Salicylic Acid Signals Plant Defence against Cadmium Toxicity

1
Institute of Environment, Resource, Soil and Fertilizer, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
2
Key Laboratory of Environment Remediation and Ecological Health, Ministry of Education, Hangzhou 310058, China
3
College of Environmental and Resource Sciences, Zhejiang University, Hangzhou 310058, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(12), 2960; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122960
Submission received: 20 May 2019
/
Revised: 6 June 2019
/
Accepted: 13 June 2019
/
Published: 18 June 2019
(This article belongs to the Special Issue Salicylic Acid Signalling in Plants)
Abstract
:Salicylic acid (SA), as an enigmatic signalling molecule in plants, has been intensively studied to elucidate its role in defence against biotic and abiotic stresses. This review focuses on recent research on the role of the SA signalling pathway in regulating cadmium (Cd) tolerance in plants under various SA exposure methods, including pre-soaking, hydroponic exposure, and spraying. Pretreatment with appropriate levels of SA showed a mitigating effect on Cd damage, whereas an excessive dose of exogenous SA aggravated the toxic effects of Cd. SA signalling mechanisms are mainly associated with modification of reactive oxygen species (ROS) levels in plant tissues. Then, ROS, as second messengers, regulate a series of physiological and genetic adaptive responses, including remodelling cell wall construction, balancing the uptake of Cd and other ions, refining the antioxidant defence system, and regulating photosynthesis, glutathione synthesis and senescence. These findings together elucidate the expanding role of SA in phytotoxicology.

1. Introduction
Cadmium (Cd) is one of the most toxic pollutants for all living organisms with a long biological half-life [1,2]. It exists in soil naturally by weathering of the parent rocks and accumulates through anthropogenic activities, such as deposition of airborne Cd from smelting and mining, wastewater irrigation, application of Cd-contaminated phosphate fertilizers and soil amendments [3,4,5,6]. Cd is a non-essential element for plants, and its uptake by plants has posed a serious health issue to humans through the food chain. For this reason, Cd phytotoxicity is a major subject in current research on plant biology [7].
Cadmium can be absorbed easily by plant roots, and transported to plant shoots, leading to various visible toxic symptoms, such as growth retardation, wilting, leaf chlorosis, and cell death [8,9]. The mechanisms of Cd toxicity include replacing and inactivating essential elements and destroying protein structure, thereby interfering with various physiological processes, such as photosynthesis [10], respiration [11], element assimilation [12], and cell division [13]. To defend against Cd injury, plants utilize many coordinated strategies, such as binding Cd to the cell wall [14], vacuolar retention through chelation via phytochelatins (PCs) [15] and upregulation of the antioxidant system [16]. Since all of the above mechanisms are closely cross-linked, it is important for plant survival that these metabolic reactions are adjusted accordingly via regulating signals.
Salicylic acid (SA), a simple phenolic molecule, has long been recognized as a potent phytohormone that regulates plant development and defence in higher plants [17]. The synthesis of SA occurs by two distinct and compartmentalized pathways. One pathway derives from phenylalanine and takes place in the cytoplasm. First, phenylalanine ammonia-lyase (PAL) converts phenylalanine into cinnamic acid; cinnamic acid is then decarboxylated to form benzoic acid and finally undergoes 2-hydroxylation to generate SA [18]. Mutation of PAL genes in Arabidopsis results in 50% decrease in pathogen-induced SA accumulation, suggesting that the PAL pathway indeed contributes to SA biosynthesis [19]. Another biosynthetic pathway is through isochorismate synthase that catalyses the conversion of chorismate into isochorismate. [20]. In this pathway, SA is generated in chloroplasts from chorismate by the synthesis of two isochorismate synthases, ICS1 and ICS2 [21]. Analysis of SA-deficient mutants, sid2, revealed that loss of ICS1 suppresses the pathogen-induced SA accumulation [20], whereas loss of both ICSes results in further reduction of SA concentration [21].
It has been well established that SA is a key signal regulating local and systemic plant defence responses against pathogens [20,22]. In this signalling pathway, SA-binding proteins, such as SABP2, NPR3, NPR4 and NPR1 with high affinity for SA, are considered to be SA receptors that induce the expression of pathogenesis-related (PR) proteins and trigger systemic acquired resistance (SAR) [23]. Moreover, SA has shown important roles in mediating plant responses to abiotic stresses [24], including drought [25], chilling [26], osmotic stress [27], thermogenesis [28] and heavy metal toxicity [29]. Recently, the beneficial roles of SA in enhancing plant Cd tolerance, which has been reported in a wide range of plant species, have drawn much attention. However, the negative effect of SA was also noted in castor bean seedlings in which pretreatment with SA aggravated Cd damage [30]. The effects of SA on Cd-stressed plants depend on many aspects, including the application mode, the concentrations of Cd and SA and endogenous SA in the tested plants as well as the different species and developmental stages of the plants. This review covered recent studies on (1) modes of SA treatment, (2) the diverse physiological roles of SA in reducing Cd toxicity, and (3) future prospects for research concerning SA in Cd-stressed plants.
2. Salicylic Acid (SA) Treatment Methods
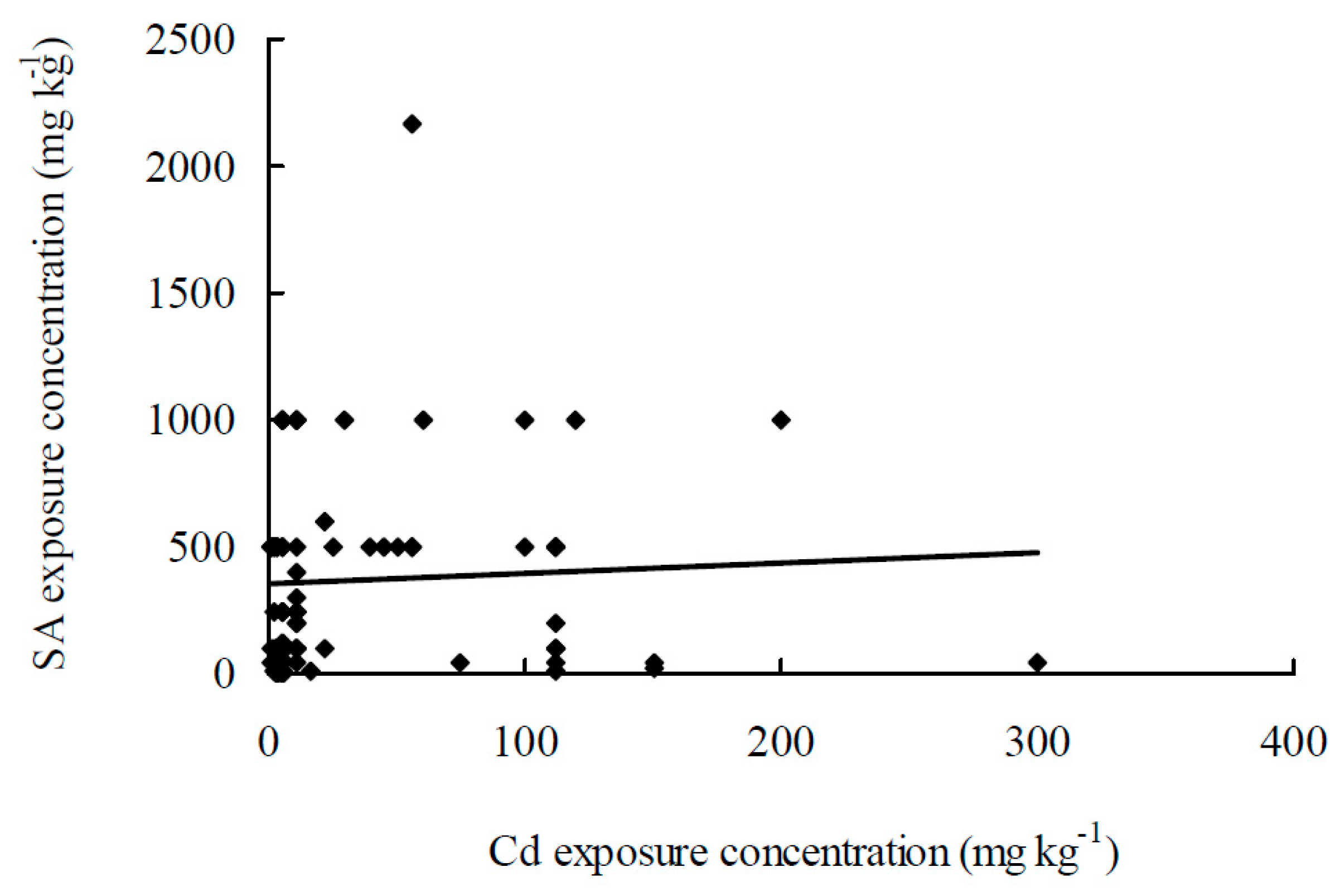
Table 1 summarizes the recent findings in the literature on the roles of SA in Cd-stressed plants. Except for one study with castor bean and three experiments with SA-deficient mutants, most of the studies show that SA alleviates the effects of Cd damage. However, very few of these literatures are actually relevant for coming to a solution for Cd toxicity. Exogenous applications of SA are mainly performed via three modes: spraying SA onto leaves, presoaking seeds with SA solution, and hydroponic treatment of roots to SA. There were 46 international publications involved in 100 pairs of exogenous SA and Cd treatments and 5 publications involved in endogenous SA treatments. Figure 1 represented SA exposure concentrations plotted against Cd exposure concentrations. Among these studies, one in which Cd and SA treatments with 560 mg L−1 and 3000 µM, respectively, was excluded because the concentrations used were too high and may be misleading. Taken together, the following information can be inferred: (1) Concentrations used in Cd treatments range from 0.56 to 300 mg L−1 (mg kg−1) with a mean of 31.6 mg L−1 (mg kg−1) and a variable coefficient of 160%; (2) concentrations of SA spray treatments range from 10 to 2170 µM with a mean of 585 µM and a variable coefficient of 104%; (3) concentrations of SA used in seed presoaking treatments range from 100 to 1000 µM with a mean of 464 µM and a variable coefficient of 70%; and (4) concentrations of SA used in hydroponic exposure treatments range from 1 to 500 µM with a mean of 148 µM and a variable coefficient of 117%.
From these studies, it was determined that the levels of SA in spray or pre-soaking treatments are generally higher than those in hydroponic treatments, suggesting that the regulation of SA levels in plants through root uptake is more efficient than that in the other two modes of application. Furthermore, each mode has high variable coefficients of concentrations either for Cd or for SA treatments. It is interesting to note that partial application of SA to different organs (seeds, leaves or roots) helps plants to establish a systemic defence against Cd toxicity.
2.1. SA Spray
Seven studies reported that SA spray alleviated Cd damage to plants. The species included potato, peppermint, oilseed rape, melon, soybean, radish, and Indian mustard. For instance, foliar application of 600 μM SA onto potato over 10 consecutive days significantly decreased the concentrations of reactive oxygen species (ROS) in leaves and stimulated the antioxidant enzyme mechanism and the related genes (StSABP2, StSOD and StAPX) under 200 μM Cd stress [31]. However, the concentrations used in SA application in the literature ranged from 10 to 2170 µM, and the frequency of applications was completely different. Such large gaps make it difficult to provide guidance for implementing SA spray treatments.
