Determination of the Role and Active Sites of PKC-Delta-Like from Lamprey in Innate Immunity

Abstract
:1. Introduction
2. Results
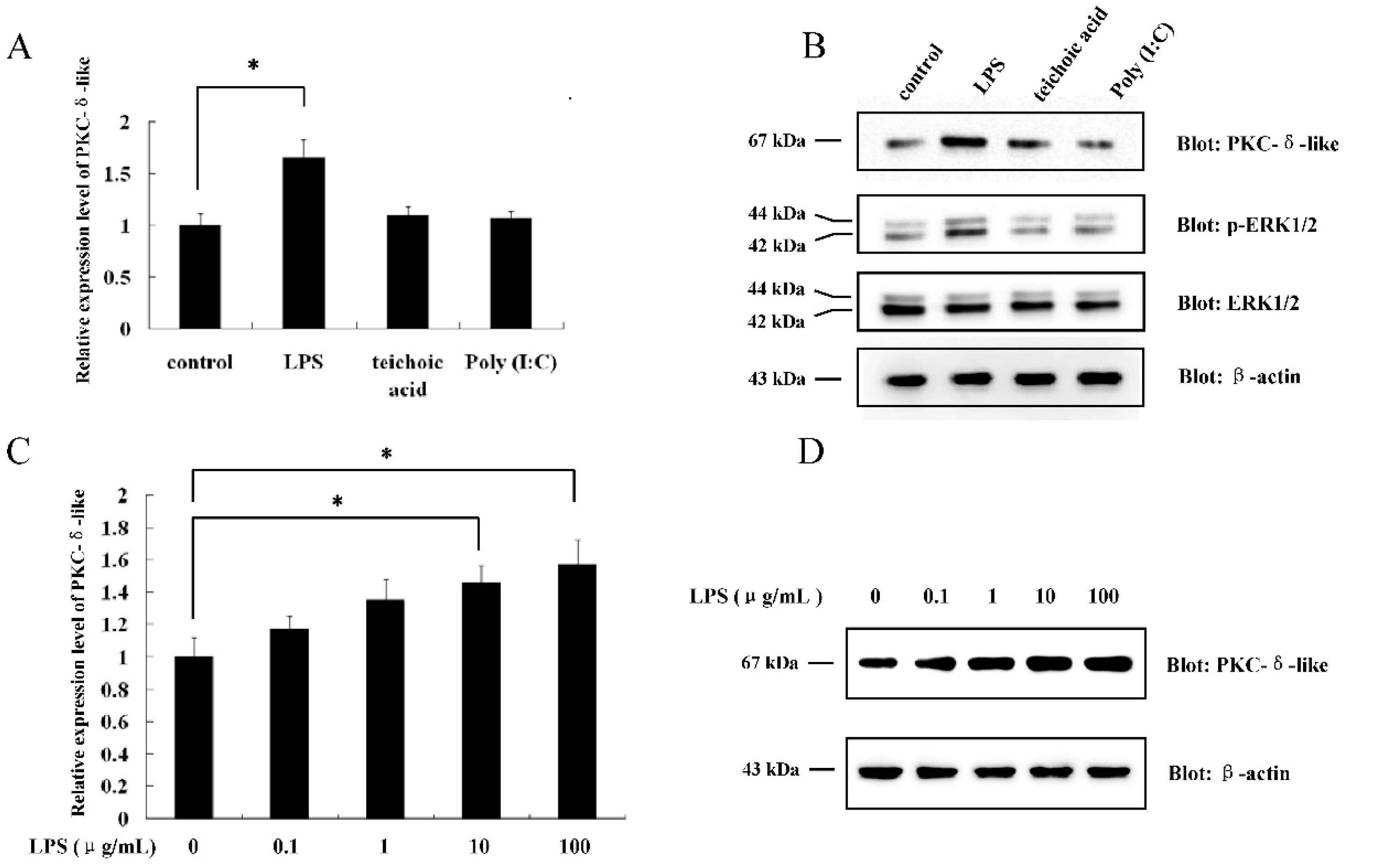
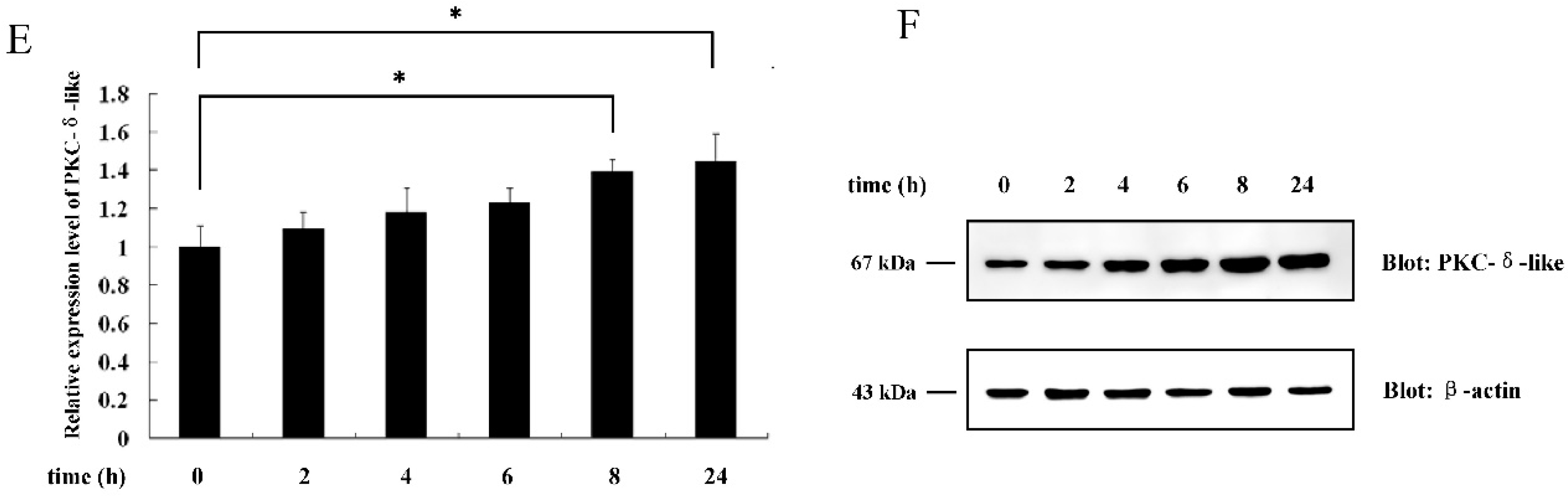
2.1. The Effect of Different Antigenic Stimuli on the Leukocytes of Lamprey
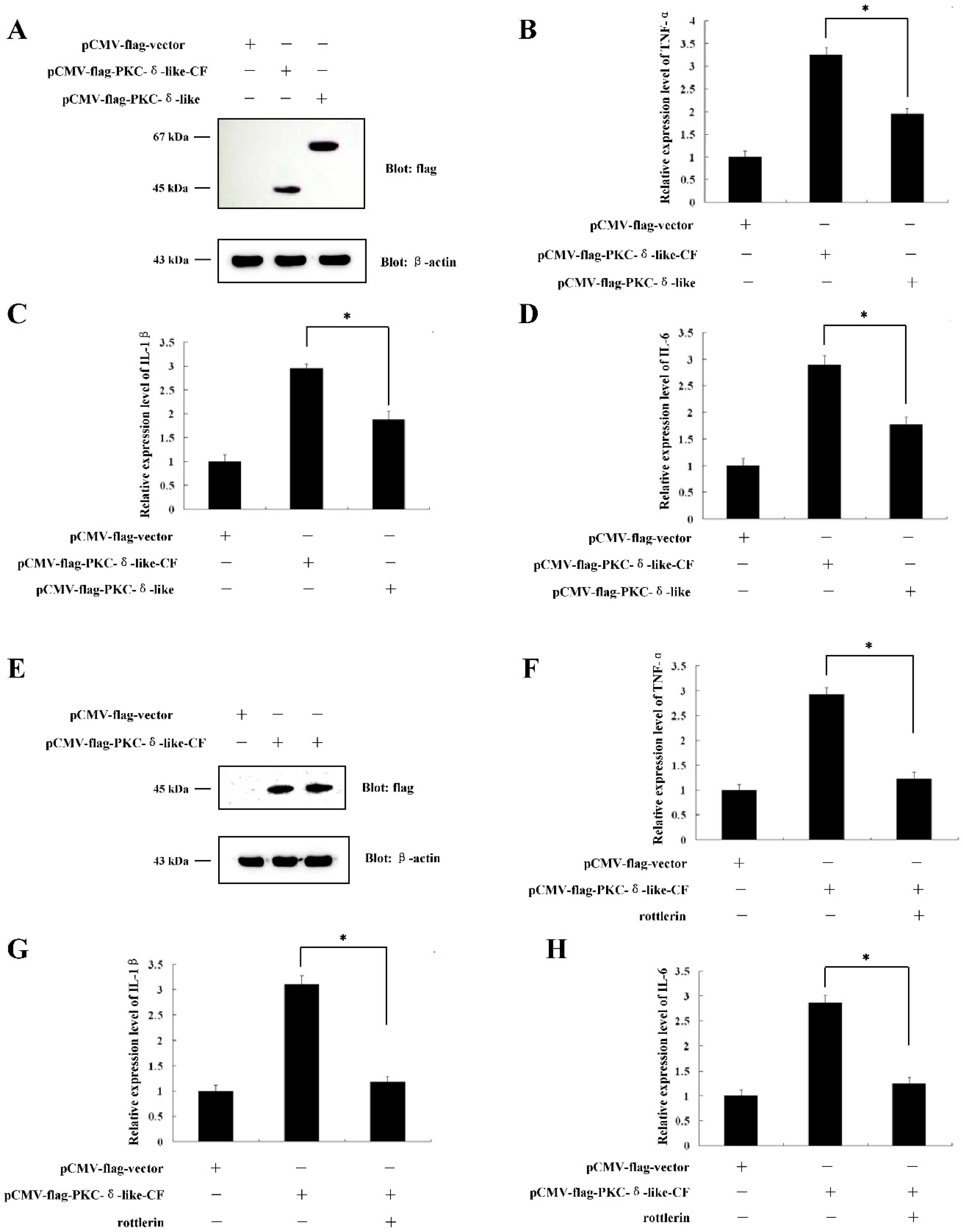
2.2. Overexpression of PKC-δ-LikeCould Induce Cellular Inflammation
2.3. The Catalytic Fragment of PKC-δ-Like Could also Stimulate Cellular Inflammation
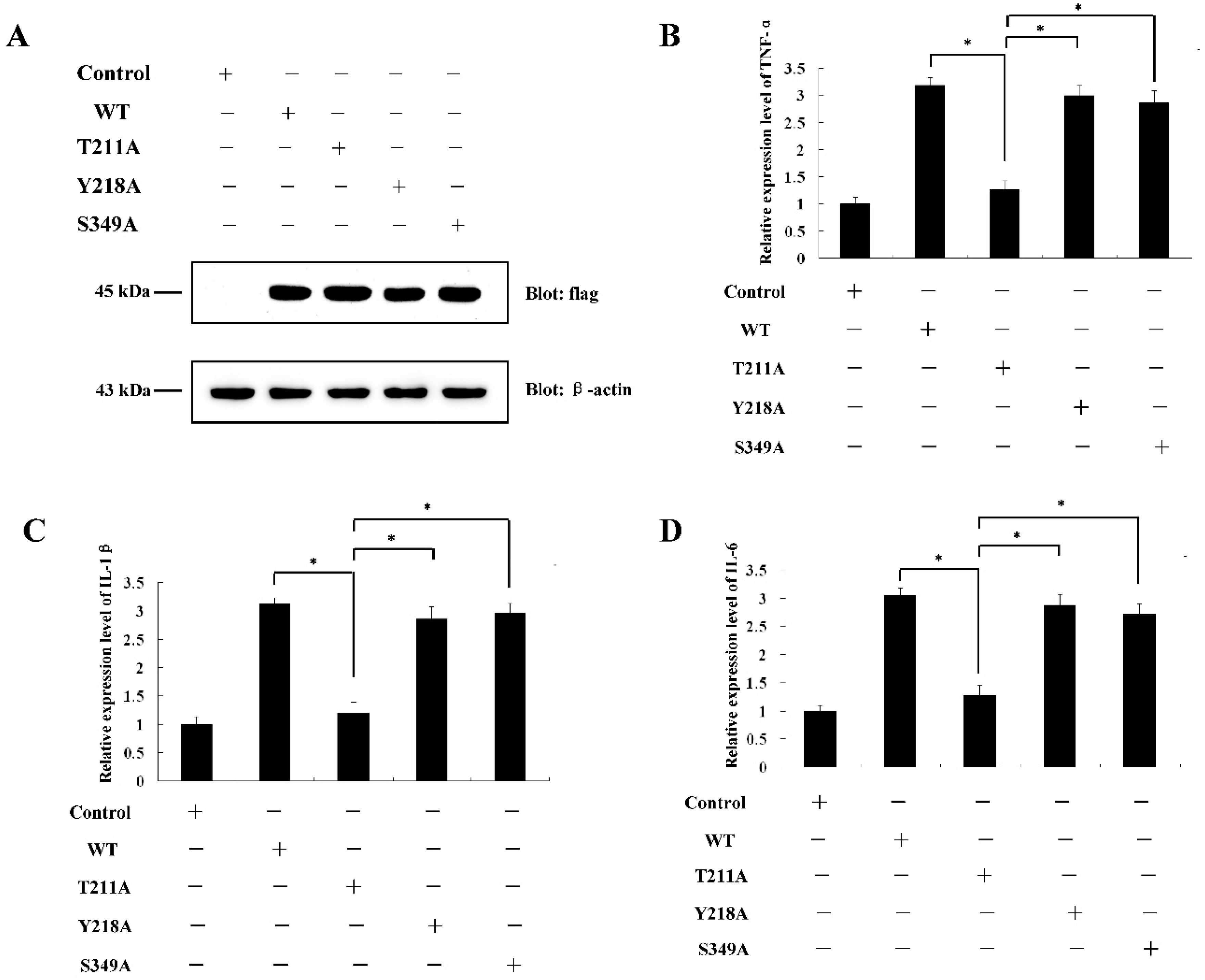
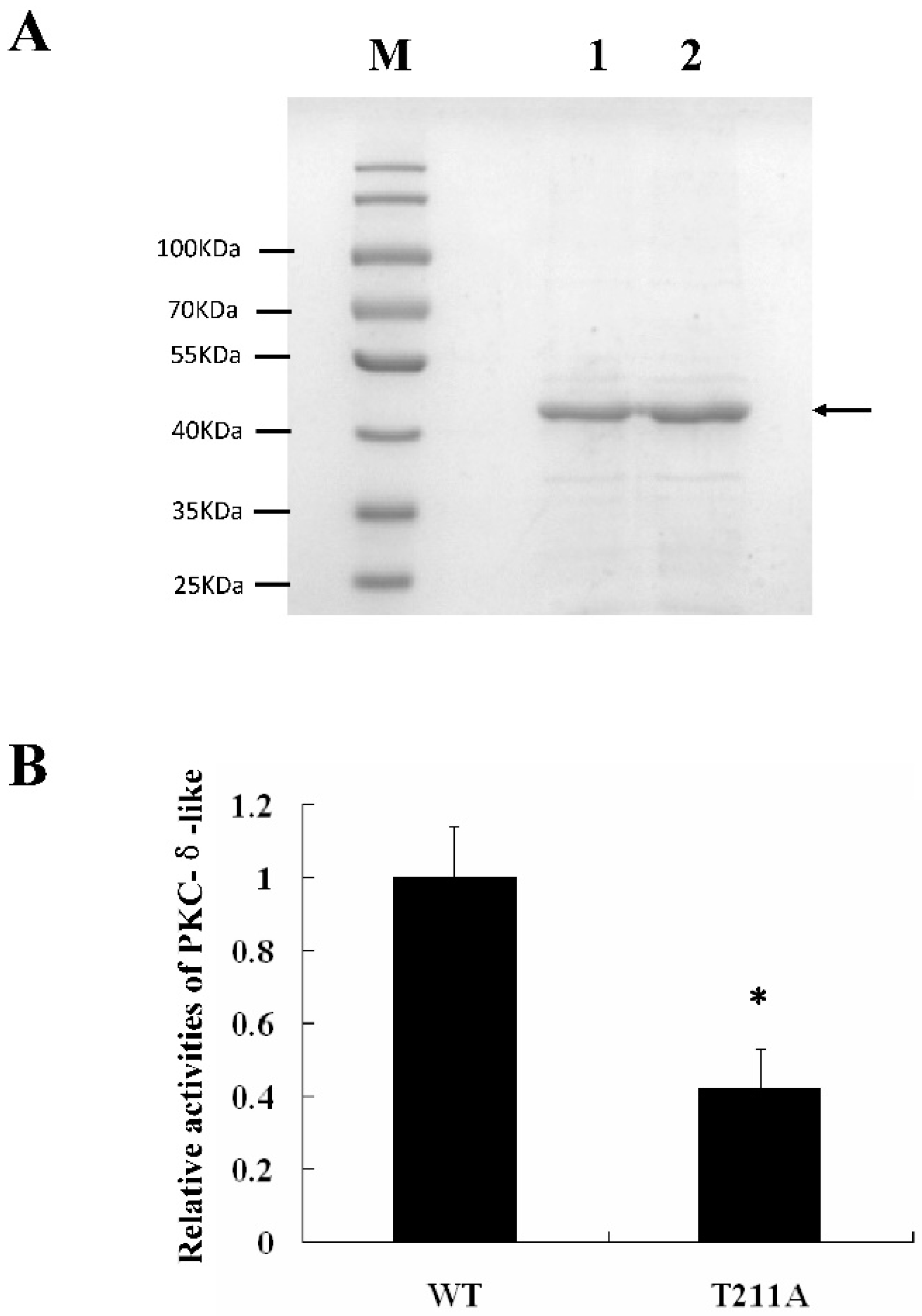
2.4. Determination of PKC-δ-Like Active Sites of the Cellular Inflammation Reaction
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Isolation of Leukocytes and Cell Culture
4.3. Real-Time PCR
4.4. Western Blotting
4.5. Cell Culture and Transfection
4.6. Construction of PKC-δ-Like-CF Mutants
4.7. PKC-δ Kinase Activity Assay
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PKC | Protein kinase C |
| L. japonica | Lampetra japonica |
| VLR | variable lymphocyte receptor |
| LPS | lipopolysaccharide |
| PKC-δ-like-CF | catalytic fragment of PKC-δ-like |
| WT | wild type of PKC-δ-like-CF |
| LRR | leucine-rich repeat |
| mRNA | messenger RNA |
| qPCR | real-time quantitative PCR |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| PMSF | phenylmethanesulphonyl fluoride |
| PVDF | polyvinylidene fluoride |
| ECL | Enhanced Chemiluminescence |
References
- Inoue, M.; Kishimoto, A.; Takai, Y.; Nishizuka, Y. Studies on a cyclic nucleotide-independent protein kinase and its proenzyme in mammalian tissues. II. Proenzyme and its activation by calcium-dependent protease from rat brain. J. Biol. Chem. 1977, 252, 7610–7616. [Google Scholar] [PubMed]
