Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases

Laboratoire des IMRCP, Université de Toulouse, CNRS UMR 5623, Université Toulouse III-Paul Sabatier, 118 Route de Narbonne, 31062 Toulouse CEDEX 09, France
Int. J. Mol. Sci. 2019, 20(10), 2407; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102407
Submission received: 26 April 2019
/
Revised: 9 May 2019
/
Accepted: 13 May 2019
/
Published: 15 May 2019
(This article belongs to the Special Issue Oxidative Stress in Neurodegenerative Diseases: Amyloid, Extracellular Vesicles and Biomarkers)
Abstract
:Increasing numbers of individuals suffer from neurodegenerative diseases, which are characterized by progressive loss of neurons. Oxidative stress, in particular, the overproduction of Reactive Oxygen Species (ROS), play an important role in the development of these diseases, as evidenced by the detection of products of lipid, protein and DNA oxidation in vivo. Even if they participate in cell signaling and metabolism regulation, ROS are also formidable weapons against most of the biological materials because of their intrinsic nature. By nature too, neurons are particularly sensitive to oxidation because of their high polyunsaturated fatty acid content, weak antioxidant defense and high oxygen consumption. Thus, the overproduction of ROS in neurons appears as particularly deleterious and the mechanisms involved in oxidative degradation of biomolecules are numerous and complexes. This review highlights the production and regulation of ROS, their chemical properties, both from kinetic and thermodynamic points of view, the links between them, and their implication in neurodegenerative diseases.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
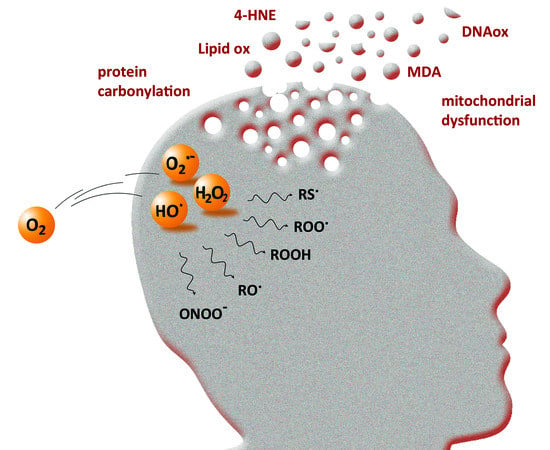
Reactive Oxygen Species (ROS) are radical or molecular species whose physical-chemical properties are well-known both on thermodynamic and kinetic points of view. They are produced from molecular oxygen, during the successive 4 steps of 1-electron reduction (reaction (1)). The reaction occurs in particular in the mitochondrial respiratory chain, where 85% of O2 is metabolized and where partially reduced O2 intermediates are produced in low quantity [1].
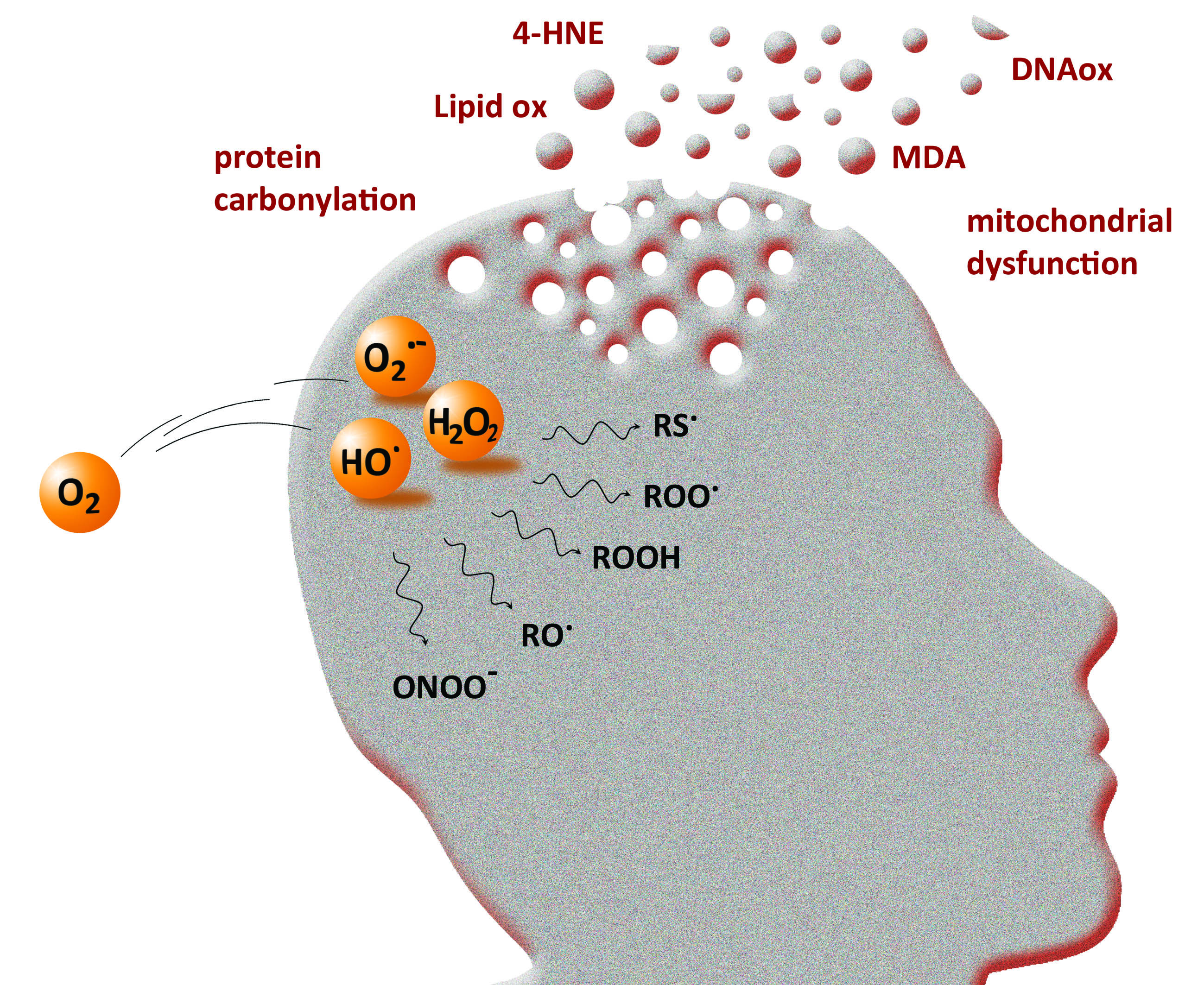
The three primary species, i.e., the superoxide anion (O2•‒), hydrogen peroxide (H2O2) and the hydroxyl radical (HO•), are called reactive oxygen species because they are oxygen-containing compounds with reactive properties. O2•‒ and HO• are commonly referred to as “free radicals”. They can react with organic substrates and lead to intermediate species able to further produce other ROS. For instance, H atom abstraction by HO• free radicals on a C-H bond leads to a carbon-centered radical, that further reacts rapidly with O2 to give a peroxyl radical RO2• (Figure 1) [2]. The latter may react with another substrate to give a new carbon-centered radical and a hydroperoxide ROOH, which may decompose into alkoxyl radical RO• in a reaction catalyzed by redox competent metal cations such as iron or copper (as occurring with heme proteins [3]). These “secondary” species are all ROS and share a similarity in structure and reactivity with the three primary species O2•‒, H2O2 and HO•. Among them, H2O2 (and hydroperoxides) is a molecular species and is supposed to be less reactive than the other radical short-lived species that are able to react with a range of targets (an exception may apply for O2•‒). However, its toxicity can be exerted via Fenton reaction in the presence of redox metal ions such as iron or copper (Figure 1), or via Haber–Weiss reaction in the presence of O2•‒ [4].
2. Production of ROS
2.1. Production of ROS In Vivo, Regulation and Oxidative Stress
ROS can be deleterious for biomolecules and lead to oxidative damages involved in several pathologies (neurodegenerative diseases, atherosclerosis, cancer and other disorders). However, they play, above all, an important role in homeostasis, cell signalization, regulation of metabolism, or memory formation via DNA methylation [5,6]. As recently reviewed, oxidative stress may be a key modulator in neurodegenerative diseases [7]. In mammalian cells, ROS are essentially produced by enzymes and are from different origins: mainly from the cytoplasmic membrane NADPH oxidase and from the enzyme complex of the mitochondrial respiratory chain, but also from sources of other organelles such as xanthine oxidase (XO), lipo- and cyclo-oxygenase, cytochromes P450 (endoplasmic reticulum) and peroxisomes. NADPH oxidase catalyzes the monoelectronic reduction of molecular oxygen, thus producing O2•‒ [8,9] that is released either outside the cell (for phagocytic cells) or inside the cell (for non-phagocytic cells) [10]. In mitochondria, ROS are produced during ATP biosynthesis which is accompanied by electron and proton transfers, with molecular oxygen as the final target. Electron leaks, which represent around 1–3% of the total electron production, may occur in complex I (NADH-ubiquinone oxidoreductase) and complex III (ubiquinol-cytochrome c oxidoreductase) of the electron transport chain and leads to the production of O2•‒ [11]. Because of the high activity of the mitochondrial respiratory chain in aerobic organisms, such a leak is the major source of ROS production in cells, more important than NAPDH oxidase (except during the activation of phagocytic cells) and XO [1]. The latter is a molybdenum enzyme, essentially located in the cytosol, that catalyzes the oxidation of hypoxanthine into xanthine and produces O2•‒, which might be further converted into H2O2 by XO (and oxidation of xanthine into uric acid) or by cytosolic Superoxide Dismutase (SOD) [12]. Xanthine oxidase is also able to convert nitrite into nitric oxide, and is thus a potential source of peroxynitrite [13]. Lipoxygenases and cyclooxygenases, which oxidize arachidonic acid into leukotrienes and prostanoids (including thromboxanes and prostaglandins), respectively, are other potential sources of ROS [14,15]. In the endoplasmic reticulum, enzymes belonging to the family of cytochromes P450 play a key role in the metabolism of drugs and other xenobiotics [16]. They reduce molecular oxygen to generate O2•‒ and H2O2, the latter being involved in the redox regulation of some essential functions of the endoplasmic reticulum [17].
In mitochondria, O2•‒ and H2O2 participate in redox signaling [18], but their production is significantly enhanced during oxidative stress conditions, as, for instance, in response to various diseases or stimuli. Oxidative stress reflects an imbalance between the production of ROS and the action of the antioxidant defense system in charge of their neutralization. They include enzymes, namely SOD that reduce O2•‒ into H2O2 [19,20], and catalase, glutathione peroxidases and thioredoxin reductase that regulate levels of H2O2 by converting it into H2O and O2 [21,22]. The selenoproteins glutathione peroxidases, among which the most abundant is the cytoplasmic and mitochondrial GPx1 [23], are also able to reduce hydroperoxides into alcohols. In addition to the enzymatic systems of defense, the regulation of the oxidative balance in vivo and the protection against oxidative attacks are also carried out by a myriad of non-enzymatic antioxidant systems, among which some are endogenous (glutathione, bilirubin, coenzyme Q, lipoic acid, melatonin, uric acid, etc.) and other ones are exogenous (α-tocopherol, ascorbic acid, carotenoids, etc.). Thus, under oxidative stress conditions, biomolecules may undergo the attack of ROS and get oxidized. Most of the time, such phenomena are deleterious for cells, but in some case, inducing an overproduction of ROS can help kill cells such as cancer cells [24].
