell. Proteom. 2012, 11. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]





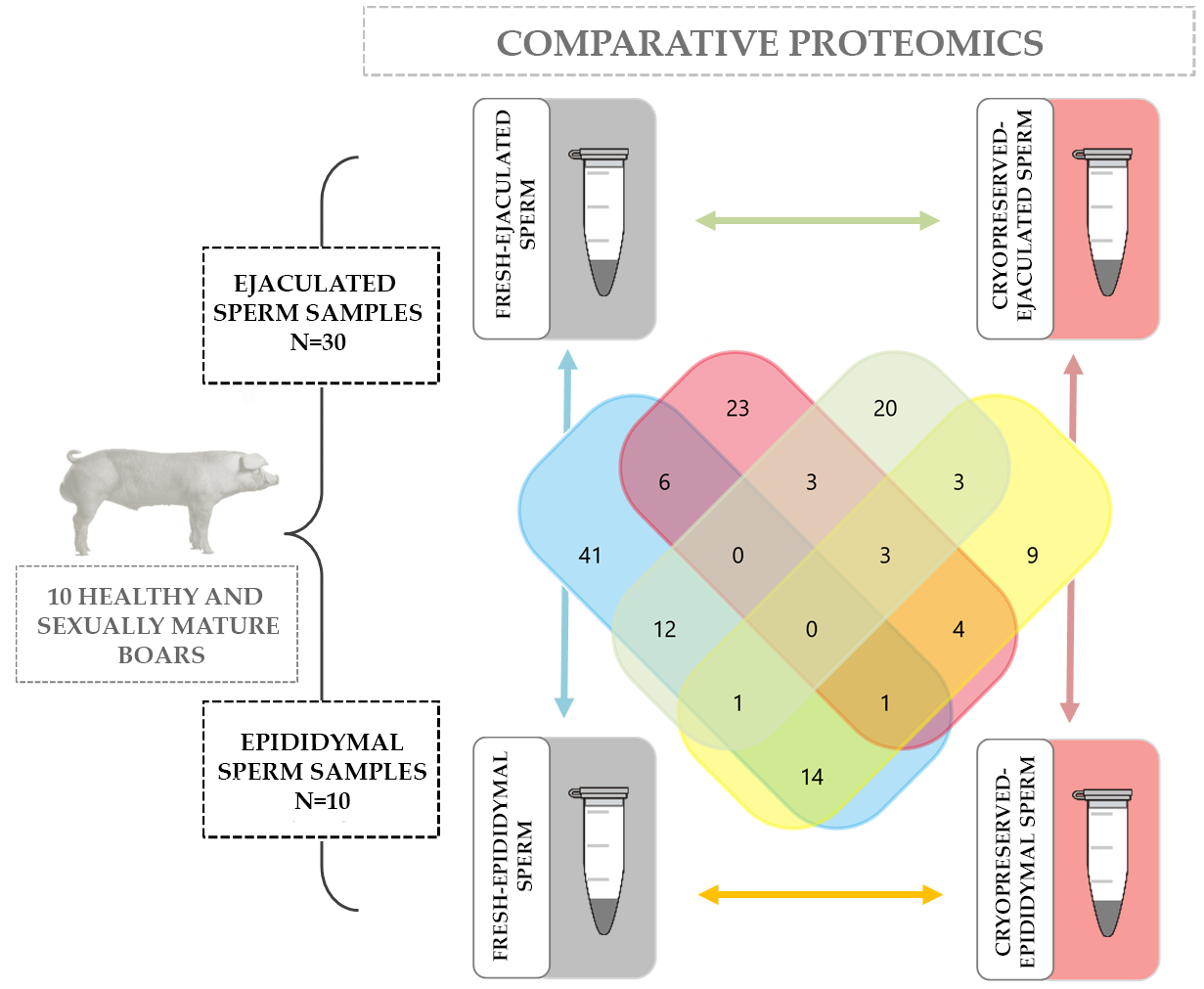{kind=link}
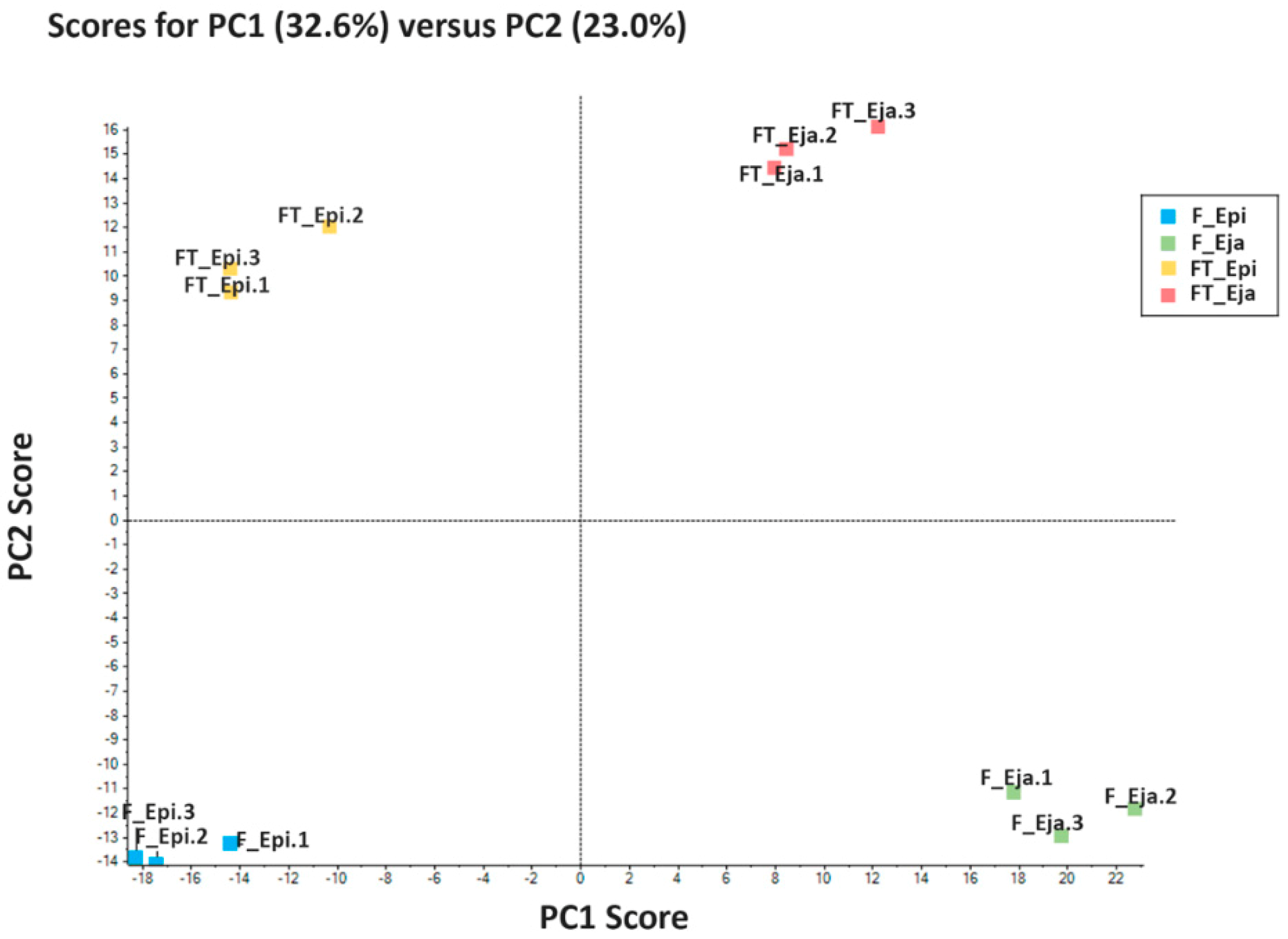{kind=link}
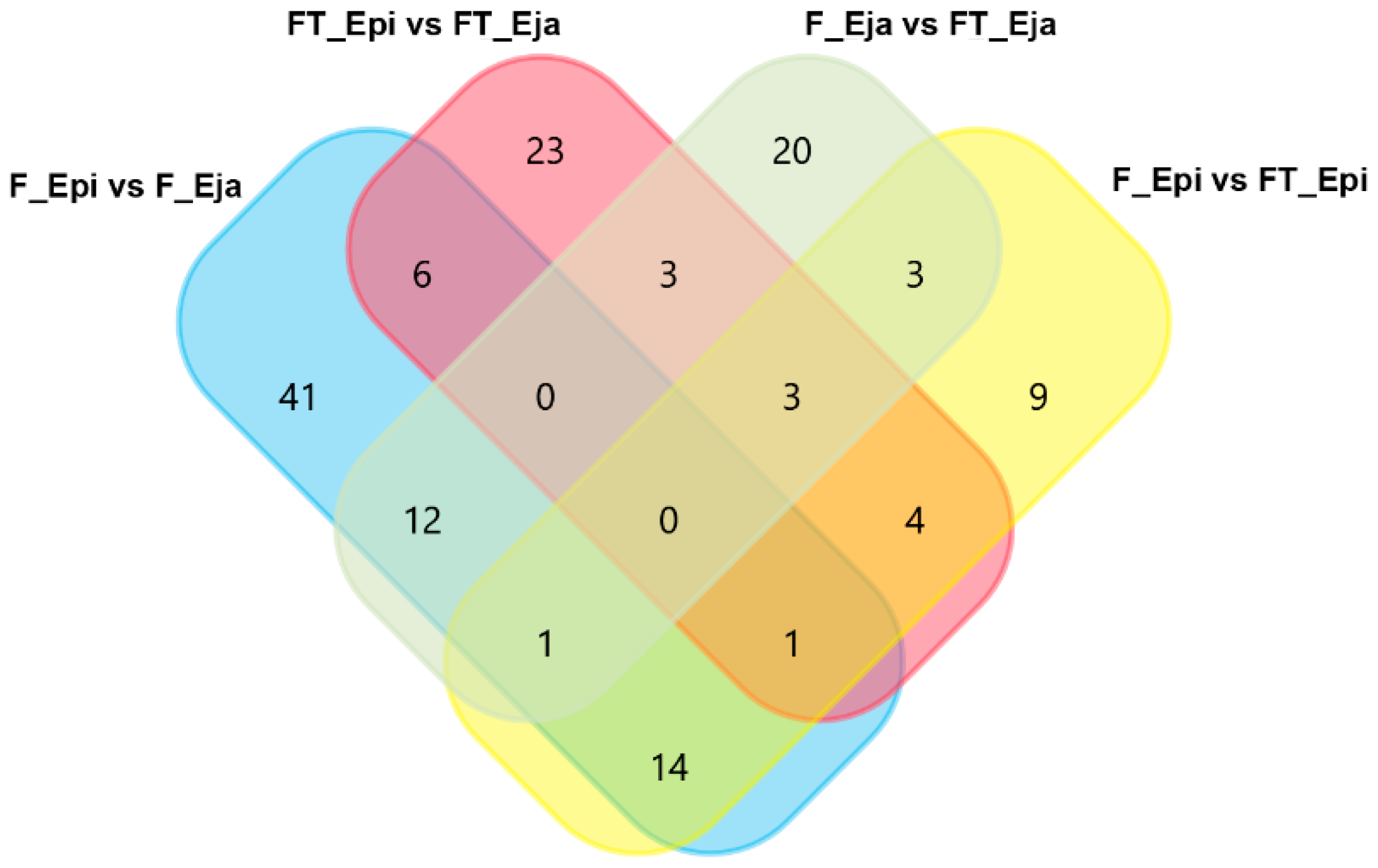{kind=link}
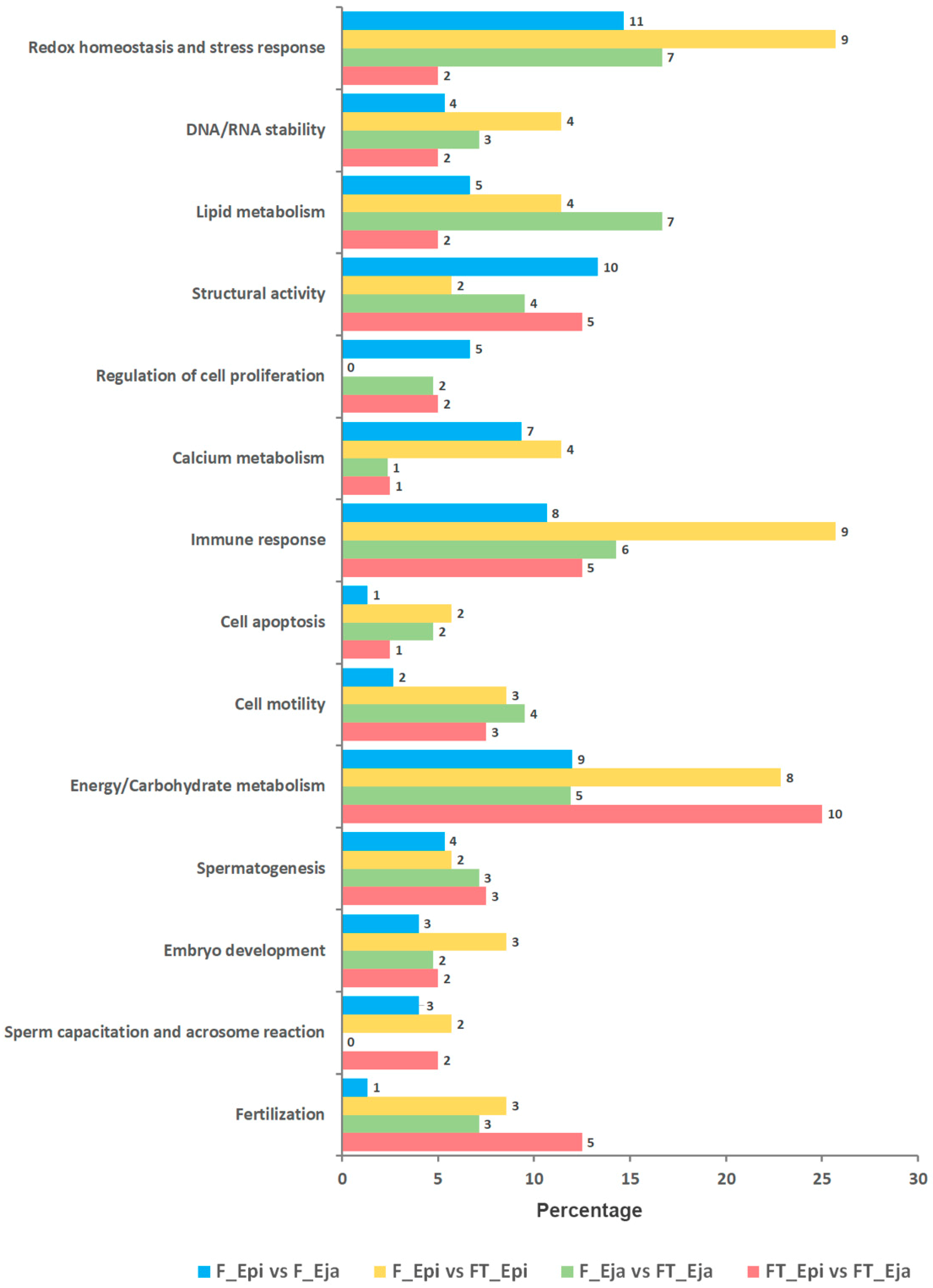{kind=link}
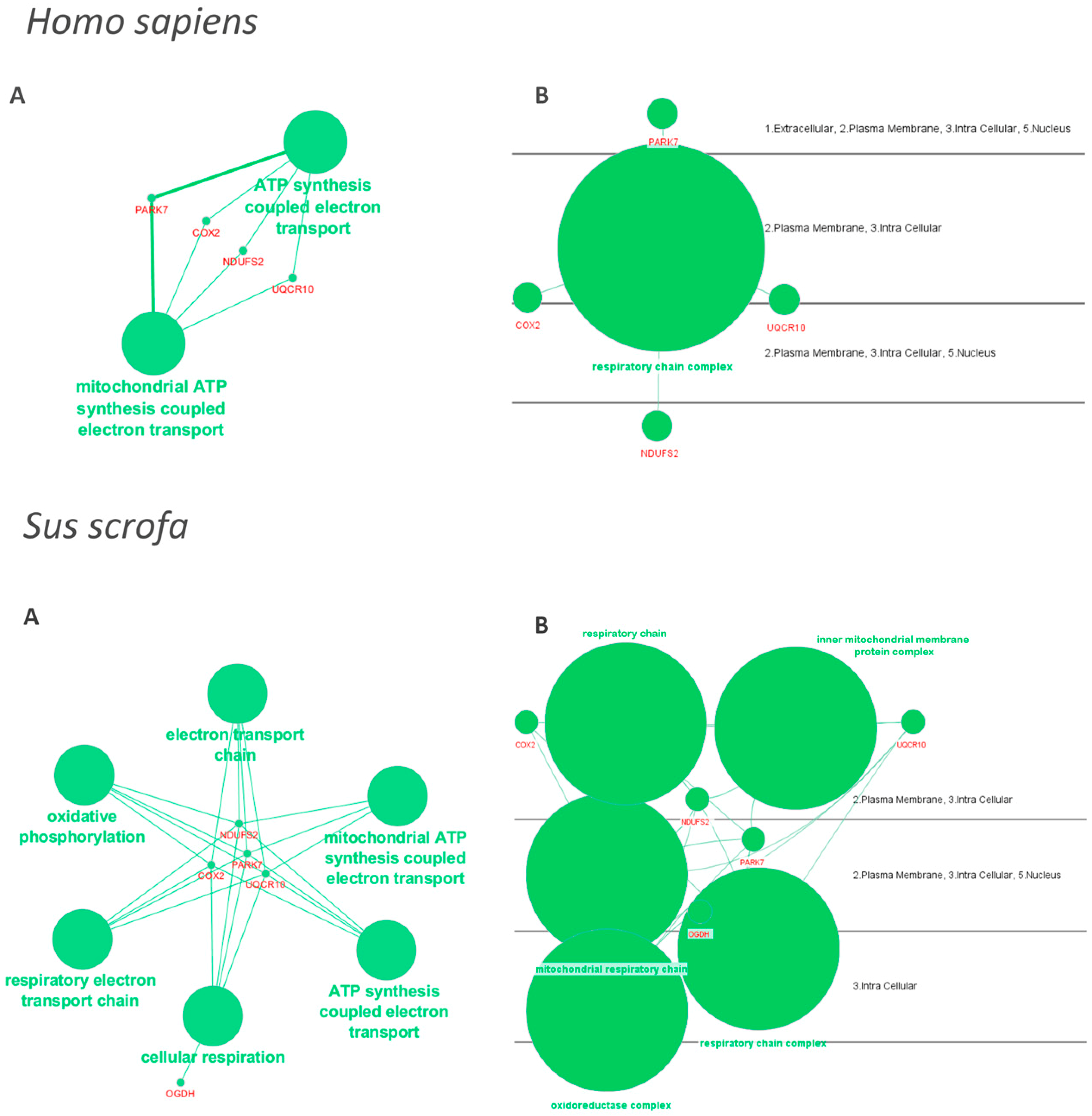{kind=link}
| Sperm Parameters 1 | Sperm Source | |
|---|---|---|
| Cauda Epididymis | Ejaculate | |
| Total motility (%) | 60.50 ± 0.63 a | 46.83 ± 1.04 b |
| Progressive motility (%) | 43.60 ± 0.70 a | 36.23 ± 1.03 b |
| Viability (%) | 62.19 ± 0.85 a | 48.40 ± 0.84 b |
| Viable sperm with high plasma membrane fluidity (%) | 3.44 ± 0.33 a | 7.83 ± 0.36 b |
| Viable sperm with high mitochondria membrane potential (%) | 52.71 ± 0.88 a | 47.99 ± 0.90 b |
| Intracellular ROS2 generation in viable sperm (fluorescent units per 106 viable sperm) | 1826.71 ± 102.04 a | 2778.35 ± 116.84 b |
| Entry Name | Protein Name | Gene Name | FC* log2 Epi vs. Eja |
|---|---|---|---|
| M3YIY8_MUSPF | Solute carrier family 25 member 3 | SLC25A3 | 5.61 |