- Takai, Y.; Kishimoto, A.; Inoue, M.; Nishizuka, Y. Studies on a cyclic nucleotide-independent protein kinase and its proenzyme in mammalian tissues. I. Purification and characterization of an active enzyme from bovine cerebellum. J. Biol. Chem. 1977, 252, 7603–7609. [Google Scholar] [PubMed]
- Ono, Y.; Fujii, T.; Ogita, K.; Kikkawa, U.; Igarashi, K.; Nishizuka, Y. Identification of three additional members of rat protein kinase C family: Delta-, epsilon- and zeta-subspecies. FEBS Lett. 1987, 226, 125–128. [Google Scholar] [CrossRef]
- Gschwendt, M.; Kittstein, W.; Marks, F. A novel type of phorbol ester-dependent protein phosphorylation in the particulate fraction of mouse epidermis. Biochem. Biophys. Res. Commun. 1986, 137, 766–774. [Google Scholar] [CrossRef]
- Mecklenbräuker, I.; Saijo, K.; Zheng, N.Y.; Leitges, M.; Tarakhovsky, A. Protein kinase Cdelta controls self-antigen-induced B-cell tolerance. Nature 2002, 416, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, A.; Nakayama, K.; Imaki, H.; Hirose, S.; Jiang, Y.; Abe, M.; Tsukiyama, T.; Nagahama, H.; Ohno, S.; Hatakeyama, S.; et al. Increased proliferation of B cells and auto-immunity in mice lacking protein kinase Cdelta. Nature 2002, 416, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Blake, R.A.; Garcia-Paramio, P.; Parker, P.J.; Courtneidge, S.A. Src promotes PKCdelta degradation. Cell Growth Differ. 1999, 10, 231–241. [Google Scholar] [PubMed]
