Kinetics of Flavoenzyme-Catalyzed Reduction of Tirapazamine Derivatives: Implications for Their Prooxidant Cytotoxicity

 , ,
, ,
Abstract
:1. Introduction
2. Results
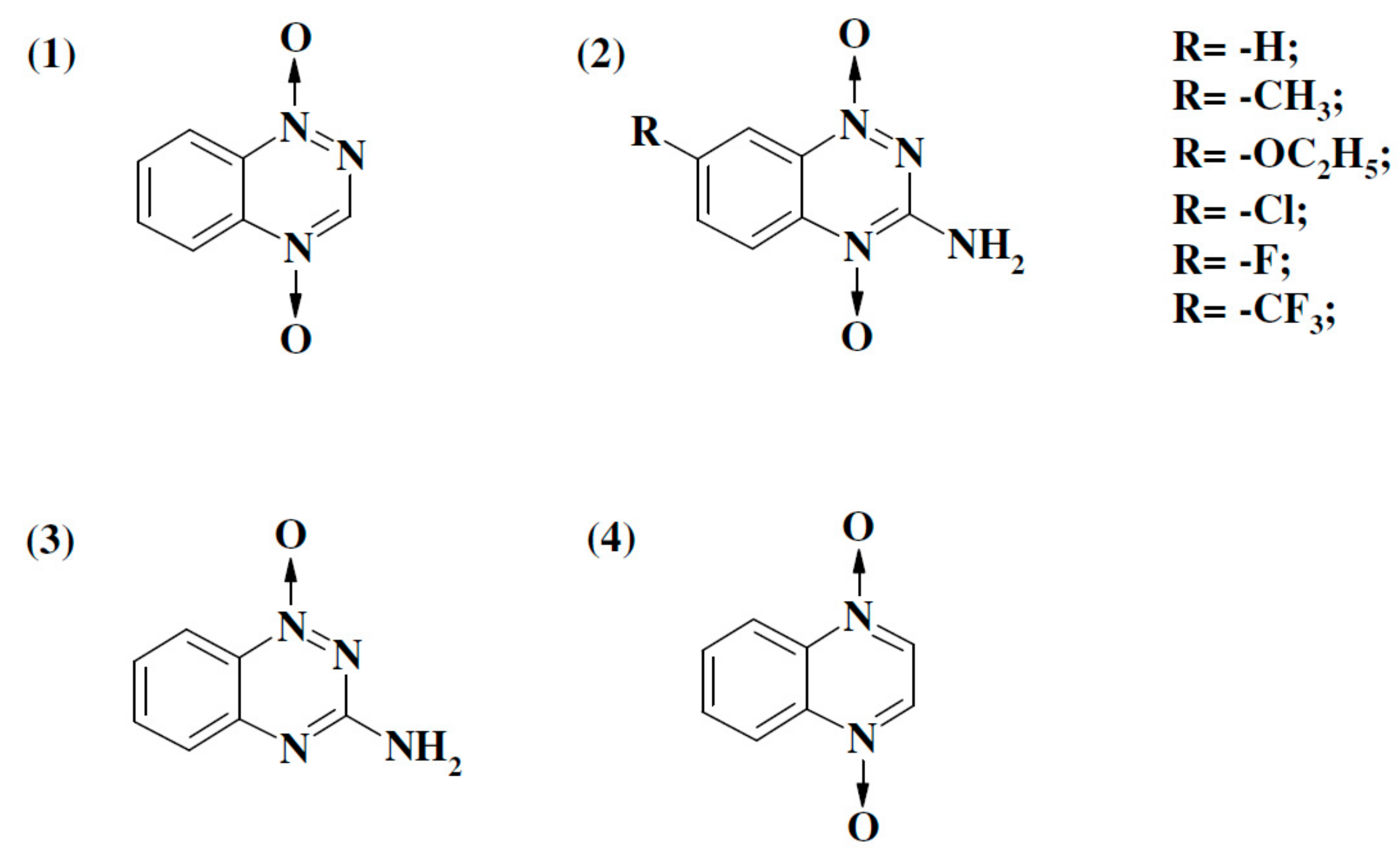
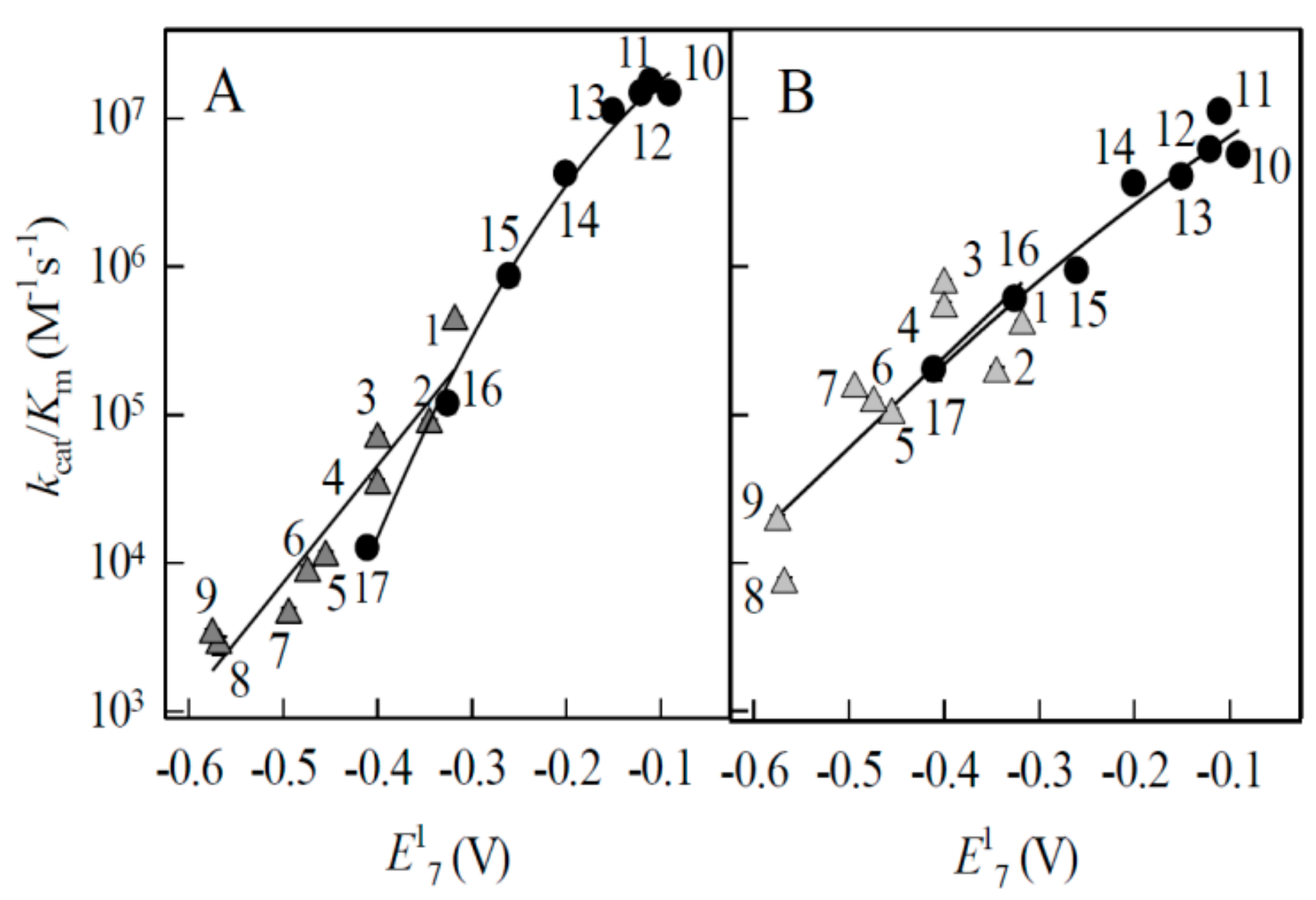
2.1. Enzymatic Single-Electron Reduction of Aromatic N-Oxides
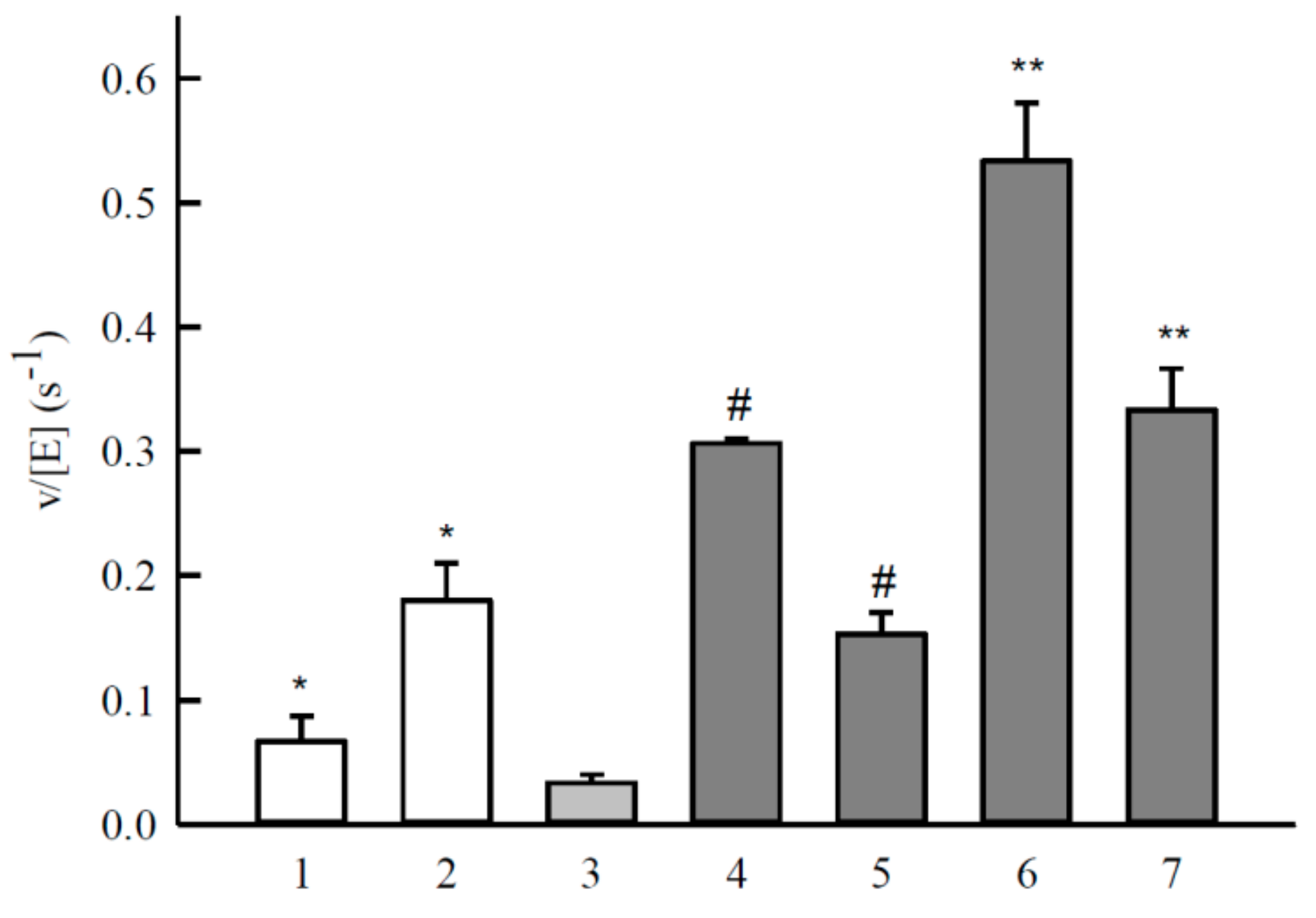
2.2. Studies of NQO1-Catalyzed Reduction of Aromatic N-Oxides
