Resveratrol Plays a Protective Role against Premature Ovarian Failure and Prompts Female Germline Stem Cell Survival


Abstract
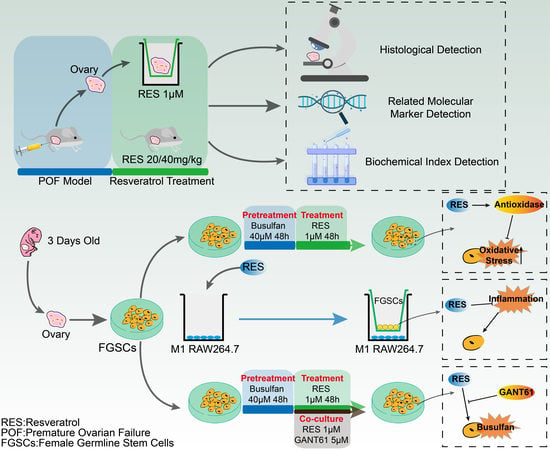
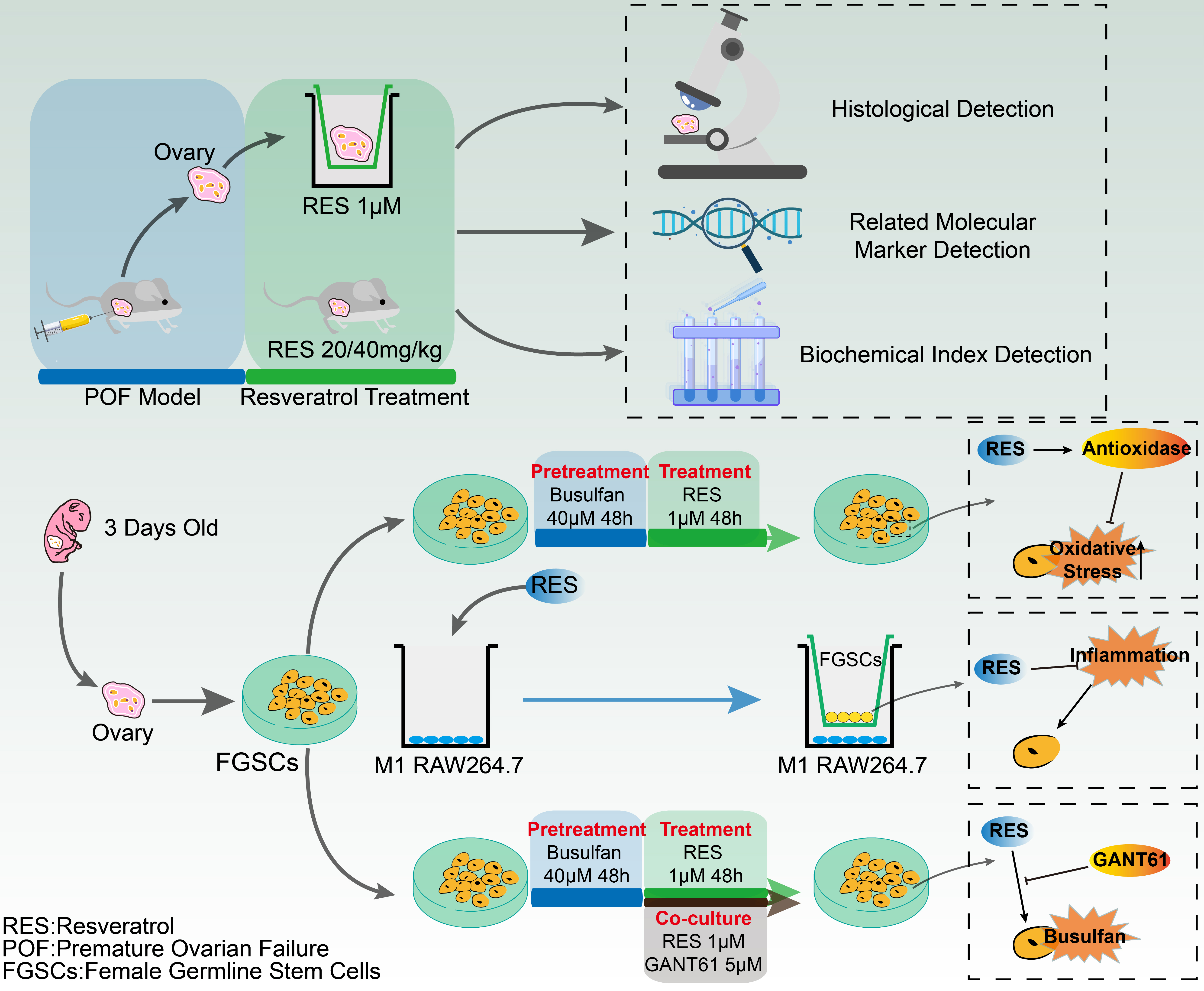
:
1. Introduction
2. Results
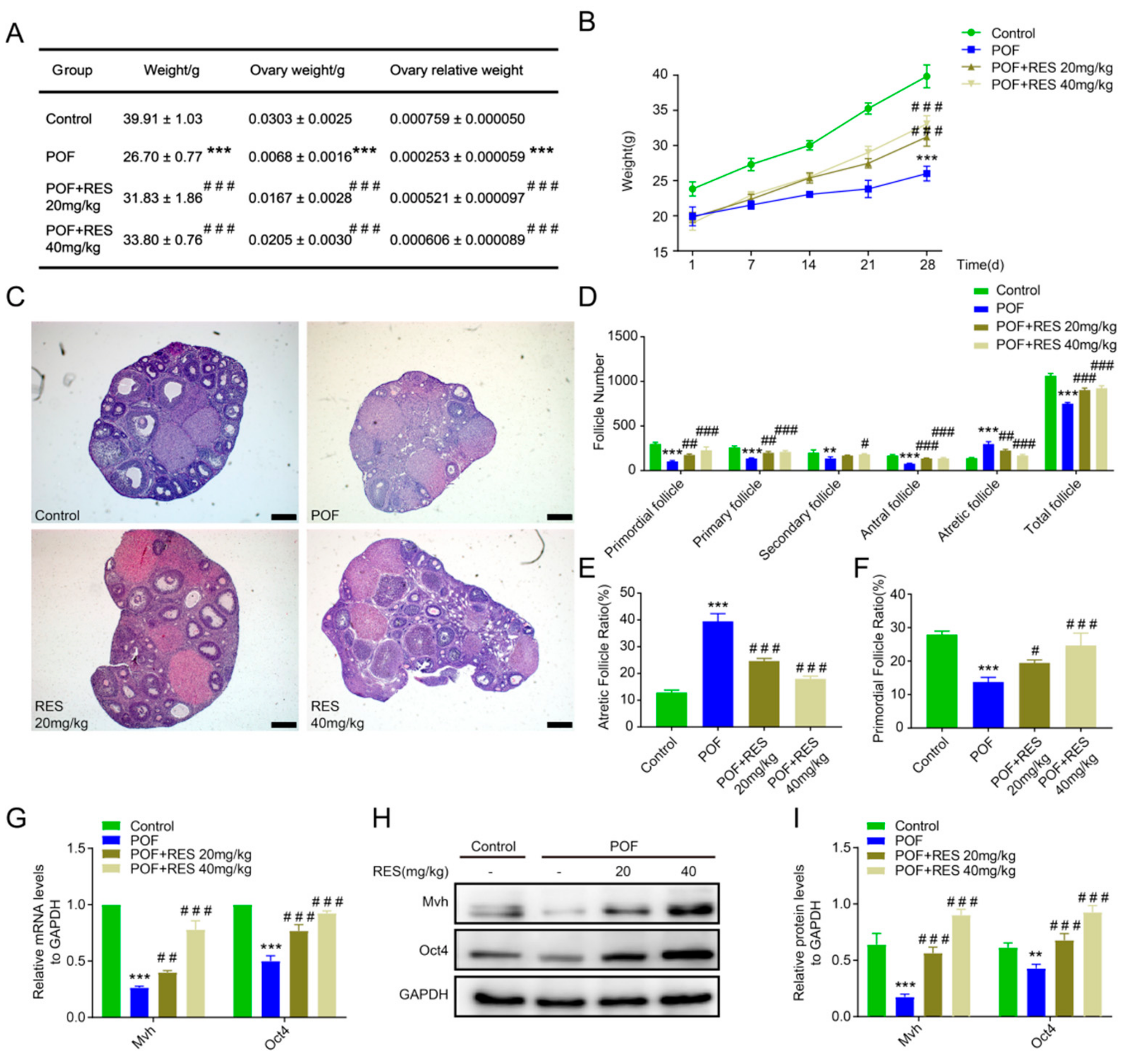
2.1. Protective Function of Resveratrol on the Follicle Number and Development in a Cyclophosphamide-Induced POF Model
2.2. Resveratrol Treatment Relieves Oxidative Stress and Benefits Germline Cells in POF Ovaries
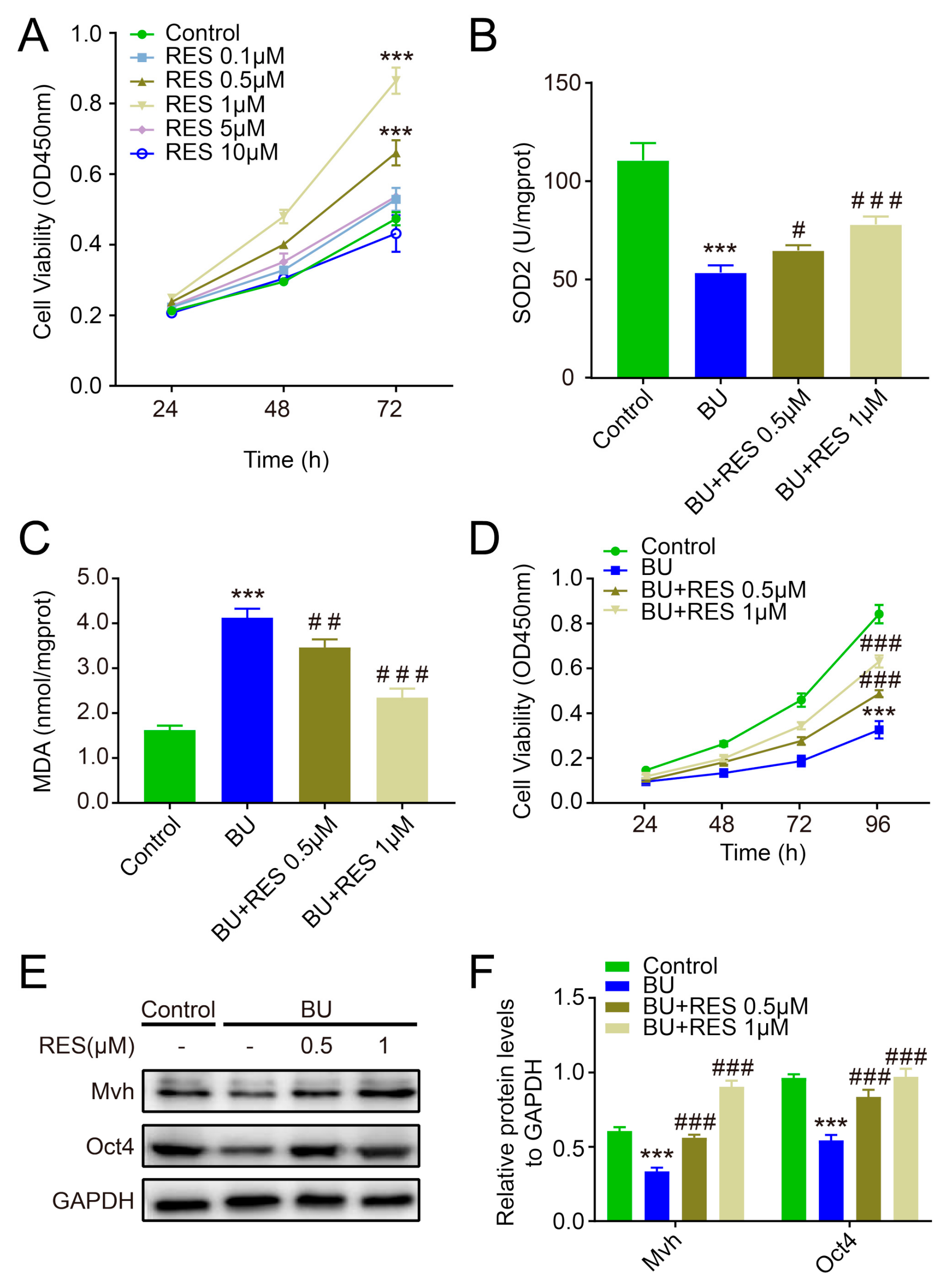
2.3. Resveratrol Enhances the Antioxidative Capacity of FGSCs and Improves Cell Survival
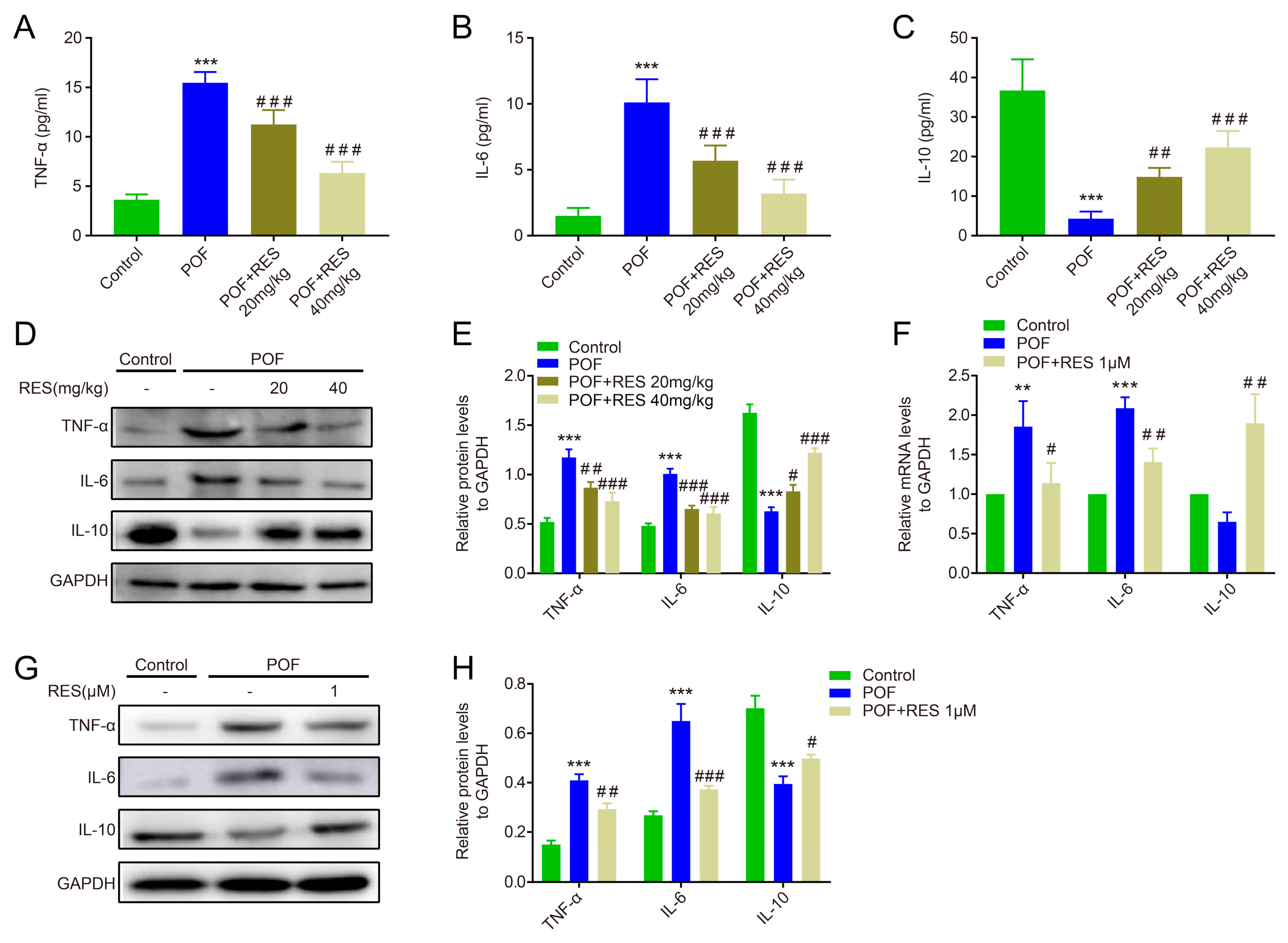
2.4. Effects of Resveratrol against Abnormal Inflammation in POF Mice
2.5. Resveratrol Improves the Inflammatory Reaction of M1 Macrophages and Promotes FGSC Survival
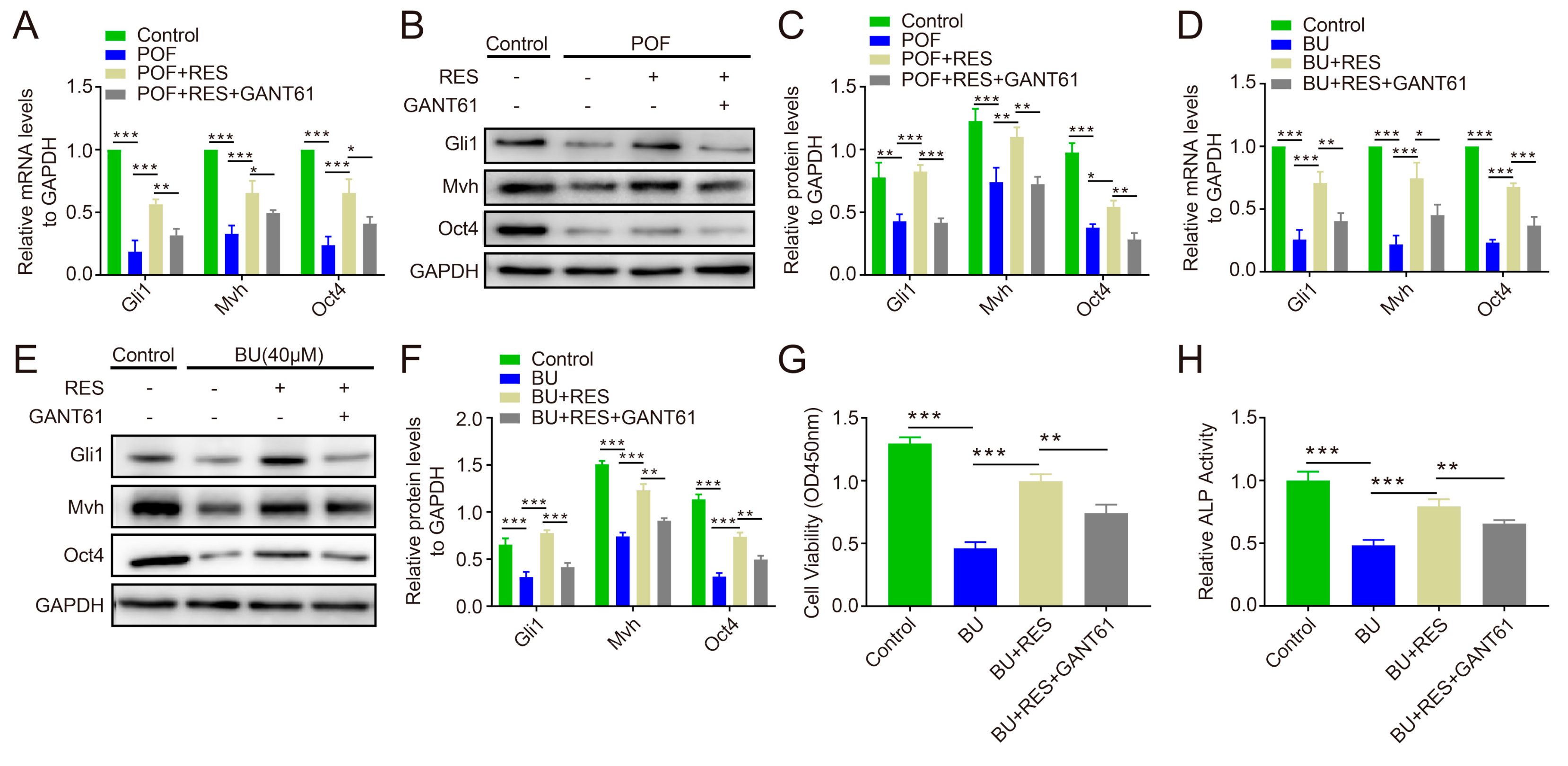
2.6. Blocking Hedgehog Pathway Activity Reverses the Protective Effect of Resveratrol on Ovarian Function and FGSC Survival
3. Discussion
4. Materials and Methods
4.1. Animals and Treatment
4.2. Ovary Extraction and Culture
4.3. HE Staining and Follicle Counting
4.4. Sample Preparation and Biochemical Analysis
4.5. Primary Isolation and Culture of FGSCs
4.6. Cell Treatment and Proliferation Activity Assay
4.7. ALP enzyme Activity Assay
4.8. Quantitative Real-Time PCR
4.9. Western Blotting
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| POF | Premature ovarian failure |
| POI | Premature ovarian insufficiency |
| CTX | Cyclophosphamide |
| BU | Busulfan |
| FSGC | Female germline stem cell |
| ESC | Embryonic stem cell |
| ROS | Reactive oxygen species |
| MSC | Mesenchymal stem cells |
| Mvh | Mouse vasa homologue |
| RES | Resveratrol |
| GPx | Glutathione peroxidase |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| GSH | Glutathione |
| MDA | Malondialdehyde |
| Hh | Hedgehog |
| TNF-α | Tumor necrosis factor α |
References