- Cho, W. Membrane targeting by C1 and C2 domains. J. Biol. Chem. 2001, 276, 32407–32410. [Google Scholar] [CrossRef] [PubMed]
- Konishi, H.; Tanaka, M.; Takemura, Y.; Matsuzaki, H.; Ono, Y.; Kikkawa, U.; Nishizuka, Y. Activation of protein kinase C by tyrosine phosphorylation in response to H2O2. Proc. Natl. Acad. Sci. USA 1997, 94, 11233–11237. [Google Scholar] [CrossRef] [PubMed]
- Kronfeld, I.; Kazimirsky, G.; Lorenzo, P.S.; Garfield, S.H.; Blumberg, P.M.; Brodie, C. Phosphorylation of protein kinase C delta on distinct tyrosine residues regulates specific cellular functions. J. Biol. Chem. 2000, 275, 35491–83549. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Pandey, P.; Sabatini, D.; Kumar, M.; Majumder, P.K.; Bharti, A.; Carmichael, G.; Kufe, D.; Kharbanda, S. Functional interaction between RAFT1/FRAP/mTOR and protein kinase cdelta in the regulation of cap-dependent initiation of translation. EMBO J. 2000, 19, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Pappa, H.; Murray-Rust, J.; Dekker, L.V.; Parker, P.J.; McDonald, N.Q. Crystal structure of the C2 domain from protein kinase C-delta. Structure 1998, 6, 885–894. [Google Scholar] [CrossRef]
- Osório, J.; Rétaux, S. The lamprey in evolutionary studies. Dev. Genes Evol. 2008, 218, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Kuratani, S.; Kuraku, S.; Murakami, Y. Lamprey as an evo-devo model: Lessons from comparative embryology and molecular phylogenetics. Genesis 2002, 34, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Nikitina, N.; Bronner-Fraser, M.; Sauka-Spengler, T. The sea lamprey Petromyzon marinus: A model for evolutionary and developmental biology. Cold Spring Harb. Protoc. 