Characterization of a New Reconstructed Full Thickness Skin Model, T-Skin™, and its Application for Investigations of Anti-Aging Compounds


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of T-Skin™ Model
2.1.1. Time Course of T-Skin™ Model Reconstruction
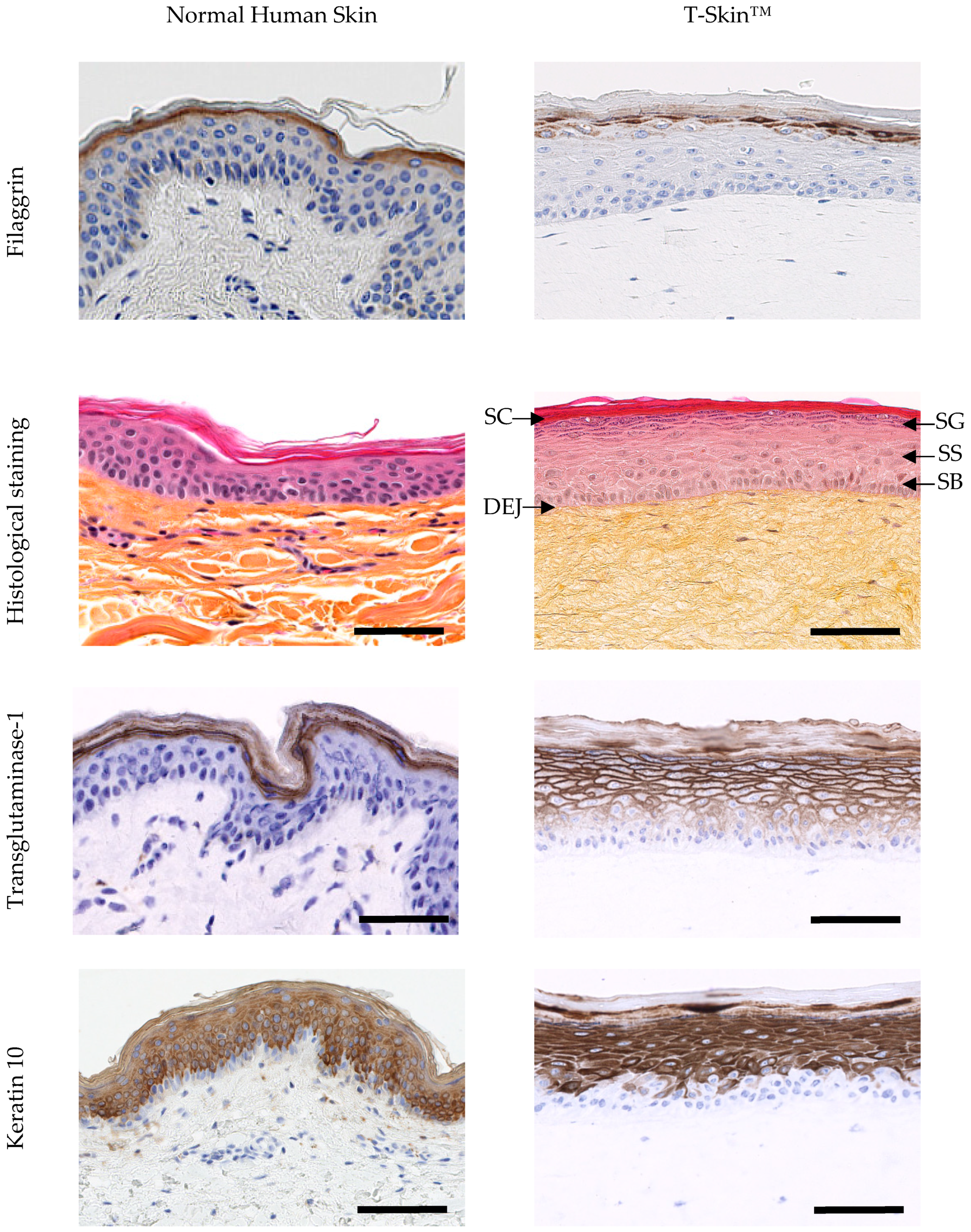
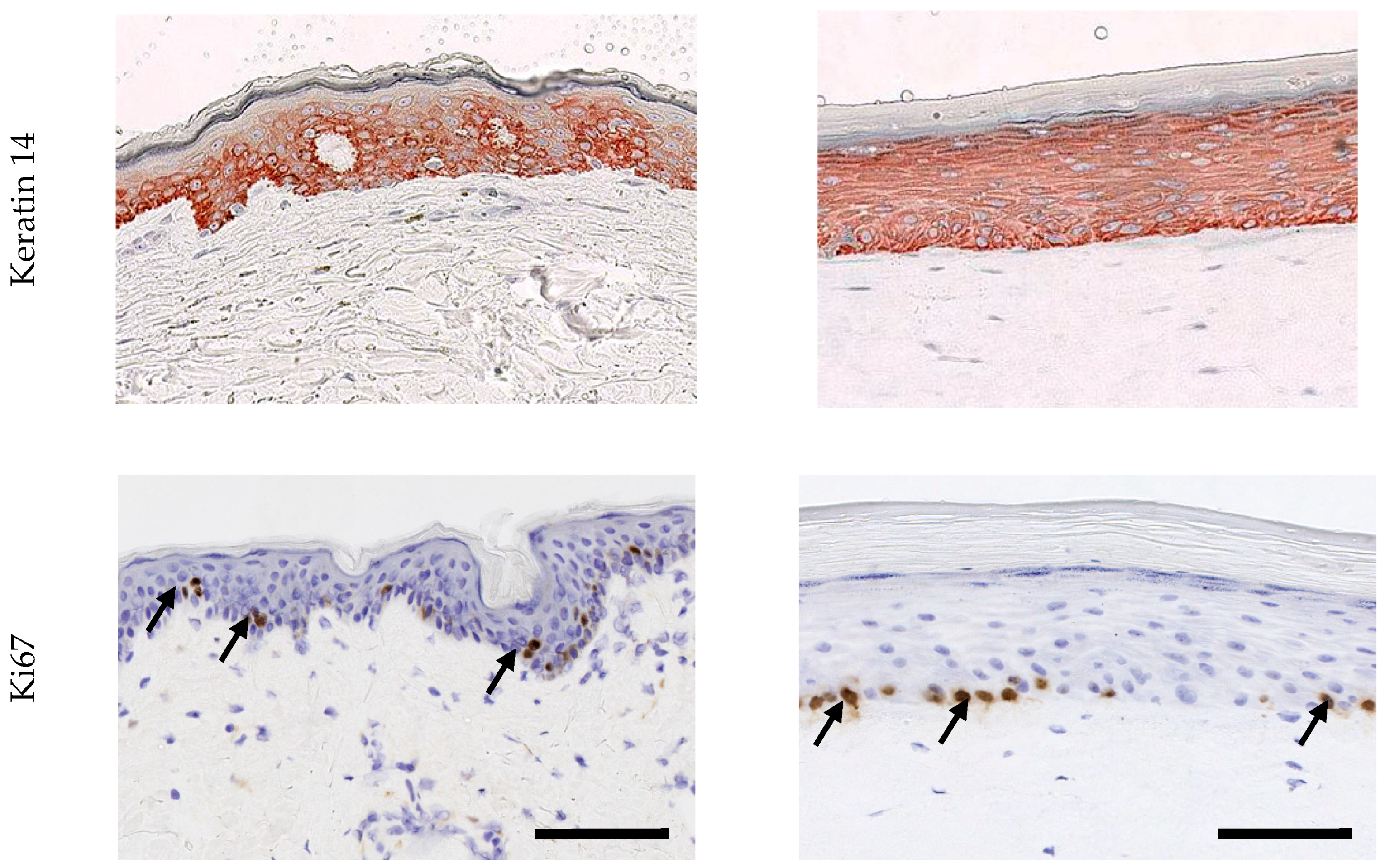
2.1.2. Epidermal Characterization
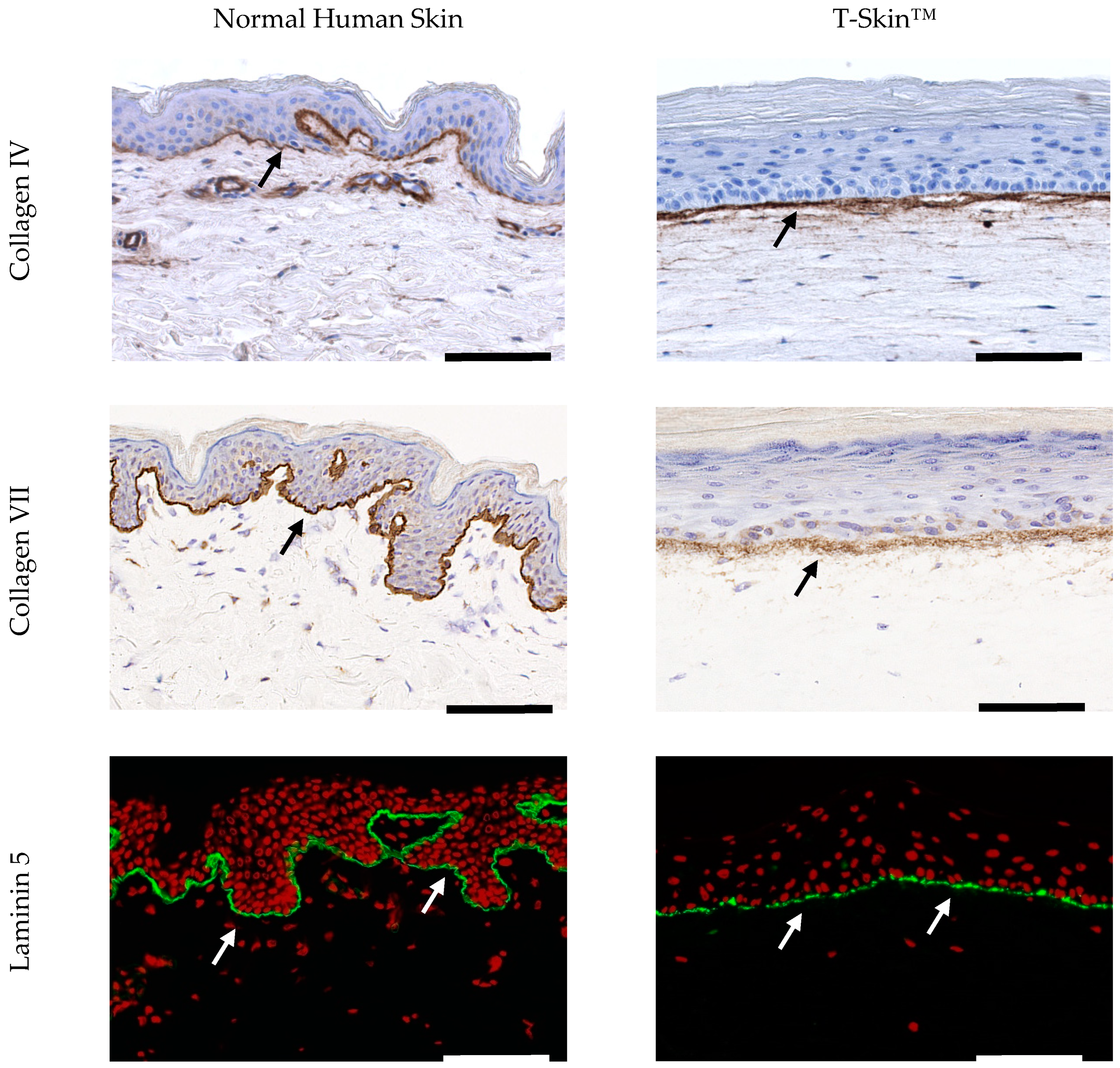
2.1.3. Dermo–Epidermal Characterization
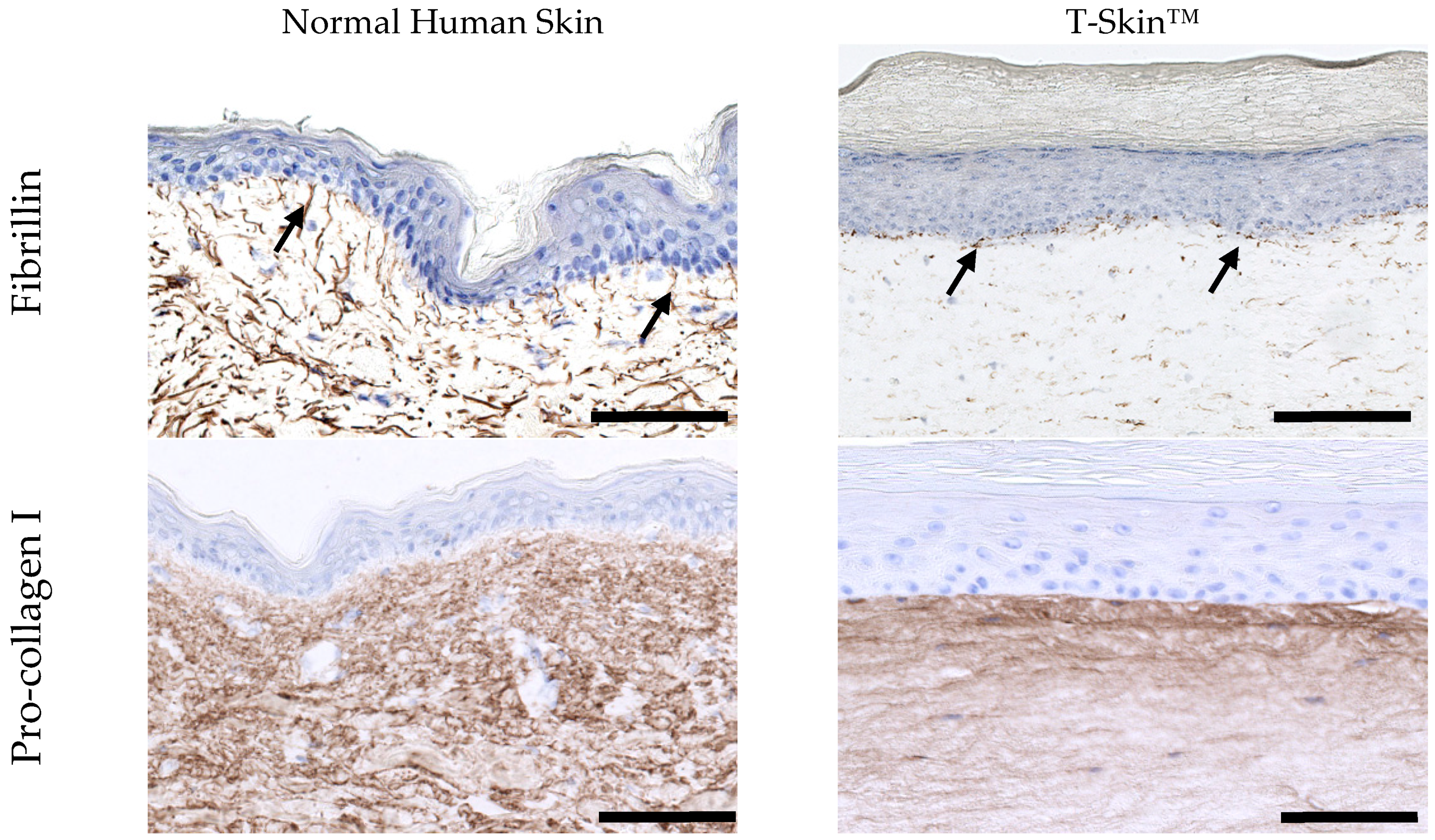
2.1.4. Dermal Characterization
2.2. Application of T-Skin™ to Anti-Aging Investigations
2.2.1. Effect of Retinol on Epidermal Markers
2.2.2. Effect of Vitamin C on Dermo–Epidermal and Dermal Markers
3. Discussion
4. Materials and Methods
4.1. T-Skin™ Preparation
4.2. Histological and Immunohistological Characterization
4.3. Cytotoxicity Measurement
4.4. Effect on Skin Aging Markers
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NHS | normal human skin |
| DEJ | dermo-epidermal junction |
| ECM | extracellular matrix |
| RHE | reconstructed human epidermal |
| FFPE | formalin-fixed paraffin-embedded block |
| SC | stratum corneum |
| SG | stratum granulosum |
| SS | stratum spinosum |
| SB | stratum basal |
References
- Burgeson RE, C.A. The dermal-epidermal junction. Curr. Opin. Cell Biol. 1997, 9, 651–658. [Google Scholar] [CrossRef]
- Green, H.; Kehinde, O.; Thomas, J. Growth of cultured human epidermal cells into multiple epithelia suitable for grafting. Proc. Natl. Acad. Sci. USA 1979, 76, 5665–5668. [Google Scholar] [CrossRef] [PubMed]
- Prunieras, M. Epidermal cell cultures as models for living epidermis. J. Investig. Dermatol. 1979, 73, 135–137. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.; Sher, S.; Hull, B.; Merrill, C.; Rosen, S.; Chamson, A.; Asselineau, D.; Dubertret, L.; Coulomb, B.; Lapiere, C.; et al. The reconstitution of living skin. J. Investig. Dermatol. 1983, 81, 2–10. [Google Scholar] [CrossRef]
