Revealing Adenosine A2A-Dopamine D2 Receptor Heteromers in Parkinson’s Disease Post-Mortem Brain through a New AlphaScreen-Based Assay

 , , and
, , and
Abstract
:1. Introduction
2. Results
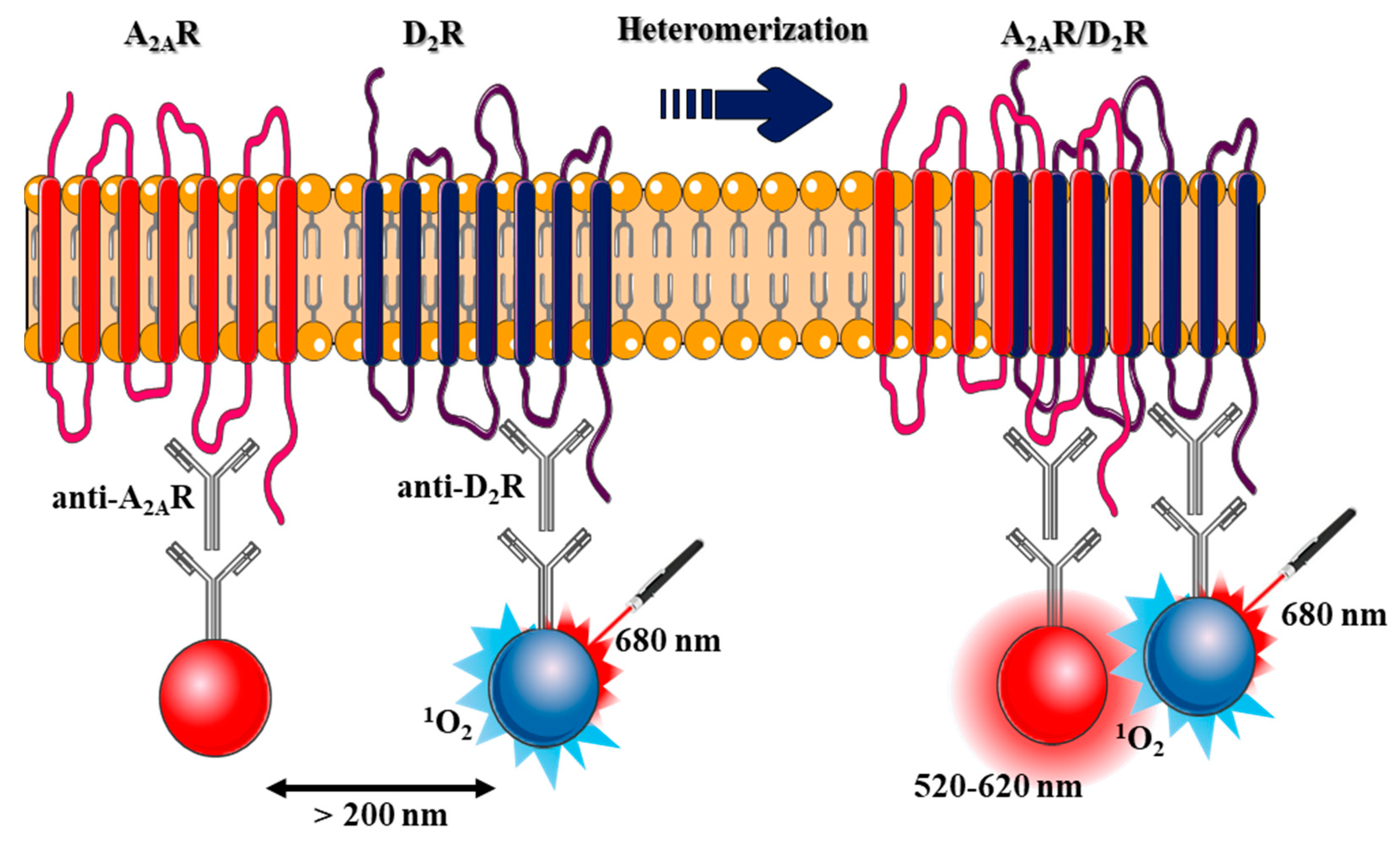
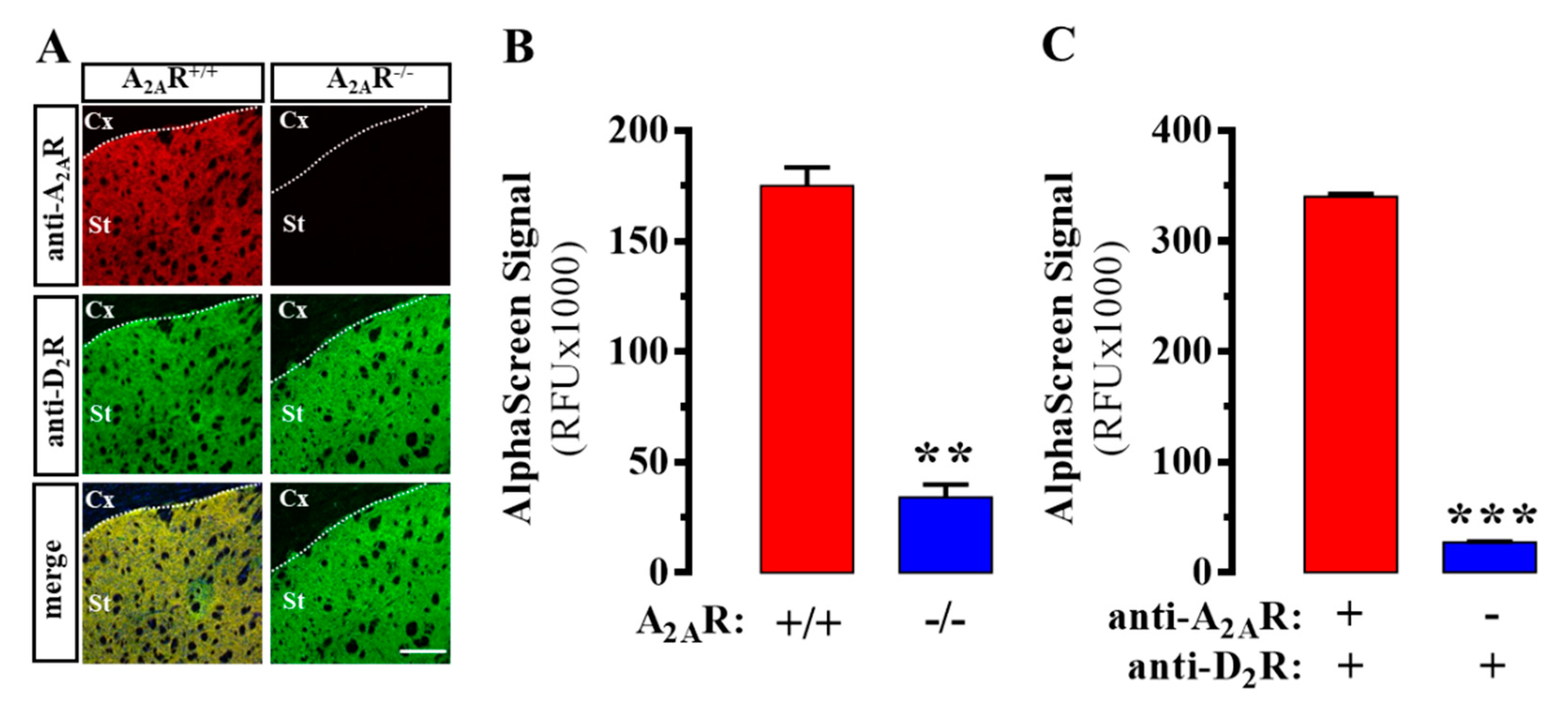
2.1. Design and Validation of the AlphaScreen Assay
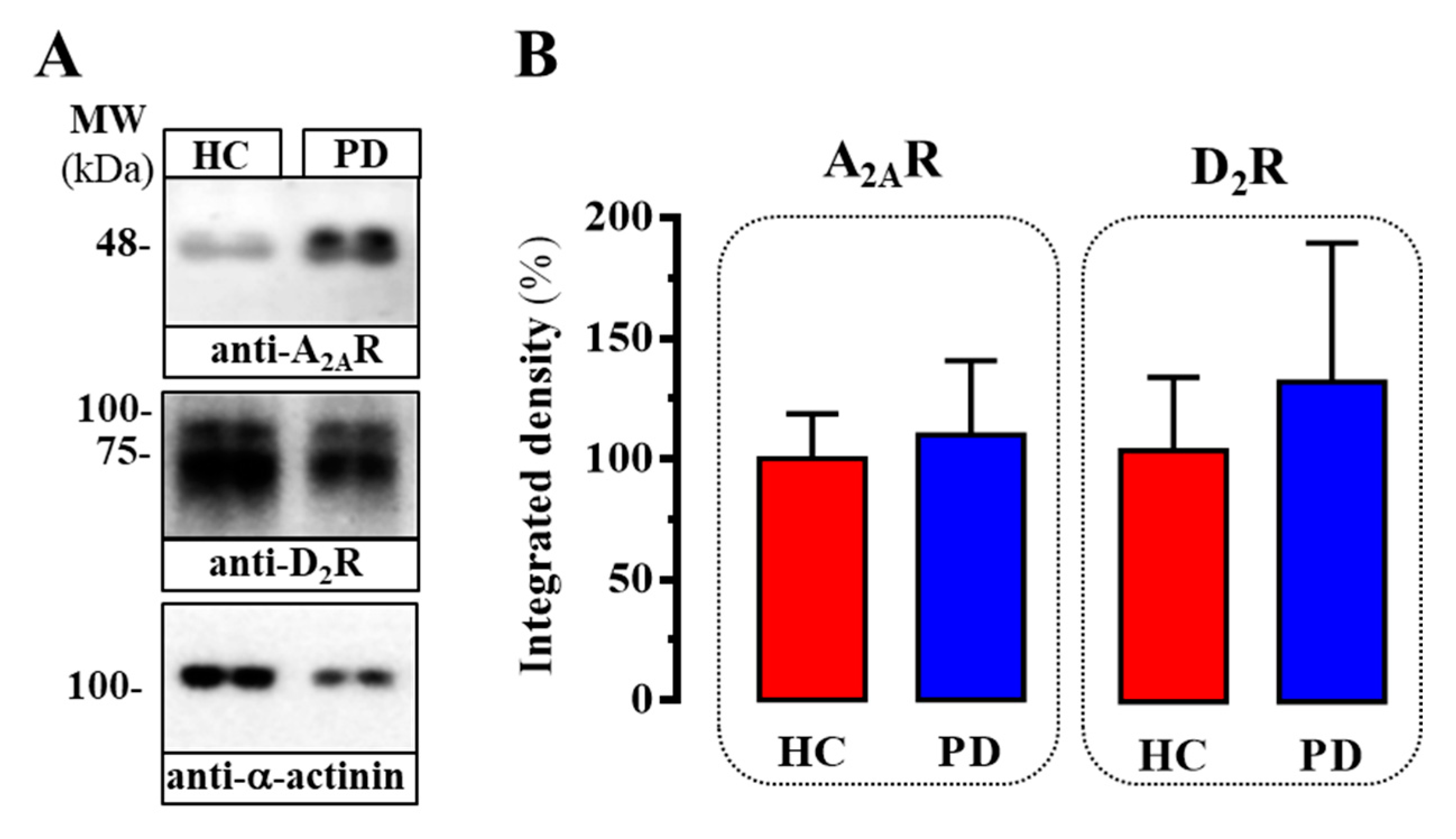
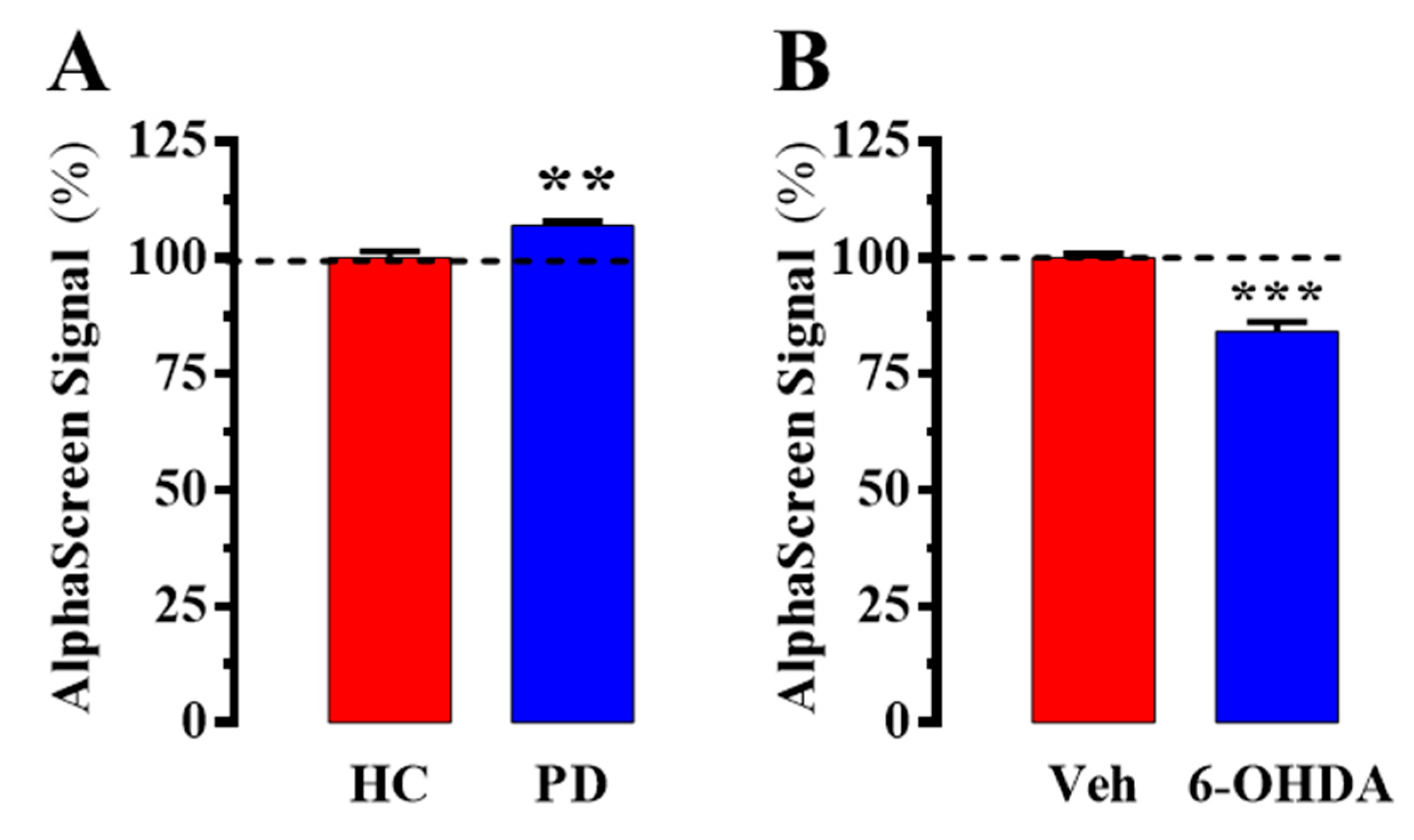
2.2. Evaluation of D2R/A2AR Oligomerization in PD Patients
3. Discussion
4. Materials and Methods
4.1. Human Brain Samples
4.2. Animals
4.3. Surgery
4.4. Membrane Preparation
4.5. Gel Electrophoresis and Immunoblotting
4.6. AlphaScreen Assay
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GPCR | G protein-coupled receptor |
| D2R | Dopamine D2 receptor |
| A2AR | Adenosine A2A receptor |
| HC | Healthy control |
| PD | Parkinson’s disease |
| 6-OHDA | 6-hydroxydopamine |
| S/B | Signal-to-background |
| HTS | High throughput screening |
| LBD | Lewy body disease |
| l-DOPA | 3,4-Dihydroxy-l-phenylalanine |
| BCA | Bicinchoninic acid assay |
| SDS/PAGE | Sodium dodecyl sulfate/polyacrylamide gel electrophoresis |
References
- Ciruela, F.; Vallano, A.; Arnau, J.M.; Sanchez, S.; Borroto-Escuela, D.O.; Agnati, L.F.; Fuxe, K.; Fernandez-Duenas, V. G protein-coupled receptor oligomerization for what? J. Recept. Signal Transduct. Res. 2010, 30, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.D.; Clarke, W.P.; von Zastrow, M.; Nichols, D.E.; Kobilka, B.; Weinstein, H.; Javitch, J.A.; Roth, B.L.; Christopoulos, A.; Sexton, P.M.; et al. Functional selectivity and classical concepts of quantitative pharmacology. J. Pharmacol. Exp. Ther. 2007, 320, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rozenfeld, R.; Devi, L.A. Receptor heterodimerization leads to a switch in signaling: beta-arrestin2-mediated ERK activation by mu-delta opioid receptor heterodimers. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2007, 21, 2455–2465. [Google Scholar] [CrossRef] [PubMed]