2009, 2009, pdb-emo113. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, C.T.; Saha, N.R.; Zapata, A. Evolution and development of immunological structures in the lamprey. Curr. Opin. Immunol. 2007, 19, 535–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrin, B.R.; Cooper, M.D. Alternative adaptive immunity in jawless vertebrates. J. Immunol. 2010, 185, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Pancer, Z.; Cooper, M.D. The evolution of adaptive immunity. Annu. Rev. Immunol. 2006, 24, 497–518. [Google Scholar] [CrossRef] [PubMed]
- Hirano, M.; Das, S.; Guo, P.; Cooper, M.D. The evolution of adaptive immunity in vertebrates. Adv. Immunol. 2011, 109, 125–157. [Google Scholar] [PubMed]
- Boehm, T.; McCurley, N.; Sutoh, Y.; Schorpp, M.; Kasahara, M.; Cooper, M.D. VLR-based adaptive immunity. Annu. Rev. Immunol. 2012, 30, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.D.; Alder, M.N. The evolution of adaptive immune systems. Cell 2006, 124, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhu, S.; Zhao, H.; Li, Q. Identification and characterisation of lamprey protein kinase C delta-like gene. Sci. Rep. 2017, 7, 12214. [Google Scholar] [CrossRef]
- Pang, Y.; Li, C.; Wang, S.; Ba, W.; Yu, T.; Pei, G.; Bi, D.; Liang, H.; Pan, X.; Zhu, T.; et al. A novel protein derived from lamprey supraneural body tissue with efficient cytocidal actions against tumor cells. Cell Commun. Signal. 2017, 15, 42. [Google Scholar] [CrossRef]