- Sukur, Y.E.; Kivancli, I.B.; Ozmen, B. Ovarian aging and premature ovarian failure. J. Turk. Ger. Gynecol. Assoc. 2014, 15, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Rudnicka, E.; Kruszewska, J.; Klicka, K.; Kowalczyk, J.; Grymowicz, M.; Skorska, J.; Pieta, W.; Smolarczyk, R. Premature ovarian insufficiency-aetiopathology, epidemiology, and diagnostic evaluation. Prz. Menopauzalny 2018, 17, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Sklar, C.A.; Mertens, A.C.; Mitby, P.; Whitton, J.; Stovall, M.; Kasper, C.; Mulder, J.; Green, D.; Nicholson, H.S.; Yasui, Y.; et al. Premature menopause in survivors of childhood cancer: A report from the childhood cancer survivor study. J. Natl. Cancer Inst. 2006, 98, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Ferrarini, E.; Russo, L.; Fruzzetti, F.; Agretti, P.; De Marco, G.; Dimida, A.; Gianetti, E.; Simoncini, T.; Simi, P.; Baldinotti, F.; et al. Clinical characteristics and genetic analysis in women with premature ovarian insufficiency. Maturitas 2013, 74, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.N.; Zhang, C.J.; Diao, H.L.; Zhang, Y. Protective Effects of Curcumin against Sodium Arsenite-induced Ovarian Oxidative Injury in a Mouse Model. Chin. Med. J. 2017, 130, 1026–1032. [Google Scholar] [CrossRef] [PubMed]
- Yin, N.; Zhao, W.; Luo, Q.; Yuan, W.; Luan, X.; Zhang, H. Restoring Ovarian Function with Human Placenta-Derived Mesenchymal Stem Cells in Autoimmune-Induced Premature Ovarian Failure Mice Mediated by Treg Cells and Associated Cytokines. Reprod. Sci. 2018, 25, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Bedoschi, G.; Navarro, P.A.; Oktay, K. Chemotherapy-induced damage to ovary: Mechanisms and clinical impact. Future Oncol. 2016, 12, 2333–2344. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Akbari Asbagh, F. Pathogenesis and causes of premature ovarian failure: An update. Int. J. Fertil. Steril. 2011, 5, 54–65. [Google Scholar]
- Melekoglu, R.; Ciftci, O.; Eraslan, S.; Cetin, A.; Basak, N. Beneficial effects of curcumin and capsaicin on cyclophosphamide-induced premature ovarian failure in a rat model. J. Ovarian Res. 2018, 11, 33. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, M.; Yao, X.; Li, T.; Wang, Q.; Lai, D. Human amniotic epithelial cells inhibit granulosa cell apoptosis induced by chemotherapy and restore the fertility. Stem Cell Res. 2015, 6, 152. [Google Scholar] [CrossRef]