| Q1RLJ4_PIG | Prepro-beta-defensin 129 protein | DEFB129 | 4.15 |
| F1S086_PIG | Solute carrier family 25 (Aspartate/glutamate carrier), member 12 | SLC25A12 | 2.93 |
| A0A286ZQW4_PIG | Nucleoporin 210 like | NUP210L | 2.83 |
| F1RRA6_PIG | Solute carrier family 25 member 31 | SLC25A31 | 2.66 |
| W5P5P6_SHEEP | Calcium regulated heat stable protein 1 | CARHSP1 | 2.63 |
| L8ISZ2_9CETA | Uncharacterized protein | M91_05957 | 2.51 |
| G1PY44_MYOLU | Saccharopine dehydrogenase (putative) | SCCPDH | 2.46 |
| H9H013_HORSE | Solute carrier family 25 member 1 | SLC25A1 | 2.34 |
| U3CNV1_CALJA | ADP/ATP translocase 3 | SLC25A6 | 2.31 |
| I3LBP3_PIG | Maestro heat like repeat family member 2B | MROH2B | 2.25 |
| I3LR09_PIG | Cell division cycle 14B | CDC14B | 2.22 |
| F1RLC5_PIG | Ferritin | FTMT | 2.18 |
| A0A287BD08_PIG | Uncharacterized protein | _ | 2.06 |
| F1SKK4_PIG | Transmembrane protein 89 | TMEM89 | 1.95 |
| F1S8P1_PIG | Uncharacterized protein | SCCPDH | 1.9 |
| W5PLN5_SHEEP | Uncharacterized protein | _ | 1.9 |
