Insulin Sensitivity Is Associated with Lipoprotein Lipase (LPL) and Catenin Delta 2 (CTNND2) DNA Methylation in Peripheral White Blood Cells in Non-Diabetic Young Women

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Anthropometric and Biochemical Characteristics of the Participants
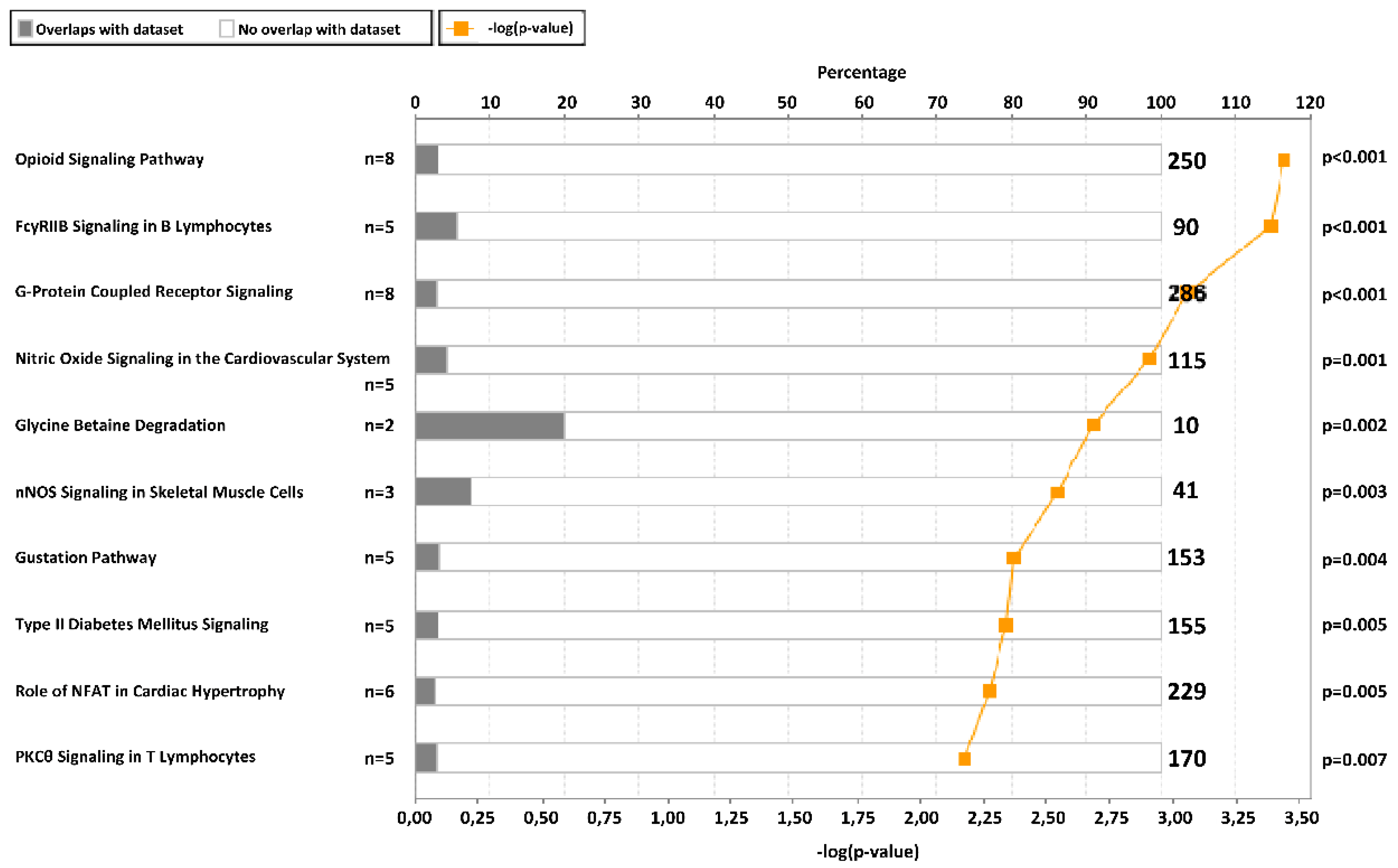
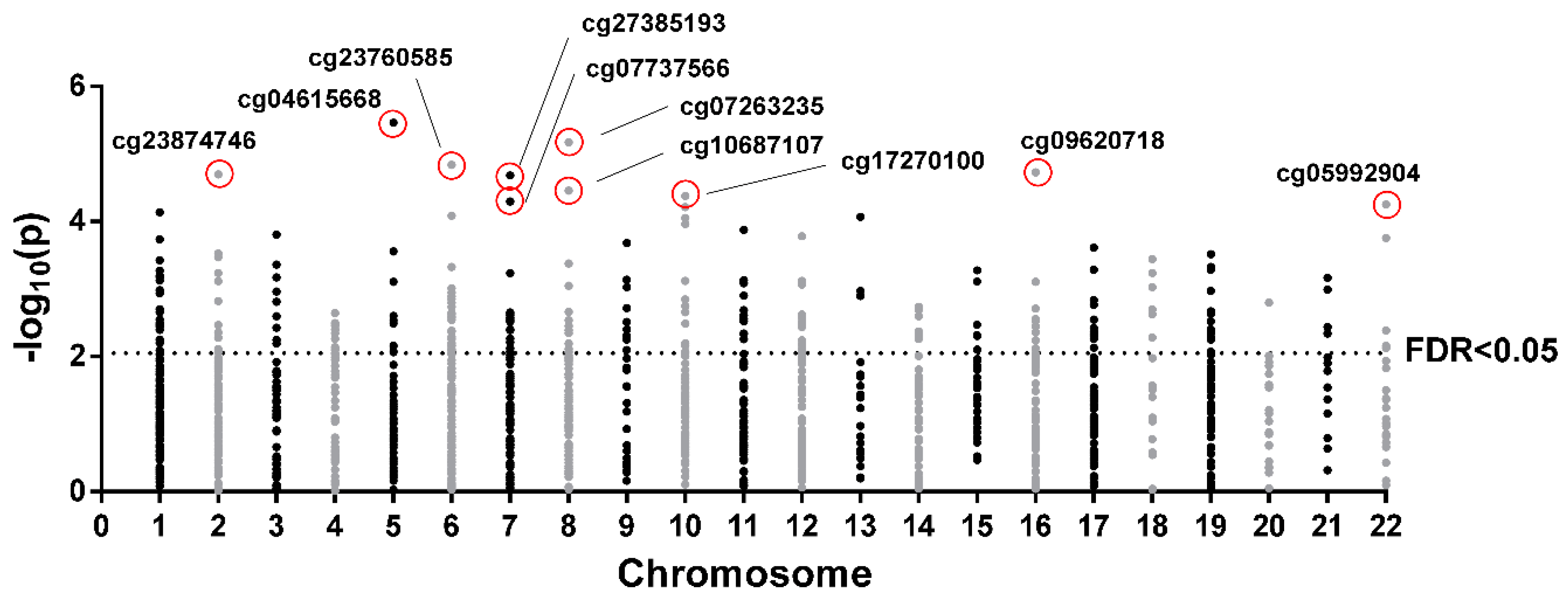
2.2. CpG Sites Selection and Ingenuity Pathway Analysis
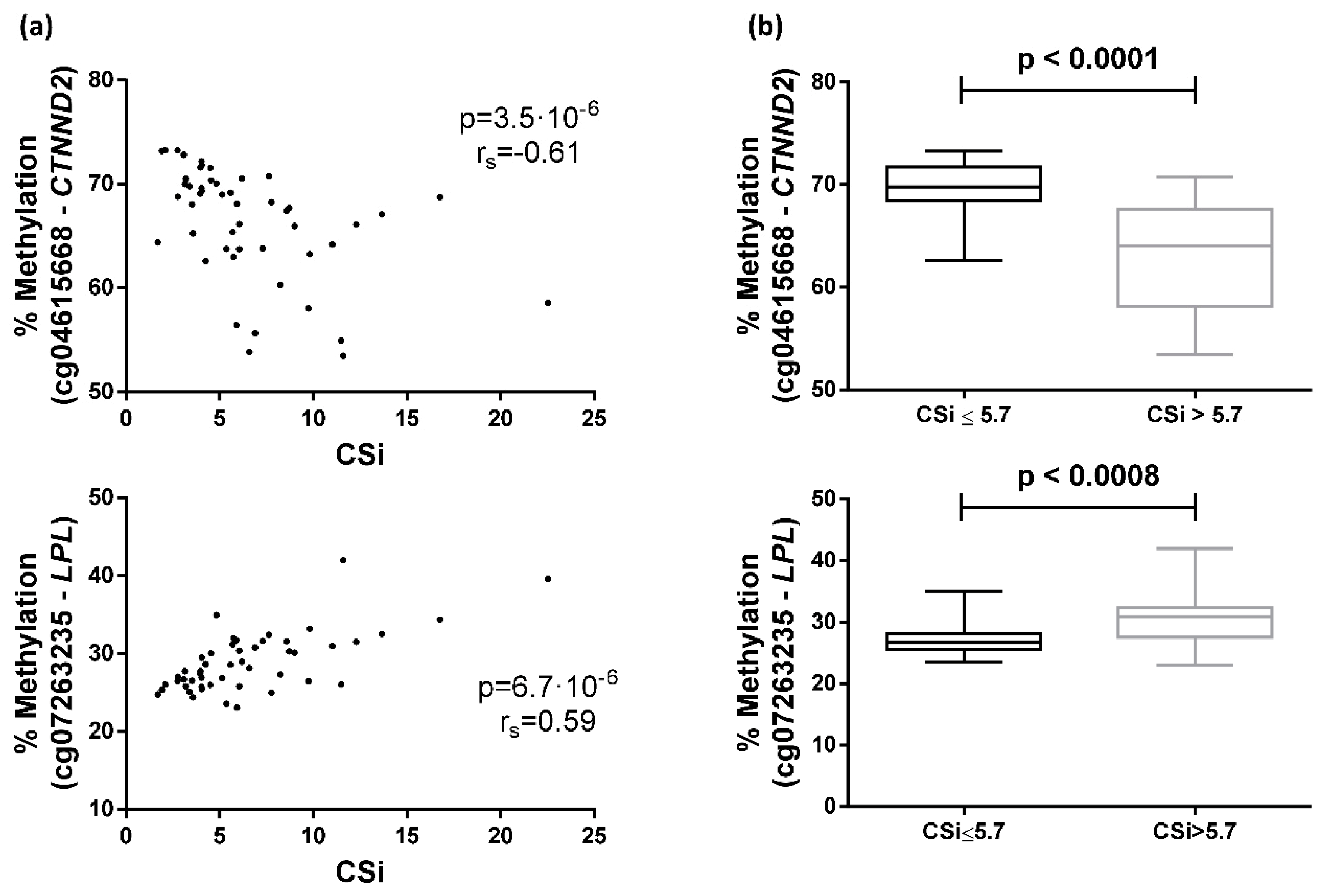
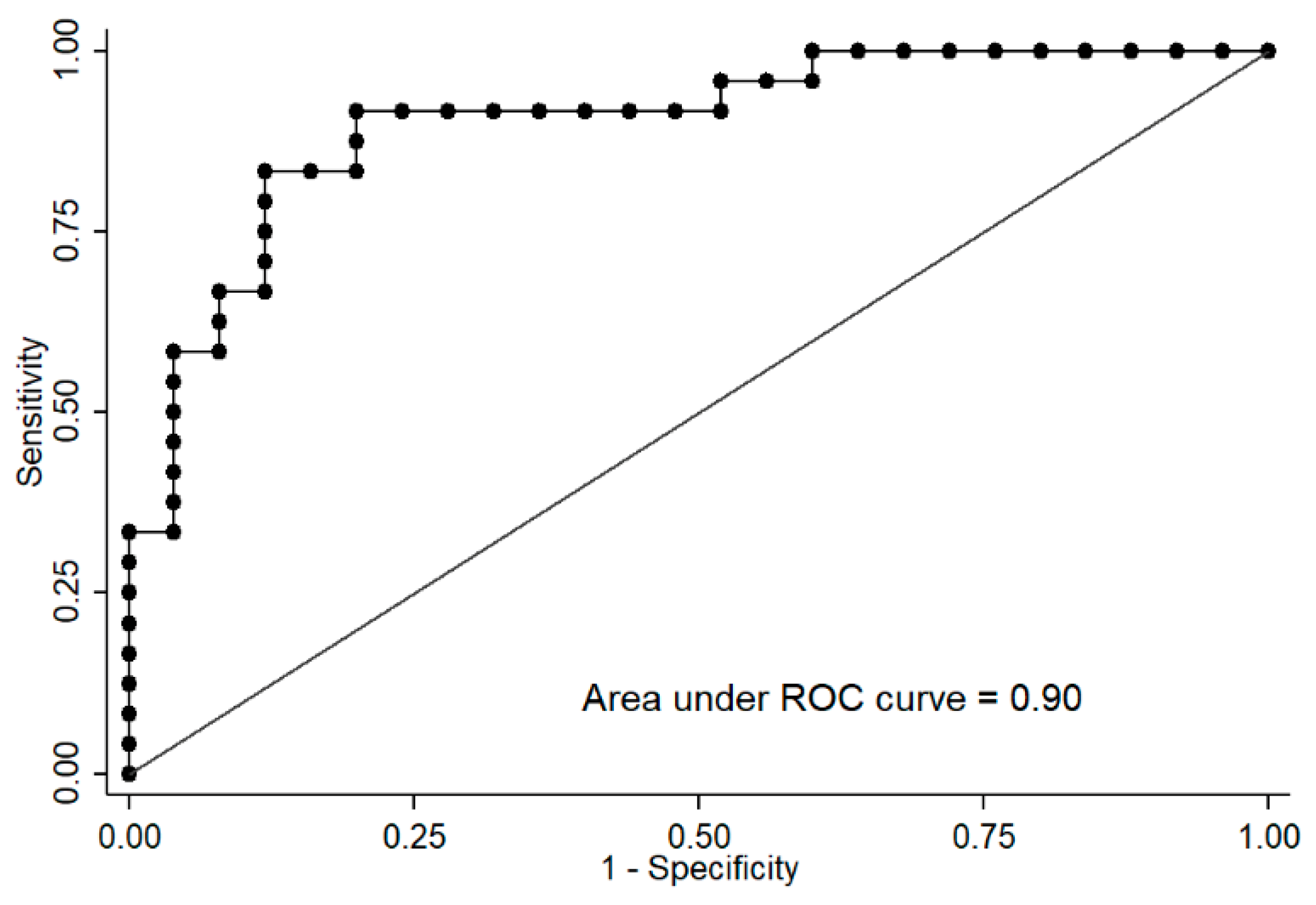
2.3. Differences between Groups Separated by the Median of CSi Values
2.4. Correlation with Other Variables
3. Discussion
4. Materials and Methods
4.1. Anthropometry, Blood Pressure and Biochemical Determinations
4.2. DNA Methylation Analysis
4.3. Treatment of Methylation Raw Data
4.4. Statistical Analysis
4.5. Ingenuity Pathway Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AIR | Acute insulin response |
| AUC | Area under the curve |
| CREB1 | Cyclic AMP-responsive element-binding protein 1 |
| CSi | Calculated insulin sensitivity index |
| DI | Disposition index |
| EWAS | Epigenome-wide association study |
| FDR | False discovery rate |
| GEDYMET | Genetics, dysglycemia and metabolism |
| GIP | Glucose-dependent insulinotropic polypeptide |
| GWAS | Genome-wide association study |
| HOMA-IR | Homeostasis model assessment - insulin resistance index |
| HOMA-S | HOMA-insulin sensitivity index |
| IPA | Ingenuity pathway analysis |
| IVGTT | Intravenous glucose tolerance test |
| LPL | Lipoprotein lipase |
| MENA | Methyl Epigenome Network Association |
| PWBCs | Peripheral white blood cells |
| ROC | Receiver operating characteristic |
| SNP | Single nucleotide polymorphism |
| T2D | Type 2 diabetes |
| TORC2 | CREB/cAMP-responsive CREB coactivator 2 |
References
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2014, 37, S81–S90. [Google Scholar] [CrossRef] [PubMed]
- Conn, V.S.; Koopman, R.J.; Ruppar, T.M.; Phillips, L.J.; Mehr, D.R.; Hafdahl, A.R. Insulin Sensitivity Following Exercise Interventions: Systematic Review and Meta-Analysis of Outcomes Among Healthy Adults. J. Prim. Care Community Health 2014, 5, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Meier, J.J.; Veldhuis, J.D.; Butler, P.C. Pulsatile Insulin Secretion Dictates Systemic Insulin Delivery by Regulating Hepatic Insulin Extraction in Humans. Diabetes 2005, 54, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Tobin, J.D.; Andres, R. Glucose clamp technique: A method for quantifying insulin secretion and resistance. Am. J. Physiol. 1979, 237, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Cobelli, C.; Toffolo, G.M.; Dalla Man, C.; Campioni, M.; Denti, P.; Caumo, A.; Butler, P.; Rizza, R. Assessment of beta-cell function in humans, simultaneously with insulin sensitivity and hepatic extraction, from intravenous and oral glucose tests. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1–E15. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Prigeon, R.L.; McCulloch, D.K.; Boyko, E.J.; Bergman, R.N.; Schwartz, M.W.; Neifing, J.L.; Ward, W.K.; Beard, J.C.; Palmer, J.P.; et al. Quantification of the relationship between insulin sensitivity and beta-cell function in human subjects. Evidence for a hyperbolic function. Diabetes 1993, 42, 1663–1672. [Google Scholar] [CrossRef]
- Cersosimo, E.; Solis-Herrera, C.; Trautmann, M.E.; Malloy, J.; Triplitt, C.L. Assessment of Pancreatic beta-Cell Function: Review of Methods and Clinical Applications. Curr. Diabetes Rev. 2014, 10, 2–42. [Google Scholar] [CrossRef]
- Tura, A.; Sbrignadello, S.; Succurro, E.; Groop, L.; Sesti, G.; Pacini, G. An empirical index of insulin sensitivity from short IVGTT: Validation against the minimal model and glucose clamp indices in patients with different clinical characteristics. Diabetologia 2010, 53, 144–152. [Google Scholar] [CrossRef]
- Bergman, R.N.; Ader, M.; Huecking, K.; Van Citters, G. Accurate assessment of beta-cell function: The hyperbolic correction. Diabetes 2002, 51, S212–S220. [Google Scholar] [CrossRef]
- Santos, J.L.; Yevenes, I.; Cataldo, L.R.; Morales, M.; Galgani, J.; Arancibia, C.; Vega, J.; Olmos, P.; Flores, M.; Valderas, J.P.; et al. Development and assessment of the disposition index based on the oral glucose tolerance test in subjects with different glycaemic status. J. Physiol. Biochem. 2016, 72, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Utzschneider, K.M.; Prigeon, R.L.; Faulenbach, M.V.; Tong, J.; Carr, D.B.; Boyko, E.J.; Leonetti, D.L.; McNeely, M.J.; Fujimoto, W.Y.; Kahn, S.E. Oral disposition index predicts the development of future diabetes above and beyond fasting and 2-h glucose levels. Diabetes Care 2009, 32, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Sookoian, S.; Pirola, C.J. Epigenetics of Insulin Resistance: An Emerging Field in Translational Medicine. Curr. Diabetes Rep. 2013, 13, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, B.; Irvin, M.R.; Sha, J.; Zhi, D.; Aslibekyan, S.; Absher, D.; Tiwari, H.K.; Kabagambe, E.K.; Ordovas, J.M.; Arnett, D.K. Epigenome-Wide Association Study of Fasting Measures of Glucose, Insulin, and HOMA-IR in the Genetics of Lipid Lowering Drugs and Diet Network Study. Diabetes 2014, 63, 801–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriebel, J.; Herder, C.; Rathmann, W.; Wahl, S.; Kunze, S.; Molnos, S.; Volkova, N.; Schramm, K.; Carstensen-Kirberg, M.; Waldenberger, M.; et al. Association between DNA Methylation in Whole Blood and Measures of Glucose Metabolism: KORA F4 Study. PLoS ONE 2016, 11, e0152314. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.C.; Loh, M.; Lehne, B.; Drong, A.; Kriebel, J.; Motta, V.; Wahl, S.; Elliott, H.R.; Rota, F.; Scott, W.R.; et al. Epigenome-wide association of DNA methylation markers in peripheral blood from Indian Asians and Europeans with incident type 2 diabetes: A nested case-control study. Lancet Diabetes Endocrinol. 2015, 3, 526–534. [Google Scholar] [CrossRef]
- Kulkarni, H.; Kos, M.Z.; Neary, J.; Dyer, T.D.; Kent, J.W.J.; Goring, H.H.; Cole, S.A.; Comuzzie, A.G.; Almasy, L.; Mahaney, M.C.; et al. Novel epigenetic determinants of type 2 diabetes in Mexican-American families. Hum. Mol. Genet. 2015, 24, 5330–5344. [Google Scholar] [CrossRef] [PubMed]
- Al Muftah, W.A.; Al-Shafai, M.; Zaghlool, S.B.; Visconti, A.; Tsai, P.C.; Kumar, P.; Spector, T.; Bell, J.; Falchi, M.; Suhre, K. Epigenetic associations of type 2 diabetes and BMI in an Arab population. Clin. Epigenetics 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Dayeh, T.; Tuomi, T.; Almgren, P.; Perfilyev, A.; Jansson, P.A.; De Mello, V.D.; Pihlajamaki, J.; Vaag, A.; Groop, L.; Nilsson, E.; et al. DNA methylation of loci withinABCG1andPHOSPHO1in blood DNA is associated with future type 2 diabetes risk. Epigenetics 2016, 11, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.; Drong, A.; Lehne, B.; Loh, M.; Scott, W.R.; Kunze, S.; Tsai, P.C.; Ried, J.S.; Zhang, W.; Yang, Y.; et al. Epigenome-wide association study of body mass index, and the adverse outcomes of adiposity. Nature 2017, 541, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Muka, T.; Nano, J.; Voortman, T.; Braun, K.V.E.; Ligthart, S.; Stranges, S.; Bramer, W.M.; Troup, J.; Chowdhury, R.; Dehghan, A.; et al. The role of global and regional DNA methylation and histone modifications in glycemic traits and type 2 diabetes: A systematic review. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Sun, B.; Li, X.; Zhu, C. DNA methylation landscapes in the pathogenesis of type 2 diabetes mellitus. Nutr. Metab. 2018, 14, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Sagesaka, H.; Sato, Y.; Someya, Y.; Tamura, Y.; Shimodaira, M.; Miyakoshi, T.; Hirabayashi, K.; Koike, H.; Yamashita, K.; Watada, H.; et al. Type 2 Diabetes: When Does It Start? J. Endocr. Soc. 2018, 2, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Szabo, M.; Mate, B.; Csep, K.; Benedek, T. Epigenetic Modifications Linked to T2D, the Heritability Gap, and Potential Therapeutic Targets. Biochem. Genet. 2018, 56, 553–574. [Google Scholar] [CrossRef] [PubMed]