2.2. Production of ROS In Vitro
Several commonly used methods are available for producing ROS in vitro, either based on metal-catalyzed production or not, and capable for some to selectively produce ROS. Water radiolysis is one of them and consists of irradiating water with γ-rays of 60Co or 137Cs (or X-ray). The initial energy deposition leads in situ to the generation of the primary radical and molecular species HO•, H•, eaq‒ (solvated electron), H2O2, H2 and H+, with well-known radiolytic yields of production [25]. The cumulated amount of ROS produced is directly linked to the radiation dose (expressed in Gy), which is dependent on the time the sample is exposed to the radiation source: the longer the exposure, the higher the radiation dose. Thus, it is easy to modulate the amount of ROS produced. A second advantage lies in the possibility of selecting ROS for a specific attack on a substrate: in aerated solutions ([O2] ≈ 2 × 10−4 mol L−1 in water), O2•‒, HO• and H2O2 are generated [26,27], whereas O2•‒ or HO• are selectively produced (along with H2O2) in 0.1 M sodium formate aqueous solution [28,29] or N2O-saturated water [30,31], respectively. For the diluted solution (below 10−2 mol·L−1), no direct interaction of radiation with the substrate occurs [25] and the latter is only oxidized by the ROS produced by water radiolysis. The production of ROS in vitro may also be achieved through the xanthine/xanthine oxidase system, an enzymatic way of selectively producing O2•‒ [32]. The selective production of HO• is usually obtained by the Fenton reaction where Fe2+ reduces H2O2 into HO• and HO‒ (Figure 1). In this case, ROS are generated by a metal-catalyzed reaction and the resulting oxidative damages are often site-directed, in particular when biomolecules are able to coordinate metal ions [33]. The same applies when ROS are produced by the Cu2+/ascorbate system, able to successively generate O2•‒, H2O2 and HO• [34,35,36]. For such systems, and unlike gamma radiolysis, the reaction continues as long as there are reagents, although it can be stopped in some cases [36,37]. The modulation of the production of ROS is more difficult to implement.
3. Chemical Properties and Reactivity of ROS
3.1. The Superoxide Anion
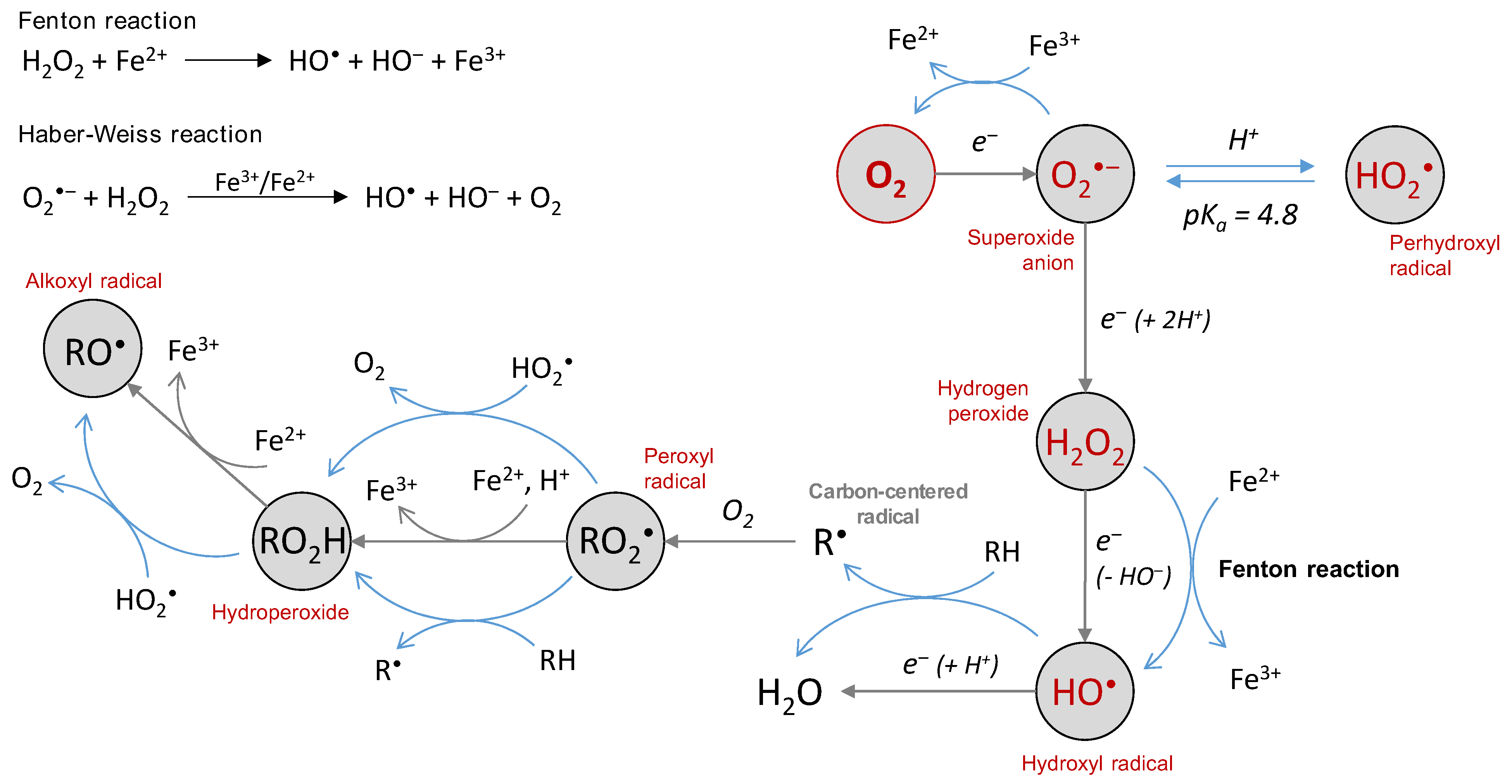
The superoxide anion is generated by the first 1-electron reduction of oxygen. At low pH, it is protonated and called perhydroxyl radical, with pKa(HO2•/O2•‒) = 4.8 [38] (Figure 1). There are two redox standard potentials for O2•‒, showing that it can act as a reductant (E°’(O2/O2•‒) = −0.33 V) or as an oxidant (E°’(O2•‒/H2O2) = 0.93 V) [39]. The 1-electron reduction of oxygen is not thermodynamically favored compared to its complete reduction (4 electrons, E°’(O2/H2O) = 0.81 V). Redox potentials also show that O2•‒ disproportionation and reduction of H2O2 by O2•‒ [40] (Haber–Weiss reaction, Figure 2) are thermodynamically spontaneous reactions.
Despite the relatively high values of its redox potential, O2•‒ is not a good reductant nor a good oxidant towards most of the biological substrates because of low rate constant values (usually below 102 L·mol−1·s−1) [38]. Some exception applies as O2•‒ is able to react with a few favored targets, with the rate constant ranging from 105 to 109 L·mol−1·s−1 [1]: cytochrome c, ascorbate and SOD (for which O2•‒ is the substrate). Recently, cytochrome c was used as a probe to demonstrate that O2•‒ was produced as an intermediate by the system Cu(I)-Aβ/O2 [41]. The perhydroxyl radical is more reactive (E°’(HO2•/H2O2) = 1.48 V) and able to oxidize polyunsaturated fatty acids such as linoleic, linolenic or arachidonic acids (k = 1.18 × 103, 1.70 × 103 and 3.05 × 103 L·mol−1·s−1) [42]. It is also engaged in the conversion of the peroxyl radical to hydroperoxide (Figure 2) and then to the alkoxyl radical [43]. The protonated form of O2•‒ could thus be the reactive one even if it is present at low concentrations at physiological pH. The toxicity of O2•‒ in a biological context is rather indirect since it is involved in the generation of highly-reactive secondary species. In the Haber–Weiss reaction (Figure 2), O2•‒ reacts with H2O2 to produce HO• radicals. The reaction is thermodynamically favored, but not kinetically [40,44,45], and needs to be catalyzed by iron. Disproportionation of perhydroxyl and of perhydroxyl/superoxide radicals (Figure 2) also represent a part of indirect toxicity of the superoxide anion as a potential source of H2O2. The rate constant is 6 × 105 L·mol−1·s−1 at pH 7, thus, the reaction is relevant under physiological conditions. The disproportionation of O2•‒, while thermodynamically spontaneous, is not kinetically favored [38]. Finally, the reaction of O2•‒ with •NO (k = 1.9 × 1010 L·mol−1·s−1) [46] to generate the highly-reactive peroxynitrite ONOO‒ is another reaction conferring an indirect toxicity to O2•‒, in particular towards DNA, proteins and lipids [47,48]. Peroxynitrite is able to nitrate tyrosine or tryptophan residues, or to oxidize methionine residues [49,50,51].
The reactivity of the superoxide anion does not always lead to a deleterious effect towards biomolecules as it is also able to help to fight against oxidative damages. Recently, Muñoz-Rugeles et al. [52] have shown that the superoxide anion is able to repair oxidized DNA by transferring one electron to the guanosyl radical of a single-stranded DNA. However, such an involvement in unusual chemical processes remains almost unexplored.
3.2. The Hydroxyl Radical
The hydroxyl radical is the most powerful oxidant among the ROS, with a potential of E°’(HO•/H2O) = 2.34 V [39]. At very low pH, HO• converts into its conjugate base O•‒ (pKa(HO•/O•‒) = 11.9), the oxide radical, which is less reactive [53] but not relevant at physiological pH. Reactions of HO• radicals with most substrates are diffusion-controlled (rate constants of 1010 L·mol−1·s−1) as, for example, with biological molecules such as DNA bases, aromatic amino acids, albumin, hemoglobin, linoleate or ascorbate [53,54,55]. Thus, HO• radicals are engaged in fast reactions, with an activation energy close to zero, meaning that they are not able to diffuse, have a very short lifetime (few 10−6 s) and free course (few 10−8 m), and are weakly selective towards molecular targets. A side consequence of this high reactivity is that the disproportionation of HO• radicals, even if kinetically favored (k ≈ 5 × 109 L·mol−1·s−1) [54], remains a rather infrequent event in biological conditions, the probability of collision between two hydroxyl radicals being very low.