| F1SAS2_PIG | Acyl-CoA synthetase short chain family member 1 | ACSS1 | 1.86 |
| F1RX00_PIG | Amine oxidase | MAOA | 1.84 |
| A0A287AN57_PIG | Lipocalin 12 | LCN12 | 1.83 |
| I3LPR1_PIG | Cancer/testis antigen 83 | CT83 | 1.81 |
| A0A287ATC1_PIG | Uncharacterized protein | _ | 1.75 |
| A0A287BJG7_PIG | Uncharacterized protein | _ | 1.74 |
| Q2I373_PIG | Fascin | FSCN3 | 1.73 |
| F1RVC1_PIG | Glycerol kinase 2 | GK2 | 1.71 |
| H0V2U0_CAVPO | Frataxin | FXN | 1.66 |
| A0A287A9I6_PIG | SAMM50 sorting and assembly machinery component | SAMM50 | 1.65 |
| A0A287BSC3_PIG | Alkaline phosphatase | ALPL | 1.62 |
| F1S7L6_PIG | UBX domain protein 6 | UBXN6 | 1.62 |
| F1SN95_PIG | Nucleoporin 155 | NUP155 | 1.6 |
| M3VZT4_FELCA | Dynein axonemal intermediate chain 1 | DNAI1 | 1.57 |
| A0A287D4X0_ICTTR | Dynein light chain roadblock | DYNLRB2 | −2.02 |
| I6R469_PIG | Calcium binding tyrosine-(Y)-phosphorylation regulated transcript variant 3 | CABYR | −2.17 |
| F1RG35_PIG | STIP1 homology and U-box containing protein 1 | STUB1 | −2.42 |