- OECD. Test No. 431: In Vitro Skin Corrosion: Reconstructed Human Epidermis (RHE) Test Method; OECD Publishing: Paris, France, 2016; ISBN 9789264264618. [Google Scholar]
- OECD. Test No. 439: In Vitro Skin Irritation: Reconstructed Human Epidermis Test Method; OECD Publishing: Paris, France, 2015; ISBN 9789264242845. [Google Scholar]
- Bernerd, F.; Vioux, C.; Asselineau, D. Evaluation of the protective effect of sunscreens on in vitro reconstructed human skin exposed to UVB or UVA irradiation. Photochem. Photobiol. 2000, 71, 314–320. [Google Scholar] [CrossRef]
- Bernerd, F.; Asselineau, D. An organotypic model of skin to study photodamage and photoprotection in vitro. J. Am. Acad. Dermatol. 2008, 58, 155–159. [Google Scholar] [CrossRef]
- Pageon, H.; Zucchi, H.; Asselineau, D. Distinct and complementary roles of papillary and reticular fibroblasts in skin morphogenesis and homeostasis. Eur. J. Dermatol. 2012, 22, 324–332. [Google Scholar]
- Pageon, H.; Zucchi, H.; Rousset, F.; Monnier, V.M.; Asselineau, D. Skin aging by glycation: Lessons from the reconstructed skin model. Clin. Chem. Lab. Med. 2014, 52, 169–174. [Google Scholar] [CrossRef]
- Pageon, H.; Zucchi, H.; Dai, Z.; Sell, D.R.; Strauch, C.M.; Monnier, V.M.; Asselineau, D. Biological Effects Induced by Specific Advanced Glycation End Products in the Reconstructed Skin Model of Aging. BioRes. Open Access 2015, 4, 54–64. [Google Scholar] [CrossRef]
- Mewes, K.R.; Raus, M.; Bernd, A.; Zoller, N.N.; Sattler, A.; Graf, R. Elastin expression in a newly developed full-thickness skin equivalent. Skin Pharmacol. Physiol. 2007, 20, 85–95. [Google Scholar] [CrossRef]
- Hamada, H.; Shimoda, K.; Horio, Y.; Onoa, T.; Hosoda, R.; Nakayama, N.; Uranod, K. Pterostilbene and Its Glucoside Induce Type XVII Collagen Expression. Nat. Prod. Commun. 2017, 12, 85–86. [Google Scholar] [CrossRef]