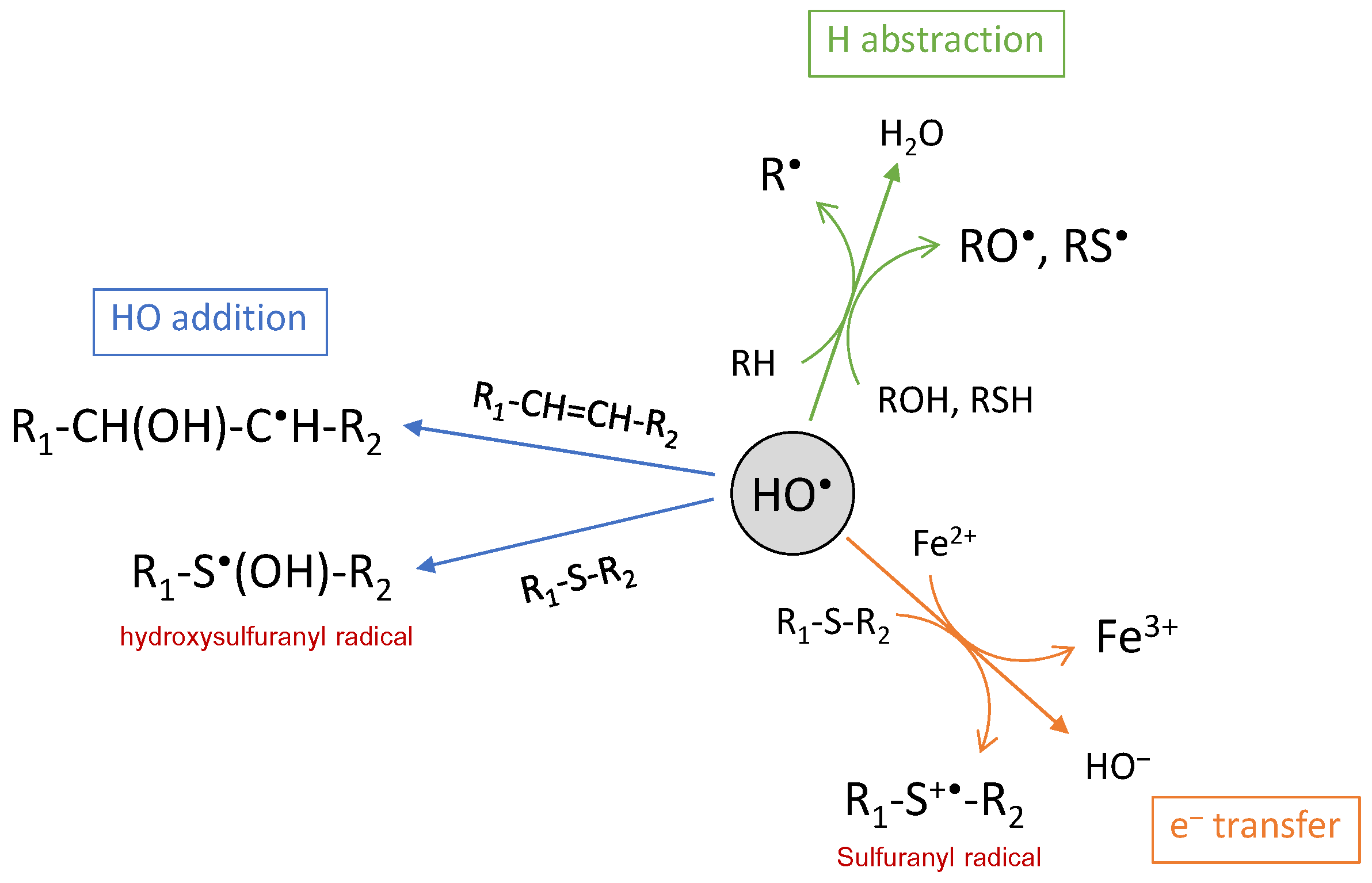
There are three ways of action for the HO• radical: electron abstraction, hydrogen abstraction and double bond addition (Figure 3). The HO• radical is electrophilic and has a strong affinity for electron-rich sites of molecules, in particular for aromatic or sulfur-containing molecules. This is illustrated by the rate constants of reaction with amino acids, ranging from 107 L·mol−1·s−1 for Gly to 1010 L·mol−1·s−1 for His, Trp or Cys [53]. Addition reactions are usually faster than H atom abstraction [56], except with Cys where H abstraction from the thiol group is faster [57,58]. HO• addition is commonly involved in biomolecule oxidation, such as oxidation of guanine into 8-oxoguanine [59], of histidine into 2-oxohistidine [60,61] or of tryptophan into N-formylkynurenine and kynurenine [43,62]. Hydroxyl radical addition may also occur on the sulfur atom, in particular, from methionine residue, leading to the hydroxysulfuranyl radical as the intermediate species and, finally, to methionine sulfoxide and sulfone as the end-products [63]. Methionine also undergoes electron abstraction when reacting with HO•, thus generating the sulfuranyl radical cation that is able to further evolve. In this case, oxidation leads to irreversible biological damages, contrary to the oxidation of methionine into methionine sulfoxide for which reversibility is ensured through methionine sulfoxide reductases (MsrA and MsrB) [64]. Electron abstraction is also observed with inorganic substrates such as ferrous ions or halides, with high rate constants [55]. The last pathway for HO• reaction is H-abstraction, for which numerous and various biomolecules are targets as, for instance, polyunsaturated fatty acids such as linoleate [65,66] or arachidonate [67,68], sulfur-containing, basic and aromatic amino acid residues from protein and peptides [69,70], or 2-deoxyribose and DNA bases [71]. Most of the time, H abstraction leads to a carbon-centered radical that either further reacts fast with molecular oxygen to generate a peroxyl radical or, in the absence of oxygen, is engaged in a biradical reaction generating a carbon-carbon bond [72]. However, abstraction may also occur on the hydroxyl or thiol functional groups, leading to oxygen- or sulfur-centered radicals [73]. Such mechanisms are observed for protein and peptide cross-linking via bityrosine formation [70,74] or disulfide bridge formation [75].
3.3. Hydrogen Peroxide
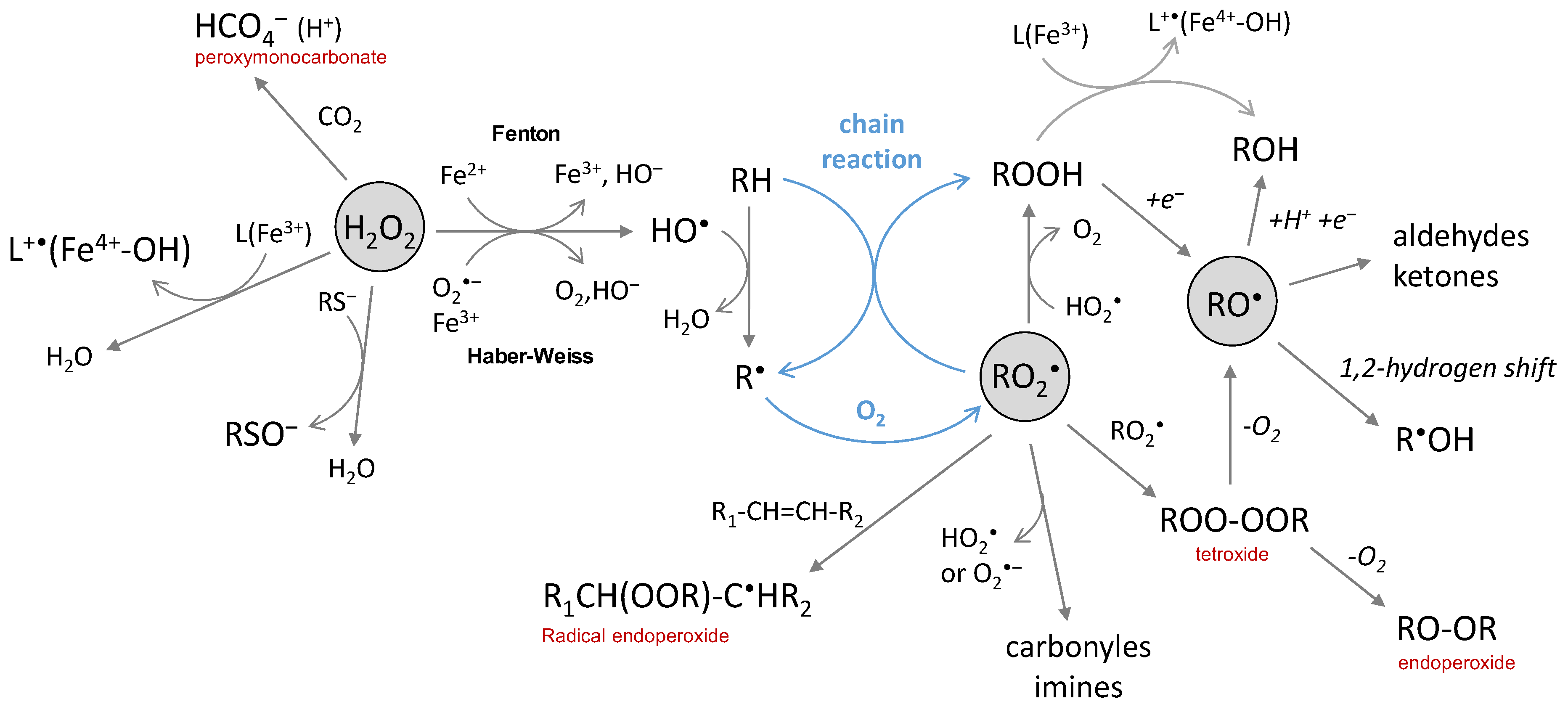
Hydrogen peroxide is produced by the two-electron reduction of molecular oxygen. Its conjugate base HOO‒ is a strong nucleophile but not relevant at physiological pH because of a high pKa value (pKa(H2O2/HOO‒) = 11.6). H2O2 is either a reductant or an oxidant in one-electron transfer reactions. The latter is not thermodynamically favored in biological conditions (E°’(O2•‒/H2O2) = 0.93 V and E°’(H2O2/HO•) = 0.30 V) [39] but H2O2 can act as an oxidant if catalysis by metal ions takes place (Fenton and Haber–Weiss reactions). It is rather engaged in two-electron transfer reactions, with a high potential (E°’(H2O2/H2O) = 1.32 V) in physiological conditions. It is more oxidizing than hypochlorous acid and peroxynitrite (E°’(ClO‒/Cl‒) = 1.28 V and E°’(ONOO‒/NO2‒) = 1.20 V). However, it reacts only poorly with most biological molecules because of a high activation energy barrier, oxidation by H2O2 being kinetically driven. Thus, the strongest oxidizing power of hydrogen peroxide comes indirectly from its metal-catalyzed conversion into HO• radicals by the Fenton and Haber–Weiss reactions (Figure 4).
In proteins, H2O2 reacts as a two-electron oxidant towards sulfur-containing residues (cysteine and methionine) but with a low rate constant (k = 2.9 L·mol−1·s−1 for cysteine) [76]. For thiols, the reaction is exclusive to the thiolate anion, thus the reactivity at physiological pH is dependent on pKa values. It leads to sulfenic acid (RSOH) as the initial product, able to be oxidized one more time by H2O2 into sulfinic acid (RSO2H) or to react with thiols to form disulfides. The highest rate constants are observed for the thiol proteins peroxiredoxins and glutathione peroxidases that react with H2O2 several orders of magnitude faster (~107 L·mol−1·s−1) [77]. Such a difference is explained by the polarization of the O-O bond of H2O2 by hydrogen bonding into the protein that facilitates the electrophilic attack on the thiolate. Pyruvate oxidation in acetate and carbon dioxide by H2O2 is also biologically relevant because of a rate constant of 2.2 L·mol−1·s−1 [78] and a pyruvate intracellular concentration of 0.1–0.5 mM (competitive with most thiols).
The toxicity of H2O2 can also be expressed indirectly. The reaction with bicarbonate leads to peroxymonocarbonate (HCO4‒) species that react approx. 300 times faster than H2O2 with thiols and sulfide (Figure 4) [79,80,81]. Only a few percent of H2O2 is present as peroxymonocarbonate in a physiological bicarbonate buffer since the reaction is an equilibrium (K = 0.32) [81]; it can be accelerated by carbonic anhydrase [79], thus enhancing the physiological relevance of the reaction. However, the most deleterious effect of H2O2 comes from its reaction with transition metals able to generate highly reactive radical species or activated metal complexes. The widely-known example is the production of hydroxyl radicals from hydrogen peroxide by the Fenton reaction (Figure 4), which involves iron ions as the metal catalyst. The reaction may be catalyzed by other redox competent metal ions (and, in this case, is called a “Fenton-like” reaction) as, for example, by copper complexed to the amyloid-beta peptide (Aβ) [82]. With the Fe3+ of heme proteins, the reaction of H2O2 is fast (k = 107–108 L·mol−1·s−1) [83] and give rise to Fe4+-oxoferryl porphyrin radical cation, able to transfer one electron to the surrounding protein [3,84], resulting in a formation of a protein radical that further evolves. The question of whether H2O2 is directly converted into HO• radicals or whether intermediates of higher oxidation states of the metal are produced has been debated in the past years. Both are possibly involved, depending on circumstances, but they are all strong oxidative species and the products resulting from their reaction with a substrate should be similar. When metal ions are coordinated to a biological molecule, the reaction may be different from Fenton chemistry since metal-catalyzed oxidation (MCO) is site-directed. Such a case is observed, for instance, for protein, DNA or the Aβ peptide in iron- or copper-catalyzed oxidation in the presence of ascorbate [36,85,86].