| A0A287BNS1_PIG | Chromosome 10 open reading frame 82 | C10orf82 | −2.45 |
| F1S137_PIG | Pro-epidermal growth factor | EGF | −3.7 |
| A0A2C9F3H7_PIG | Dipeptidyl peptidase 4 | DPP4 | −3.83 |
| I3LH70_PIG | Collagen type XVIII alpha 1 chain | COL18A1 | −3.95 |
| Q8WNW8_PIG | Nexin-1 | PN-1 | −6.99 |
| I7HJH6_PIG | Seminal plasma sperm motility inhibitor | AQN-3 SPMI | −7.01 |
| A0A286ZY95_PIG | Fibronectin 1 | FN1 | −9.54 |
| Entry Name | Protein Name | Gene Name | FC* log2 | |
|---|---|---|---|---|
| F vs. FT_Epi | F vs. FT_Eja | |||
| K7GMV8_PIG | Ectonucleotide pyrophosphatase/phosphodiesterase 3 | ENPP3 | 3.74 | _ |
| M3W2V1_FELCA | Thioredoxin | TXN | 3.6 | _ |
| E1CAJ5_PIG | Protein disulfide-isomerase | grp-58 | 3.15 | _ |
| D0G6X8_PIG | Beta-hexosaminidase | HEXB | 2.9 | _ |
| W5P708_SHEEP | Actinin alpha 4 | ACTN4 | 2.87 | _ |
| G9F6X8_PIG | Protein disulfide-isomerase | P4HB | 2.74 | _ |
| F7HXH1_CALJA | Phosphoglycerate mutase | PGAM2 | 2.27 | _ |
| V9HWB4_HUMAN | Epididymis secretory sperm binding protein Li 89n | HEL-S-89n | 2.23 | _ |