- Guitart, X.; Navarro, G.; Moreno, E.; Yano, H.; Cai, N.S.; Sanchez-Soto, M.; Kumar-Barodia, S.; Naidu, Y.T.; Mallol, J.; Cortes, A.; et al. Functional Selectivity of Allosteric Interactions within G Protein-Coupled Receptor Oligomers: The Dopamine D1-D3 Receptor Heterotetramer. Mol. Pharmacol. 2014, 86, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Sahlholm, K.; Gómez-Soler, M.; Valle-León, M.; López-Cano, M.; Taura, J.J.; Ciruela, F.; Fernández-Dueñas, V. Antipsychotic-Like Efficacy of Dopamine D2 Receptor-Biased Ligands is Dependent on Adenosine A2A Receptor Expression. Mol. Neurobiol. 2018, 55, 4952–4958. [Google Scholar] [CrossRef]
- De Lau, L.M.L.; Breteler, M.M.B. Epidemiology of Parkinson’s disease. Lancet. Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef]
- Ascherio, A.; Schwarzschild, M.A. The epidemiology of Parkinson’s disease: risk factors and prevention. Lancet Neurol. 2016, 15, 1257–1272. [Google Scholar] [CrossRef]
- Fuxe, K.; Marcellino, D.; Borroto-Escuela, D.O.; Guescini, M.; Fernandez-Duenas, V.; Tanganelli, S.; Rivera, A.; Ciruela, F.; Agnati, L.F. Adenosine-dopamine interactions in the pathophysiology and treatment of CNS disorders. CNS Neurosci. Ther. 2010, 16, e18–e42. [Google Scholar] [CrossRef]