- Zou, K.; Yuan, Z.; Yang, Z.; Luo, H.; Sun, K.; Zhou, L.; Xiang, J.; Shi, L.; Yu, Q.; Zhang, Y.; et al. Production of offspring from a germline stem cell line derived from neonatal ovaries. Nat. Cell Biol. 2009, 11, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Bukovsky, A.; Caudle, M.R. Immunoregulation of follicular renewal, selection, POF, and menopause in vivo, vs. neo-oogenesis in vitro, POF and ovarian infertility treatment, and a clinical trial. Reprod. Biol. Endocrinol. 2012, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Wu, F.; Xue, E.; Huang, L.; Yan, P.; Pan, X.; Zhou, Y. Hypoxia preconditioning promotes bone marrow mesenchymal stem cells survival by inducing HIF-1alpha in injured neuronal cells derived exosomes culture system. Cell Death Dis. 2019, 10, 134. [Google Scholar] [CrossRef] [PubMed]
- Agrahari, G.; Sah, S.K.; Kim, T.Y. Superoxide dismutase 3 protects mesenchymal stem cells through enhanced autophagy and regulation of FoxO3a trafficking. BMB Rep. 2018, 51, 344–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.L.; Chakraborty, S.; Rajan, S.S.; Wang, R.; Huang, F. Effects of oxidative stress on mouse embryonic stem cell proliferation, apoptosis, senescence, and self-renewal. Stem Cells Dev. 2010, 19, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Huang, J.; Zhang, H.; Wang, Y.; Matesic, L.E.; Takahata, M.; Awad, H.; Chen, D.; Xing, L. Tumor necrosis factor inhibits mesenchymal stem cell differentiation into osteoblasts via the ubiquitin E3 ligase Wwp1. Stem Cells 2011, 29, 1601–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhao, Y.; Liu, Y.; Akiyama, K.; Chen, C.; Qu, C.; Jin, Y.; Shi, S. IFN-gamma and TNF-alpha synergistically induce mesenchymal stem cell impairment and tumorigenesis via NFkappaB signaling. Stem Cells 2013, 31, 1383–1395. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A. Cancer prevention and treatment with resveratrol: From rodent studies to clinical trials. Cancer Prev. Res. 2009, 2, 409–418. [Google Scholar] [CrossRef]