- Davegardh, C.; Garcia-Calzon, S.; Bacos, K.; Ling, C. DNA methylation in the pathogenesis of type 2 diabetes in humans. Mol. Metab. 2018, 14, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Su, C.F.; Chang, Y.Y.; Pai, H.H.; Liu, I.M.; Lo, C.Y.; Cheng, J.T. Infusion of beta-Endorphin Improves Insulin Resistance in Fructose-fed Rats. Horm. Metab. Res. 2004, 36, 571–577. [Google Scholar] [CrossRef]
- Li, Y.; Eitan, S.; Wu, J.; Evans, C.J.; Kieffer, B.; Sun, X.; Polakiewicz, R.D. Morphine induces desensitization of insulin receptor signaling. Mol. Cell Biol. 2003, 23, 6255–6266. [Google Scholar] [CrossRef]
- Layden, B.T.; Durai, V.; Lowe, J.W.L. G-Protein-Coupled Receptors, Pancreatic Islets, and Diabetes. Nat. Educ. 2010, 3, 13. [Google Scholar]
- Walford, G.A.; Ma, Y.; Clish, C.; Florez, J.C.; Wang, T.J.; Gerszten, R.E. Diabetes Prevention Program Research G Metabolite Profiles of Diabetes Incidence and Intervention Response in the Diabetes Prevention Program. Diabetes 2016, 65, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Sansbury, B.E.; Hill, B.G. Regulation of obesity and insulin resistance by nitric oxide. Free Radic. Boil. Med. 2014, 73, 383–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botker, H.E.; Moller, N. ON NO-the continuing story of nitric oxide, diabetes, and cardiovascular disease. Diabetes 2013, 62, 2645–2647. [Google Scholar] [CrossRef] [PubMed]
- Karimian Azari, E.; Smith, K.R.; Yi, F.; Osborne, T.F.; Bizzotto, R.; Mari, A.; Pratley, R.E.; Kyriazis, G.A. Inhibition of sweet chemosensory receptors alters insulin responses during glucose ingestion in healthy adults: A randomized crossover interventional study. Am. J. Clin. Nutr. 2017, 105, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, M.O.; Taylor, K.D.; Guo, X.; Hokanson, J.E.; Haffner, S.M.; Cui, J.; Chen, Y.D.; Wagenknecht, L.E.; Bergman, R.N.; Rotter, J.I. Haplotypes in the lipoprotein lipase gene influence fasting insulin and discovery of a new risk haplotype. J. Clin. Endocrinol. Metab. 2007, 92, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Ramos, F.J.; Langlais, P.R.; Hu, D.; Dong, L.Q.; Liu, F. Grb10 mediates insulin-stimulated degradation of the insulin receptor: A mechanism of negative regulation. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1262–E1266. [Google Scholar] [CrossRef] [PubMed]
- Horbelt, T.; Tacke, C.; Markova, M.; Herzfeld de Wiza, D.; Van de Velde, F.; Bekaert, M.; Van Nieuwenhove, Y.; Hornemann, S.; Rodiger, M.; Seebeck, N.; et al. The novel adipokine WISP1 associates with insulin resistance and impairs insulin action in human myotubes and mouse hepatocytes. Diabetologia 2018, 61, 2054–2065. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Navarrete, J.M.; Ortega, F.; Moreno, M.; Xifra, G.; Ricart, W.; Fernandez-Real, J.M. PRDM16 sustains white fat gene expression profile in human adipocytes in direct relation with insulin action. Mol. Cell Endocrinol. 