- Trifilieff, P.; Rives, M.L.; Urizar, E.; Piskorowski, R.A.; Vishwasrao, H.D.; Castrillon, J.; Schmauss, C.; Slättman, M.; Gullberg, M.; Javitch, J.A. Detection of antigen interactions ex vivo by proximity ligation assay: endogenous dopamine D2-adenosine A2A receptor complexes in the striatum. Biotechniques 2011, 51, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Bonaventura, J.; Rico, A.J.; Moreno, E.; Sierra, S.; Sánchez, M.; Luquin, N.; Farré, D.; Müller, C.E.; Martínez-Pinilla, E.; Cortés, A.; et al. l-DOPA-treatment in primates disrupts the expression of A2A adenosine-CB1 cannabinoid-D2 dopamine receptor heteromers in the caudate nucleus. Neuropharmacology 2014, 79, 90–100. [Google Scholar] [CrossRef]
- Fernández-Dueñas, V.; Taura, J.J.; Cottet, M.; Gómez-Soler, M.; López-Cano, M.; Ledent, C.; Watanabe, M.; Trinquet, E.; Pin, J.P.; Luján, R.; et al. Untangling dopamine-adenosine receptor-receptor assembly in experimental parkinsonism in rats. Dis. Models Mech. 2015, 8, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Longhena, F.; Faustini, G.; Missale, C.; Pizzi, M.; Bellucci, A. Dopamine transporter/α-synuclein complexes are altered in the post mortem caudate putamen of Parkinson’s disease: An in situ proximity ligation assay study. Int. J. Mol. Sc. 2018, 19, 1611. [Google Scholar] [CrossRef] [PubMed]
- Ullman, E.F.; Kirakossian, H.; Singh, S.; Wu, Z.P.; Irvin, B.R.; Pease, J.S.; Switchenko, A.C.; Irvine, J.D.; Dafforn, A.; Skold, C.N. Luminescent oxygen channeling immunoassay: measurement of particle binding kinetics by chemiluminescence. Proc. Natl. Acad. Sci. USA 1994, 91, 5426–5430. [Google Scholar] [CrossRef] [PubMed]
- Ullman, E.F.; Kirakossian, H.; Switchenko, A.C.; Ishkanian, J.; Ericson, M.; Wartchow, C.A.; Pirio, M.; Pease, J.; Irvin, B.R.; Singh, S.; et al. Luminescent oxygen channeling assay (LOCI): sensitive, broadly applicable homogeneous immunoassay method. Clin. Chem. 1996, 42, 1518–1526. [Google Scholar] [PubMed]