2.2. Presoaking of Seed with SA
There are more studies (19 articles) conducted on seed presoaking than on SA spray treatments. The presoaking periods ranged from 3 to 24 h, and the presoaking levels ranged from 100 to 1000 µM. Moreover, the tested plants were mainly food crops, i.e., rice, maize, wheat, barley, and bean, and industrial crops, i.e., bluegrass, flax, alfalfa, hemp, and castor bean. Similarly, most of the presoaking reports suggested that pretreatment with SA alleviated the subsequent Cd-induced damage to plant growth and the antioxidant system. Notably, presoaking method is more practical for agricultural and/or economic use than spray treatment.
2.3. Hydroponic Application
Indeed, hydroponic treatment of SA is less practicable because Cd is not a problem in hydrographic environment. However, this type of studies is needed since SA or Cd treatment can be controlled more precisely and effectively, which is beneficial to the phytotoxicology research. There were 20 articles conducted on this application method. The SA concentrations ranged from 10 to 500 µM, and the treatment period ranged from 3 to 72 h. Furthermore, the timeline of SA application was different with pretreatment, simultaneous treatment and post-treatment methods. Again, hydroponic treatment of plant roots to SA mitigated Cd toxicity systemically in plants. However, more research work is needed on the transport and signalling pathways of SA through different organs under Cd stress.
2.4. SA Mutants
In the tested experiments, the exposure times of Cd and SA were relatively short, and the levels of Cd and SA were relatively high. Hence, plants received the SA signal rapidly as an instant response to the “acute” Cd stress. Furthermore, information in the literature differs in terms of plant species, treatments and concentrations. All of the above uncertain issues may lead to unpredictable results. For example, when the treatment concentration of SA exceeded the appropriate range, oxidative toxicity occurred [77]. Recently, studies (5 articles) focused on the role of endogenous SA in regulating Cd tolerance by comparing SA-deficient or SA-accumulating mutants with wild-type plants. Surprisingly, although sid2 and NahG are both SA-deficient Arabidopsis lines, these two mutants exhibited contrasting responses. In sid2 mutants, the mutation in the gene encoding isochorismate synthase (ICS) aggravated Cd toxicity compared to the wild-type plants [72]. In contrast, the NahG transgenic lines manifested higher Cd resistance than the wild-type plants [74,75].
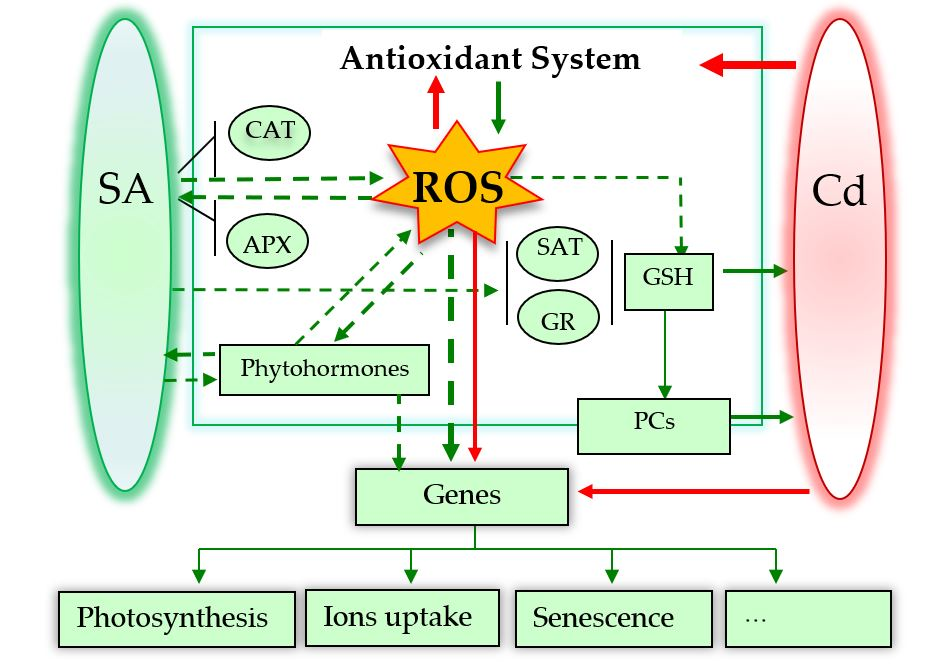
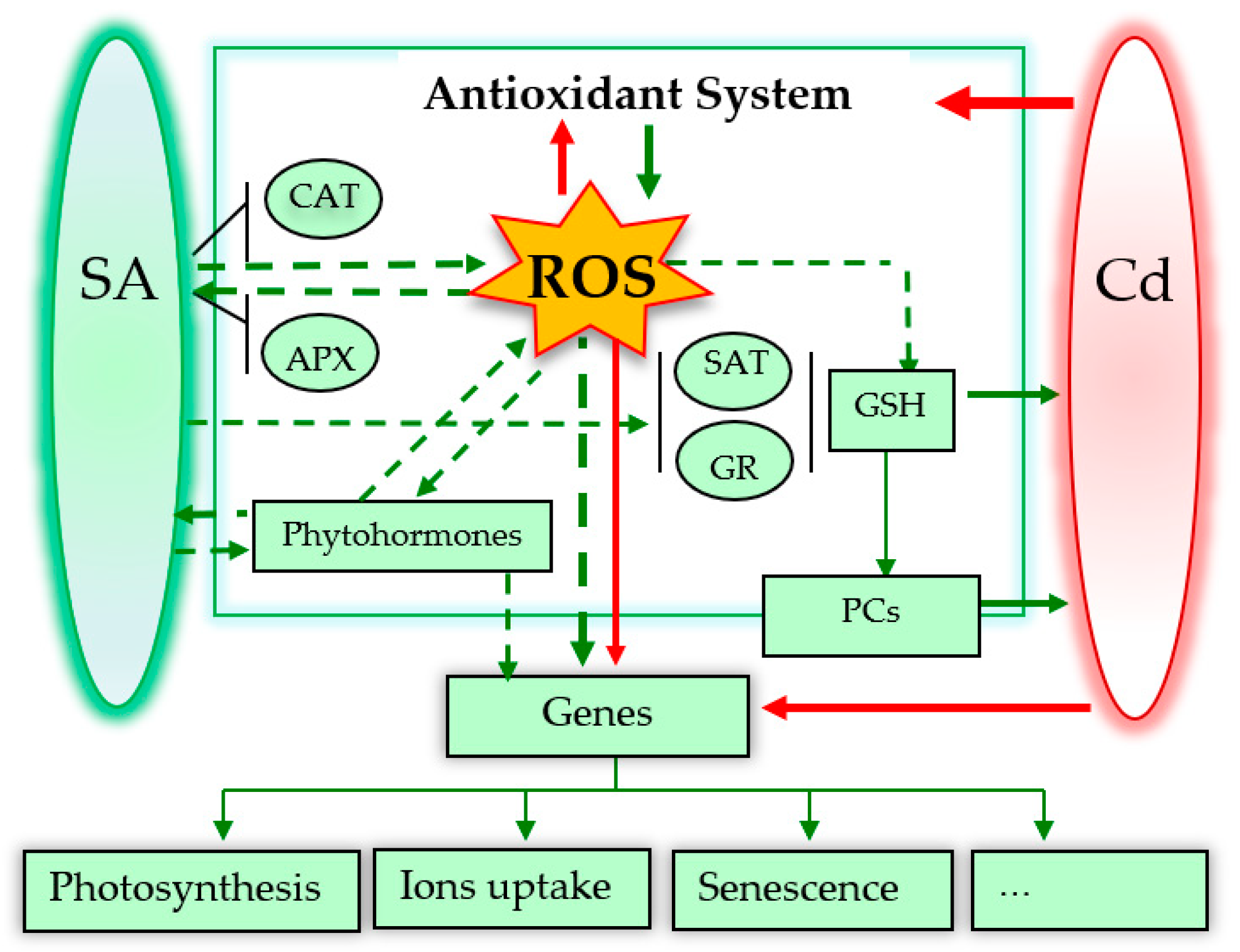
3. Possible Roles of SA in Alleviating Cadmium (Cd) Toxicity
Taking a comprehensive view of SA roles in response to Cd toxicity, this review focuses on the recent advances in the physiological and molecular mechanisms of the following aspects: plant growth, Cd immobilization and distribution, element assimilation, photosynthesis, ROS and the antioxidant defence system, glutathione, and senescence.
3.1. Plant Growth
Cadmium exposure inhibits plant growth [78]. It also causes morphological changes in leaves and roots, such as leaf chlorosis and lignification of cell walls in root tissues [79]. The Cd-induced growth inhibition is mainly due to reduction of net photosynthetic rate [80], inactivation of enzymes involved in CO2 fixation [81], disturbance of element metabolism [82], and induction of lipid peroxidation [83].
As a multifaceted phytohormone, SA mediates physiological and biochemical processes during all plant developmental stages, including seed germination, vegetative growth, seed production, and senescence [84,85]. Arabidopsis mutants with constitutively high levels of SA, such as cpr5 [86], acd6−1 [87], agd2 [88] and pi4kIIIβ1β2 [89] exhibit dwarf phenotype. In contrast, the SA-depleted Arabidopsis NahG transgenic plants have a longer vegetative stage and higher growth rate compared with wild-type plants [18]. The biochemical events involved in the regulation of cell division and growth by SA still need to be clarified [90], which may be crosslinked with AUX, ROS, Ca2+ and mitogen-activated protein kinase (MAPK) pathways [84].
In the presence of Cd, exogenous treatment with SA showed a positive effect on the growth of various plant species, such as flax [47], bluegrass [41], radish [36], and rice [67,68]. Soybean seedlings treated with 6 mg kg−1 Cd for 72 h showed retarded growth symptoms in roots, stems and leaves [91]. SA applied simultaneously at the levels of 1 μM, 10 μM and 100 μM significantly reversed these inhibitory effects. In barley, Cd exposure reduced the dry weight of shoots and roots by approximately 35%, whereas pretreatment with SA resulted in significant recovery of all the growth parameters [54].
Exogenous treatment with SA has a dose-dependent effect on plant growth, as observed in the Arabidopsis mutants with unnecessary or deficient SA levels. Presoaking treatment with 10 to 500 μM SA increased the germination of Kentucky bluegrass seeds, while the germination sharply declined under 1000 to 5000 μM SA treatments [41]. The Cd-induced inhibitory effects on ryegrass growth were significantly alleviated by low SA concentrations, but no effects were found for the high SA concentration [69]. Some plants, such as hemp [50], are vulnerable to high levels of SA but still benefit from SA treatment when they suffer from Cd toxicity. In castor bean seedlings, SA treatment significantly worsened plant growth in both the presence and absence of Cd [30].
Furthermore, inconsistent conclusions were drawn in experiments with Arabidopsis transgenic plants or mutants. The Cd-inhibited growth in Arabidopsis was aggravated by unnecessary SA in snc1 mutants and mitigated by the depletion of SA in nahG transgenic lines [74]. It seems that endogenous SA negatively regulates Arabidopsis tolerance to Cd. However, the sid2 mutants with SA-deficient phenotype showed a Cd-sensitive phenotype that manifested as having accentuated Cd-induced growth inhibition [72].