2015, 405, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Ottosson Laakso, E.; Krus, U.; Storm, P.; Prasad, R.B.; Oskolkov, N.; Ahlqvist, E.; Fadista, J.; Hansson, O.; Groop, L.; Vikman, P. Glucose-Induced Changes in Gene Expression in Human Pancreatic Islets: Causes or Consequences of Chronic Hyperglycemia. Diabetes 2017, 66, 3013–3028. [Google Scholar] [CrossRef] [Green Version]
- Graae, A.S.; Grarup, N.; Ribel Madsen, R.; Lystbaek, S.H.; Boesgaard, T.; Staiger, H.; Fritsche, A.; Wellner, N.; Sulek, K.; Kjolby, M.; et al. ADAMTS9 Regulates Skeletal Muscle Insulin Sensitivity Through Extracellular Matrix Alterations. Diabetes 2019, 68, 502–514. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Williams, K.J. NOX4 pathway as a source of selective insulin resistance and responsiveness. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1236–1245. [Google Scholar] [CrossRef]
- Lu, Q.; Aguilar, B.J.; Li, M.; Jiang, Y.; Chen, Y.H. Genetic alterations of delta-catenin/NPRAP/Neurojungin (CTNND2): Functional implications in complex human diseases. Hum. Genet. 2016, 135, 1107–1116. [Google Scholar] [CrossRef]
- Turner, T.N.; Sharma, K.; Oh, E.C.; Liu, Y.P.; Collins, R.L.; Sosa, M.X.; Auer, D.R.; Brand, H.; Sanders, S.J.; Moreno De Luca, D.; et al. Loss of delta-catenin function in severe autism. Nature 2015, 520, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Diabetes Genetics Initiative of Broad Institute of Harvard and MIT, Lund University, and Novartis Institutes of BioMedical Research; Saxena, R.; Voight, B.F.; Lyssenko, V.; Burtt, N.P.; de Bakker, P.I.; Chen, H.; Roix, J.J.; Kathiresan, S.; Hirschhorn, J.N.; et al. Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science 2007, 316, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Zeggini, E.; Weedon, M.N.; Lindgren, C.M.; Frayling, T.M.; Elliott, K.S.; Lango, H.; Timpson, N.J.; Perry, J.R.; Rayner, N.W.; Freathy, R.M.; et al. Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science 2007, 316, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.S.; Heard Costa, N.; Cupples, L.A.; Dupuis, J.; Vasan, R.S.; Atwood, L.D. Genome-wide association to body mass index and waist circumference: The Framingham Heart Study 100K project. BMC Med. Genet. 2007, 8, S18. [Google Scholar] [CrossRef] [PubMed]
- Pingitore, P.; Lepore, S.M.; Pirazzi, C.; Mancina, R.M.; Motta, B.M.; Valenti, L.; Berge, K.E.; Retterstol, K.; Leren, T.P.; Wiklund, O.; et al. Identification and characterization of two novel mutations in the LPL gene causing type I hyperlipoproteinemia. J. Clin. Lipidol. 2016, 10, 816–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kersten, S. Physiological regulation of lipoprotein lipase. Biochim. Biophys. Acta 2014, 1841, 919–933. [Google Scholar] [CrossRef] [PubMed]