- Eglen, R.M.; Reisine, T.; Roby, P.; Rouleau, N.; Illy, C.; Bossé, R.; Bielefeld, M. The Use of AlphaScreen Technology in HTS: Current Status. Curr. Chem. Genom. 2008, 1, 2–10. [Google Scholar] [CrossRef]
- Yasgar, A.; Jadhav, A.; Simeonov, A.; Coussens, N.P. AlphaScreen-Based Assays: Ultra-High-Throughput Screening for Small-Molecule Inhibitors of Challenging Enzymes and Protein-Protein Interactions. Methods Mol. Biol. 2016, 1439, 77–98. [Google Scholar] [Green Version]
- Zhang, J.H.; Chung, T.D.Y.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Wydra, K.; Li, X.; Rodriguez, D.; Carlsson, J.; Jastrzębska, J.; Filip, M.; Fuxe, K. Disruption of A2AR-D2R Heteroreceptor Complexes After A2AR Transmembrane 5 Peptide Administration Enhances Cocaine Self-Administration in Rats. Mol. Neurobiol. 2018, 55, 7038–7048. [Google Scholar] [CrossRef] [Green Version]
- Taura, J.; Valle-León, M.; Sahlholm, K.; Watanabe, M.; Van Craenenbroeck, K.; Fernández-Dueñas, V.; Ferré, S.; Ciruela, F. Behavioral control by striatal adenosine A2A-dopamine D2 receptor heteromers. Genes Brain Behav. 2018, 17, e12432. [Google Scholar] [CrossRef]
- Fuxe, K.; Borroto-Escuela, D.O.; Romero-Fernandez, W.; Palkovits, M.; Tarakanov, A.O.; Ciruela, F.; Agnati, L.F. Moonlighting proteins and protein-protein interactions as neurotherapeutic targets in the G protein-coupled receptor field. Neuropsychopharmacology 2014, 39, 131–135. [Google Scholar] [CrossRef]
- Fuxe, K.; Agnati, L.F.; Jacobsen, K.; Hillion, J.; Canals, M.; Torvinen, M.; Tinner-Staines, B.; Staines, W.; Rosin, D.; Terasmaa, A.; et al. Receptor heteromerization in adenosine A2A receptor signaling: relevance for striatal function and Parkinson’s disease. Neurology 2003, 61, S19–S23. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.H.; El Baz, R.; Cuconati, A.; Arthos, J.; Jain, P.; Khan, Z.K. A novel high-throughput screening assay to identify inhibitors of HIV-1 gp120 protein interaction with DC-SIGN. J. Antivir. Antiretrovir. 2011, 3, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R. Parkinson’s disease: From pathogenesis to pharmacogenomics. Int. J. Mol. Sci. 2017, 18, 551. [Google Scholar] [CrossRef] [PubMed]
- Bohush, A.; Niewiadomska, G.; Filipek, A. Role of Mitogen Activated Protein Kinase Signaling in Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 2973. [Google Scholar] [CrossRef] [PubMed]
- Comi, C.; Ferrari, M.; Marino, F.; Magistrelli, L.; Cantello, R.; Riboldazzi, G.; Bianchi, M.L.E.; Bono, G.; Cosentino, M. Polymorphisms of dopamine receptor genes and risk of l-dopa–induced dyskinesia in parkinson’s disease. Int. J. Mol. Sci. 2017, 18, 242. [Google Scholar] [CrossRef] [PubMed]
- Piggott, M.A.; Marshall, E.F.; Thomas, N.; Lloyd, S.; Court, J.A.; Jaros, E.; Burn, D.; Johnson, M.; Perry, R.H.; McKeith, I.G.; et al. Striatal dopaminergic markers in dementia with Lewy bodies, Alzheimer’s and Parkinson’s diseases: rostrocaudal distribution. Brain 1999, 122, 1449–1468. [Google Scholar] [CrossRef] [PubMed]
- Ahlskog, J.E.; Richelson, E.; Nelson, A.; Kelly, P.J.; Okazaki, H.; Tyce, G.M.; van Heerden, J.A.; Stoddard, S.L.; Carmichael, S.W. Reduced D2 dopamine and muscarinic cholinergic receptor densities in caudate specimens from fluctuating parkinsonian patients. Ann. Neurol. 1991, 30, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, P.D.; Perry, R.H.; Crossman, A.R. A detailed anatomical analysis of neurotransmitter receptors in the putamen and caudate in Parkinson’s disease and Alzheimer’s disease. Neurosci. Lett. 1994, 169, 68–72. [Google Scholar] [CrossRef]