3.4. Peroxyl Radicals, Hydroperoxides and Alkoxyl Radicals
Peroxyl radicals are secondary species generated by the addition of molecular oxygen on carbon-centered radicals ([O2] ≈ 2 × 10−4 mol·L−1 in aerated aqueous solution), whose rate constant usually range between 108 and 109 L·mol−1·s−1 [2]. They can also be produced in the absence of oxygen by metal-induced conversion of hydroperoxides [87]. Hydroperoxides are generated from peroxyl radicals by reaction with HO2• or by H abstraction from another molecule, and may further react with HO2• or a metal ion to generate alkoxyl radicals (Figure 4). The latter can also be generated from peroxyl radicals via a tetroxide. Peroxyl and alkoxyl radicals are oxidant species, with relatively high redox standard potentials of E°’(RO2•/RO2H) = 1.00 V and E°’(RO•/ROH) = 1.60 V, respectively [88].
Peroxyl radicals react faster than the superoxide anion with numerous biological substrates (DNA, lipids, proteins); rate constants [2] ranging from 102 to 108 L·mol−1·s−1. Even if they are much less reactive than the hydroxyl radical, they share some similarity in their mode of reaction as they are able to either be engaged in electron abstraction, H atom abstraction or addition on double-bonds (Figure 4) [89]. In the latter case, intra- or intermolecular reactions lead to the formation of the radical endoperoxide ROOR• species. Peroxyl radicals with the α-hydroxyl or α-amino groups can also undergo rapid unimolecular elimination of HO2•/O2•‒, leading to carbonyl or imine group formation [2,90,91]. Peroxyl radicals ROO• can undergo dimerization with other peroxyl radicals R’OO• and yield tetroxide species ROO-OOR’; the reaction is also possible between peroxyl radicals and HO2•, as observed for thymine [59]. Tetroxide is an unstable species and their subsequent decomposition yields carbonyl groups and alcohol, accompanied by the loss of molecular oxygen [3].
Among the very diverse reactions that peroxyl radicals can initiate, some are of particular importance because they contribute to the degradation of cell membranes induced by lipid peroxidation. Once a carbon-centered radical has been generated on a fatty acid moiety, it reacts fast with molecular oxygen to yield a peroxyl radical, able to abstract an H atom from another fatty acid moiety to give birth to another carbon-centered radical. This H abstraction is facilitated by the proximity of the two fatty acid chains within the lipid bilayers of cell membranes. In such a condition, a chain reaction starts and is propagated by the R• and RO2• radicals. The chain reaction stops either when there are no more lipids, no more oxygen or when peroxide radicals react with a lipid-soluble antioxidant, such as α-tocopherol or carotenoids [92,93].
Primary and secondary alkoxyl radicals undergo a rapid 1,2-hydrogen shift, resulting in the generation of α-hydroxyalkyl radicals, in competition with the intramolecular 1,5-hydrogen shift and the formation of alcohol by intermolecular H abstraction [94,95]. In some cases, in particular, when a 1,2-hydrogen shift is not possible (tertiary alkoxyl radicals), β-fragmentation reactions occur and yield aldehydes and ketones [96] with relatively high rate constants in aqueous solutions (k > 106 s−1) [97,98].
4. The Implication of ROS in Neurodegenerative Diseases
The high consumption of molecular oxygen and the high content of polyunsaturated fatty acid, strongly sensitive to peroxidation, make the brain a particularly vulnerable tissue to oxidative stress [99]. The latter is a modulator of neurodegenerative diseases (recently reviewed in Reference [7]). Peroxidation products of fatty acids are among the biomarkers of oxidative stress in neurodegenerative diseases such as Alzheimer’s disease (AD), Parkinson’s disease (PD) and amyotrophic lateral sclerosis (ALS), along with protein carbonylation and nitration, DNA and RNA oxidative damages [100,101,102,103,104]. Neurodegenerative diseases are commonly associated with abnormal protein aggregation. In AD, the Aβ peptide is found aggregated in senile plaques (composed of Aβ fibrils and metal ions) and hyperphosphorylated Tau in neurofibrillary tangles. In PD, aggregation of α-synuclein leads to Lewy bodies inclusions, while the aggregation of the huntingtin protein and copper/zinc superoxide dismutase are involved in Huntington’s disease (HD) and ALS, respectively. Such abnormal protein aggregation is able to induce oxidative stress via mitochondria dysfunction and ROS production [105,106,107], leading to chronic inflammation, and play an important role in neurodegeneration.
In AD, an imbalance between the production of ROS and the reduced activity of enzymes responsible for ROS scavenging leads to oxidative damages on biomolecules, and on the Aβ peptide itself [108,109,110]. The link between oxidative stress and the amyloid beta peptide has been recently reviewed [111]. Because copper is present in relatively high levels in the brain and because of the ability of the Aβ peptide to chelate metal ions, Aβ-copper is a potential direct source of ROS in the presence of ascorbate and molecular oxygen. Reybier et al. [41] have shown that the superoxide anion is generated as an intermediate during H2O2 production by Aβ-copper. No direct link has yet been established between the production of ROS by Aβ-copper and oxidation of biological material in vivo. However, increased levels of lipids, protein and DNA oxidation have been reported to be associated with elevated levels of Aβ, whereas low Aβ-content brain regions do not present high concentrations of oxidative stress markers [112,113,114,115]. Lipid peroxidation is one of the events associated with AD, which might be involved in the phospholipid imbalance observed in the brain of AD patients [116,117]. Malondialdehyde (MDA) and 4-hydroxynonenal (4-HNE) are two aldehydes commonly found in high levels in AD brains [118]. The compound 4-HNE is toxic for neurons by causing apoptosis or by altering the microtubule structure [119,120], but is also prone to react with lipoid acid [121] and to form adducts with proteins (target amino acids are cysteine, histidine and lysine) [122] detected in AD brains [123]. In particular, adducts with Tau were found to modify its conformation and to favor neurofibrillary tangles formation [124]. The compound 4-HNE is generated by the non-enzymatic oxidation of polyunsaturated omega-6 fatty acids, such as arachidonate or linoleate. It is a direct consequence of the peroxidation of lipids by ROS since it is generated by the degradation of lipid hydroperoxides [125]—hydroperoxyoctadecadioenoate (HPODE) from linoleate or hydroperoxyeicosatetraenoate (HPETE) from arachidonate (Figure 5). F2- and F4-isoprostanes, which are generated by peroxidation of arachidonate, are other markers of oxidative stress in AD and found in elevated levels in the brain of AD patients [126,127,128].
Protein oxidation has been evidenced by high levels of carbonylated proteins in the brain areas the most involved in AD (i.e., hippocampus and parietal cortex) [114,129]. Several molecular mechanisms have been proposed for protein carbonylation, some of them being induced by direct ROS attacks and leading to protein cleavage via an alkoxyde radical formation (Figure 5). The target proteins are, among others, those involved in glucose metabolism and ATP synthesis [130], such as ATP synthase [131], pyruvate kinase, phosphoglucose mutase, α-enolase, malate dehydrogenase or glyceraldehyde-3-phosphate dehydrogenase (see Reference [132] for a review). Modifications detected include carbonylation, nitration and HNE-adducts formation. Like protein carbonylation, oxidative damages of DNA bases may result from a direct attack of ROS. Increased levels of 8-oxo-2-dehydroguanine, 8-hydroxyadenine and 5-hydroxyuracil have been reported in the temporal, parietal and frontal lobes of AD brains [133,134], along with 8-hydroxyguanine in the hippocampus of patients with preclinical stages of AD [135]. The high levels of oxidized DNA bases are detected in neurons where lipids and protein oxidation are also increased [136].
In PD, the involvement of ROS and oxidative stress might be one of the major factors causing the disease. Dopaminergic neurons of the substantia nigra, where the basal level of free radicals is important [137], are particularly sensitive to degeneration. Elevated levels of oxidized lipids and proteins have been detected in the substantia nigra of PD patients [138,139]. Additionally, an increase of 8-hydroxy-2′-deoxyguanosine and 8-hydroxyguanine levels, two markers of DNA oxidation, was observed [140]. As in AD, 4-HNE-modified proteins have also been detected in PD [141]. Thus, AD and PD share some similarities regarding the biomarkers of the oxidative stress detected. In PD, metal ion release (e.g., Fe2+) would be an important mechanism of neurodegeneration, through ROS production and dopamine oxidation. High levels of iron ions, in conjunction with the production of H2O2 via dopamine oxidation (enzymatically by monoamine oxidases, would lead to an overproduction of ROS and thus to oxidative stress conditions. Non-enzymatic oxidation of dopamine is involved in free radicals production and in elevation of free iron levels in dopaminergic cells [142,143,144]. Oxidative modification of proteins in PD may also have an impact on their propensity to aggregate. Surgucheva et al. [145] have shown that oxidation of γ-synuclein enhanced the formation of annular oligomers that accumulate in cells and that can initiate α-synuclein aggregation.
In other diseases such as ALS or HD, the link with oxidative stress is also evidenced, even if the mechanisms involved in their etiology are not fully understood. In ALS, oxidative stress is evidenced by elevated levels of MDA, 4-HNE, advanced oxidation protein products, isoprostanoids and 8-hydroxy-2′-deoxyguanosine [146,147,148,149,150]. Oxidative stress is also coupled to mitochondrial damages and dysfunction, each exacerbating the other, and to RNA dysmetabolism and unfolded protein aggregates formation [151,152]. As in other neurodegenerative diseases, copper and iron homeostasis is disturbed in ALS and elevated levels of these redox-competent metal ions could participate in ROS production [153]. Strong evidence exists also for early oxidative stress in HD, coupled with mitochondrial dysfunction, but it is still not clear whether oxidative stress is a cause or a consequence of HD. As for ALS, metal dyshomeostasis, evidenced by high levels of iron and copper in post-mortem brain tissues of HD patients [154], would participate in ROS production via Fenton chemistry. Increases of nuclear and mitochondrial DNA 8-hydroxy-2′-deoxyguanosine were detected in the blood and serum of HD patients [155,156], along with DNA double-strand breaks, a potential result of free radical damage [157,158]. Lipid peroxidation (high levels of MDA and cytoplasmic lipofuscin), protein carbonylation and nitration (an increase of 3-nitrotyrosine levels) are also observed in HD [159,160].