- He, P.P.; Jiang, T.; Ouyang, X.P.; Liang, Y.Q.; Zou, J.Q.; Wang, Y.; Shen, Q.Q.; Liao, L.; Zheng, X.L. Lipoprotein lipase: Biosynthesis, regulatory factors, and its role in atherosclerosis and other diseases. Clin. Chim. Acta 2018, 480, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, I.J.; Eckel, R.H.; Abumrad, N.A. Regulation of fatty acid uptake into tissues: Lipoprotein lipase- and CD36-mediated pathways. J. Lipid Res. 2009, 50, S86–S90. [Google Scholar] [CrossRef]
- Farese, R.V., Jr.; Yost, T.J.; Eckel, R.H. Tissue-specific regulation of lipoprotein lipase activity by insulin/glucose in normal-weight humans. Metabolism 1991, 40, 214–216. [Google Scholar] [CrossRef]
- Yost, T.J.; Jensen, D.R.; Haugen, B.R.; Eckel, R.H. Effect of dietary macronutrient composition on tissue-specific lipoprotein lipase activity and insulin action in normal-weight subjects. Am. J. Clin. Nutr. 1998, 68, 296–302. [Google Scholar] [CrossRef]
- Kim, J.K.; Fillmore, J.J.; Chen, Y.; Yu, C.; Moore, I.K.; Pypaert, M.; Lutz, E.P.; Kako, Y.; Velez-Carrasco, W.; Goldberg, I.J.; et al. Tissue-specific overexpression of lipoprotein lipase causes tissue-specific insulin resistance. Proc. Natl. Acad. Sci. USA 2001, 98, 7522–7527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodarzi, M.O.; Guo, X.; Taylor, K.D.; Quinones, M.J.; Saad, M.F.; Yang, H.; Hsueh, W.A.; Rotter, J.I. Lipoprotein lipase is a gene for insulin resistance in Mexican Americans. Diabetes 2004, 53, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Houde, A.A.; St Pierre, J.; Hivert, M.F.; Baillargeon, J.P.; Perron, P.; Gaudet, D.; Brisson, D.; Bouchard, L. Placental lipoprotein lipase DNA methylation levels are associated with gestational diabetes mellitus and maternal and cord blood lipid profiles. J. Dev. Orig. Health Dis. 2014, 5, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Castellano Castillo, D.; Moreno Indias, I.; Fernandez Garcia, J.C.; Alcaide Torres, J.; Moreno Santos, I.; Ocana, L.; Gluckman, E.; Tinahones, F.; Queipo Ortuno, M.I.; Cardona, F. Adipose Tissue LPL Methylation is Associated with Triglyceride Concentrations in the Metabolic Syndrome. Clin. Chem. 2018, 64, 210–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drogan, D.; Boeing, H.; Janke, J.; Schmitt, B.; Zhou, Y.; Walter, J.; Pischon, T.; Tierling, S. Regional distribution of body fat in relation to DNA methylation within the LPL, ADIPOQ and PPARgamma promoters in subcutaneous adipose tissue. Nutr. Diabetes 2015, 5, e168. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Nian, C.; McIntosh, C.H. GIP increases human adipocyte LPL expression through CREB and TORC2-mediated trans-activation of the LPL gene. J. Lipid Res. 2010, 51, 3145–3157. [Google Scholar] [CrossRef] [Green Version]