3.2. Cd Immobilization in the Cell Wall
The plant cell wall, as a complex composed of sugars, proteins and phenols, is the first barrier against Cd toxicity and limits Cd translocation into the cytosol [78]. The hemicellulose and pectin in the cell wall are major components for Cd deposition due to their negative charges [92]. Findings between hyperaccumulating and non-hyperaccumulating ecotypes of Sedm alfredii show that roots with higher cell-wall polysaccharides and activity of pectin methylesterase are more impermeable to Cd [93]. Using energy-dispersive X-ray microanalysis, Cd binding to the cell wall was directly visualized in Thlaspi caerulescens [14]. In addition, long-term Cd treatments may interfere synthesis and composition of cell wall, such as inducing lignification, which in turn impact Cd sequestration in a more complicated way [94]. Whereas for SA, the signalling pathway is also involved in regulation of cell wall components. Genetic research showed that pmr4 mutants are resistant to pathogens due to mutation of a callose synthase, while blocking the SA signaling pathway restore the susceptibility, suggesting callose or callose synthase negatively regulates the SA pathway [95]. SA application altered the lignin and hemicellulose composition of cell wall in Brachypodium distachyon by increase in caffeic acid, ferulic acid and p-coumaric acid content [96].
Therefore, it is hypothesized that pretreatment with SA may increase Cd accumulation in the cell wall and prevent Cd translocation into other cell organelles. Until now, only a few experiments with peanut and ryegrass have shown that SA treatment reduced Cd influx by rearrangement of the cell wall composition [60,62,69]. It has been reported that either SA pretreatment or Cd treatment alone strengthened the root cell wall in rice [97]. However, it was also found that SA treatment with Cd application failed to synergistically affect the cell wall construction or the activity of enzymes related to lignin synthesis, such as polyphenol oxidase (PPO), peroxidase (POD) and PAL. This might be because the strong toxicity of Cd maximized the process of lignification, which concealed the SA signalling role in cell wall construction.
3.3. Cd Uptake and Translocation
The effect of SA on the construction of the cell wall is closely related to Cd uptake and translocation, but the results are still controversial. Some studies have indicated that the treatment of the plant with SA could decrease Cd assimilation and root-to-shoot translocation. For instance, SA application substantially reduced Cd uptake and ameliorated Cd-induced growth inhibition in radish roots [36]. Pretreatment of flax with SA significantly decreased the Cd levels in different organs of the plant [47]. This was also reported in other plant species, i.e., Kentucky bluegrass [41], ryegrass [62,69], alfalfa [65], oilseed rape [33], and wheat [98]. The suppression of Cd uptake and translocation might be due to SA-induced reduction in the uptake, inhibition or activation of element translocators that dislocate Cd into vacuoles [50,91].
However, the role of SA signalling in preventing Cd transport between plant organs is not always physiological. SA and Cd applied simultaneously increased Cd assimilation in soybean [91]. In SA-pretreated barley, tissue Cd contents were unaltered both in vacuoplasts and mesophyll cells at the organ and the whole-plant level [54]. An interesting examination of Cd translocation was conducted using a split-root system with rice [67]. In this system, an appreciable amount of Cd was transferred from Cd-treated root parts to non-Cd-treated root parts. However, pretreatment with 10 μM SA of the whole rice roots did not restrict but promoted this transport process. As for SA mutants, in sid2 and NahG, SA deficiency did not influence Cd assimilation either in shoots or in roots, indicating that SA might not mediate an avoidance mechanism in plants [72,75].
3.4. Element Uptake
Cadmium uptake by plants involves the competition of nutrients by using the same membrane transporters, hence interfering with ionic homeostasis [54,78]. Expression studies with the Fe transporter cloned from Arabidopsis, IRT1, facilitate the Cd influx across the root-cell plasma membrane [99]. Expression of the wheat cDNA LCT1 in S. cerevisiae mediates both Ca and Cd transport into the cytosol of plant cells [100].
The beneficial role of SA could be attributed to its maintenance of the optimal nutrition status of plants. For instance, the Cd-induced disturbance in ion uptake, including K, Fe, Ca, Mg, Mn, and Zn uptake, was alleviated by SA treatment, in flax [47], ryegrass [69], rice [101], peanut [60], Kentucky bluegrass [41] and oilseed rape [33]. This could be explained by the alterations of plasma membrane properties by SA, increasing the activity of H+-ATPase [102], which facilitates the assimilation of nutrients under Cd toxicity.
The effect of SA on element uptake is especially relevant for K, which plays a key role in regulating H+-ATPase in the plasma membrane. As early as 1981, the inhibitory function of SA was observed for K absorption in oat roots [103]. However, under Cd stress, a positive correlation between SA and K was noted in soybean seedlings [91] and the SA mutant, sid2 [72], and SA further interfered with the uptake of Fe and Mg in roots and shoots. The interaction between SA and Cd is also involved in the assimilation of S, the key element of sulfhydryl groups that chelate Cd ions into less vulnerable organelles in plant cells. In the absence of Cd, SA treatment increased the S content of barley roots [54]. In contrast, the S levels were much higher in the sid2 mutant leaves than in the wild-type plants under Cd-free conditions [72].
3.5. Photosynthesis
The mineral nutrient stress induced by Cd results in severe alterations in photosynthesis in terms of chloroplast structure, chlorophyll concentration, and activities of carboxylating enzymes [51]. Meanwhile, SA may act as an important photosynthesis regulator by the influence of RuBisCO activity, contribution to light acclimation and redox homeostasis, and the function of the stomatal switch [84].
Pretreatment with SA prevented Cd-induced chlorophyll destruction in maize [51], soybean [64], oilseed rape [33] and flax [47]. Conversely, depletion of SA further lowered the chlorophyll concentrations in Cd-treated sid2 mutants [72]. The Cd-inhibited activity of RuBisCo and carbonic anhydrase was recovered by exposure to 0.1 mM SA in peppermint [32]. Moreover, SA application increased the carotenoid synthesis of soybean seedlings, whereas it decreased the flavonoid content under Cd stress [35]. The alleviating role of SA could be due to the restored K contents in leaves, strengthening stomatal closure [72], which is synchronized with the net photosynthetic rate, transpiration, capability for CO2 fixation and inhibition of the activities of chlorophyll-degrading enzymes [35,50,62].
In addition, non-stomatal factors signalled by SA also play roles in the maintenance of photosynthetic capacity under Cd stress. The application of SA to Cd-treated barley leaf slices significantly slowed the decreasing trends in the photosynthetic yield of photosystem II (PSII) [54]. Spraying 0.1 mM SA onto melon leaves induced an increase in Fv/Fm in Cd-stressed plants, indicating that SA improved PSII efficiency [34]. Similarly, treatment with SA in rice prevented the unnecessary energy transference from PSII to PSI induced by Cd toxicity [55]. In this study, SA increased the cyclic electron transport around PSI in thylakoid membranes and protected the Mn-cluster of the oxygen-evolving complex from Cd damage.
The photosynthetic response to SA is both dose- and species-dependent. Pretreatment with 250 and 500 μM SA in castor bean leaves failed to affect chlorophyll levels but aggravated the negative effect on photosynthesis induced by Cd [30], which might have been associated with an increase in stomatal limitation. The reduced SA level in nahG plants resulted in the maintenance of photosynthetic efficiency by low photoinhibition under Cd stress [71]. The response to endogenous SA in Cd tolerance might be associated with the regulation of photosynthetic electron transport, starch degradation and PSII structures at the transcriptional level.
3.6. Reactive Oxygen Species (ROS) and Antioxidant Defence System
The production of ROS in plant cell is an unavoidable consequence of oxygen metabolism, especially during respiration and photosynthesis [104]. The major mode of Cd toxicity in plants is to induce ROS production and results in oxidative injuries in plants [30]. Although Cd does not participate directly in cellular redox reactions (such as Fenton reaction), it can indirectly elevate ROS accumulation in the cellular environment by the disturbance of electron transport, destroying the structure of antioxidant enzymes and interfering with antioxidant molecule synthesis. Cd indirectly modulates the activity of the plasma membrane NADPH oxidase, increasing the formation of O2•− and H2O2, which has been found in tobacco [105], rice [106], and lupine roots [107]. Meanwhile, the enhanced demand for glutathione (GSH) for Cd chelation causes rapid loss in antioxidative defence [16].
It is well known that SA signals plant resistance through modulation of ROS metabolism, especially H2O2. The mode of SA action involves binding directly to CAT and ascorbate peroxidase (APX), two major H2O2-scavenging enzymes [108], inhibiting their activities in plants. This finding was also confirmed in sid2 mutants, SA-deficient Arabidopsis plants, whose leaf CAT activity is higher than that in wild-type plants [72]. Using the DAB staining method, the SA-induced accumulation of H2O2 was visualized in rice leaves [66]. SA may regulate H2O2 accumulation through a self-amplifying feedback loop in Arabidopsis. In this process, SA acts as an electron donor of CAT that effectively slows down the peroxidation cycle, hence sharply decreasing the efficiency of H2O2 elimination [108]. Another important mechanism may occur in mitochondria. SA blocks electron flow from the substrate dehydrogenises to the ubiquinone pool and triggers H2O2 generation [109].
Both Cd stress and SA accumulation can elevate H2O2 production in cells. Therefore, SA treatment may aggravate the oxidative stress induced by Cd toxicity. Surprisingly, a large body of studies found that SA treatment alleviates Cd-induced oxidative stress in plants. In fact, H2O2 has a dual role in plant biology as both a toxic byproduct and a key regulator against many abiotic stresses in plants [110]. The increase in H2O2 status stimulated by SA pretreatment acts as a crucial message to “set up” the antioxidant system and then induces plants to resist subsequent Cd stress. The beneficial effects of SA were found in most of the studies in which SA treatment was performed, either by spraying, presoaking, or by hydroponic incubation, in advance of the application of Cd stress (Table 1). For example, pretreatment with 10 µM SA for 72 h initially caused H2O2 accumulation in vitro in rice roots. Correspondingly, the levels of GSH, non-protein thiols (NPT), and ascorbic acid (AsA) and the activities of CAT, superoxide dismutase (SOD) and POD were elevated compared to those of non-SA treated roots during the subsequent Cd exposure period [67,68]. The expression level of selected genes (StSABP2, StSOD and StAPX) was enhanced in SA-treated potato plants under Cd stress [31]. Pretreatment with SA improved the antioxidant capacity and tolerance of Cd-induced oxidative stress in rice leaves, which was similar to the effects of H2O2 pretreatment [66]. Moreover, the role of SA in mitigating Cd damage was also confirmed in many other species, such as flax [39], mustard [37], oilseed rape [33], ryegrass [62,69], melon [34], etc.