5. Concluding Remarks
An important feature shared by most of the neurodegenerative diseases is the presence of oxidative damages that link them to oxidative stress. The latter is supposed to be an early event in the etiology of some diseases since the biomarkers of oxidation appear early in their development [132]. An overproduction of ROS is considered to have a major contribution in oxidative damages undergone by biomolecules, including lipids, proteins and DNA. The intrinsic chemical properties of ROS make them formidable weapons against most biomolecules. Among them, because of its diffusion-controlled reactivity with most of the biological material, HO• may be considered as a nuclear weapon compared to O2•‒ and H2O2. These last two react directly only with few specific targets (e.g., SOD and catalase). However, they are strong deleterious species because of (i) their ability to be engaged in Fenton and Haber-Weiss reactions, two metal-catalyzed reactions that lead to HO• production, (ii) their lack of direct reactivity which gives them the possibility to spread to areas where metal levels are high. So, most of the time, final oxidative damages on biomolecules could be considered as resulting from HO• attacks. This could particularly apply to neurodegenerative diseases where metal dyshomeostasis takes place and where elevated levels of redox-competent metal ions—such as iron or copper—are observed. In this context, better control of ROS homeostasis would be important for neuron survival. This could be achieved, among others, by developing antioxidant-based strategies. This is the reason why many studies have focused and are still focusing on possible therapeutic approaches based on antioxidant strategies, either by the administration of antioxidant in the form of plant extracts or nutraceuticals [161,162] or by reinforcing the antioxidant defense system in vivo [163]. Antioxidant therapy-based strategies to fight against neurodegenerative diseases have shown promising results in preclinical trials but only a few clinical trials have been conducted and the benefit of such a therapy is still under debate [164]. In this context, and because most of the mechanisms underlying the etiology of neurodegenerative diseases have still not been elucidated, all efforts to better understand, through basic research, the causes of disease development will increase the global knowledge and will help to develop novel therapeutic strategies.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 3rd ed.; Oxford science Publications: Oxford, UK, 1999. [Google Scholar]
- Neta, P.; Huie, R.E.; Ross, A.B. Rate constants for reactions of peroxyl radicals in fluid solutions. J. Phys. Chem. Ref. Data 1990, 19, 413–513. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Generation and propagation of radical reactions on proteins. Biochim. Et Biophys. Acta 2001, 1504, 196–219. [Google Scholar] [CrossRef] [Green Version]
- Haber, F.; Weiss, J. The catalytic decomposition of hydrogen peroxide by iron salts. Proc. R. Soc. Lond. 1934, 147, 332–351. [Google Scholar]
- Rhee, S.G. Redox signaling: Hydrogen peroxide as intracellular messenger. Exp. Mol. Med. 1999, 31, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhuang, Z.; Wang, W.; He, L.; Wu, H.; Cao, Y.; Pan, F.; Zhao, J.; Hu, Z.; Sekhar, C.; et al. OGG1 is essential in oxidative stress induced DNA demethylation. Cell Signal. 2016, 28, 1163–1171. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Vignais, P.V. The superoxide-generating NADPH oxidase: Structural aspects and activation mechanism. Cell. Mol. Life Sci. 2002, 59, 1428–1459. [Google Scholar] [CrossRef]
- Finkel, T. Redox-dependent signal transduction. FEBS Lett. 2000, 476, 52–54. [Google Scholar] [CrossRef] [Green Version]
- Souza, H.P.; Laurindo, F.R.; Ziegelstein, R.C.; Berlowitz, C.O.; Zweier, J.L. Vascular NAD(P)H oxidase is distinct from the phagocytic enzyme and modulates vascular reactivity control. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H658–H667. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Schmidt, H.M.; Kelley, E.E.; Straub, A.C. The impact of xanthine oxidase (XO) on hemolytic diseases. Redox Biol. 2019, 21, 101072. [Google Scholar] [CrossRef]
- Godber, B.L.; Doel, J.J.; Sapkota, G.P.; Blake, D.R.; Stevens, C.R.; Eisenthal, R.; Harrison, R. Reduction of nitrite to nitric oxide catalyzed by xanthine oxidoreductase. J. Biol. Chem. 2000, 275, 7757–7763. [Google Scholar] [CrossRef]
- Bonizzi, G.; Piette, J.; Merville, M.P.; Bours, V. Cell type-specific role for reactive oxygen species in nuclear factor-kappaB activation by interleukin-1. Biochem. Pharmacol. 2000, 59, 7–11. [Google Scholar] [CrossRef]
- van der Donk, W.A.; Tsai, A.L.; Kulmacz, R.J. The cyclooxygenase reaction mechanism. Biochemistry 2002, 41, 15451–15458. [Google Scholar] [CrossRef]
- Estabrook, R.W. A passion for P450s (rememberances of the early history of research on cytochrome P450). Drug Metab. Dispos. Biol. Fate Chem. 2003, 31, 1461–1473. [Google Scholar] [CrossRef]
- Coon, M.J.; Ding, X.X.; Pernecky, S.J.; Vaz, A.D. Cytochrome P450: Progress and predictions. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1992, 6, 669–673. [Google Scholar] [CrossRef]
- Brand, M.D. Mitochondrial generation of superoxide and hydrogen peroxide as the source of mitochondrial redox signaling. Free Radic. Biol. Med. 2016, 100, 14–31. [Google Scholar] [CrossRef]
- Hsu, J.L.; Hsieh, Y.; Tu, C.; O’Connor, D.; Nick, H.S.; Silverman, D.N. Catalytic properties of human manganese superoxide dismutase. J. Biol. Chem. 1996, 271, 17687–17691. [Google Scholar] [CrossRef]
- Oury, T.D.; Crapo, J.D.; Valnickova, Z.; Enghild, J.J. Human extracellular superoxide dismutase is a tetramer composed of two disulphide-linked dimers: A simplified, high-yield purification of extracellular superoxide dismutase. Biochem. J. 1996, 317 (Pt 1), 51–57. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Stanley, B.A.; Sivakumaran, V.; Shi, S.; McDonald, I.; Lloyd, D.; Watson, W.H.; Aon, M.A.; Paolocci, N. Thioredoxin reductase-2 is essential for keeping low levels of H(2)O(2) emission from isolated heart mitochondria. J. Biol. Chem. 2011, 286, 33669–33677. [Google Scholar] [CrossRef] [PubMed]
- Ursini, F.; Maiorino, M.; Brigelius-Flohe, R.; Aumann, K.D.; Roveri, A.; Schomburg, D.; Flohe, L. Diversity of glutathione peroxidases. Methods Enzym. 1995, 252, 38–53. [Google Scholar]
- Lee, C.M.; Lee, J.; Nam, M.J.; Choi, Y.S.; Park, S.H. Tomentosin Displays Anti-Carcinogenic Effect in Human Osteosarcoma MG-63 Cells via the Induction of Intracellular Reactive Oxygen Species. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Spinks, J.W.T.; Woods, R.J. Water and inorganic aqueous systems. In Introduction to Radiation Chemistry; John Wiley & Sons: New York, NY, USA, 1990; pp. 243–313. [Google Scholar]
- Bielski, B.H.J. Re-Evaluation of Spectral and Kinetic-Properties of Ho2 and (0--)2 Free-Radicals. Photochem. Photobiol. 1978, 28, 645–649. [Google Scholar] [CrossRef]
- Thomas, J.K. The rate constants for H atom reactions in aqueous solutions 1. J. Phys. Chem. 1963, 67, 2593–2595. [Google Scholar] [CrossRef]
- Matthews, R.W.; Sangster, D.F. Measurement by Benzoate Radiolytic Decarboxylation of Relative Rate Constants for Hydroxyl Radical Reactions. J. Phys. Chem. 1965, 69, 1938–1946. [Google Scholar] [CrossRef]
- Rabani, J.; Matheson, M.S. The Pulse Radiolysis of Aqueous Solutions of Potassium Ferrocyanide1. J. Phys. Chem. 1966, 70, 761–769. [Google Scholar] [CrossRef]
- Janata, E.; Schuler, R.H. Rate constant for scavenging eaq- in nitrous oxide-saturated solutions. J. Phys. Chem. 1982, 86, 2078–2084. [Google Scholar] [CrossRef]
- Asmus, K.D.; Fendler, J.H. Reaction of sulfur hexafluoride with hydrated electrons. J. Phys. Chem. 1968, 72, 4285–4289. [Google Scholar] [CrossRef]
- Hille, R.; Hall, J.; Basu, P. The mononuclear molybdenum enzymes. Chem. Rev. 2014, 114, 3963–4038. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, C.; Xu, H.H.; Liu, Y.N.; Zhou, F. Binding of alpha-synuclein with Fe(III) and with Fe(II) and biological implications of the resultant complexes. J. Inorg. Biochem. 2010, 104, 365–370. [Google Scholar] [CrossRef]
- Miotto, M.C.; Rodriguez, E.E.; Valiente-Gabioud, A.A.; Torres-Monserrat, V.; Binolfi, A.; Quintanar, L.; Zweckstetter, M.; Griesinger, C.; Fernandez, C.O. Site-specific copper-catalyzed oxidation of alpha-synuclein: Tightening the link between metal binding and protein oxidative damage in Parkinson’s disease. Inorg. Chem. 2014, 53, 4350–4358. [Google Scholar] [CrossRef] [PubMed]
- Cheignon, C.; Faller, P.; Testemale, D.; Hureau, C.; Collin, F. Metal-catalyzed oxidation of Abeta and the resulting reorganization of Cu binding sites promote ROS production. Metallomics 2016, 8, 1081–1089. [Google Scholar] [CrossRef]
- Cassagnes, L.E.; Herve, V.; Nepveu, F.; Hureau, C.; Faller, P.; Collin, F. The catalytically active copper-amyloid-Beta state: Coordination site responsible for reactive oxygen species production. Angew. Chem. Int. Ed. Engl. 2013, 52, 11110–11113. [Google Scholar] [CrossRef]
- Cheignon, C.; Hureau, C.; Collin, F. Real-time evolution of A beta(40) metal-catalyzed oxidation reveals Asp1 as the main target and a dependence on metal binding site. Inorg. Chim. Acta 2018, 472, 111–118. [Google Scholar] [CrossRef]
- Bielski, B.H.J.; Cabelli, D.E.; Arudi, R.L.; Ross, A.B. Reactivity of HO2/O2− Radicals in Aqueous Solution. J. Phys. Chem. Ref. Data 1985, 14, 1041–1100. [Google Scholar] [CrossRef]
- Wardman, P. Reduction Potentials of One-Electron Couples Involving Free-Radicals in Aqueous-Solution. J. Phys. Chem. Ref. Data 1989, 18, 1637–1755. [Google Scholar] [CrossRef]
- Ferradini, C.; Foos, J.; Houee, C.; Pucheault, J. Reaction between Superoxide Anion and Hydrogen-Peroxide. Photochem. Photobiol. 1978, 28, 697–700. [Google Scholar] [CrossRef]
- Reybier, K.; Ayala, S.; Alies, B.; Rodrigues, J.V.; Bustos Rodriguez, S.; La Penna, G.; Collin, F.; Gomes, C.M.; Hureau, C.; Faller, P. Free Superoxide is an Intermediate in the Production of H2O2 by Copper(I)-Abeta Peptide and O2. Angew. Chem. Int. Ed. 2016, 55, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Bielski, B.H.J.; Arudi, R.L.; Sutherland, M.W. A Study of the Reactivity of Ho2/O2- with Unsaturated Fatty-Acids. J. Biol. Chem. 1983, 258, 4759–4761. [Google Scholar] [PubMed]
- Stadtman, E.R.; Levine, R.L. Free radical-mediated oxidation of free amino acids and amino acid residues in proteins. Amino Acids 2003, 25, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Rigo, A.; Stevanato, R.; Finazzi-Agro, A.; Rotilio, G. An attempt to evaluate the rate of the Haber-Weiss reaction by using OH radical scavengers. FEBS Lett. 1977, 80, 130–132. [Google Scholar] [CrossRef]
- Weinstein, J.; Bielski, B.H.J. Kinetics of the Interaction of Ho2 and O2-Radicals with Hydrogen-Peroxide - Haber-Weiss Reaction. J. Am. Chem. Soc. 1979, 101, 58–62. [Google Scholar] [CrossRef]
- Kissner, R.; Nauser, T.; Bugnon, P.; Lye, P.G.; Koppenol, W.H. Formation and properties of peroxynitrite as studied by laser flash photolysis, high-pressure stopped-flow technique, and pulse radiolysis. Chem. Res. Toxicol. 1997, 10, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, G.L.; Pryor, W.A. The formation of peroxynitrite in vivo from nitric oxide and superoxide. Chem. Biol. Interact. 1995, 96, 203–206. [Google Scholar] [CrossRef]
- Packer, M.A.; Porteous, C.M.; Murphy, M.P. Superoxide production by mitochondria in the presence of nitric oxide forms peroxynitrite. Biochem. Mol. Biol. Int. 1996, 40, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Uchida, K.; Kawakishi, S. Oxidative fragmentation of collagen and prolyl peptide by Cu(II)/H2O2. Conversion of proline residue to 2-pyrrolidone. J. Biol. Chem. 1992, 267, 23646–23651. [Google Scholar] [PubMed]
- Pryor, W.A.; Squadrito, G.L. The chemistry of peroxynitrite: A product from the reaction of nitric oxide with superoxide. Am. J. Physiol. 1995, 268, L699–L722. [Google Scholar] [CrossRef] [PubMed]
- Berlett, B.S.; Friguet, B.; Yim, M.B.; Chock, P.B.; Stadtman, E.R. Peroxynitrite-mediated nitration of tyrosine residues in Escherichia coli glutamine synthetase mimics adenylylation: Relevance to signal transduction. Proc. Natl. Acad. Sci. USA 1996, 93, 1776–1780. [Google Scholar] [CrossRef]
- Munoz-Rugeles, L.; Galano, A.; Alvarez-Idaboy, J.R. The other side of the superoxide radical anion: Its ability to chemically repair DNA oxidized sites. Chem. Commun. 2018, 54, 13710–13713. [Google Scholar] [CrossRef]
- Buxton, G.V.; Greenstock, C.L.; Helman, W.P.; Ross, A.B. Critical-Review of Rate Constants for Reactions of Hydrated Electrons, Hydrogen-Atoms and Hydroxyl Radicals (.OH/.O-) in Aqueous-Solution. J. Phys. Chem. Ref. Data 1988, 17, 513–886. [Google Scholar] [CrossRef]
- Farhataziz, R.A.; Ross, A. Selected specific rates of reactions of transients from water in aqueous solution. III. Hydroxyl Radic. Perhydroxyl Radic. Radic. Ions Nsrds-Nbs 1977, 59. [Google Scholar]
- Dorfman, L.M.; Adams, G.E. Reactivity of the hydroxyl radical in aqueous solutions; DTIC Document: 1973.