- Stellavato, A.; Corsuto, L.; D’Agostino, A.; La Gatta, A.; Diana, P.; Bernini, P.; De Rosa, M.; Schiraldi, C. Hyaluronan Hybrid Cooperative Complexes as a Novel Frontier for Cellular Bioprocesses Re-Activation. PLoS ONE 2016, 11, e0163510. [Google Scholar] [CrossRef]
- Ackermann, K.; Borgia, S.L.; Korting, H.C.; Mewes, K.R.; Schafer-Korting, M. The Phenion full-thickness skin model for percutaneous absorption testing. Skin Pharmacol. Physiol. 2010, 23, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Batheja, P.; Song, Y.; Wertz, P.; Michniak-Kohn, B. Effects of growth conditions on the barrier properties of a human skin equivalent. Pharm. Res. 2009, 26, 1689–1700. [Google Scholar] [CrossRef] [PubMed]
- Meloni, M.; Farina, A.; de Servi, B. Molecular modifications of dermal and epidermal biomarkers following UVA exposures on reconstructed full-thickness human skin. Photochem. Photobiol. Sci. 2010, 9, 439–447. [Google Scholar] [CrossRef]
- Ding, X.; Cheng, L.; Chen, W.; Zhou, F.; Dou, X.; Zhang, B.; Zhu, B. Integrative analysis of gene expression in response to low-dose ionizing radiation in a human skin model. Med. Oncol. 2015, 32, 621. [Google Scholar] [CrossRef] [PubMed]
- Yunis, R.; Albrecht, H.; Kalanetra, K.M.; Wu, S.; Rocke, D.M. Genomic characterization of a three-dimensional skin model following exposure to ionizing radiation. J. Radiat. Res. 2012, 53, 860–875. [Google Scholar] [CrossRef] [Green Version]
- Jackh, C.; Blatz, V.; Fabian, E.; Guth, K.; van Ravenzwaay, B.; Reisinger, K.; Landsiedel, R. Characterization of enzyme activities of Cytochrome P450 enzymes, Flavin-dependent monooxygenases, N-acetyltransferases and UDP-glucuronyltransferases in human reconstructed epidermis and full-thickness skin models. Toxicol In Vitro 2011, 25, 1209–1214. [Google Scholar] [CrossRef]