Although a number of reports on the benefits of SA signalling exist in the literature, some questions have not yet been fully answered. SA treatment alleviated Cd toxicity in barley but lowered the activities of antioxidant enzymes [54]. SA could act as an oxidant at high levels due to its capability to increase H2O2 generation and oxidation of proteins in plant leaves [77]. In contrast, it was reported that SA may act as a direct scavenger to reduce excessive H2O2 injury in the pea during Cd stress [111]. In addition, SA may directly signal the expression of defence-related enzymes, such as haem oxygenase−1 (HO−1), to alleviate Cd-triggered oxidative toxicity by reestablishing redox homeostasis [64]. Moreover, the crosstalk and response pathways between SA and other phytohormones should also be noted. SA combined with NO treatment synergistically counteracted Cd-induced oxidative damage in peanut and ryegrass [60,62]. Under Cd stress, pretreatment of wheat seedlings with SA significantly promoted the synthesis of dehydrins, the abscisic acid (ABA)-signalled proteins that can neutralize and bind unnecessary H2O2 [98].
The results of the SA mutants studies are still contradictory. snc1 mutants with high intrinsic levels of SA possess high POD activity, which can generate a large amount of ROS and manifest the Cd-sensitive phenotype [74]. In contrast, SA deficiency in NahG transgenic lines mitigates the oxidative stress induced by Cd toxicity [74,75]. In contrast, sid2 mutants with an SA-deficient phenotype aggravated Cd-induced oxidative damage compared with their wild-type plants.
3.7. Glutathione and Chelation
Glutathione (γ-Glu-Cys-Gly, GSH) is one of the most important reducing equivalents in plants, protecting plants against Cd-induced oxidative damage. Furthermore, it is also a key molecular compound or a basic component of phytochelatins (PCs) involved in Cd chelation and thereby confines Cd to less sensitive organelles, such as vacuoles [15,112].
A series of genetic reports using single mutants or transformants have shown direct evidence that endogenous SA signalling is linked to GSH biosynthesis. The catalase-deficient Arabidopsis mutant, cat 2, induces SA levels and a wide range of SA-dependent responses alongside the upregulation of GSH [113]. In contrast, attenuation of GSH levels was correlated with decreased SA contents in cat2 atrbohF compared with cat2 [114]. It has been shown that the GSH concentrations were much lower in Cd-stressed sid2 leaves than in wild-type plants [72]. Low GSH levels were also found in other SA-deficient mutants, such as npr1−1 and mpk4−1 [115,116]. Furthermore, the exogenous application of SA has been shown to enhance S assimilation in barley roots [54] and elevate GSH content in some plant species, such as peppermint [32], flax [45], peanut [60] and rice [68].
The regulatory role of SA in GSH biosynthesis may be related to serine acetyltransferase (SAT) transcription, the precursor gene that catalyses cysteine formation. Increased free SA levels, both by genetics and by exogenous application, lead to an increased specific activity of SAT and GSH in Arabidopsis [117]. Under Cd stress, although depletion of SA in sid2 mutants significantly enhanced the uptake of S, a key element for GSH construction, down-regulated transcription of SAT-c and SAT-p in sid2 blocked the process of GSH biosynthesis and resulted in lower GSH levels compared with the wild-type plants [72]. Glutathione synthetase (GSHS) is a rate-limiting enzyme that catalyses the second step of GSH synthesis in plants. The expression of the Cd-induced LcGSHS transcript, a GSHS gene isolated from L. chinense, is controlled by the endogenous SA-dependent pathway and results in greater GSH accumulation and Cd tolerance in transgenic Arabidopsis [73]. Furthermore, SA has also been shown to mediate the synthesis of glutathione reductase (GR1), the pivotal enzyme for regenerating and maintaining GSH in the reduced state [118]. During Cd exposure, SA deficiency significantly decreased GR1 transcription in sid2 mutants and resulted in lower GSH levels and GR activity compared with the wild-type plants [72]. However, another SA-deficient transgenic line, NahG, manifested high GSH accumulation and a high GSH/GSSG ratio in the presence of Cd [74,75]. These inconsistent results suggest that the role of SA in regulating GSH synthesis under Cd stress requires further investigation.
3.8. Senescence
Cd toxicity accelerates the ageing process in plant cells referred to as senescence. The Cd-induced morphological changes associated with senescence were shown in pine roots [16]. Cd exposure accelerated the senescence process of Arabidopsis, as indicated by an increase in SAG12 expression, a typical senescence marker gene [119].
Although senescence is a negative physiological process, it is one of the most important stages that plants undergo to maintain organ homeostasis and to escape from unfavourable conditions. Under biotic stress, SA is well known to induce senescence of infected tissues to build up a physical barrier against the spread of pathogens. In SA-deficient Arabidopsis plants, the transcript of SAG12 was considerably reduced or undetectable [88]. Whether treatment with Cd and SA manifests antagonistic or synergistic effects on plant senescence is still unclear. Generally, Cd distributes unevenly in field environment. Therefore, some parts of plant root may suffer Cd toxicity severely but some parts may not. An interesting hypothesis is that SA might accelerate Cd-induced senescence of stressed root parts, and then benefit the whole plant to elude Cd damage by adjusting the root growth direction towards the non-Cd contaminated environment. To test this hypothesis, a split-root experiment was conducted in which half of the roots were exposed to Cd while the other half were not exposed [67]. They found that Cd treatment caused senescence of the stressed part of the roots and stimulated the growth of the non-stressed part of the roots. However, SA pretreatment had no effect on the senescence of the Cd-exposed roots and did not lower the Cd uptake, which might be because the low level of exogenous SA used was insufficient to trigger senescence. Nonetheless, this hypothesis can be tested by using Arabidopsis mutants, such as snc−1 with constitutively high concentrations of endogenous SA, since this mutant manifests a senescence phenotype and lower uptake of Cd compared to the wild-type plants [74].
4. Future Insights and Conclusions
Research over the past 20 years has strongly indicated that SA is a very promising molecule for the reduction of Cd toxicity in plants. Here, we reviewed reports describing the promoting role of SA in Cd resistance under various treatment methods, including pre-soaking, hydroponic exposure, and spraying. Figure 2 proposed the possible roles of SA in alleviating Cd toxicity to plants. However, there still remains a contradiction between the effects of SA at low and high doses. Furthermore, several unsolved and central questions concerning homeostasis, gene expression, and crosstalk with other phytohormones are still not fully understood.
4.1. SA Homeostasis
It is well known that plants can increase their intracellular concentration of SA to combat various environmental stresses, including Cd toxicity [120]. Exposure of Cd for 7 days increased the SA levels of leaves in young maize seedlings [121]. Compared with SA-deficient mutants, Cd stress significantly increased the SA level in wild-type Arabidopsis, indicating that Cd-induced SA synthesis occurs through the NahG- or SID2- pathways [72,74,75]. Exogenous SA treatment highlights the expression of OsWRKY45 and increases the endogenous concentrations of SA in Cd-stressed rice and Lemna minor plants [57,66]. Under biotic stress, the mechanism of SA transport in plant cells has been illuminated. During pathogen attack, the hydroxyl group of SA is conjugated by glucose, resulting in formation of the SA glucoside (SAG). Then, SAG is actively transported from the cytosol into the vacuole, where it functions as an inactive storage form that can release free SA [122]. However, the balance of SA homeostasis in vivo under Cd stress still needs to be further monitored.
4.2. SA-Related Gene Expression
The Cd toxicity and SA signal may share the same origins in regard to gene expression. Characterization and identification of the SA receptor during Cd stress are highly anticipated.
4.2.1. Nonexpressor of Pathogenesis-Related (NPR) Protein
Nonexpressor of PR (NPR1 and NPR3/4), the canonical signal transducer of SA, regulates many gene expressions in seed germination, flowering, and senescence processes [123]. It has been uncovered that both NPR1 and NPR3/4 are SA receptors in regulating SA-mediated plant immunity [124]. Further genetic analysis indicated that NPR1 is a transcriptional activator while NPR3/4 are repressors functioning independent of NPR1 [125]. However, few studies have focused on the issue of whether SA-induced NPR is involved in Cd tolerance in plants. Surprisingly, it has been found that an increase in plant biomass was coupled with SA accumulation in npr1−1 (a SA-deficient mutant) after 12-h-Cd exposure, suggesting that the regulation of Cd tolerance is not related to the NPR1 signalling pathway [74].
4.2.2. Mitogen-Activated Protein Kinase (MAPK)
Another important example of SA-regulated gene expression involves mitogen-activated protein kinase cascades (MAPK), which acts as a negative regulator in plant growth and development in response to endogenous and environmental cues [126]. SA accumulation in mpk4 mutants result in a severely dwarf phenotype [127]. Cd exposure quickly enhanced the kinase activity of MPK6, while the mpk6 mutation enhanced Cd tolerance by alleviating oxidative stress [128]. However, until now, no studies have been conducted to evaluate the role of SA in signalling Cd resistance through MAPK regulation.
4.2.3. ATP-Binding Cassette (ABC) Transporters
ATP-binding cassette (ABC) transporters belong to a large family that utilizes the energy of ATP binding and hydrolysis to transport elements across cellular membranes. In particular, they are involved in sequestering Cd or PC-Cd complexes into vacuoles to alleviate Cd toxicity [129]. An ABC transporter from soybean was identified that was strongly and rapidly induced by SA treatment [130]. Therefore, it is of interest to explore whether ABC transporters are involved in SA signalling and SA-induced Cd tolerance.
4.3. Crosstalk with Other Phytohormones
In addition to crosstalk with ROS transduction, the coordination between signalling of SA and other phytohormones is also an important aspect to consider [131]. SA pretreatment mitigated the Cd-induced disturbance in the levels of indoleacetic acid, cytokinins and ABA in wheat seedlings [98], and alleviated Cd toxicity in barley root tips by inhibiting auxin-mediated ROS generation [61]. It was also reported that SA combined with NO counteracted the negative effects of Cd on ryegrass plants [62]. However, the direct or indirect influence of SA signalling on the balance of plant hormones needs to be determined.
In conclusion, investigations of SA signalling roles shed new light on the approaches to enhancing Cd tolerance with phytohormones. In addition to classical methods of exogenous application, these studies can now be complemented by the creation of a new generation of SA-excessive or SA-deficient mutants. Furthermore, with the characteristics of low cost and high efficiency, SA application shows promising use in helping plants defend against Cd toxicity. Seed presoaking and spraying with SA are pragmatic approaches for this purpose. In the meantime, novel roles of SA in Cd toxicity will likely continue to be unveiled.