- Su, P.; Liu, X.; Han, Y.; Zheng, Z.; Liu, G.; Li, J.; Li, Q. Identification and characterization of a novel IκB-ε-like gene from lamprey (Lampetra japonica) with a role in immune response. Fish. Shellfish Immunol. 2013, 35, 1146–1154. [Google Scholar] [CrossRef]
- Su, P.; Liu, X.; Pang, Y.; Liu, C.; Li, R.; Zhang, Q.; Liang, H.; Wang, H.; Li, Q. The archaic roles of the lamprey NF-κB (lj-NF-κB) in innate immune responses. Mol. Immunol. 2017, 92, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.K.; Jeong, S.K.; Kim, H.S.; Choi, K.J.; Seo, J.T.; Choi, E.H.; Ahn, S.K.; Lee, S.H. Rottlerin, a specific inhibitor of protein kinase C-delta, impedes barrier repair response by increasing intracellular free calcium. J. Investig. Dermatol. 2006, 126, 1348–1355. [Google Scholar] [CrossRef] [PubMed]
- Basu, A. Involvement of protein kinase C-delta in DNA damage-induced apoptosis. J. Cell Mol. Med. 2003, 7, 341–350. [Google Scholar] [CrossRef]
- Gschwendt, M. Protein kinase C delta. Eur. J. Biochem. 1999, 259, 555–564. [Google Scholar] [CrossRef]
- Zhao, M.; Xia, L.; Chen, G.Q. Protein kinase c δ in apoptosis: A brief overview. Arch. Immunol. Ther. Exp. (Warsz.) 2012, 60, 361–372. [Google Scholar] [CrossRef]
- Huxley, T.H. The Nature of the Craniofacial Apparatus of Petromyzon. J. Anat. Physiol. 1876, 10 Pt 2, 412–429. [Google Scholar]
- Xu, Y.; Zhu, S.W.; Li, Q.W. Lamprey: A model for vertebrate evolutionary research. Zool. Res. 2016, 37, 263–269. [Google Scholar] [PubMed]