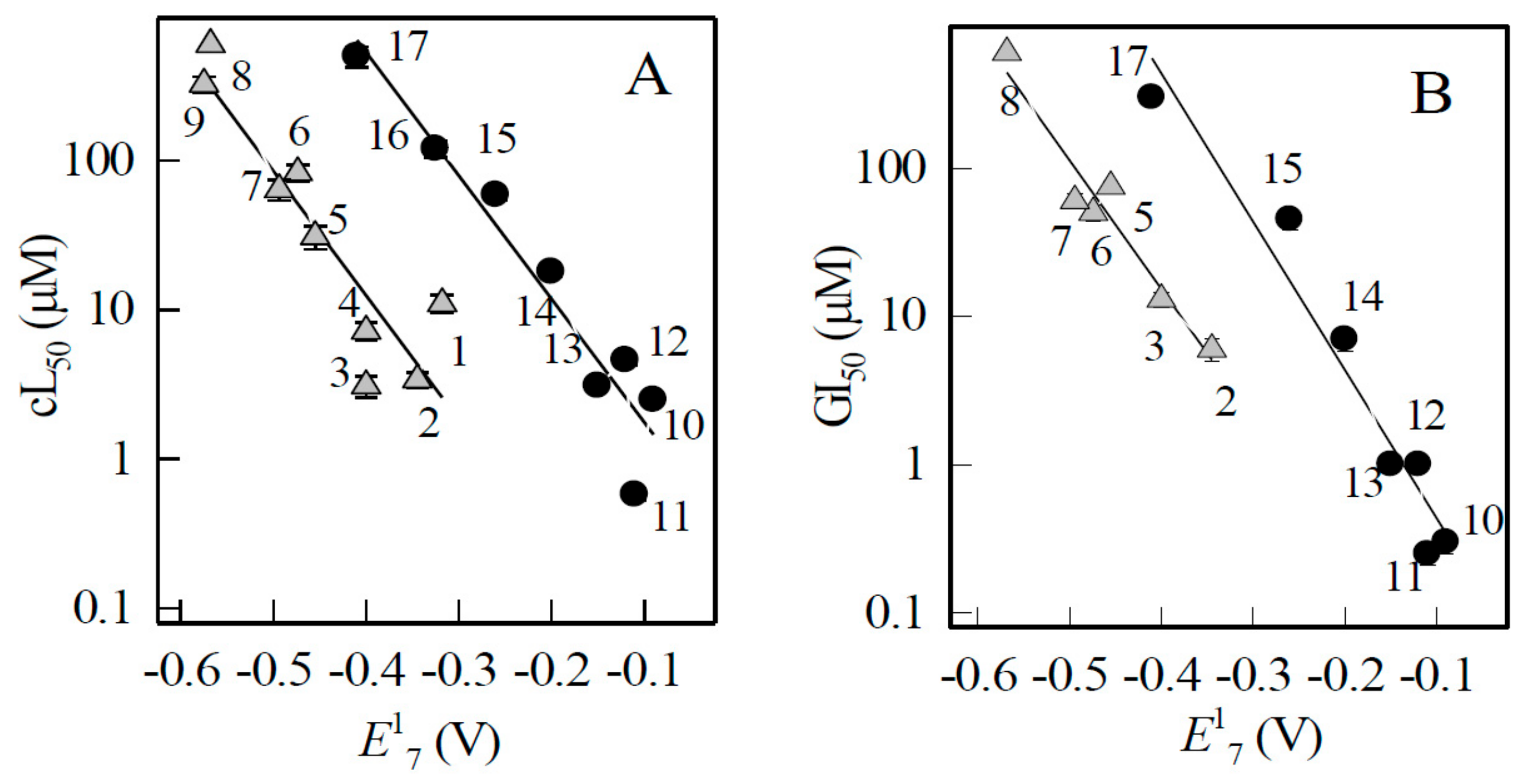
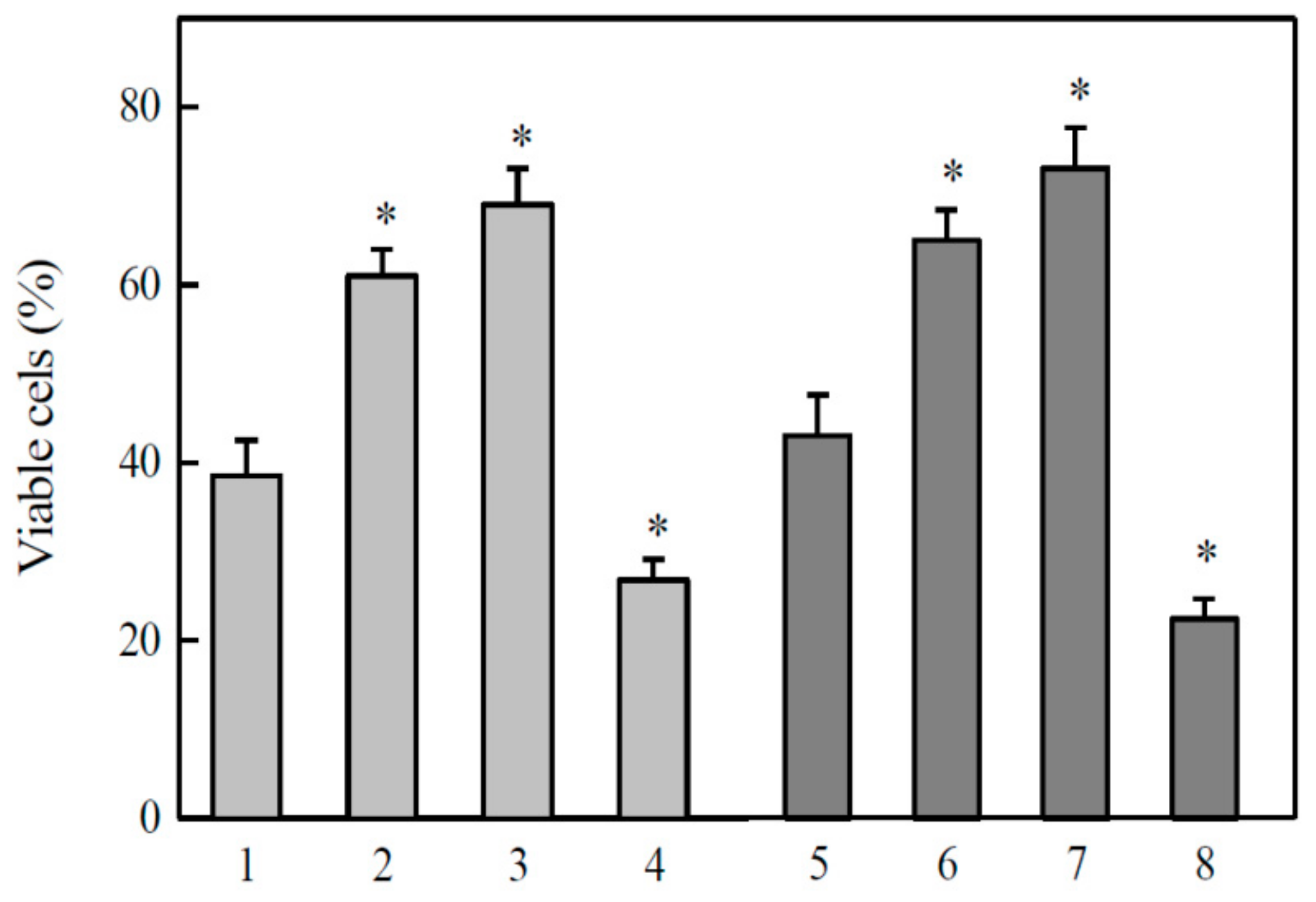
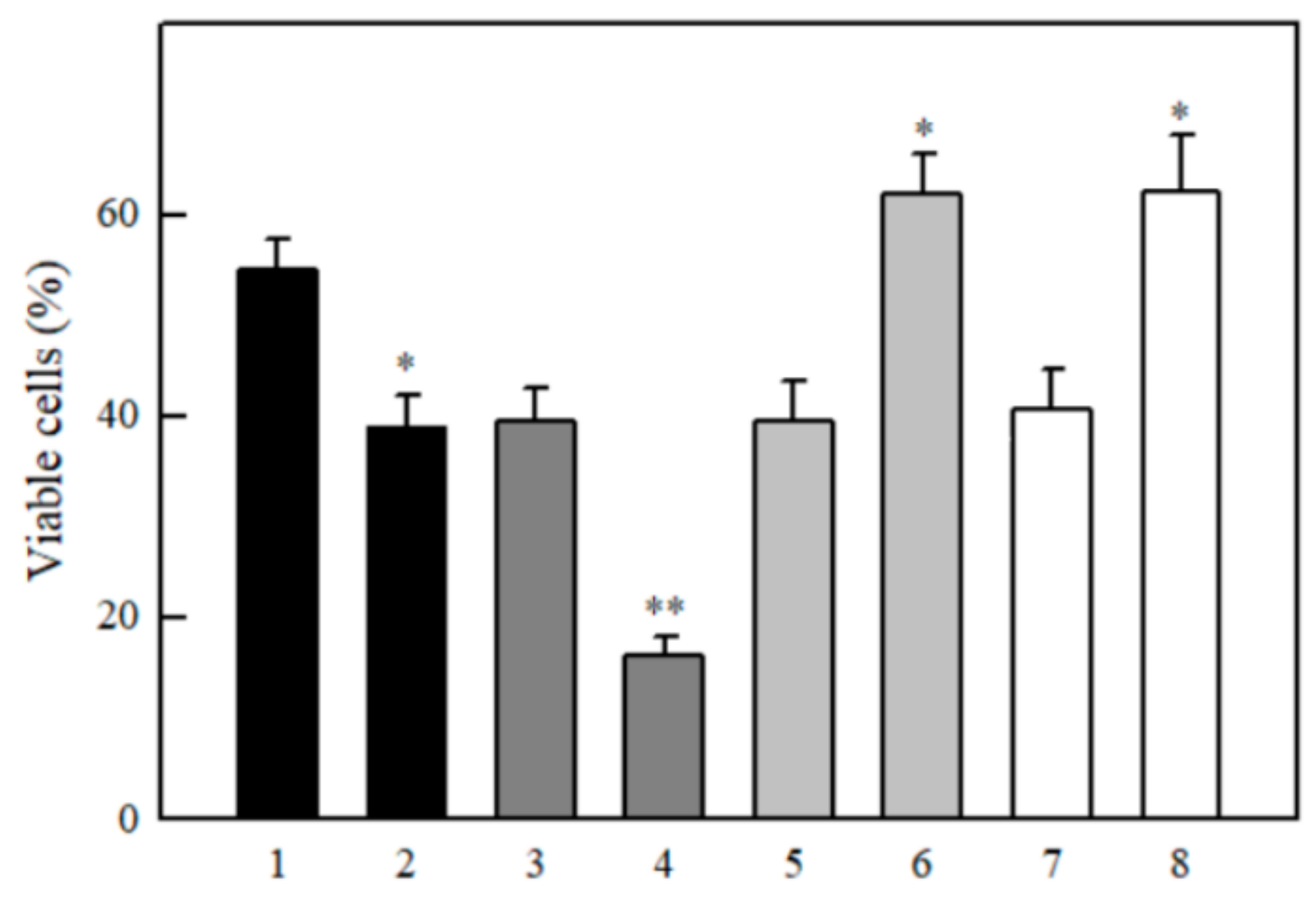
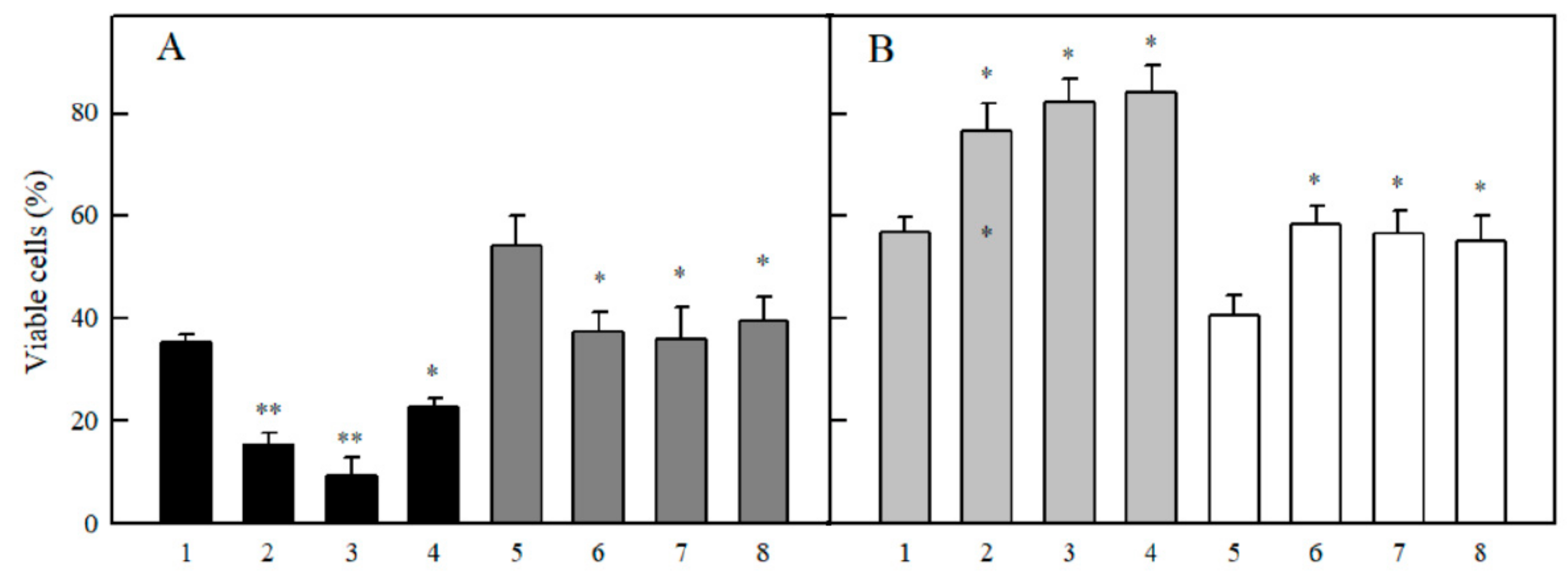
2.3. Studies of Cytotoxicity of Aromatic N-oxides
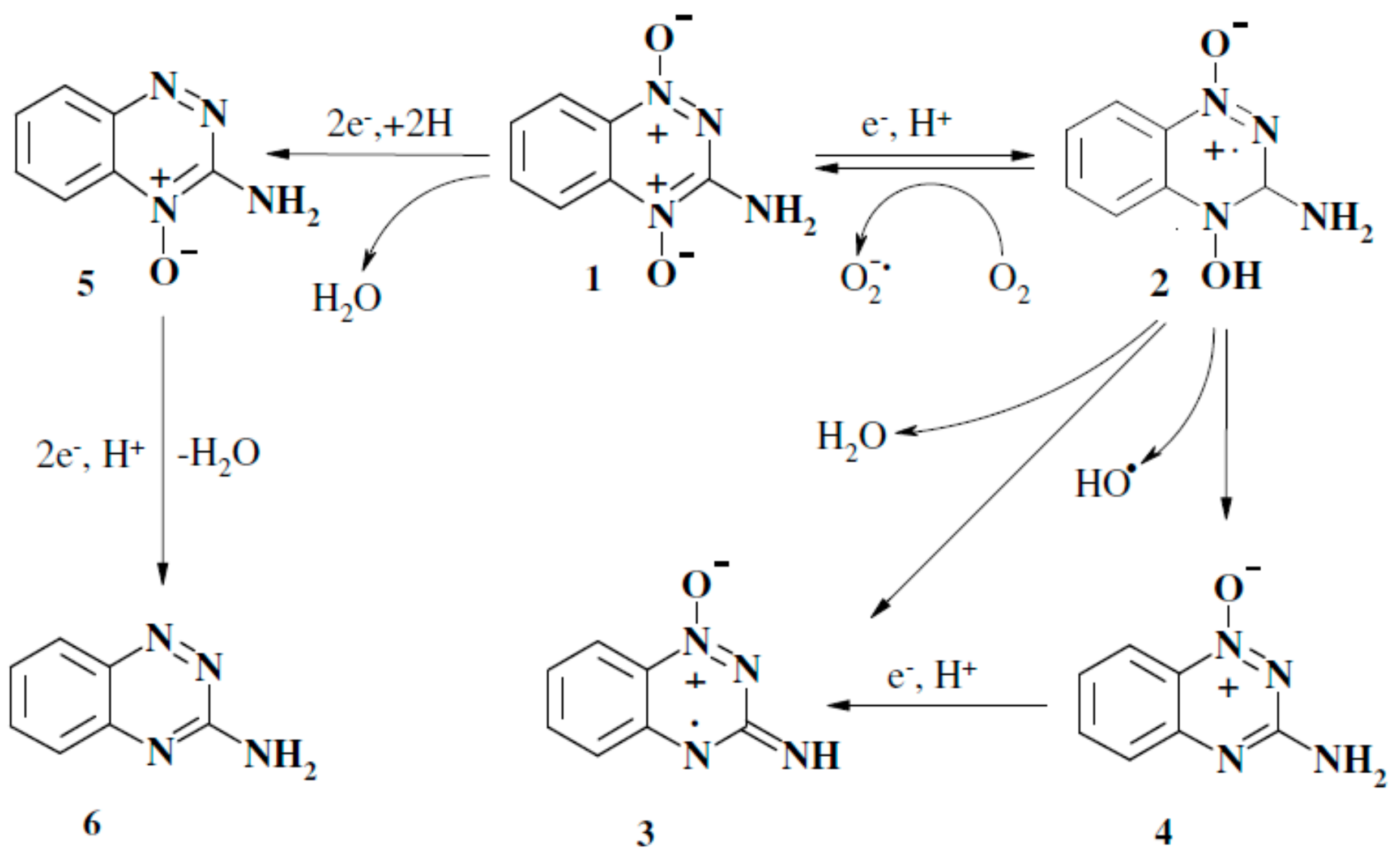
3. Discussion
4. Materials and Methods
4.1. Enzymes and Chemicals
4.2. Enzymatic Assays
4.3. Cytotoxicity Assays