Funding
Supported by Department of Science and Technology of Zhejiang Province, China (2010C33158, 2015C03020), Fundamental Research Funds for the Central Universities (2019FZJD007), State Key Laboratory Breeding Base for Zhejiang Sustainable Pest and Disease Control (2010DS700124-ZZ1802), Natural Science Foundation of China (No. 41001184), and National Key R&D Program of China (2018YFD0800202).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AsA | ascorbic acid |
| ABA | abscisic acid |
| ABC transporters | ATP-binding cassette transporters |
| APX | ascorbate peroxidase; |
| Cd | cadmium; |
| CAT | catalase; |
| GR | glutathione reductase |
| GSH | glutathione |
| GSHS | glutathione synthetase |
| HO−1 | haem oxygenase−1 |
| MAPK | mitogen-activated protein kinase |
| NPR1 | Nonexpressor of PR1 |
| NPT | non-protein thiols |
| PAL | phenylalanine ammonia-lyase |
| PCs | phytochelatins |
| POD | peroxidase |
| PPO | polyphenol oxidase |
| PSII | photosystem II |
| ROS | reactive oxygen species |
| SA | salicylic acid |
| SAT | serine acetyltransferase |
| SOD | superoxide dismutase |
References
- Vig, K.; Megharaj, M.; Sethunathan, N.; Naidu, R. Bioavailability and toxicity of cadmium to microorganisms and their activities in soil: A review. Adv. Environ. Res. 2003, 8, 121–135. [Google Scholar] [CrossRef]
- Gong, B.; Nie, W.; Yan, Y.; Gao, Z.; Shi, Q. Unravelling cadmium toxicity and nitric oxide induced tolerance in Cucumis sativus: Insight into regulatory mechanisms using proteomics. J. Hazard. Mater. 2017, 336, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, Y.; Meng, A.; Li, Q.; Cheng, H. Experimental and thermodynamic investigation on transfer of cadmium influenced by sulfur and chlorine during municipal solid waste (MSW) incineration. J. Hazard. Mater. 2008, 153, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Ma, Y.; Zhang, S.; Wei, D.; Zhu, Y. An inventory of trace element inputs to agricultural soils in China. J. Environ. Manag. 2009, 90, 2524–2530. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.L. Cadmium and phosphorous fertilizers: The issues and the science. Procedia Eng. 2014, 83, 52–59. [Google Scholar] [CrossRef]
- Zhou, J.; Wan, H.; He, J.; Lyu, D.; Li, H. Integration of cadmium accumulation, subcellular distribution, and physiological responses to understand cadmium tolerance in apple rootstocks. Front. Plant Sci. 2017, 8, 966. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S.; Aarts, M.G.M.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef]
- Daud, M.K.; Sun, Y.; Dawood, M.; Hayat, Y.; Variath, M.T.; Wu, Y.X.; Mishkat, U.; Najeeb, U.; Zhu, S. Cadmium-induced functional and ultrastructural alterations in roots of two transgenic cotton cultivars. J. Hazard. Mater. 2009, 161, 463–473. [Google Scholar] [CrossRef]
- Song, Y.; Jin, L.; Wang, X. Cadmium absorption and transportation pathways in plants. Int. J. Phytoremediat. 2017, 19, 133–141. [Google Scholar] [CrossRef]
- Lysenko, E.A.; Klaus, A.A.; Pshybytko, N.L.; Kusnetsov, V.V. Cadmium accumulation in chloroplasts and its impact on chloroplastic processes in barley and maize. Photosynth. Res. 2015, 125, 291–303. [Google Scholar] [CrossRef]
- Hasan, S.A.; Hayat, S.; Ahmad, A. Screening of tomato (Lycopersicon esculentum) cultivars against cadmium through shotgun approach. J. Plant Interact. 2009, 3, 187–201. [Google Scholar] [CrossRef]
- Bertoli, A.C.; Cannata, M.G.; Carvalho, R.; Bastos, A.R.R.; Freitas, M.P.; Dos Santos Augusto, A. Lycopersicon esculentum submitted to Cd-stressful conditions in nutrition solution: Nutrient contents and translocation. Ecotoxicol. Environ. Saf. 2012, 86, 176–181. [Google Scholar] [CrossRef]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Palme, K.J.; Jansen, M.A.K. Stress-induced morphogenic responses: Growing out of trouble? Trends Plant Sci. 2007, 12, 98–105. [Google Scholar] [CrossRef]
- Wójcik, M.; D’Haen, V.J.; Tukiendorf, A. Cadmium tolerance in Thlaspi caerulescens II. localization of cadmium in Thlaspi caerulescens. Environ. Exp. Bot. 2005, 53, 163–171. [Google Scholar] [CrossRef]
- Seth, C.S.; Remans, T.; Keunen, E.; Jozefczak, M.; Gielen, H.; Opdenakker, K.; Weyens, N.; Vangronsveld, J.; Cuypers, A. Phytoextraction of toxic metals: A central role for glutathione. Plant Cell Environ. 2012, 35, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Schützendübel, A.; Schwanz, P.; Teichmann, T.; Gross, K.; Langenfeld-Heyser, R.; Godbold, D.L.; Polle, A. Cadmium-induced changes in antioxidative systems, hydrogen peroxide content, and differentiation in scots pine roots. Plant Physiol. 2001, 127, 887–898. [Google Scholar] [CrossRef]
- Raskin, I. Role of salicylic acid in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Abreu, M.E.; Munné-Bosch, S. Salicylic acid deficiency in NahG transgenic lines and sid2 mutants increases seed yield in the annual plant Arabidopsis thaliana. J. Exp. Bot. 2009, 60, 1261–1271. [Google Scholar] [CrossRef]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef]
- Wildermuth, M.C.; Dewdney, J.; Wu, G.; Ausubel, F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defense. Nature 2001, 414, 562–565. [Google Scholar] [CrossRef]
- Garcion, C.; Lohmann, A.; Lamodiere, E.; Catinot, J.; Buchala, A.; Doermann, P.; Metraux, J.P. Characterization and biological function of the ISOCHORISMATE SYNTHASE2 gene of Arabidopsis. Plant Physiol. 2008, 147, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, D.A.; Shah, J.; Klessig, D.F. Salicylic acid and disease resistance in plants. Crit. Rev. Plant Sci. 1997, 18, 547–575. [Google Scholar] [CrossRef]
- Kumar, D. Salicylic acid signaling in disease resistance. Plant Sci. 2014, 228, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.; Kovacs, V.; Szalai, G.; Soos, V.; Ma, X.; Liu, H.; Mei, H.; Janda, T. Salicylic acid and abiotic stress responses in rice. J. Agron. Crop Sci. 2014, 200, 1–11. [Google Scholar] [CrossRef]
- Chini, A.; Grant, J.J.; Seki, M.; Shinozaki, K.; Loake, G.J. Drought tolerance established by enhanced expression of the CC–NBS–LRR gene, ADR1, requires salicylic acid, EDS1 and ABI1. Plant J. 2004, 38, 810–822. [Google Scholar] [CrossRef]
- Janda, T.; Szalai, G.; Tari, I.; Páldi, E. Hydroponic treatment with salicylic acid decreases the effects of chilling injury in maize (Zea mays L.) plants. Planta 1999, 208, 175–180. [Google Scholar] [CrossRef]
- Szalai, G.; Janda, T. Effect of salt stress on the salicylic acid synthesis in young maize (Zea mays L.) Plants. J. Agron. Crop Sci. 2009, 195, 165–171. [Google Scholar] [CrossRef]
- Dat, J.F.; Foyer, C.H.; Scott, I.M. Changes in salicylic acid and antioxidants during induced thermotolerance in mustard seedlings. Plant Physiol. 1998, 118, 1455–1461. [Google Scholar] [CrossRef]
- Horváth, E.; Szalai, G.; Janda, T. Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 2007, 26, 290–300. [Google Scholar] [CrossRef]
- Liu, C.F.; Guo, J.L.; Cui, Y.L.; Lü, T.F.; Zhang, X.H.; Shi, G.R. Effects of cadmium and salicylic acid on growth, spectral reflectance and photosynthesis of castor bean seedlings. Plant Soil 2011, 344, 131–141. [Google Scholar] [CrossRef]
- Li, Q.; Wang, G.; Wang, Y.; Yang, D.; Guan, C.; Ji, J. Foliar application of salicylic acid alleviate the cadmium toxicity by modulation the reactive oxygen species in potato. Ecotoxicol. Environ. Saf. 2019, 172, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Jaleel, H.; Sadiq, Y.; Khan, M.M.A.; Shabbir, A. Response of exogenous salicylic acid on cadmium induced photosynthetic damage, antioxidant metabolism and essential oil production in peppermint. Plant Growth Regul. 2018, 86, 273–286. [Google Scholar] [CrossRef]
- Ali, E.; Maodzeka, A.; Hussain, N.; Shamsi, I.H.; Jiang, L. The alleviation of cadmium toxicity in oilseed rape (Brassica napus) by the application of salicylic acid. Plant Growth Regul. 2015, 75, 641–655. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Yang, S.; Chen, Y. Salicylic acid alleviates cadmium-induced inhibition of growth and photosynthesis through upregulating antioxidant defense system in two melon cultivars (Cucumis melo, L.). Protoplasma 2015, 252, 911–924. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Ma, L.J.; Bu, N.; Li, Y.Y.; Zhang, L.H. Effects of salicylic acid pre-treatment on cadmium and/or UV-B stress in soybean seedlings. Biol. Plant. 2014, 58, 195–199. [Google Scholar] [CrossRef]
- Raza, S.H.; Shafiq, F. Exploring the role of salicylic acid to attenuate cadmium accumulation in radish (Raphanus sativus). Int. J. Agric. Biol. 2013, 15, 547–552. [Google Scholar]
- Ahmad, P.; Nabi, G.; Ashraf, M. Cadmium-induced oxidative damage in mustard (Brassica juncea (L.) czern. & coss.) plants can be alleviated by salicylic acid. S. Afr. J. Bot. 2011, 77, 36–44. [Google Scholar]
- Roychoudhury, A.; Ghosh, S.; Paul, S.; Mazumdar, S.; Das, G.; Das, S. Pre-treatment of seeds with salicylic acid attenuates cadmium chloride-induced oxidative damages in the seedlings of mungbean (Vigna radiata, L. wilczek). Acta Physiol. Plant. 2016, 38, 11. [Google Scholar] [CrossRef]
- Belkadhi, A.; De Haro, A.; Obregon, S.; Chaïbi, W.; Djebali, W. Positive effects of salicylic acid pretreatment on the composition of flax plastidial membrane lipids under cadmium stress. Environ. Sci. Pollut. Res. 2015, 22, 1457–1467. [Google Scholar] [CrossRef]
- Belkadhi, A.; De Haro, A.; Soengas, P.; Obregόn, S.; Cartea, M.E.; Chaïbi, W.; Djebali, W. Salicylic acid increases tolerance to oxidative stress induced by hydrogen peroxide accumulation in leaves of cadmium-exposed flax (Linum usitatissimum L.). J. Plant Interact. 2014, 9, 647–654. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, L.; Mao, P.C.; Jia, Y.Q.; Shi, Y.J. Role of exogenous salicylic acid in alleviating cadmium-induced toxicity in Kentucky bluegrass. Biochem. Syst. Ecol. 2013, 50, 269–276. [Google Scholar] [CrossRef]
- Agami, R.A.; Mohamed, G.F. Exogenous treatment with indole-3-acetic acid and salicylic acid alleviates cadmium toxicity in wheat seedlings. Ecotoxicol. Environ. Saf. 2013, 94, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Szalai, G.; Krantev, A.; Yordanova, R.; Popova, L.P.; Janda, T. Influence of salicylic acid on phytochelatin synthesis in Zea mays during Cd stress. Turk. J. Bot. 2013, 37, 708–714. [Google Scholar]