- Von Sonntag, C. The Chemical Basis of Radiation Biology; Taylor and Francis: Abingdon, UK, 1987; p. 515. [Google Scholar]
- Von Sonntag, C. Free-radical reactions involving thiols and disulphides. In Sulfur-Centered Reactive Intermediates in Chemistry and Biology; Chatgilialoglu, C., Asmus, K.-D., Eds.; Plenum Press: New York, NY, USA, 1990; pp. 359–366. [Google Scholar]
- Armstrong, D.A. Application of pulse radiolysis for the study of short-lived sulphur species. In Sulfur-Centered Reactive Intermediates in Chemistry and Biology; Chatgilialoglu, K.-D.A., Ed.; Plenum Press: New York, NY, USA, 1990; pp. 121–134. [Google Scholar]
- Cadet, J.; Delatour, T.; Douki, T.; Gasparutto, D.; Pouget, J.P.; Ravanat, J.L.; Sauvaigo, S. Hydroxyl radicals and DNA base damage. Mutat. Res. 1999, 424, 9–21. [Google Scholar] [CrossRef]
- Schoneich, C. Mechanisms of metal-catalyzed oxidation of histidine to 2-oxo-histidine in peptides and proteins. J. Pharm. Biomed. Anal. 2000, 21, 1093–1097. [Google Scholar] [CrossRef]
- Uchida, K.; Kawakishi, S. 2-Oxo-histidine as a novel biological marker for oxidatively modified proteins. FEBS Lett. 1993, 332, 208–210. [Google Scholar] [CrossRef] [Green Version]
- Berlett, B.S.; Levine, R.L.; Stadtman, E.R. Comparison of the effects of ozone on the modification of amino acid residues in glutamine synthetase and bovine serum albumin. J. Biol. Chem. 1996, 271, 4177–4182. [Google Scholar] [CrossRef]
- Schoneich, C.; Pogocki, D.; Hug, G.L.; Bobrowski, K. Free radical reactions of methionine in peptides: Mechanisms relevant to beta-amyloid oxidation and Alzheimer’s disease. J. Am. Chem. Soc. 2003, 125, 13700–13713. [Google Scholar] [CrossRef]
- Hoshi, T.; Heinemann, S. Regulation of cell function by methionine oxidation and reduction. J. Physiol. 2001, 531, 1–11. [Google Scholar] [CrossRef]
- Hasegawa, K.; Patterson, L.K. Pulse-Radiolysis Studies in Model Lipid Systems: Formation and Behavior of Peroxy Radicals in Fatty-Acids. Photochem. Photobiol. 1978, 28, 817–823. [Google Scholar] [CrossRef]
- Gardner, H.W. Oxygen radical chemistry of polyunsaturated fatty acids. Free Radic. Biol. Med. 1989, 7, 65–86. [Google Scholar] [CrossRef]
- Yin, H.; Havrilla, C.M.; Gao, L.; Morrow, J.D.; Porter, N.A. Mechanisms for the formation of isoprostane endoperoxides from arachidonic acid. “Dioxetane” intermediate versus beta-fragmentation of peroxyl radicals. J. Biol. Chem. 2003, 278, 16720–16725. [Google Scholar] [CrossRef]
- Greco, A.; Minghetti, L.; Levi, G. Isoprostanes, novel markers of oxidative injury, help understanding the pathogenesis of neurodegenerative diseases. Neurochem. Res. 2000, 25, 1357–1364. [Google Scholar] [CrossRef]
- Stadtman, E.R. Protein oxidation in aging and age-related diseases. Ann. N. Y. Acad. Sci. 2001, 928, 22–38. [Google Scholar] [CrossRef]
- Davies, K.J.; Delsignore, M.E.; Lin, S.W. Protein damage and degradation by oxygen radicals. II. Modification of amino acids. J. Biol. Chem. 1987, 262, 9902–9907. [Google Scholar] [PubMed]
- Breen, A.P.; Murphy, J.A. Reactions of oxyl radicals with DNA. Free Radic. Biol. Med. 1995, 18, 1033–1077. [Google Scholar] [CrossRef]
- Stadtman, E.R. Oxidation of free amino acids and amino acid residues in proteins by radiolysis and by metal-catalyzed reactions. Annu. Rev. Biochem. 1993, 62, 797–821. [Google Scholar] [CrossRef] [PubMed]
- Lal, M. Radiation-Induced Oxidation of Sulfhydryl Molecules in Aqueous-Solutions - a Comprehensive Review. Radiat. Phys. Chem. 1994, 43, 595–611. [Google Scholar] [CrossRef]
- Atwood, C.S.; Perry, G.; Zeng, H.; Kato, Y.; Jones, W.D.; Ling, K.Q.; Huang, X.; Moir, R.D.; Wang, D.; Sayre, L.M.; et al. Copper mediates dityrosine cross-linking of Alzheimer’s amyloid-beta. Biochemistry 2004, 43, 560–568. [Google Scholar] [CrossRef]
- Garrison, W.M. Reaction-Mechanisms in the Radiolysis of Peptides, Polypeptides, and Proteins. Chem. Rev. 1987, 87, 381–398. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Metodiewa, D. Reactivity of biologically important thiol compounds with superoxide and hydrogen peroxide. Free Radic. Biol. Med. 1999, 27, 322–328. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Hampton, M.B. Thiol chemistry and specificity in redox signaling. Free Radic. Biol. Med. 2008, 45, 549–561. [Google Scholar] [CrossRef]
- Vasquez-Vivar, J.; Denicola, A.; Radi, R.; Augusto, O. Peroxynitrite-mediated decarboxylation of pyruvate to both carbon dioxide and carbon dioxide radical anion. Chem. Res. Toxicol. 1997, 10, 786–794. [Google Scholar] [CrossRef]
- Bakhmutova-Albert, E.V.; Yao, H.; Denevan, D.E.; Richardson, D.E. Kinetics and mechanism of peroxymonocarbonate formation. Inorg. Chem. 2010, 49, 11287–11296. [Google Scholar] [CrossRef] [PubMed]
- Trindade, D.F.; Cerchiaro, G.; Augusto, O. A role for peroxymonocarbonate in the stimulation of biothiol peroxidation by the bicarbonate/carbon dioxide pair. Chem. Res. Toxicol. 2006, 19, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.E.; Yao, H.R.; Frank, K.M.; Bennett, D.A. Equilibria, kinetics, and mechanism in the bicarbonate activation of hydrogen peroxide: Oxidation of sulfides by peroxymonocarbonate. J. Am. Chem. Soc. 2000, 122, 1729–1739. [Google Scholar] [CrossRef]
- Kowalik-Jankowska, T.; Ruta, M.; Wisniewska, K.; Lankiewicz, L.; Dyba, M. Products of Cu(II)-catalyzed oxidation in the presence of hydrogen peroxide of the 1-10, 1-16 fragments of human and mouse beta-amyloid peptide. J. Inorg. Biochem. 2004, 98, 940–950. [Google Scholar] [CrossRef]
- Davies, M.J.; Hawkins, C.L.; Pattison, D.I.; Rees, M.D. Mammalian heme peroxidases: From molecular mechanisms to health implications. Antioxid. Redox Signal. 2008, 10, 1199–1234. [Google Scholar] [CrossRef]
- Davies, M.J.; Dean, R.T. Radical-Mediated Protein Oxidation: From Chemistry to Medicine; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Chevion, M. A site-specific mechanism for free radical induced biological damage: The essential role of redox-active transition metals. Free Radic. Biol. Med. 1988, 5, 27–37. [Google Scholar] [CrossRef]
- Stadtman, E.R. Metal ion-catalyzed oxidation of proteins: Biochemical mechanism and biological consequences. Free Radic. Biol. Med. 1990, 9, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.J.; Fu, S.; Dean, R.T. Protein hydroperoxides can give rise to reactive free radicals. Biochem. J. 1995, 305, 643–649. [Google Scholar] [CrossRef] [Green Version]
- Buettner, G.R.; Jurkiewicz, B.A. Catalytic metals, ascorbate and free radicals: Combinations to avoid. Radiat. Res. 1996, 145, 532–541. [Google Scholar] [CrossRef]
- Von Sonntag, C. Peroxyl Radicals in Aqueous Media. In Oxygen Radicals in Biology and Medicine; Simic, M.G., Taylor, K.A., Ward, J.F., von Sonntag, C., Eds.; Springer: Berlin/Heidelberg, Germany, 1988; Volume 49. [Google Scholar]
- Von Sonntag, C.; Schuchmann, H.-P. Peroxyl radicals in aqueous solution. In Peroxyl Radicals; Alfassi, Z.B., Ed.; John Wiley and Sons: Chichester, UK, 1997; pp. 173–234. [Google Scholar]
- Bothe, E.; Schuchmann, M.N.; Schultefrohlinde, D.; Vonsonntag, C. Ho2 Elimination from Alpha-Hydroxyalkylperoxyl Radicals in Aqueous-Solution. Photochem. Photobiol. 1978, 28, 639–644. [Google Scholar] [CrossRef]
- Niki, E. Role of vitamin E as a lipid-soluble peroxyl radical scavenger: In vitro and in vivo evidence. Free Radic. Biol. Med. 2014, 66, 3–12. [Google Scholar] [CrossRef]
- El-Agamey, A.; McGarvey, D.J. Peroxyl radical reactions with carotenoids in microemulsions: Influence of microemulsion composition and the nature of peroxyl radical precursor. Free Radic. Biol. Med. 2016, 90, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, B.C.; Holmes, R.G.G.; Laue, H.A.H.; Norman, R.O.C. Electron-Spin Resonance Studies. 50. Reactions of Alkoxyl Radicals Generated from Alkyl Hydroperoxides and Titanium(Iii) Ion in Aqueous-Solution. J. Chem. Soc. Perk. Trans. 1976, 2, 1047–1052. [Google Scholar] [CrossRef]
- Gilbert, B.C.; David, P.; Marshall, R.; Norman, R.O.C.; Pineda, N.; Williams, P.S. Electron-Spin Resonance Studies. 61. The Generation and Reactions of the Tert-Butoxyl Radical in Aqueous-Solution. J. Chem. Soc. Perk. Trans. 1981, 2, 1392–1400. [Google Scholar] [CrossRef]
- Headlam, H.A.; Davies, M.J. Beta-scission of side-chain alkoxyl radicals on peptides and proteins results in the loss of side-chains as aldehydes and ketones. Free Radic. Biol. Med. 2002, 32, 1171–1184. [Google Scholar] [CrossRef]
- Bors, W.; Tait, D.; Michel, C.; Saran, M.; Erbenruss, M. Reactions of Alkoxy Radicals in Aqueous-Solutions. Isr. J. Chem. 1984, 24, 17–24. [Google Scholar] [CrossRef]
- Erbenruss, M.; Michel, C.; Bors, W.; Saran, M. Absolute Rate Constants of Alkoxyl Radical Reactions in Aqueous-Solution. J. Phys. Chem. 1987, 91, 2362–2365. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Hyde, J.S. Concentration of oxygen in lipid bilayers using a spin-label method. Biophys. J. 1983, 41, 283–286. [Google Scholar] [CrossRef]