4.4. Statistical Analysis and Calculations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADR | NADPH:adrenodoxin reductase |
| ADX | Adrenodoxin |
| ArN→O | Heteroaromatic N-oxide |
| BCNU | 1,3-bis(2-chloroethyl)-1-nitrosourea |
| cL50 | Concentration for 50% cell survival |
| DPPD | N,N’-diphenyl-p-phenylene diamine |
| E17 | Single-electron reduction midpoint potential |
| GI50 | Concentration for 50% inhibition of maximal cell proliferation |
| kcat | Catalytic constant |
| kcat/Km | Bimolecular rate constant |
| LD50 | Median lethal dose |
| log D | Octanol/water distribution coefficient at pH 7.0 |
| NQO1 | NAD(P)H:quinone oxidoreductase |
| P-450 | Cytochrome P-450 |
| P-450R | NADPH:cytochrome P-450 reductase |
| TPZ | Tirapazamine |
Appendix A
References
- Wardman, P.; Dennis, M.F.; Everett, S.A.; Patel, K.B.; Stratford, M.R.L.; Tracy, M. Radicals from one-electron reduction of nitro compounds, aromatic N-oxides and quinones: The kinetic basis for hypoxia-selective, bioreductive drugs. Biochem. Soc. Symp. 1995, 61, 171–194. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Gates, K.S. Enzyme-activated generation of reactive oxygen species from