- Sinkeler, S.J.; Kwakernaak, A.J.; Bakker, S.J.; Shahinfar, S.; Esmatjes, E.; de Zeeuw, D.; Navis, G.; Lambers Heerspink, H.J. Creatinine excretion rate and mortality in type 2 diabetes and nephropathy. Diabetes Care 2013, 36, 1489–1494. [Google Scholar] [CrossRef]
- Bao, X.; Gu, Y.; Zhang, Q.; Liu, L.; Meng, G.; Wu, H.; Xia, Y.; Shi, H.; Wang, H.; Sun, S.; et al. Low serum creatinine predicts risk for type 2 diabetes. Diabetes Metab. Res. Rev. 2018, 34, e3011. [Google Scholar] [CrossRef]
- Takeuchi, M.; Imano, H.; Muraki, I.; Shimizu, Y.; Hayama Terada, M.; Kitamura, A.; Okada, T.; Kiyama, M.; Iso, H. Serum creatinine levels and risk of incident type 2 diabetes mellitus or dysglycemia in middle-aged Japanese men: A retrospective cohort study. Vet. Rec. 2018, 6, e000492. [Google Scholar] [CrossRef]
- Bacos, K.; Gillberg, L.; Volkov, P.; Olsson, A.H.; Hansen, T.; Pedersen, O.; Gjesing, A.P.; Eiberg, H.; Tuomi, T.; Almgren, P.; et al. Blood-based biomarkers of age-associated epigenetic changes in human islets associate with insulin secretion and diabetes. Nat. Commun. 2016, 7, 11089. [Google Scholar] [CrossRef] [Green Version]
- Crujeiras, A.B.; Diaz Lagares, A.; Sandoval, J.; Milagro, F.I.; Navas Carretero, S.; Carreira, M.C.; Gomez, A.; Hervas, D.; Monteiro, M.P.; Casanueva, F.F.; et al. DNA methylation map in circulating leukocytes mirrors subcutaneous adipose tissue methylation pattern: A genome-wide analysis from non-obese and obese patients. Sci. Rep. 2017, 7, 41903. [Google Scholar] [CrossRef] [PubMed]
- Marcelli Tourvieille, S.; Hubert, T.; Pattou, F.; Vantyghem, M.C. Acute insulin response (AIR): Review of protocols and clinical interest in islet transplantation. Diabetes Metab. 2006, 32, 295–303. [Google Scholar] [CrossRef]
- Kahn, S.E. The relative contributions of insulin resistance and beta-cell dysfunction to the pathophysiology of Type 2 diabetes. Diabetologia 2003, 46, 3–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arpón, A.; Riezu Boj, J.I.; Milagro, F.I.; Razquin, C.; Martinez Gonzalez, M.A.; Corella, D.; Estruch, R.; Casas, R.; Fito, M.; Ros, E.; et al. Adherence to Mediterranean diet is associated with methylation changes in inflammation-related genes in peripheral blood cells. J. Physiol. Biochem. 2017, 73, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touleimat, N.; Tost, J. Complete pipeline for Infinium® Human Methylation 450K BeadChip data processing using subset quantile normalization for accurate DNA methylation estimation. Epigenomics 2012, 4, 325–341. [Google Scholar] [CrossRef]