- Beal, M.F.; Ferrante, R.J.; Browne, S.E.; Matthews, R.T.; Kowall, N.W.; Brown, R.H., Jr. Increased 3-nitrotyrosine in both sporadic and familial amyotrophic lateral sclerosis. Ann. Neurol. 1997, 42, 644–654. [Google Scholar] [CrossRef]
- Ferrante, R.J.; Browne, S.E.; Shinobu, L.A.; Bowling, A.C.; Baik, M.J.; MacGarvey, U.; Kowall, N.W.; Brown, R.H., Jr.; Beal, M.F. Evidence of increased oxidative damage in both sporadic and familial amyotrophic lateral sclerosis. J. Neurochem. 1997, 69, 2064–2074. [Google Scholar] [CrossRef]
- Giasson, B.I.; Duda, J.E.; Murray, I.V.; Chen, Q.; Souza, J.M.; Hurtig, H.I.; Ischiropoulos, H.; Trojanowski, J.Q.; Lee, V.M. Oxidative damage linked to neurodegeneration by selective alpha-synuclein nitration in synucleinopathy lesions. Science 2000, 290, 985–989. [Google Scholar] [CrossRef]
- Hensley, K.; Maidt, M.L.; Yu, Z.; Sang, H.; Markesbery, W.R.; Floyd, R.A. Electrochemical analysis of protein nitrotyrosine and dityrosine in the Alzheimer brain indicates region-specific accumulation. J. Neurosci. 1998, 18, 8126–8132. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Reed, T.; Newman, S.F.; Sultana, R. Roles of amyloid beta-peptide-associated oxidative stress and brain protein modifications in the pathogenesis of Alzheimer’s disease and mild cognitive impairment. Free Radic. Biol. Med. 2007, 43, 658–677. [Google Scholar] [CrossRef]
- Selfridge, J.E.; E., L.; Lu, J.; Swerdlow, R.H. Role of mitochondrial homeostasis and dynamics in Alzheimer’s disease. Neurobiol. Dis. 2013, 51, 3–12. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, B. Oxidative stress and the pathogenesis of Alzheimer’s disease. Oxid. Med. Cell. Longev. 2013, 2013, 316523. [Google Scholar] [CrossRef]
- Hands, S.; Sajjad, M.U.; Newton, M.J.; Wyttenbach, A. In vitro and in vivo aggregation of a fragment of huntingtin protein directly causes free radical production. J. Biol. Chem. 2011, 286, 44512–44520. [Google Scholar] [CrossRef]
- Markesbery, W.R. Oxidative Stress Hypothesis in Alzheimer’s Disease. Free Radic. Biol. Med. 1997, 23, 134–147. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Drake, J.; Pocernich, C.; Castegna, A. Evidence of oxidative damage in Alzheimer’s disease brain: Central role for amyloid β-peptide. Trends Mol. Med. 2001, 7, 548–554. [Google Scholar] [CrossRef]
- Naslund, J.; Schierhorn, A.; Hellman, U.; Lannfelt, L.; Roses, A.D.; Tjernberg, L.O.; Silberring, J.; Gandy, S.E.; Winblad, B.; Greengard, P.; et al. Relative abundance of Alzheimer A beta amyloid peptide variants in Alzheimer disease and normal aging. Proc. Natl. Acad. Sci. USA 1994, 91, 8378–8382. [Google Scholar] [CrossRef]
- Cheignon, C.; Tomas, M.; Bonnefont-Rousselot, D.; Faller, P.; Hureau, C.; Collin, F. Oxidative stress and the amyloid beta peptide in Alzheimer’s disease. Redox Biol. 2018, 14, 450–464. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Lauderback, C.M. Lipid peroxidation and protein oxidation in Alzheimer’s disease brain: Potential causes and consequences involving amyloid beta-peptide-associated free radical oxidative stress. Free Radic. Biol. Med. 2002, 32, 1050–1060. [Google Scholar] [CrossRef]
- Sultana, R.; Boyd-Kimball, D.; Poon, H.F.; Cai, J.; Pierce, W.M.; Klein, J.B.; Merchant, M.; Markesbery, W.R.; Butterfield, D.A. Redox proteomics identification of oxidized proteins in Alzheimer’s disease hippocampus and cerebellum: An approach to understand pathological and biochemical alterations in AD. Neurobiol. Aging 2006, 27, 1564–1576. [Google Scholar] [CrossRef]
- Hensley, K.; Hall, N.; Subramaniam, R.; Cole, P.; Harris, M.; Aksenov, M.; Aksenova, M.; Gabbita, S.P.; Wu, J.F.; Carney, J.M.; et al. Brain regional correspondence between Alzheimer’s disease histopathology and biomarkers of protein oxidation. J. Neurochem. 1995, 65, 2146–2156. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Yatin, S.M.; Varadarajan, S.; Koppal, T. Amyloid beta-peptide-associated free radical oxidative stress, neurotoxicity, and Alzheimer’s disease. Methods Enzym. 1999, 309, 746–768. [Google Scholar]
- Nitsch, R.M.; Blusztajn, J.K.; Pittas, A.G.; Slack, B.E.; Growdon, J.H.; Wurtman, R.J. Evidence for a membrane defect in Alzheimer disease brain. Proc. Natl. Acad. Sci. USA 1992, 89, 1671–1675. [Google Scholar] [CrossRef]
- Svennerholm, L.; Gottfries, C.G. Membrane lipids, selectively diminished in Alzheimer brains, suggest synapse loss as a primary event in early-onset form (type I) and demyelination in late-onset form (type II). J. Neurochem. 1994, 62, 1039–1047. [Google Scholar] [CrossRef]
- Markesbery, W.R.; Lovell, M.A. Four-hydroxynonenal, a product of lipid peroxidation, is increased in the brain in Alzheimer’s disease. Neurobiol. Aging 1998, 19, 33–36. [Google Scholar] [CrossRef]
- Neely, M.D.; Sidell, K.R.; Graham, D.G.; Montine, T.J. The lipid peroxidation product 4-hydroxynonenal inhibits neurite outgrowth, disrupts neuronal microtubules, and modifies cellular tubulin. J. Neurochem. 1999, 72, 2323–2333. [Google Scholar] [CrossRef]
- Keller, J.N.; Pang, Z.; Geddes, J.W.; Begley, J.G.; Germeyer, A.; Waeg, G.; Mattson, M.P. Impairment of glucose and glutamate transport and induction of mitochondrial oxidative stress and dysfunction in synaptosomes by amyloid beta-peptide: Role of the lipid peroxidation product 4-hydroxynonenal. J. Neurochem. 1997, 69, 273–284. [Google Scholar] [CrossRef]
- Hardas, S.S.; Sultana, R.; Clark, A.M.; Beckett, T.L.; Szweda, L.I.; Murphy, M.P.; Butterfield, D.A. Oxidative modification of lipoic acid by HNE in Alzheimer disease brain. Redox Biol. 2013, 1, 80–85. [Google Scholar] [CrossRef]
- Gegotek, A.; Skrzydlewska, E. Biological effect of protein modifications by lipid peroxidation products. Chem. Phys. Lipids 2019, 221, 46–52. [Google Scholar] [CrossRef]
- Sayre, L.M.; Zelasko, D.A.; Harris, P.L.; Perry, G.; Salomon, R.G.; Smith, M.A. 4-Hydroxynonenal-derived advanced lipid peroxidation end products are increased in Alzheimer’s disease. J. Neurochem. 1997, 68, 2092–2097. [Google Scholar] [CrossRef]
- Liu, Q.; Smith, M.A.; Avila, J.; DeBernardis, J.; Kansal, M.; Takeda, A.; Zhu, X.; Nunomura, A.; Honda, K.; Moreira, P.I.; et al. Alzheimer-specific epitopes of tau represent lipid peroxidation-induced conformations. Free Radic. Biol. Med. 2005, 38, 746–754. [Google Scholar] [CrossRef]
- Pryor, W.A.; Porter, N.A. Suggested mechanisms for the production of 4-hydroxy-2-nonenal from the autoxidation of polyunsaturated fatty acids. Free Radic. Biol. Med. 1990, 8, 541–543. [Google Scholar] [CrossRef]
- Montine, T.J.; Markesbery, W.R.; Morrow, J.D.; Roberts, L.J., 2nd. Cerebrospinal fluid F2-isoprostane levels are increased in Alzheimer’s disease. Ann. Neurol. 1998, 44, 410–413. [Google Scholar] [CrossRef]
- Nourooz-Zadeh, J.; Liu, E.H.; Yhlen, B.; Anggard, E.E.; Halliwell, B. F4-isoprostanes as specific marker of docosahexaenoic acid peroxidation in Alzheimer’s disease. J. Neurochem. 1999, 72, 734–740. [Google Scholar] [CrossRef]
- Reich, E.E.; Markesbery, W.R.; Roberts, L.J., 2nd; Swift, L.L.; Morrow, J.D.; Montine, T.J. Brain regional quantification of F-ring and D-/E-ring isoprostanes and neuroprostanes in Alzheimer’s disease. Am. J. Pathol. 2001, 158, 293–297. [Google Scholar] [CrossRef]
- Smith, M.A.; Perry, G.; Richey, P.L.; Sayre, L.M.; Anderson, V.E.; Beal, M.F.; Kowall, N. Oxidative damage in Alzheimer’s. Nature 1996, 382, 120–121. [Google Scholar] [CrossRef] [PubMed]
- Tramutola, A.; Lanzillotta, C.; Perluigi, M.; Butterfield, D.A. Oxidative stress, protein modification and Alzheimer disease. Brain Res. Bull. 2016. [Google Scholar] [CrossRef] [PubMed]
- Terni, B.; Boada, J.; Portero-Otin, M.; Pamplona, R.; Ferrer, I. Mitochondrial ATP-synthase in the entorhinal cortex is a target of oxidative stress at stages I/II of Alzheimer’s disease pathology. Brain Pathol. 2010, 20, 222–233. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Boyd-Kimball, D. Redox proteomics and amyloid beta-peptide: Insights into Alzheimer disease. J. Neurochem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mecocci, P.; MacGarvey, U.; Beal, M.F. Oxidative damage to mitochondrial DNA is increased in Alzheimer’s disease. Ann. Neurol. 1994, 36, 747–751. [Google Scholar] [CrossRef]
- Gabbita, S.P.; Lovell, M.A.; Markesbery, W.R. Increased nuclear DNA oxidation in the brain in Alzheimer’s disease. J. Neurochem. 1998, 71, 2034–2040. [Google Scholar] [CrossRef]
- Lovell, M.A.; Soman, S.; Bradley, M.A. Oxidatively modified nucleic acids in preclinical Alzheimer’s disease (PCAD) brain. Mech. Ageing Dev. 2011, 132, 443–448. [Google Scholar] [CrossRef]