- Reisinger, K.; Blatz, V.; Brinkmann, J.; Downs, T.R.; Fischer, A.; Henkler, F.; Hoffmann, S.; Krul, C.; Liebsch, M.; Luch, A.; et al. Validation of the 3D Skin Comet assay using full thickness skin models: Transferability and reproducibility. Mutat. Res. 2018, 827, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Schmid, T.E.; Dollinger, G.; Hable, V.; Greubel, C.; Zlobinskaya, O.; Michalski, D.; Molls, M.; Roper, B. Relative biological effectiveness of pulsed and continuous 20 MeV protons for micronucleus induction in 3D human reconstructed skin tissue. Radiother. Oncol. 2010, 95, 66–72. [Google Scholar] [CrossRef]
- Silva, R.; Ferreira, H.; Matama, T.; Gomes, A.C.; Cavaco-Paulo, A. Wound-healing evaluation of entrapped active agents into protein microspheres over cellulosic gauzes. Biotechnol. J. 2012, 7, 1376–1385. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.; Gomes, A.C.; Cavaco-Paulo, A. Novel silk fibroin/elastin wound dressings. Acta Biomater. 2012, 8, 3049–3060. [Google Scholar] [CrossRef] [Green Version]
- Kuchler, S.; Wolf, N.B.; Heilmann, S.; Weindl, G.; Helfmann, J.; Yahya, M.M.; Stein, C.; Schafer-Korting, M. 3D-wound healing model: Influence of morphine and solid lipid nanoparticles. J. Biotechnol. 2010, 148, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Orlov, M.D.; Chernyavsky, A.I.; Arredondo, J.; Grando, S.A. Synergistic actions of pemphigus vulgaris IgG, Fas-ligand and tumor necrosis factor-alpha during induction of basal cell shrinkage and acantholysis. Autoimmunity 2006, 39, 557–562. [Google Scholar] [CrossRef]
- Lee, S.; Dong, D.X.; Jindal, R.; Maguire, T.; Mitra, B.; Schloss, R.; Yarmush, M. Predicting full thickness skin sensitization using a support vector machine. Toxicol In Vitro 2014, 28, 1413–1423. [Google Scholar] [CrossRef] [Green Version]
- Asselineau, D.; Ricois, S.; Pageon, H.; Zucchi, H. Reconstructed Skin to Create In Vitro Flexible Models of Skin Aging: New Results and Prospects. In Textbook of Aging Skin; Springer: Heidelberg, Berlin, 2017; pp. 1203–1228. [Google Scholar]
- Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Makrantonaki, E.; Zouboulis, C.C. Skin anti-aging strategies. Derm. Endocrinol. 2012, 4, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Telang, P.S. Vitamin C in dermatology. Indian Dermatol. Online J. 2013, 4, 143–146. [Google Scholar] [CrossRef]
- Farris, P.K. Topical Vitamin C: A Useful Agent for Treating Photoaging and Other Dermatologic Conditions. Dermatol. Surg. 2005, 31, 814–817. [Google Scholar] [CrossRef]
- Haftek, M.; Mac-Mary, S.; Bitoux MA, L.; Creidi, P.; Seité, S.; Rougier, A.; Humbert, P. Clinical, biometric and structural evaluation of the long-term effects of a topical treatment with ascorbic acid and madecassoside in photoaged human skin. J. Investig. Dermatol. 2001, 116, 853–859. [Google Scholar] [CrossRef]
- Pullar, J.M.; Carr, A.C.; Vissers, M. The Roles of Vitamin C in Skin Health. Nutrients 2017, 9, 866. [Google Scholar] [CrossRef]
- Purkis, P.E.; Steel, J.B.; Mackenzie, I.C.; Nathrath, W.B.; Leigh, I.M.; Lane, E.B. Antibody markers of basal cells in complex epithelia. J. Cell Sci. 1990, 97, 39–50. [Google Scholar] [PubMed]
- Pruniéras, M.; Regnier, M.; Woodley, D. Methods for cultivation of keratinocytes with an air-liquid interface. J. Investig. Dermatol. 1983, 81, 28–33. [Google Scholar] [CrossRef]
- Heenen, M.; Thiriar, S.; Noël, J.C.; Galand, P. Ki-67 Immunostaining of Normal Human Epidermis: Comparison with 3H-Thymidine Labelling and PCNA Immunostaining. Dermatology 1998, 197, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Lippens, S.; Denecker, G.; Ovaere, P.; Vandenabeele, P.; Declercq, W. Death penalty for keratinocytes: Apoptosis versus cornification. Cell Death Differ. 2005, 12, 1497–1508. [Google Scholar] [CrossRef] [PubMed]