- Pancer, Z.; Amemiya, C.T.; Ehrhardt, G.R.; Ceitlin, J.; Gartland, G.L.; Cooper, M.D. Somatic diversification of variable lymphocyte receptors in the agnathan sea lamprey. Nature 2004, 430, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wu, F.; Datta, R.; Kharbanda, S.; Kufe, D. Interaction between protein kinase C delta and the c-Abl tyrosine kinase in the cellular response to oxidative stress. J. Biol. Chem. 2000, 275, 7470–7473. [Google Scholar] [CrossRef] [PubMed]
- Szallasi, Z.; Denning, M.F.; Chang, E.Y.; Rivera, J.; Yuspa, S.H.; Lehel, C.; Olah, Z.; Anderson, W.B.; Blumberg, P.M. Development of a rapid approach to identification of tyrosine phosphorylation sites: Application to PKC delta phosphorylated upon activation of the high affinity receptor for IgE in rat basophilic leukemia cells. Biochem. Biophys. Res. Commun. 1995, 214, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, W.H.; Parekh, D.B.; Le Good, J.A.; Whelan, R.D.; Kelly, J.J.; Frech, M.; Hemmings, B.A.; Parker, P.J. Rapamycin-sensitive phosphorylation of PKC on a carboxy-terminal site by an atypical PKC complex. Curr. Biol. 1999, 9, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Altman, A.; Kong, K.F. Protein kinase C enzymes in the hematopoietic and immune systems. Annu. Rev. Immunol. 2016, 34, 511–538. [Google Scholar] [CrossRef] [PubMed]
- Platten, M.; Eitel, K.; Wischhusen, J.; Dichgans, J.; Weller, M. Involvement of protein kinase Cdelta and extracellular signal-regulated kinase-2 in the suppression of microglial inducible nitric oxide synthase expression by N-[3,4-dimethoxycinnamoyl]-anthranilic acid (tranilast). Biochem. Pharmacol. 2003, 66, 1263–1270. [Google Scholar] [CrossRef]