- Nunomura, A.; Perry, G.; Pappolla, M.A.; Wade, R.; Hirai, K.; Chiba, S.; Smith, M.A. RNA oxidation is a prominent feature of vulnerable neurons in Alzheimer’s disease. J. Neurosci. 1999, 19, 1959–1964. [Google Scholar] [CrossRef]
- Jenner, P. Oxidative stress in Parkinson’s disease. Ann. Neurol. 2003, 53 (Suppl. 3), S26–S38. [Google Scholar] [CrossRef]
- Bosco, D.A.; Fowler, D.M.; Zhang, Q.; Nieva, J.; Powers, E.T.; Wentworth, P., Jr.; Lerner, R.A.; Kelly, J.W. Elevated levels of oxidized cholesterol metabolites in Lewy body disease brains accelerate alpha-synuclein fibrilization. Nat. Chem. Biol. 2006, 2, 249–253. [Google Scholar] [CrossRef]
- Alam, Z.I.; Daniel, S.E.; Lees, A.J.; Marsden, D.C.; Jenner, P.; Halliwell, B. A generalised increase in protein carbonyls in the brain in Parkinson’s but not incidental Lewy body disease. J. Neurochem. 1997, 69, 1326–1329. [Google Scholar] [CrossRef] [PubMed]
- Nakabeppu, Y.; Tsuchimoto, D.; Yamaguchi, H.; Sakumi, K. Oxidative damage in nucleic acids and Parkinson’s disease. J. Neurosci. Res. 2007, 85, 919–934. [Google Scholar] [CrossRef]
- Selley, M.L. (E)-4-hydroxy-2-nonenal may be involved in the pathogenesis of Parkinson’s disease. Free Radic. Biol. Med. 1998, 25, 169–174. [Google Scholar] [CrossRef]
- Halliwell, B. The wanderings of a free radical. Free Radic. Biol. Med. 2009, 46, 531–542. [Google Scholar] [CrossRef]
- Jellinger, K.; Kienzl, E.; Rumpelmair, G.; Riederer, P.; Stachelberger, H.; Ben-Shachar, D.; Youdim, M.B. Iron-melanin complex in substantia nigra of parkinsonian brains: An x-ray microanalysis. J. Neurochem. 1992, 59, 1168–1171. [Google Scholar] [CrossRef]
- Fahn, S.; Cohen, G. The oxidant stress hypothesis in Parkinson’s disease: Evidence supporting it. Ann. Neurol. 1992, 32, 804–812. [Google Scholar] [CrossRef]
- Surgucheva, I.; Sharov, V.S.; Surguchov, A. gamma-Synuclein: Seeding of alpha-synuclein aggregation and transmission between cells. Biochemistry 2012, 51, 4743–4754. [Google Scholar] [CrossRef]
- Mendez, E.F.; Sattler, R. Biomarker development for C9orf72 repeat expansion in ALS. Brain Res. 2015, 1607, 26–35. [Google Scholar] [CrossRef]
- Smith, R.G.; Henry, Y.K.; Mattson, M.P.; Appel, S.H. Presence of 4-hydroxynonenal in cerebrospinal fluid of patients with sporadic amyotrophic lateral sclerosis. Ann. Neurol. 1998, 44, 696–699. [Google Scholar] [CrossRef]
- Bogdanov, M.; Brown, R.H.; Matson, W.; Smart, R.; Hayden, D.; O’Donnell, H.; Flint Beal, M.; Cudkowicz, M. Increased oxidative damage to DNA in ALS patients. Free Radic. Biol. Med. 2000, 29, 652–658. [Google Scholar] [CrossRef]
- Miller, E.; Morel, A.; Saso, L.; Saluk, J. Isoprostanes and neuroprostanes as biomarkers of oxidative stress in neurodegenerative diseases. Oxid. Med. Cell. Longev. 2014, 2014, 572491. [Google Scholar] [CrossRef]
- Mitsumoto, H.; Santella, R.M.; Liu, X.; Bogdanov, M.; Zipprich, J.; Wu, H.C.; Mahata, J.; Kilty, M.; Bednarz, K.; Bell, D.; et al. Oxidative stress biomarkers in sporadic ALS. Amyotroph. Lateral Scler. 2008, 9, 177–183. [Google Scholar] [CrossRef]
- Bozzo, F.; Mirra, A.; Carri, M.T. Oxidative stress and mitochondrial damage in the pathogenesis of ALS: New perspectives. Neurosci. Lett. 2017, 636, 3–8. [Google Scholar] [CrossRef]
- Blokhuis, A.M.; Groen, E.J.; Koppers, M.; van den Berg, L.H.; Pasterkamp, R.J. Protein aggregation in amyotrophic lateral sclerosis. Acta Neuropathol. 2013, 125, 777–794. [Google Scholar] [CrossRef] [Green Version]
- Lovejoy, D.B.; Guillemin, G.J. The potential for transition metal-mediated neurodegeneration in amyotrophic lateral sclerosis. Front. Aging Neurosci. 2014, 6, 173. [Google Scholar] [CrossRef]
- Rosas, H.D.; Chen, Y.I.; Doros, G.; Salat, D.H.; Chen, N.K.; Kwong, K.K.; Bush, A.; Fox, J.; Hersch, S.M. Alterations in brain transition metals in Huntington disease: An evolving and intricate story. Arch. Neurol. 2012, 69, 887–893. [Google Scholar] [CrossRef]
- Hersch, S.M.; Gevorkian, S.; Marder, K.; Moskowitz, C.; Feigin, A.; Cox, M.; Como, P.; Zimmerman, C.; Lin, M.; Zhang, L.; et al. Creatine in Huntington disease is safe, tolerable, bioavailable in brain and reduces serum 8OH2’dG. Neurology 2006, 66, 250–252. [Google Scholar] [CrossRef]
- Chen, C.M.; Wu, Y.R.; Cheng, M.L.; Liu, J.L.; Lee, Y.M.; Lee, P.W.; Soong, B.W.; Chiu, D.T. Increased oxidative damage and mitochondrial abnormalities in the peripheral blood of Huntington’s disease patients. Biochem. Biophys. Res. Commun. 2007, 359, 335–340. [Google Scholar] [CrossRef]
- Stack, E.C.; Matson, W.R.; Ferrante, R.J. Evidence of oxidant damage in Huntington’s disease: Translational strategies using antioxidants. Ann. N. Y. Acad. Sci. 2008, 1147, 79–92. [Google Scholar] [CrossRef]
- Dragunow, M.; Faull, R.L.; Lawlor, P.; Beilharz, E.J.; Singleton, K.; Walker, E.B.; Mee, E. In situ evidence for DNA fragmentation in Huntington’s disease striatum and Alzheimer’s disease temporal lobes. Neuroreport 1995, 6, 1053–1057. [Google Scholar] [CrossRef]
- Browne, S.E.; Ferrante, R.J.; Beal, M.F. Oxidative stress in Huntington’s disease. Brain Pathol. 1999, 9, 147–163. [Google Scholar] [CrossRef]
- Tunez, I.; Sanchez-Lopez, F.; Aguera, E.; Fernandez-Bolanos, R.; Sanchez, F.M.; Tasset-Cuevas, I. Important role of oxidative stress biomarkers in Huntington’s disease. J. Med. Chem. 2011, 54, 5602–5606. [Google Scholar] [CrossRef]
- LeBars, P.L.; Katz, M.M.; Berman, N.; Itil, T.M.; Freedman, A.M.; Schatzberg, A.F. A placebo-controlled, double-blind, randomized trial of an extract of Ginkgo biloba for dementia. J. Am. Med. Assoc. 1997, 278, 1327–1332. [Google Scholar]
- Purushothuman, S.; Nandasena, C.; Peoples, C.L.; El Massri, N.; Johnstone, D.M.; Mitrofanis, J.; Stone, J. Saffron Pre-Treatment Offers Neuroprotection to Nigral and Retinal Dopaminergic Cells of MPTP-Treated mice. J. Parkinson Dis. 2013, 3, 77–83. [Google Scholar]
- Barkats, M.; Millecamps, S.; Abrioux, P.; Geoffroy, M.C.; Mallet, J. Overexpression of glutathione peroxidase increases the resistance of neuronal cells to Abeta-mediated neurotoxicity. J. Neurochem. 2000, 75, 1438–1446. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, T.; Ziegler, A.C.; Dimitrion, P.; Zuo, L. Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications. Oxid. Med. Cell. Longev. 2017, 2017, 2525967. [Google Scholar] [CrossRef]
Figure 1.
The chemical basis of Reactive Oxygen Species (ROS) generation—primary radical and molecular species are produced by incomplete reduction of molecular oxygen and can further react with an organic substrate to generate substrate-derived ROS. Metal ions are engaged in electron transfer (through metalloenzymes in vivo), but also involved in both Fenton and Haber-Weiss reactions, and in the reduction of hydroperoxide into alkoxyl radical.
Figure 1.
The chemical basis of Reactive Oxygen Species (ROS) generation—primary radical and molecular species are produced by incomplete reduction of molecular oxygen and can further react with an organic substrate to generate substrate-derived ROS. Metal ions are engaged in electron transfer (through metalloenzymes in vivo), but also involved in both Fenton and Haber-Weiss reactions, and in the reduction of hydroperoxide into alkoxyl radical.

Figure 2.
The chemical reactions of the superoxide and the perhydroxyl radicals.

Figure 3.
The chemical reactions of the hydroxyl radical.

Figure 4.
The chemical reactions of hydrogen peroxide, peroxyl and alkoxyl radicals.

Figure 5.
The direct involvement of ROS in lipid peroxidation and protein carbonylation. (A) the mechanism of 4-HNE formation from ROS-induced polyunsaturated omega-6 fatty acid peroxidation (from Pryor and Porter [125]); 4-HNE is able to form adducts with lipoic acid, proteins (C, H and K residues) and DNA bases; (B) ROS-induced protein carbonylation and cleavage (from Stadtman and Levine [43]).
Figure 5.
The direct involvement of ROS in lipid peroxidation and protein carbonylation. (A) the mechanism of 4-HNE formation from ROS-induced polyunsaturated omega-6 fatty acid peroxidation (from Pryor and Porter [125]); 4-HNE is able to form adducts with lipoic acid, proteins (C, H and K residues) and DNA bases; (B) ROS-induced protein carbonylation and cleavage (from Stadtman and Levine [43]).

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Collin, F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 2407. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102407
AMA Style
Collin F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. International Journal of Molecular Sciences. 2019; 20(10):2407. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102407
Chicago/Turabian StyleCollin, Fabrice. 2019. "Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases" International Journal of Molecular Sciences 20, no. 10: 2407. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102407
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.