- Nemes, Z.; Steinert, P.M. Bricks and mortar of the epidermal barrier. Exp. Mol. Med. 1999, 31, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef]
- Iizuka, R.; Chiba, K.; Imajoh-Ohmi, S. A novel approach for the detection of proteolytically activated transglutaminase 1 in epidermis using cleavage site-directed antibodies. J. Investig. Dermatol. 2003, 121, 457–464. [Google Scholar] [CrossRef]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens—Structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef]
- Shapiro, S.D.; Endicott, S.K.; Province, M.A.; Pierce, J.A.; Campbell, E.J. Marked longevity of human lung parenchymal elastic fibers deduced from prevalence of D-aspartate and nuclear weapons-related radiocarbon. J. Clin. Investig. 1991, 87, 1828–1834. [Google Scholar] [CrossRef]
- Cotta-Pereira, G.; Rodrigo, G.; Bittencourt-Sampaio, S. Oxytalan, elaunin, and elastic fibers in the human skin. J. Investig. Dermatol. 1976, 66, 143–148. [Google Scholar] [CrossRef]
- Haynes, S.L.; Shuttleworth, C.A.; Kielty, C.M. Keratinocytes express fibrillin and assemble microfibrils:implications for dermal matrix organization. Br. J. Dermatol. 1997, 137, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Marionnet, C.; Pierrard, C.; Vioux-Chagnoleau, C.; Sok, J.; Asselineau, D.; Bernerd, F. Interactions between fibroblasts and keratinocytes in morphogenesis of dermal epidermal junction in a model of reconstructed skin. J. Investig. Dermatol. 2006, 126, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Fleischmajer, R.; Utani, A.; MacDonald, E.D.; Perlish, J.S.; Pan, T.C.; Chu, M.L.; Nomizu, M.; Ninomiya, Y.; Yamada, Y. Initiation of skin basement membrane formation at the epidermodermal interface involves assembly of laminins through binding to cell membrane receptors. J. Cell Sci. 1998, 111, 1929–1940. [Google Scholar] [PubMed]