- Baig, M.S.; Liu, D.; Muthu, K.; Roy, A.; Saqib, U.; Naim, A.; Faisal, S.M.; Srivastava, M.; Saluja, R. Heterotrimeric complex of p38 MAPK, PKCδ, and TIRAP is required for AP1 mediated inflammatory response. Int. Immunopharmacol. 2017, 48, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.C.; Kim, S.H.; Jeong, M.W.; Baek, N.I.; Kim, K.T. Effect of rottlerin, a PKC-delta inhibitor, on TLR-4-dependent activation of murine microglia. Biochem. Biophys. Res. Commun. 2005, 337, 110–115. [Google Scholar] [CrossRef]
- Tiwari, R.L.; Singh, V.; Singh, A.; Barthwal, M.K. IL-1R-associated kinase-1 mediates protein kinase Cδ-induced IL-1β production in monocytes. J. Immunol. 2011, 187, 2632–2645. [Google Scholar] [CrossRef]
- Kontny, E.; Kurowska, M.; Szczepańska, K.; Maśliński, W. Rottlerin, a PKC isozyme-selective inhibitor, affects signaling events and cytokine production in human monocytes. J. Leukoc. Biol. 2000, 67, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Loegering, D.J.; Lennartz, M.R. Protein kinase C and toll-like receptor signaling. Enzyme Res. 2011, 2011, 537821. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Song, Y.; Chen, C.; Lu, Z.Z.; Zhang, Y. Stimulation of adenosine A(2B) receptors induces interleukin-6 secretion in cardiac fibroblasts via the PKC-delta-P38 signalling pathway. Br. J. Pharmacol. 2010, 159, 1598–1607. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) |
|---|---|
| Realtime-PCR | |
| PKC-δ-like-forward | GCATCTCCACGGAACGAC |
| PKC-δ-like-reverse | CCACCTCCACCTTCTCAACT |
| Lamprey-GAPDH-forward | ACCCCTTCATTGACCTGGAGTA |