| K7GRY0_PIG | Ubiquitin like modifier activating enzyme 1 | UBA1 | 2.08 | _ |
| M3WST2_FELCA | Glutaminyl-peptide cyclotransferase | QPCT | 3.26 | 2.6 |
| A0A287A5M7_PIG | Aldose reductase | AKR1B1 | 1.74 | 1.97 |
| F1SAT2_PIG | Cystatin | CST11 | 1.52 | 1.58 |
| MSMB_PIG | Beta-microseminoprotein | MSMB PSP94 | _ | 2.85 |
| A0A287AXJ7_PIG | Inositol-3-phosphate synthase 1 | ISYNA1 | _ | 2.82 |
| F7E460_HORSE | Uncharacterized protein | RELCH | _ | 2.64 |
| A0A287BM88_PIG | Lipocalin 8 | LCN8 | _ | 2.32 |
| Q307R2_RABIT | Peptidyl-prolyl cis-trans isomerase (Fragment) | PPIA | _ | 2.23 |
| A0A1B2TT55_PIG | Aspartate aminotransferase | GOT1 | _ | 1.97 |
| U3F9K8_CALJA | 26S proteasome non-ATPase regulatory subunit 1 isoform 1 | PSMD1 | _ | 1.83 |
| Q0R678_PIG | DJ-1 protein | PARK7 | _ | 1.71 |
| Q8WNR3_PIG | Arylsulfatase A | AS-A | _ | 1.52 |
| A0A287ARR1_PIG | Ferritin | FTH1 | _ | 1.51 |
| Q2EN79_PIG | Ubiquinol-cytochrome c reductase complex O | UQCR10 | _ | −1.69 |
| F1SDZ2_PIG | Uncharacterized protein | GALC | _ | −1.77 |