- Poulsen, M.M.; Fjeldborg, K.; Ornstrup, M.J.; Kjaer, T.N.; Nohr, M.K.; Pedersen, S.B. Resveratrol and inflammation: Challenges in translating pre-clinical findings to improved patient outcomes. Biochim. Biophys. Acta 2015, 1852, 1124–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwachukwu, J.C.; Srinivasan, S.; Bruno, N.E.; Parent, A.A.; Hughes, T.S.; Pollock, J.A.; Gjyshi, O.; Cavett, V.; Nowak, J.; Garcia-Ordonez, R.D.; et al. Resveratrol modulates the inflammatory response via an estrogen receptor-signal integration network. Elife 2014, 3, e02057. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Daiber, A.; Forstermann, U.; Li, H. Antioxidant effects of resveratrol in the cardiovascular system. Br. J. Pharmacol. 2017, 174, 1633–1646. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.; Bath, S.; Elbarbry, F. An Organ System Approach to Explore the Antioxidative, Anti-Inflammatory, and Cytoprotective Actions of Resveratrol. Oxidative Med. Cell. Longev. 2015, 2015, 803971. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Jiang, X.; Lai, Y.; Liu, Y.; Zhang, Z. Resveratrol protects against arsenic trioxide-induced oxidative damage through maintenance of glutathione homeostasis and inhibition of apoptotic progression. Environ. Mol. Mutagen. 2015, 56, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Akbari Asbagh, F. The role of autoimmunity in premature ovarian failure. Iran. J. Reprod. Med. 2015, 13, 461–472. [Google Scholar] [PubMed]
- Semba, R.D.; Nicklett, E.J.; Ferrucci, L. Does accumulation of advanced glycation end products contribute to the aging phenotype? J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65, 963–975. [Google Scholar] [CrossRef]
- Park, E.J.; Pezzuto, J.M. The pharmacology of resveratrol in animals and humans. Biochim. Biophys. Acta 2015, 1852, 1071–1113. [Google Scholar] [CrossRef] [Green Version]