- Varani, K.; Vincenzi, F.; Tosi, A.; Gessi, S.; Casetta, I.; Granieri, G.; Fazio, P.; Leung, E.; MacLennan, S.; Granieri, E.; et al. A2A adenosine receptor overexpression and functionality, as well as TNF-alpha levels, correlate with motor symptoms in Parkinson’s disease. FASEB J. 2010, 24, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Mattila, P.M.; Röyttä, M.; Lönnberg, P.; Marjamäki, P.; Helenius, H.; Rinne, J.O. Choline acetytransferase activity and striatal dopamine receptors in Parkinson’s disease in relation to cognitive impairment. Acta Neuropathol. 2001, 102, 160–166. [Google Scholar] [PubMed]
- McFarland, N.R.; Burdett, T.; Desjardins, C.A.; Frosch, M.P.; Schwarzschild, M.A. Postmortem brain levels of urate and precursors in Parkinson’s disease and related disorders. Neurodegener. Dis. 2013, 12, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Villar-Menéndez, I.; Porta, S.; Buira, S.P.; Pereira-Veiga, T.; Díaz-Sánchez, S.; Albasanz, J.L.; Ferrer, I.; Martín, M.; Barrachina, M. Increased striatal adenosine A2A receptor levels is an early event in Parkinson’s disease-related pathology and it is potentially regulated by miR-34b. Neurobiol. Dis. 2014, 69, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Ledent, C.; Vaugeois, J.M.; Schiffmann, S.N.; Pedrazzini, T.; El Yacoubi, M.; Vanderhaeghen, J.J.; Costentin, J.; Heath, J.K.; Vassart, G.; Parmentier, M. Aggressiveness, hypoalgesia and high blood pressure in mice lacking the adenosine A2a receptor. Nature 1997, 388, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.D.; Gebhart, G.F.; Gonder, J.C.; Keeling, M.E.; Kohn, D.F. Special report: the 1996 guide for the care and use of laboratory animals. ILAR J.Natl. Res. Counc. Inst. Lab. Anim. Resour. 