- Belkadhi, A.; De Haro, A.; Soengas, P.; Obregόn, S.; Cartea, M.E.; Djebali, W.; Chaïbi, W. Salicylic acid improves root antioxidant defense system and total antioxidant capacities of flax subjected to cadmium. OMICS J. Integr. Biol. 2013, 17, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Belkadhi, A.; Hediji, H.; Abbes, Z.; Djebali, W.; Chaibi, W. Influence of salicylic acid pre-treatment on cadmium tolerance and its relationship with non-protein thiol production in flax root. Afr. J. Biotechnol. 2012, 11, 9788–9796. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, H.; Xia, Y.; Wang, G.; Xu, L.; Shen, Z. Exogenous application of salicylic acid alleviates cadmium toxicity and reduces hydrogen peroxide accumulation in root apoplasts of Phaseolus aureus and Vicia sativa. Plant Cell Rep. 2011, 30, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Belkhadi, A.; Hediji, H.; Abbes, Z.; Nouairi, I.; Barhoumi, Z.; Zarrouk, M.; Chaïbi, W.; Djebali, W. Effects of exogenous salicylic acid pre-treatment on cadmium toxicity and leaf lipid content in Linum usitatissimum, L. Ecotoxicol. Environ. Saf. 2010, 73, 1004–1011. [Google Scholar] [CrossRef]
- Moussa, H.R.; Elgamal, S.M. Effect of salicylic acid pretreatment on cadmium toxicity in wheat. Biol. Plant. 2010, 54, 315–320. [Google Scholar] [CrossRef]
- Dražić, G.; Mihailovic, N. Salicylic acid modulates accumulation of Cd in seedlings of Cd-tolerant and Cd-susceptible soybean genotypes. Arch. Biol. Sci. 2009, 61, 431–439. [Google Scholar] [CrossRef]
- Shi, G.R.; Cai, Q.S.; Liu, Q.Q.; Wu, L. Salicylic acid-mediated alleviation of cadmium toxicity in hemp plants in relation to cadmium uptake, photosynthesis, and antioxidant enzymes. Acta Physiol. Plant. 2009, 31, 969–977. [Google Scholar] [CrossRef]
- Krantev, A.; Yordanova, R.; Janda, T.; Szalai, G.; Popova, L. Treatment with salicylic acid decreases the effect of cadmium on photosynthesis in maize plants. J. Plant Physiol. 2008, 165, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Patra, H.K. Effect of salicylic acid potentiates cadmium-induced oxidative damage in oryza sativa, l. leaves. Acta Physiol. Plant. 2007, 29, 567–575. [Google Scholar] [CrossRef]
- Choudhury, S.; Panda, S.K. Role of salicylic acid in regulating cadmium induced oxidative stress in Oryza sativa L. roots. Bulg. J. Plant Physiol. 2004, 30, 95–110. [Google Scholar]
- Metwally, A.; Finkemeier, I.; Georgi, M.; Dietz, K.J. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol. 2003, 132, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Yotsova, E.K.; Dobrikova, A.G.; Stefanov, M.A.; Kouzmanova, M.; Apostolova, E.L. Improvement of the rice photosynthetic apparatus defence under cadmium stress modulated by salicylic acid supply to roots. Theor. Exp. Plant Phys. 2018, 30, 57–70. [Google Scholar] [CrossRef]
- Gu, C.S.; Yang, Y.H.; Shao, Y.F.; Wu, K.W.; Liu, Z.L. The effects of exogenous salicylic acid on alleviating cadmium toxicity in nymphaea tetragona georgi. S. Afr. J. Bot. 2018, 114, 267–271. [Google Scholar] [CrossRef]
- Lu, Q.; Zhang, T.; Zhang, W.; Su, C.; Yang, Y.; Hu, D.; Xu, Q. Alleviation of cadmium toxicity in Lemna minor by exogenous salicylic acid. Ecotoxicol. Environ. Saf. 2018, 147, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Gondor, O.K.; Pál, M.; Darkó, É.; Janda, T.; Szalai, G. Salicylic acid and sodium salicylate alleviate cadmium toxicity to different extents in maize (Zea mays L.). PLoS ONE 2016, 11, e0160157. [Google Scholar] [CrossRef]
- Singh, I.; Shah, K. Evidences for suppression of cadmium induced oxidative stress in presence of sulphosalicylic acid in rice seedlings. Plant Growth Regul. 2015, 76, 99–110. [Google Scholar] [CrossRef]
- Xu, L.L.; Fan, Z.Y.; Dong, Y.J.; Kong, J.; Bai, X.Y. Effects of exogenous salicylic acid and nitric oxide on physiological characteristics of two peanut cultivars under cadmium stress. Biol. Plant. 2015, 59, 171–182. [Google Scholar] [CrossRef]
- Tamás, L.; Mistrík, I.; Alemayehu, A.; Zelinová, V.; Bočová, B.; Huttová, J. Salicylic acid alleviates cadmium-induced stress responses through the inhibition of Cd-induced auxin-mediated reactive oxygen species production in barley root tips. J. Plant Physiol. 2015, 173, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liang, X.; Dong, Y.; Xu, L.; Zhang, X.; Kong, J.; Liu, S. Effects of exogenous salicylic acid and nitric oxide on physiological characteristics of perennial ryegrass under cadmium stress. J. Plant Growth Regul. 2013, 32, 721–731. [Google Scholar] [CrossRef]
- Saidi, I.; Ayouni, M.; Dhieb, A.; Chtourou, Y.; Chaïbi, W.; Djebali, W. Oxidative damages induced by short-term exposure to cadmium in bean plants: Protective role of salicylic acid. S. Afr. J. Bot. 2013, 85, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Noriega, G.; Caggiano, E.; Lecube, M.L.; Santa-Cruz, D.; Batlle, A.; Tomaro, M.; Balestrasse, K.B. The role of salicylic acid in the prevention of oxidative stress elicited by cadmium in soybean plants. Biometals 2012, 25, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Le, L.; Gao, Z.; Wu, H.; Xie, Y.; Shen, W. Haem oxygenase-1 is involved in salicylic acid-induced alleviation of oxidative stress due to cadmium stress in Medicago sativa. J. Exp. Bot. 2012, 63, 5521–5534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, Y.Y.; Chen, C.Y.; Huang, W.D.; Chinghuei, K. Salicylic acid-mediated hydrogen peroxide accumulation and protection against Cd toxicity in rice leaves. Plant Soil 2010, 329, 327–337. [Google Scholar] [CrossRef]
- Guo, B.; Liang, Y.; Zhu, Y. Does salicylic acid regulate antioxidant defense system, cell death, cadmium uptake and partitioning to acquire cadmium tolerance in rice? J. Plant Physiol. 2009, 166, 20–31. [Google Scholar] [CrossRef]
- Guo, B.; Liang, Y.; Zhu, Y.; Zhao, F. Role of salicylic acid in alleviating oxidative damage in rice roots (Oryza sativa) subjected to cadmium stress. Environ. Pollut. 2007, 147, 743–749. [Google Scholar] [CrossRef]
- Bai, X.; Dong, Y.; Kong, J.; Xu, L.; Liu, S. Effects of application of salicylic acid alleviates cadmium toxicity in perennial ryegrass. Plant Growth Regul. 2015, 75, 695–706. [Google Scholar] [CrossRef]
- Pál, M.; Szalai, G.; Horváth, E.; Janda, T.; Páldi, E. Effect of salicylic acid during heavy metal stress. Acta Biol. Szeged. 2002, 46, 119–120. [Google Scholar]
- Wang, Y.Y.; Wang, Y.; Li, G.Z.; Hao, L. Salicylic acid-altering Arabidopsis plant response to cadmium exposure: Underlying mechanisms affecting antioxidation and photosynthesis-related processes. Ecotoxicol. Environ. Saf. 2019, 169, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Liu, C.; Li, H.; Yi, K.; Ding, N.; Li, N.; Lin, Y.; Fu, Q. Endogenous salicylic acid is required for promoting cadmium tolerance of Arabidopsis by modulating glutathione metabolisms. J. Hazard. Mater. 2016, 316, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.; Ji, J.; Jia, C.; Guan, W.; Li, X.; Jin, C.; Wang, G. A GSHS-like gene from Lycium chinense maybe regulated by cadmium induced endogenous salicylic acid and overexpression of this gene enhances tolerance to cadmium stress in Arabidopsis. Plant Cell Rep. 2015, 34, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Sun, L.; Ma, C.; Li, L.; Li, G.; Hao, L. Reducing basal salicylic acid enhances Arabidopsis tolerance to lead or cadmium. Plant Soil 2013, 372, 309–318. [Google Scholar] [CrossRef]
- Zawoznik, M.S.; Groppa, M.D.; Tomaro, M.L.; Benavides, M.P. Endogenous salicylic acid potentiates cadmium-induced oxidative stress in Arabidopsis thaliana. Plant Sci. 2007, 173, 190–197. [Google Scholar] [CrossRef]
- Guo, B. Role of salicylic acid in mitigating cadmium toxicity in plants. In Cadmium Toxicity and Tolerance in Plants; Hasanuzzaman, M., Prasad, M.N.V., Fujita, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Chapter 14; pp. 349–374. [Google Scholar]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Influence of salicylic acid on H2O2 production, oxidative stress, and H2O2-metabolizing enzymes salicylic acid-mediated oxidative damage requires H2O2. Plant Physiol. 1997, 115, 137–149. [Google Scholar] [CrossRef]
- Sanità di Toppi, L.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Souza, V.L.; Almeida, A.A.F.D.; Lima, S.G.C.; Cascardo, J.; Silva, D.D.C.; Mangabeira, P.A.O.; Gomes, F.P. Morphophysiological responses and programmed cell death induced by cadmium in Genipa Americana L. (Rubiaceae). Biometals 2011, 24, 59–71. [Google Scholar] [CrossRef]
- Qian, H.; Li, J.; Pan, X.; Jiang, H.; Sun, L.; Fu, Z. Photoperiod and temperature influence cadmium’s effects on photosynthesis-related gene transcription in Chlorella vulgaris. Ecotoxicol. Environ. Saf. 2010, 73, 1202–1206. [Google Scholar] [CrossRef]
- Perfus-Barbeoch, L.; Leonhardt, N.; Vavaddeur, A.; Forestier, C. Heavy metal toxicity: Cadmium permeates through calcium channels and disturbs the plant water status. Plant J. 2002, 32, 539–548. [Google Scholar] [CrossRef]
- Boussama, N.; Ouariti, O.; Suzuki, A.; Ghorbal, M.H. Cd-stress on nitrogen assimilation. J. Plant Physiol. 1998, 155, 310–317. [Google Scholar] [CrossRef]
- Iannone, M.F.; Rosales, E.P.; Groppa, M.D.; Benavides, M.P. Reactive oxygen species formation and cell death in catalase-deficient tobacco leaf disks exposed to cadmium. Protoplasma 2010, 245, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Vicente, R.S.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.; MacKerness, S.A.; Page, T.; John, C.F.; Murphy, A.M.; Carr, J.P.; Buchanan-Wollaston, V. Salicylic acid has a role in regulating gene expression during leaf senescence. Plant J. 2000, 23, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Bowling, S.A.; Clarke, J.D.; Liu, Y.; Klessig, D.F.; Dong, X. The cpr5 mutant of Arabidopsis expresses both NPR1-dependent and NPR1-independent resistance. Plant Cell 1997, 9, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Rate, D.N.; Cuenca, J.V.; Bowman, G.R.; Guttman, D.S.; Greenberg, J.T. The gain-of-function Arabidopsis acd6 mutant reveals novel regulation and function of the salicylic acid signaling pathway in controlling cell death, defenses and cell growth. Plant Cell 1999, 11, 1695–1708. [Google Scholar] [CrossRef]