| A0A0K0KW08_PIG | L-lactate dehydrogenase | LDHAL6B | _ | −1.81 |
| F1S1A8_PIG | NADH:ubiquinone oxidoreductase core subunit S2 | NDUFS2 | _ | −1.92 |
| F1RWZ8_PIG | Dual specificity phosphatase 21 | DUSP21 | _ | −1.97 |
| I3LNF2_PIG | Dynein axonemal heavy chain 1 | DNAH1 | _ | −2.2 |
| F1S0P1_PIG | Regulator of G protein signaling 22 | RGS22 | _ | −2.22 |
| K9IVI1_PIG | 2-oxoglutarate dehydrogenase, mitochondrial | OGDH | _ | −2.45 |
| V5KX18_PIG | Cytochrome c oxidase subunit 2 | COX2 | _ | −2.51 |
| G3QF37_GORGO | Chromosome 9 open reading frame 84 | C9ORF84 | _ | −3.8 |
| Entry Name | Protein Name | Gene Name | FC* log2 Epi vs. Eja |
|---|---|---|---|
| A0A287BPZ5_PIG | Uncharacterized protein | IQCN | 3.08 |
| A0A2I3T063_PANTR | Uncharacterized protein | _ | 2.68 |
| A0A287BCS4_PIG | Pitrilysin metallopeptidase 1 | PITRM1 | 2.63 |
| I3LJA4_PIG | Uncharacterized protein | _ | 2.15 |
| A0A286ZN09_PIG | Uncharacterized protein | _ | 1.9 |
| A0A287AI93_PIG | Uncharacterized protein | IQCN | 1.8 |