- Mukherjee, S.; Date, A.; Patravale, V.; Korting, H.C.; Roeder, A.; Weindl, G. Retinoids in the treatment of skin aging: An overview of clinical efficacy and safety. Clin. Interv. Aging 2006, 1, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Voorhees, J.J. Molecular mechanisms of retinoid actions in skin. FASEB J. 1996, 10, 1002–1013. [Google Scholar] [CrossRef] [PubMed]
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased Collagen Production in Chronologically Aged Skin: Roles of Age-Dependent Alteration in Fibroblast Function and Defective Mechanical Stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef] [PubMed]
- Nusgens, B.V.; Humbert, P.; Rougier, A.; Colige, A.C.; Haftek, M.; Lambert, C.A.; Richard, A.; Creidi, P.; Lapière, C.M. Topically applied vitamin C enhances the mRNA level of collagens I and III, their processing enzymes and tissue inhibitor of matrix metalloproteinase 1 in the human dermis. J. Investig. Dermatol. 2001, 116, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Feru, J.; Delobbe, E.; Ramont, L.; Brassart, B.; Terryn, C.; Dupont-Deshorgue, A.; Garbar, C.; Monboisse, J.C.; Maquart, F.X.; Brassart-Pasco, S. Aging decreases collagen IV expression in vivo in the dermo-epidermal junction and in vitro in dermal fibroblasts: Possible involvement of TGF-β1. Eur. J. Dermatol. 2016, 26, 350–360. [Google Scholar]
- Marionnet, C.; Vioux-Chagnoleau, C.; Pierrard, C.; Sok, J.; Asselineau, D.; Bernerd, F. Morphogenesis of dermal–epidermal junction in a model of reconstructed skin: Beneficial effects of vitamin C. Exp. Dermatol. 2006, 15, 625–633. [Google Scholar] [CrossRef]
- Boyce, S.T.; Supp, A.P.; Swope, V.B.; Warden, G.D. Vitamin C Regulates Keratinocyte Viability, Epidermal Barrier, and Basement Membrane In Vitro, and Reduces Wound Contraction After Grafting of Cultured Skin Substitutes. J. Investig. Dermatol. 