| Lamprey-GAPDH-reverse | TGCTTACCCCATGGGATGTT |
| TNF-α-forward | GTCTCAGCCTCTTCTCATTC |
| TNF-α-reverse | CATAGAACTGATGAGAGGGA |
| IL-1β-forward | AAATACCTGTGGCCTTGGGC |
| IL-1β-reverse | CTTGGGATCCACACTCTCCAG |
| IL-6-forward | GAGTCCTTCAGAGAGATACAG |
| IL-6-reverse | CTGTGACTCCAGCTTATCTG |
| human-GAPDH-forward | TGGCCAAGGTCATCCATGACAAC |
| human-GAPDH-reverse | TCCAGAGGGGCCATCCACAGTCTTCTG |
| Mutation | |
| F-T211A | AGGAAACCTCGCCACCGCGTTCTGCGGCACCCCGG |
| R-T211A | CCGGGGTGCCGCAGAACGCGGTGGCGAGGTTTCCT |
| F-Y218A | CTGCGGCACCCCGGACGCCATCGCACCCGAGATCT |
| R-Y218A | AGATCTCGGGTGCGATGGCGTCCGGGGTGCCGCAG |
| F-S349A | CGAGTCCCCGCAGGTGGCGTGCCGAGGAAACAGCA |
| R-S349A | TGCTGTTTCCTCGGCACGCCACCTGCGGGGACTCG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Zhao, H.; Tian, Y.; Ren, K.; Zheng, N.; Li, Q. Determination of the Role and Active Sites of PKC-Delta-Like from Lamprey in Innate Immunity. Int. J. Mol. Sci. 2019, 20, 3357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133357
Xu Y, Zhao H, Tian Y, Ren K, Zheng N, Li Q. Determination of the Role and Active Sites of PKC-Delta-Like from Lamprey in Innate Immunity. International Journal of Molecular Sciences. 2019; 20(13):3357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133357
Chicago/Turabian StyleXu, Yang, Huan Zhao, Yang Tian, Kaixia Ren, Nan Zheng, and Qingwei Li. 2019. "Determination of the Role and Active Sites of PKC-Delta-Like from Lamprey in Innate Immunity" International Journal of Molecular Sciences 20, no. 13: 3357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133357