| W5Q086_SHEEP | cAMp-dependent protein kinase type I-alpha regulatory subunit | PRKAR1A | −1.6 |
| ENPL_PIG | Endoplasmin | HSP90B1 | −1.61 |
| G1M5S6_AILME | Cytochrome b-c1 complex subunit 6 | LOC100469420 | −1.77 |
| F1S1R1_PIG | Cylicin 1 OS = Sus scrofa | CYLC1 | −2.22 |
| I3LNH3_PIG | Neutral alpha-glucosidase AB | GANAB | −2.28 |
| A7VK02_PIG | Transmembrane protease, serine 2 | TMPRSS2 | −2.32 |
| W5Q770_SHEEP | Uncharacterized protein | _ | −2.42 |
| W5QAX3_SHEEP | Actin gamma 1 | ACTG1 | −2.74 |
| A0A286XIX4_CAVPO | Fibronectin 1 | FN1 | −3.29 |
| A0A250Y012_CASCN | Glia-derived nexin | SERPINE2 | −3.3 |
| A0A287B423_PIG | Uncharacterized protein | SPESP1 | −3.52 |
| M3Z880_MUSPF | Peptidyl-prolyl cis-trans isomerase | PPIA | −3.64 |
| Q29057_PIG | Glutathione S-transferase | GST | −3.67 |
| F1PLM3_CANLF | Tubulin alpha chain | TUBA1A | −4.18 |
| F1RT83_PIG | Uncharacterized protein | SDCBP | −4.91 |
| Q4R0H6_PIG | Spermadhesin PSP-I | PSP-I | −5.72 |
| A0A0A8IK66_PIG | Heparin-binding protein WGA16 | WGA16 | −6.58 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez-Patiño, C.; Barranco, I.; Li, J.; Padilla, L.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J.; Parrilla, I. Cryopreservation Differentially Alters the Proteome of Epididymal and Ejaculated Pig Spermatozoa. Int. J. Mol. Sci. 2019, 20, 1791. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071791
Perez-Patiño C, Barranco I, Li J, Padilla L, Martinez EA, Rodriguez-Martinez H, Roca J, Parrilla I. Cryopreservation Differentially Alters the Proteome of Epididymal and Ejaculated Pig Spermatozoa. International Journal of Molecular Sciences. 2019; 20(7):1791. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071791
Chicago/Turabian StylePerez-Patiño, Cristina, Isabel Barranco, Junwei Li, Lorena Padilla, Emilio A. Martinez, Heriberto Rodriguez-Martinez, Jordi Roca, and Inmaculada Parrilla. 2019. "Cryopreservation Differentially Alters the Proteome of Epididymal and Ejaculated Pig Spermatozoa" International Journal of Molecular Sciences 20, no. 7: 1791. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071791