1997, 38, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Benito, P.; Fernández-Dueñas, V.; Carmona, M.; Escobar, L.A.; Torrejón-Escribano, B.; Aso, E.; Ciruela, F.; Ferrer, I. Locus coeruleus at asymptomatic early and middle Braak stages of neurofibrillary tangle pathology. Neuropathol. Appl. Neurobiol. 2017, 43, 373–392. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parkinson’s Disease Subjects (n = 6) and Matched Controls (n = 6) | ||||
|---|---|---|---|---|
| Group | Gender (M/F) | Age (years) | PMI (h) | pH |
| PD | 4M/2F | 71.3 ± 13.1 | 6.8 ± 3.1 | 6.3 ± 0.1 |
| HC | 4M/2F | 72.5 ± 9.2 | 5.1 ± 1.5 | 6.2 ± 0.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Dueñas, V.; Gómez-Soler, M.; Valle-León, M.; Watanabe, M.; Ferrer, I.; Ciruela, F. Revealing Adenosine A2A-Dopamine D2 Receptor Heteromers in Parkinson’s Disease Post-Mortem Brain through a New AlphaScreen-Based Assay. Int. J. Mol. Sci. 2019, 20, 3600. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143600
Fernández-Dueñas V, Gómez-Soler M, Valle-León M, Watanabe M, Ferrer I, Ciruela F. Revealing Adenosine A2A-Dopamine D2 Receptor Heteromers in Parkinson’s Disease Post-Mortem Brain through a New AlphaScreen-Based Assay. International Journal of Molecular Sciences. 2019; 20(14):3600. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143600
Chicago/Turabian StyleFernández-Dueñas, Víctor, Maricel Gómez-Soler, Marta Valle-León, Masahiko Watanabe, Isidre Ferrer, and Francisco Ciruela. 2019. "Revealing Adenosine A2A-Dopamine D2 Receptor Heteromers in Parkinson’s Disease Post-Mortem Brain through a New AlphaScreen-Based Assay" International Journal of Molecular Sciences 20, no. 14: 3600. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143600