- Chinwe, G.S.; Azuka, O.I.; Adaeze, N.C. Resveratrol supplementation rescues pool of growing follicles and ovarian stroma from Cisplatin-induced toxicity on the ovary in Sprague-Dawley rats: An experimental study. Int. J. Reprod. Biomed. 2018, 16, 19–30. [Google Scholar]
- Furat Rencber, S.; Kurnaz Ozbek, S.; Eraldemir, C.; Sezer, Z.; Kum, T.; Ceylan, S.; Guzel, E. Effect of resveratrol and metformin on ovarian reserve and ultrastructure in PCOS: An experimental study. J. Ovarian Res. 2018, 11, 55. [Google Scholar] [CrossRef]
- Hadjzadeh, M.A.R.; Alikhani, V.; Hosseinian, S.; Zarei, B.; Keshavarzi, Z. The Effect of Melatonin against Gastric Oxidative Stress and Dyslipidemia in Streptozotocin-Induced Diabetic Rats. Acta Endocrinol. Buchar. 2018, 14, 453–458. [Google Scholar] [CrossRef] [PubMed]
- De Freitas Carvalho, M.M.; Lage, N.N.; de Souza Paulino, A.H.; Pereira, R.R.; de Almeida, L.T.; da Silva, T.F.; de Brito Magalhaes, C.L.; de Lima, W.G.; Silva, M.E.; Pedrosa, M.L.; et al. Effects of acai on oxidative stress, ER stress, and inflammation-related parameters in mice with high fat diet-fed induced NAFLD. Sci. Rep. 2019, 9, 8107. [Google Scholar] [CrossRef] [PubMed]
- Niringiyumukiza, J.D.; Cai, H.; Chen, L.; Li, Y.; Wang, L.; Zhang, M.; Xu, X.; Xiang, W. Protective properties of glycogen synthase kinase-3 inhibition against doxorubicin-induced oxidative damage to mouse ovarian reserve. Biomed. Pharmacother. 2019, 116, 108963. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Ma, W.; Huang, C.; Ding, J.; Cui, D.; Zhang, M. Expression Pattern of Mouse Vasa Homologue (MVH) in the Ovaries of C57BL/6 Female Mice. Med. Sci. Monit. 2016, 22, 2656–2663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Kajiura-Kobayashi, H.; Nagahama, Y. Differential expression of vasa homologue gene in the germ cells during oogenesis and spermatogenesis in a teleost fish, tilapia, Oreochromis niloticus. Mech. Dev. 2000, 99, 139–142. [Google Scholar] [CrossRef]