- Team, R. RStudio: Integrated Development for R. Available online: http://www.rstudio.com/ (accessed on 5 June 2018).
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinform. 2012, 13, 86. [Google Scholar] [CrossRef]
- Volkmar, M.; Dedeurwaerder, S.; Cunha, D.A.; Ndlovu, M.N.; Defrance, M.; Deplus, R.; Calonne, E.; Volkmar, U.; Igoillo Esteve, M.; Naamane, N.; et al. DNA methylation profiling identifies epigenetic dysregulation in pancreatic islets from type 2 diabetic patients. EMBO J. 2012, 31, 1405–1426. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | N | Median (IQR) |
|---|---|---|
| Age (y) | 57 | 25 (22–30) |
| Weight (kg) | 57 | 59.5 (56.0–64.2) |
| Height (m) | 57 | 1.59 (1.56–1.63) |
| Body mass index (kg/m2) | 57 | 23.4 (21.8–25.4) |
| Plasma total cholesterol (mg/dL) | 56 | 167.5 (148.0–196.5) |
| Plasma HDL cholesterol (mg/dL) | 56 | 63.0 (51.5–71.5) |
| Plasma LDL cholesterol (mg/dL) | 56 | 84.0 (67.5–103.5) |
| Plasma triglycerides (mg/dL) | 56 | 92.5 (68.0–131.0) |
| Systolic blood pressure (mmHg) | 57 | 113 (104–119) |
| Diastolic blood pressure (mmHg) | 57 | 70 (65–75) |
| Fasting glucose (mg/dL) | 49 | 78 (74–82) |
| Fasting insulin (IU/µmL) | 49 | 7.0 (5.8–9.1) |
| HOMA-S | 48 | 71.5 (55.5–88.5) |
| Calculated insulin sensitivity (CSi) | 49 | 5.7 (4.0–8.2) |
| Acute insulin release (AIR) | 49 | 538.7 (398.5–718.6) |
| IVGTT-based disposition index (DI) | 49 | 2792.6 (2023.4–4136.4) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arpón, A.; Santos, J.L.; Milagro, F.I.; Cataldo, L.R.; Bravo, C.; Riezu-Boj, J.-I.; Martínez, J.A. Insulin Sensitivity Is Associated with Lipoprotein Lipase (LPL) and Catenin Delta 2 (CTNND2) DNA Methylation in Peripheral White Blood Cells in Non-Diabetic Young Women. Int. J. Mol. Sci. 2019, 20, 2928. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122928
Arpón A, Santos JL, Milagro FI, Cataldo LR, Bravo C, Riezu-Boj J-I, Martínez JA. Insulin Sensitivity Is Associated with Lipoprotein Lipase (LPL) and Catenin Delta 2 (CTNND2) DNA Methylation in Peripheral White Blood Cells in Non-Diabetic Young Women. International Journal of Molecular Sciences. 2019; 20(12):2928. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122928
Chicago/Turabian StyleArpón, Ana, José L. Santos, Fermín I. Milagro, Luis Rodrigo Cataldo, Carolina Bravo, José-Ignacio Riezu-Boj, and J. Alfredo Martínez. 2019. "Insulin Sensitivity Is Associated with Lipoprotein Lipase (LPL) and Catenin Delta 2 (CTNND2) DNA Methylation in Peripheral White Blood Cells in Non-Diabetic Young Women" International Journal of Molecular Sciences 20, no. 12: 2928. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122928