- Rate, D.N.; Greenberg, J.T. The Arabidopsis aberrant growth and death2 mutant shows resistance to Pseudomonas syringae and reveals a role for NPR1 in suppressing hypersensitive cell death. Plant J. 2001, 27, 203–211. [Google Scholar] [CrossRef]
- Šašek, V.; Janda, M.; Delage, E.; Puyaubert, J.; Guivarc’h, A.; López Maseda, E.; Dobrev, P.I.; Caius, J.; Bóka, K.; Valentová, O.; et al. Constitutive salicylic acid accumulation in pi4kIIIβ1β2 Arabidopsis plants stunts rosette but not root growth. New Phytol. 2014, 203, 805–816. [Google Scholar] [CrossRef]
- Janda, M.; Ruelland, E. Magical mystery tour: Salicylic acid signalling. Environ. Exp. Bot. 2015, 114, 117–128. [Google Scholar] [CrossRef]
- Drazic, G.; Mihailovic, N. Modification of cadmium toxicity in soybean seedlings by salicylic acid. Plant Sci. 2005, 168, 511–517. [Google Scholar] [CrossRef]
- Zhu, X.F.; Lei, G.J.; Jiang, T.; Liu, Y.; Li, G.X.; Zheng, S.J. Cell wall polysaccharides are involved in P-deficiency-induced Cd exclusion in Arabidopsis thaliana. Planta 2012, 236, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Tao, Q.; Shohag, M.J.I.; Yang, X.; Sparks, D.L.; Liang, Y. Root cell wall polysaccharides are involved in cadmium hyperaccumulation in Sedum alfredii. Plant Soil 2015, 389, 387–399. [Google Scholar] [CrossRef]
- Parrotta, L.; Guerriero, G.; Sergeant, K.; Cai, G.; Hausman, J.F. Target or barrier? The cell wall of early- and later-diverging plants vs cadmium toxicity: Differences in the response mechanisms. Front. Plant Sci. 2015, 6, 133. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.T.; Stein, M.; Hou, B.H.; Vogel, J.P.; Edwards, H.; Somerville, S.C. Loss of a callose synthase results in salicylic acid-dependent disease resistance. Science 2003, 301, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Napoleão, T.A.; Soares, G.; Vital, C.E.; Bastos, C.; Castro, R.; Loureiro, M.E.; Giordano, A. Methyl jasmonate and salicylic acid are able to modify cell wall but only salicylic acid alters biomass digestibility in the model grass Brachypodium distachyon. Plant Sci. 2017, 263, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Liang, Y.C.; Li, Z.J.; Guo, W. Role of salicylic acid in alleviating cadmium toxicity in rice roots. J. Plant Nutr. 2007, 30, 427–439. [Google Scholar] [CrossRef]
- Shakirova, F.M.; Allagulova, C.R.; Maslennikova, D.R.; Klyuchnikova, E.O.; Avalbaev, A.M.; Bezrukova, M.V. Salicylic acid-induced protection against cadmium toxicity in wheat plants. Environ. Exp. Bot. 2016, 122, 19–28. [Google Scholar] [CrossRef]
- Cohen, C.K.; Fox, T.C.; Garvin, D.F.; Kochian, L.V. The role of iron-deficiency stress responses in stimulating heavy-metal transport in plants. Plant Physiol. 1998, 116, 1063–1072. [Google Scholar] [CrossRef]
- Clemens, S.; Antosiewicz, D.M.; Ward, J.M.; Schachtman, D.P.; Schroeder, J.I. The plant cdna LCT1 mediates the uptake of calciumand cadmium in yeast. Proc. Natl. Acad. Sci. USA 1998, 95, 12043–12048. [Google Scholar] [CrossRef]
- Fatima, R.N.; Javed, F.; Wahid, A. Salicylic acid modifies growth performance and nutrient status of rice (Oryza sativa) under cadmium stress. Int. J. Agric. Biol. 2014, 16, 1083–1090. [Google Scholar]
- Gordon, L.K.; Minibayeva, F.V.; Rakhmatullina, D.F.; Alyabyev, A.J.; Ogorodnikova, T.I.; Loseva, N.L.; Valitova, Y.N. Heat production of wheat roots induced by the disruption of proton gradient by salicylic acid. Thermochim. Acta 2004, 422, 101–104. [Google Scholar] [CrossRef]
- Harper, J.R.; Balke, N.E. Characterization of the inhibition of k absorption in oat roots by salicylic acid. Plant Physiol. 1981, 68, 1349–1353. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Garnier, L.; Simon-Plas, F.; Thuleau, P.; Agnel, J.P.; Blein, J.P.; Ranjeva, R.; Montillet, J.L. Cd affects tobacco cells by a series of three waves of reactive oxygen species that contribute to cytotoxicity. Plant Cell Environ. 2006, 29, 1956–1969. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.M.; Chien, P.S.; Huang, J. Distinct signaling pathways for induction of MAP kinase activities by cadmium and copper in rice roots. J. Exp. Bot. 2007, 58, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J.; Deckert, J.; Rucińska-Sobkowiak, R.; Gzyl, J.; Pawlak-Sprada, S.; Abramowski, D.; Jelonek, T.; Gwóźdź, E.A. Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellowem lupine plants. Plant Physiol. Biochem. 2012, 58, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Durner, J.; Klessig, D.F. Salicylic acid is a modulator of tobacco and mammalian catalases. J. Biol. Chem. 1996, 271, 28492–28501. [Google Scholar] [CrossRef]
- Norman, C.; Howell, K.A.; Millar, A.H.; Whelan, J.M.; Day, D.A. Salicylic acid is an uncoupler and inhibitor of mitochondrial electron transport. Plant Physiol. 2004, 134, 492–501. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef]
- Popova, L.P.; Maslenkova, L.T.; Yordanova, R.Y.; Ivanova, A.P.; Krantev, A.P.; Szalai, G.; Janda, T. Exogenous treatment with salicylic acid attenuates cadmium toxicity in pea seedlings. Plant Physiol. Biochem. 2009, 47, 224–231. [Google Scholar] [CrossRef]
- Szalai, G.; Kellős, T.; Galiba, G.; Kocsy, G. Glutathione as an antioxidant and regulatory molecule in plants subjected to abiotic stresses. J. Plant Growth Regul. 2009, 28, 66–80. [Google Scholar] [CrossRef]
- Chaouch, S.; Queval, G.; Vanderauwera, S.; Mhamdi, A.; Vandorpe, M.; Langlois-Meurinne, M.; Van Breusegem, F.; Saindrenan, P.; Noctor, G. Peroxisomal hydrogen peroxide is coupled to biotic defense responses by ISOCHORISMATE SYNTHASE 1 in a daylength-related manner. Plant Physiol. 2010, 153, 1692–1705. [Google Scholar] [CrossRef] [PubMed]
- Chaouch, S.; Queval, G.; Noctor, G. AtRbohF is a crucial modulator of defence-associated metabolism and a key actor in the interplay between intracellular oxidative stress and pathogenesis responses in Arabidopsis. Plant J. 2012, 69, 613–627. [Google Scholar] [CrossRef] [PubMed]
- Mateo, A.; Funck, D.; Muhlenbock, P.; Kular, B.; Mullineaux, P.M.; Karpinski, S. Controlled levels of salicylic acid are required for optimal photosynthesis and redox homeostasis. J. Exp. Bot. 2006, 57, 1795–1807. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Sejir, C.; Amna, M.; Guillaume, Q.; Bernd, Z.; Graham, N. Functional analysis of Arabidopsis mutants points to novel roles for glutathione in coupling H2O2 to activation of salicylic acid accumulation and signaling. Antioxid. Redox Signal. 2012, 18, 2106–2121. [Google Scholar]
- Freeman, J.L.; Garcia, D.; Kim, D.; Hopf, A.; Salt, D.E. Constitutively elevated salicylic acid signals glutathione-mediated nickel tolerance in Thlaspi nickel hyperaccumulators. Plant Physiol. 2005, 137, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Hager, J.; Chaouch, S.; Queval, G.; Yi, H.; Taconnat, L.; Saindrenan, P.; Gouia, H.; Issakidis-Bourguet, E.; Renou, J.P.; et al. Arabidopsis glutathione reductase1 plays a crucial role in leaf responses to intracellular hydrogen peroxide and in ensuring appropriate gene expression through both salicylic acid and jasmonic acid signaling pathways. Plant Physiol. 2010, 153, 1144–1160. [Google Scholar] [CrossRef] [PubMed]
- Michele, R.D.; Vurro, E.; Rigo, C.; Costa, A.; Elviri, L.; Valentin, M.D.; Careri, M.; Zottini, M.; Sanità di Toppi, L.; Lo Schiavo, F. Nitric oxide is involved in cadmium-induced programmed cell death in Arabidopsis suspension cultures. Plant Physiol. 2009, 150, 217–228. [Google Scholar] [CrossRef]
- Kovács, V.; Gondor, O.K.; Szalai, G.; Darkó, É.; Majláth, I.; Janda, T.; Pál, M. Synthesis and role of salicylic acid in wheat varieties with different levels of cadmium tolerance. J. Hazard. Mater. 2014, 280, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Pál, M.; Horváth, E.; Janda, T.; Páldi, E.; Szalai, G. Cadmium stimulates the accumulation of salicylic acid and its putative precursors in maize (Zea mays L.) plants. Physiol. Plant. 2005, 125, 356–364. [Google Scholar] [CrossRef]
- Dean, J.V.; Mohammed, L.A.; Fitzpatrick, T. The formation, vacuolar localization, and tonoplast transport of salicylic acid glucose conjugates in tobacco cell suspension cultures. Planta 2005, 221, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Vanacker, H.; Lu, H.; Rate, D.N.; Greenberg, J.T. A role for salicylic acid and npr1 in regulating cell growth in Arabidopsis. Plant J. 2001, 28, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Sun, T.; Ao, K.; Peng, Y.; Zhang, Y.; Li, X.; Zhang, Y. Opposite roles of salicylic acid receptors NPR1 and NPR 3/ NPR 4 in transcriptional regulation of plant immunity. Cell 2018, 173, 1454–1467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Klessig, D.F. MAPK cascades in plant defense signaling. Trends Plant Sci. 2001, 6, 520–527. [Google Scholar] [CrossRef]
- Brodersen, P.; Petersen, M.; Nielsen, H.B.; Zhu, S.; Newman, M.A.; Shokat, K.M.; Rietz, S.; Parker, J.; Mundy, J. Arabidopsis MAP kinase 4 regulates salicylic acid- and jasmonic acid/ethylene-dependent responses via EDS1 and PAD4. Plant J. 2006, 47, 532–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.W.; Mao, Q.Q.; Luo, B.F.; Lin, X.Y.; Du, S.T. Mutation of mpk6 enhances cadmium tolerance in Arabidopsis plants by alleviating oxidative stress. Plant Soil 2013, 371, 387–396. [Google Scholar] [CrossRef]
- Bovet, L.; Feller, U.; Martinoia, E. Possible involvement of plant ABC transporters in cadmium detoxification: A cDNA sub-microarray approach. Environ. Int. 2005, 31, 263–267. [Google Scholar] [CrossRef]
- Eichhorn, H.; Klinghammer, M.; Becht, P.; Tenhaken, R. Isolation of a novel ABC-transporter gene from soybean induced by salicylic acid. J. Exp. Bot. 2006, 57, 2193–2201. [Google Scholar] [CrossRef] [Green Version]
- Pál, M.; Janda, T.; Szalai, G. Abscisic acid may alter the salicylic acid-related abiotic stress response in maize. J. Agron. Crop Sci. 2011, 197, 368–377. [Google Scholar] [CrossRef]
Figure 1.