- Noce, T.; Okamoto-Ito, S.; Tsunekawa, N. Vasa homolog genes in mammalian germ cell development. Cell Struct. Funct. 2001, 26, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Pesce, M.; Scholer, H.R. Oct-4: Control of totipotency and germline determination. Mol. Reprod. Dev. 2000, 55, 452–457. [Google Scholar] [CrossRef]
- Chahvar, S.T.; Al-Shawaf, T.; Tranquilli, A.L. Pharmacologic ovarian preservation in young women undergoing chemotherapy. Curr. Med. Chem. 2014, 21, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Banu, S.K.; Stanley, J.A.; Sivakumar, K.K.; Arosh, J.A.; Burghardt, R.C. Resveratrol protects the ovary against chromium-toxicity by enhancing endogenous antioxidant enzymes and inhibiting metabolic clearance of estradiol. Toxicol. Appl. Pharmacol. 2016, 303, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Huminiecki, L.; Horbanczuk, J. The functional genomic studies of resveratrol in respect to its anti-cancer effects. Biotechnol. Adv. 2018, 36, 1699–1708. [Google Scholar] [CrossRef]
- Ozturk, Y.; Gunaydin, C.; Yalcin, F.; Naziroglu, M.; Braidy, N. Resveratrol Enhances Apoptotic and Oxidant Effects of Paclitaxel through TRPM2 Channel Activation in DBTRG Glioblastoma Cells. Oxidative Med. Cell. Longev. 2019, 2019, 13. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.D.; Yang, J.L.; Zhang, W.L.; Liu, D.X. Resveratrol inhibits oral squamous cell carcinoma through induction of apoptosis and G2/M phase cell cycle arrest. Tumor Biol. 2016, 37, 2871–2877. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, M.E.S.; Gouveia, B.B.; Barberino, R.S.; Menezes, V.G.; Macedo, T.J.S.; Cavalcante, A.Y.P.; Monte, A.P.O.; Santos, J.M.S.; Matos, M.H.T. Resveratrol promotes in vitro activation of ovine primordial follicles by reducing DNA damage and enhancing granulosa cell proliferation via phosphatidylinositol 3-kinase pathway. Reprod. Domest. Anim. 2018, 53, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, L. Mechanism of resveratrol in improving ovarian function in a rat model of premature ovarian insufficiency. J. Obstet. Gynaecol. Res. 2018, 44, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wei, R.; Sun, Y.; Xia, Q.; Xie, W.; Song, H.; Wang, W.; Zou, K. AKT3 Is a Pivotal Molecule of Cadherin-22 and GDNF Family Receptor-alpha1 Signal Pathways Regulating Self-Renewal in Female Germline Stem Cells. Stem Cells 2019. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Lee, T.L.; Hu, B.; Li, J.; Li, X.; Zhao, X.; Hou, C.; Zhang, C.; He, L.; Huang, X.; et al. Molecular characteristics of early-stage female germ cells revealed by RNA sequencing of low-input cells and analysis of genome-wide DNA methylation. DNA Res. 2019, 26, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.J.; Yeh, Y.C.; Shang, W.J.; Wu, G.J.; Liu, J.Y.; Chen, C.H. Protective effect of a gonadotropin-releasing hormone analogue on chemotherapeutic agent-induced ovarian gonadotoxicity: A mouse model. Eur. J. Obstet. Gynecol. Reprod. Biol. 2010, 149, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ran, L.; Zhang, W.; Leng, X.; Wang, K.; Liu, G.; Song, J.; Wang, Y.; Zhang, X.; Wang, Y.; et al. FOXS1 is regulated by GLI1 and miR-125a-5p and promotes cell proliferation and EMT in gastric cancer. Sci. Rep. 2019, 9, 5281. [Google Scholar] [CrossRef] [PubMed]
- Riaz, S.K.; Ke, Y.; Wang, F.; Kayani, M.A.; Malik, M.F.A. Influence of SHH/GLI1 axis on EMT mediated migration and invasion of breast cancer cells. Sci. Rep. 2019, 9, 6620. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Yin, N.; Zhang, L.; Yuan, W.; Zhao, W.; Luan, X.; Zhang, H. Role of SDF-1/CXCR4 and cytokines in the development of ovary injury in chemotherapy drug induced premature ovarian failure mice. Life Sci. 2017, 179, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Park, M.R.; Choi, Y.J.; Kwon, D.N.; Park, C.; Bui, H.T.; Gurunathan, S.; Cho, S.G.; Song, H.; Seo, H.G.; Min, G.; et al. Intraovarian transplantation of primordial follicles fails to rescue chemotherapy injured ovaries. Sci. Rep. 2013, 3, 1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.G.; Luo, L.L.; Xu, J.J.; Zhuang, X.L.; Kong, X.X.; Fu, Y.C. Effects of plant polyphenols on ovarian follicular reserve in aging rats. Biochem. Cell Biol. 2010, 88, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Gonnissen, A.; Isebaert, S.; McKee, C.M.; Muschel, R.J.; Haustermans, K. The Effect of Metformin and GANT61 Combinations on the Radiosensitivity of Prostate Cancer Cells. Int. J. Mol. Sci. 2017, 18, 339. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′-3′) | Gene Number |