2002, 118, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Suwabe, H.; Serizawa, A.; Kajiwara, H.; Ohkido, M.; Tsutsumi, Y. Degenerative processes of elastic fibers in sun-protected and sun-exposed skin: Immunoelectron microscopic observation of elastin, fibrillin-1, amyloid P component, lysozyme and alpha(1)-antitrypsin. Pathol. Int. 1999, 49, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.; Michelet, J.F.; Simonetti, L.; Fiat, F.; Garrigues, A.; Potter, A.; Segot, E.; Watson, R.E.; Griffiths, C.E.; de Lacharrière, O. In vitro and in vivo studies with tetra-hydro-jasmonic acid (LR2412) reveal its potential to correct signs of skin ageing. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 415–423. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bataillon, M.; Lelièvre, D.; Chapuis, A.; Thillou, F.; Autourde, J.B.; Durand, S.; Boyera, N.; Rigaudeau, A.-S.; Besné, I.; Pellevoisin, C. Characterization of a New Reconstructed Full Thickness Skin Model, T-Skin™, and its Application for Investigations of Anti-Aging Compounds. Int. J. Mol. Sci. 2019, 20, 2240. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092240
Bataillon M, Lelièvre D, Chapuis A, Thillou F, Autourde JB, Durand S, Boyera N, Rigaudeau A-S, Besné I, Pellevoisin C. Characterization of a New Reconstructed Full Thickness Skin Model, T-Skin™, and its Application for Investigations of Anti-Aging Compounds. International Journal of Molecular Sciences. 2019; 20(9):2240. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092240
Chicago/Turabian StyleBataillon, Michel, Damien Lelièvre, Adeline Chapuis, Fabienne Thillou, Jean Baptiste Autourde, Steven Durand, Nathalie Boyera, Anne-Sophie Rigaudeau, Isabelle Besné, and Christian Pellevoisin. 2019. "Characterization of a New Reconstructed Full Thickness Skin Model, T-Skin™, and its Application for Investigations of Anti-Aging Compounds" International Journal of Molecular Sciences 20, no. 9: 2240. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092240