Representation of SA exposure concentrations plotted against Cd exposure concentrations, drawn from the data reported in Table 1. * Table 1 and Figure 1 are adapted from the Reference [76].

Figure 2.
Possible roles of SA in alleviating Cd toxicity to plants. The dotted arrows mean possible signalling pathways. Red and green arrows indicate damage and positive effects, respectively. * Figure 2 is adapted from the reference [76].


{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of salicylic acid (SA) on cadmium (Cd) tolerance in plants.
| SA Treatment | Cd Treatment | Timeline | Plant Species | Main Responses * Means Negative or No Effect | Reference | |
|---|---|---|---|---|---|---|
| Spraying | 600 μM, 10 days | 22.5 mg L−1 | Simultaneous | Potato (S. tuberosum L.) | I, II, III, VII | [31] |
| 100 μM, 1 time | 30, 60 and 120 mg kg−1 (pot) | Simultaneous | Peppermint (Mentha piperita) | I, II, III | [32] | |
| 50 μM, 4 times in a 3-day interval | 75, 150, and 300 mg kg−1 | Simultaneous | Oilseed rape (Brassica napus) | II, III, V, VI | [33] | |
| 10, 50, 100, and 200 μM each day treated for 50 mL last 4 days | 44.8 mg kg−1 | Pretreatment | Melon (Cucumis melo L.) | I, II, III | [34] | |
| 500 μM, 1 time | 40 mg kg−1 | Pretreatment | Soybean (Glycine max L. cv. Liaoxing 1) | I, II, III | [35] | |
| 2170 μM 1 time | 56 and 112 mg kg−1 | Simultaneous | Radish (Raphanus sativus) | I, *IV | [36] | |
| 1000 μM for 10 mL, 45 times in a day interval | 100 and 200 mg L−1 | Simultaneous | Indian mustard (Brassica juncea) | I, II, III, IV, V | [37] | |
| Presoaking | 500 μM for 24 h. | 112 mg L−1 for 72 h. | Pretreatment | Mungbean (Vigna radiata L. Wilczek) | I, II | [38] |
| 250 or 1000 μM for 8 h | 5.6 and 11.2 mg L−1 for 10 days | Pretreatment | Flax (Linum usitatissimum L.) | Lipids | [39] | |
| 250 or 1000 μM for 8 h | 5.6 and 11.2 mg L−1 for 10 days | Pretreatment | Flax (Linum usitatissimum L.) | I, II | [40] | |
| 500 μM for 12 h | 0.56, 1.12, and 5.60 mg L−1 for 7 days | Pretreatment | Kentucky bluegrass | I, II, III, *IV, V | [41] | |
| 500 μM for 12 h | 56 and 112 mg kg−1 for 56 days | Pretreatment | Wheat (Triticum aestivum L. cv. Giza 168) | I, II, III, *IV | [42] | |
| 500 μM for 6 h | 1.12, 1.68, and 2.80 mg L−1 for 14 days | Pretreatment | Maize (Zea mays) | VI | [43] | |
| 250 or 1000 μM for 8 h | 5.6 and 11.2 mg L−1 for 10 days | Pretreatment | Flax (cv. Viking) | I, II | [44] | |
| 250 or 1000 μM for 8 h | 5.6 and 11.2 mg L−1 for 10 days | Pretreatment | Flax (Linum usitatissimum L.) | I, *IV, VI | [45] | |
| 100 μM for 12 h | 5.6 and 11.2 mg L−1 for 6 days | Pretreatment | Legume (Phaseolus aureus and Vicia sativa) | I, II | [46] | |
| 250 and 500 μM for 12 h | 5.6 mg L−1 for 12 days | Pretreatment | Bean (R. communis cv. Zibi 5) | I, III, IV | [30] | |
| 250 and 1000 μM for 12 h | 5.6 and 11.2 mg L−1 for 12 days | Pretreatment | Flax (Linum usitatissimum L.) | I, II, III, IV, V | [47] | |
| 500 μM for 20 h | 11.2, 44.8 and 112 mg kg−1 for 30 days | Pretreatment | Wheat (Triticum aestivum L.) | I, II, III | [48] | |
| 100 μM for 3 h | 3, 5, and 7 mg kg−1 for 3 days | Pretreatment | Soybean (Balkan, L608) | II, III, *IV | [49] | |
| 500 μM for 6 h | 25, 50, and 100 mg kg−1 | Pretreatment | Hemp (Cannabis sativa L.) | I, II, III, *IV | [50] | |
| 500 μM for 6 h | 1.12, 1.68, and 2.80 mg L−1 for 14 days | Pretreatment | Maize (Zea mays L., hybrid Norma) | I, II, III, IV | [51] | |
| 100 μM for 16 h | 11.2 and 112 mg L−1 for 1 day | Pretreatment | Rice (cv: Longai) | I, II | [52] | |
| 100 μM for 1, 3, 6 h | 3 and 5 mg L−1 for 7 days | Pretreatment | Alfalfa (Medicago sativa L. cv. Evropa) | I, IV, V | [49] | |
| 100 μM for 8 h | 1.12, 11.2, and 112 mg L−1 for 1 day | Pretreatment | Rice (cv: Longai) | I, II, *IV | [53] | |
| 500 μM for 6 h | 2.8 mg L−1 for 12 days | Pretreatment | Barley (Hordeum vulgare cv Gerbel) | I, II, III, IV, V, VI, VII | [54] | |
| Hydroponic application | 10 μM for 15 days | 16.8 mg L−1 for 15 days | Simultaneous | Rice (Oryza sativa L. Galileo)) | I, II, III | [55] |
| 20 μM for 1 day | 150 mg L−1 for 9 days | Pretreatment | Nymphaea tetragona Georgi | II, III, *IV,V | [56] | |
| 50 μM for 7 days | 1.12 mg L−1 for 7 days | Simultaneously | Lemna minor | II, III, IV, V | [57] | |
| 50 μM for 1 day | 11.2 mg L−1 for 8 h | Pretreatment | Wheat (Triticum aestivum L.) | I, *IV, Hormones | [33] | |
| 500 μM for 24 h | 56 mg L−1 for 1 day | Pretreatment | Maize (Zea mays L., hybrid Norma) | II, III, *IV, VI | [58] | |
| 100, 200, 300 and 400 μM for 14 days | 11.2 mg L−1 for 14 days | Simultaneous | Ryegrass (Lolium perenne L.) | I, II, III, *IV, V | [50] | |
| 50 μM for 10 days | 5.6 mg L−1 for 10 days | Simultaneous | Rice (Oryza sativa cv. HUR3022) | I, II, III | [59] | |
| 100 μM for 14 days | 22.4 mg L−1 for 14 days | Simultaneously | Peanut (Arachis hypogaea L.) | I, II, III, *IV, V | [60] | |
| 250 and 500 μM for 10 mins | 1.68 mg L−1 for 3 and 6 h | Post-treatment | Barley (Hordeum vulgare L.) cv. Slaven | I, II, Auxin | [61] | |
| 200 μM for 14 days | 11.2 mg L−1 for 14 days | Simultaneously | Ryegrass (Lolium perenne L.) | I, II, III, VI | [62] | |
| 10, 50 and 100 μM for 7 days | 2.24 mg L−1 for 3 days | Pretreatment | Bean (Phaseolus vulgaris) | I, II, III, IV, V | [63] | |
| 60, 120, 250 and 500 mM | 5.6 mg L−1 for 5 days | Pretreatment | Soybean (Glycine max L., A6445RG) | II, III, *IV, V, VI, VII (HO−1) | [64] | |
| 1, 10, and 100 μM for 72 h | 5.6 mg L−1 for 1 day | Pretreatment | Alfalfa (Medicago sativa L. cv Zhongmu No.1) | I, II, *IV, VI, VII (HO−1) | [65] | |
| 3000 μM for 3 h | 560 mg L−1 for 1 day | Pretreatment | Rice (Oryza sativa L., cv. Taichung Native 1) | II, IV | [66] | |
| 10 μM for 72 h | 5.6 mg L−1 for 6 days | Pretreatment | Rice (O. sativa cv Jiahua 1) | I, II, IV | [67] | |
| 10 μM for 72 h | 5.6 mg L−1 for 6 days | Pretreatment | Rice (O. sativa cv Jiahua 1) | I, II | [68] | |
| 10 μM for 24 h | 5.6 mg L−1 for 6 days | Pretreatment | Rice (O. sativa cv Jiahua 1) | I, II, IV | [65] | |
| 1, 10, and 100 μM | 3 and 6 mg L−1 for 3 days | Simultaneous | Soybean (Glycinemax L. cv SG1) | *I, *IV, IV | [54] | |
| 500 μM for 24 h | 2.8 mg L−1 for 10 days | Pretreatment | Barley leaves (Hordeum vulgare cv Gerbel) | I, II, III, IV, V, VI, VII | [69] | |
| 500 μM for 24 h | 56 mg L−1 for 1 day | Pretreatment and simultaneously | Maize (Zea mays L., hybrid Norma) | *I, *II, *III, *VI | [70] | |
| SA mutants | Up and down-regulating endogenesis SA | 5.6 mg L−1 for 7 days | - | NahG,snc1 | I, II, III, IV, VII | [71] |
| Down-regulating endogenesis SA | 0.56 mg L−1 for 12 days | - | Sid2 | I, II, III, IV, V, VI, VII | [72] | |
| SA accumulation | 16.8 mg L−1 for 28 days | - | Lycium chinense | II, III, IV, VII(LcGSHS) | [73] | |
| Up and down-regulating endogenesis SA | 5.6, 11.2, and 16.8 mg L−1 for 7days | - | Accumulating mutant snc1, npr1−1, Reducing mutant nahG, snc1/nahG | *I, *II, *III | [74] | |
| Down-regulating endogenesis SA | 56 mg L−1 for 5 days | - | NahG | *II, *III, *VII (CAT1) | [75] |
I Growth, II antioxidant system, III photosynthesis, IV Cd uptake, V Ion uptake, VI phytochelatins, VII SA or Cd-induced genes.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guo, B.; Liu, C.; Liang, Y.; Li, N.; Fu, Q. Salicylic Acid Signals Plant Defence against Cadmium Toxicity. Int. J. Mol. Sci. 2019, 20, 2960. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122960
AMA Style
Guo B, Liu C, Liang Y, Li N, Fu Q. Salicylic Acid Signals Plant Defence against Cadmium Toxicity. International Journal of Molecular Sciences. 2019; 20(12):2960. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122960
Chicago/Turabian StyleGuo, Bin, Chen Liu, Yongchao Liang, Ningyu Li, and Qinglin Fu. 2019. "Salicylic Acid Signals Plant Defence against Cadmium Toxicity" International Journal of Molecular Sciences 20, no. 12: 2960. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122960
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.