|---|---|---|
| Mvh | Forward:GTGTATTATTGTAGCACCAACTCG Reverse:CACCCTTGTACTATCTGTCGAACT | NM_001145885.1 |
| Oct4 | Forward: AGCTGCTGAAGCAGAAGAGG Reverse: GGTTCTCATTGTTGTCGGCT | NM_013633.3 |
| GPx | Forward: GGCTCATCTGAGCAACAAGG Reverse: CGCCGATGTTCAAGGTCAAT | NM_008160.6 |
| SOD2 | Forward: ACAACTCAGGTCGCTCTTCA Reverse: GAACCTTGGACTCCCACAGA | NM_013671.3 |
| CAT | Forward: ACATGGTCTGGGACTTCTGG Reverse: ACTGCCTCTCCATCTGCATT | NM_009804.2 |
| TNF-α | Forward: CTCATGCACCACCATCAAGG Reverse: ACCTGACCACTCTCCCTTTG | NM_013693.3 |
| IL-6 | Forward: GACTGATGCTGGTGACAACC Reverse: AGACAGGTCTGTTGGGAGTG | NM_031168.2 |
| IL-10 | Forward: AGTACAGCCGGGAAGACAAT Reverse: TCTAGGAGCATGTGGCTCTG | NM_010548.2 |
| Gli1 | Forward: CCCAATACATGCTGGTGGTG Reverse: GCAACCTTCTTGCTCACACA | NM_010296.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Zhang, Z.; Cha, L.; Li, L.; Zhu, D.; Fang, Z.; He, Z.; Huang, J.; Pan, Z. Resveratrol Plays a Protective Role against Premature Ovarian Failure and Prompts Female Germline Stem Cell Survival. Int. J. Mol. Sci. 2019, 20, 3605. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143605
Jiang Y, Zhang Z, Cha L, Li L, Zhu D, Fang Z, He Z, Huang J, Pan Z. Resveratrol Plays a Protective Role against Premature Ovarian Failure and Prompts Female Germline Stem Cell Survival. International Journal of Molecular Sciences. 2019; 20(14):3605. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143605
Chicago/Turabian StyleJiang, Yu, Zhaoyuan Zhang, Lijun Cha, Lili Li, Dantian Zhu, Zhi Fang, Zhiqiang He, Jian Huang, and Zezheng Pan. 2019. "Resveratrol Plays a Protective Role against Premature Ovarian Failure and Prompts Female Germline Stem Cell Survival" International Journal of Molecular Sciences 20, no. 14: 3605. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143605