heterocyclic N-oxides under aerobic and anaerobic conditions and its relevance to hypoxia-selective prodrugs. Chem. Res. Toxicol. 2019, 32, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Rajapakse, A.; Gallazzi, F.; Junnotula, V.; Fuchs-Knotts, T.; Glaser, R.; Gates, K.S. Isotopic labeling experiments that elucidate the mechanism of DNA strand cleavage by the hypoxia-selective antitumor agent 1,2,4-benzotriazine 1,4-di-N-oxide. Chem. Res. Toxicol. 2013, 22, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Shinde, S.S.; Maroz, A.; Hay, M.P.; Patterson, A.V.; Denny, V.A.; Anderson, R.F. Characterization of radicals formed following enzymatic reduction of 3-substituted analogues of the hypoxia-selective cytotoxin 3-amino-1,2,4-benzotriazine 1,4-dioxide (tirapazamine). J. Am. Chem. Soc. 2010, 132, 2591–2599. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.F.; Yadav, P.; Shinde, S.S.; Hong, C.R.; Pullen, S.M.; Reynisson, J.; Wilson, W.R.; Hay, M.P. Radical chemistry and cytotoxicity of bioreductive 3-substituted quinoxaline di-N-oxides. Chem. Res. Toxicol. 2016, 29, 1310–1324. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, T.; Chowdhury, G.; Fuchs, C.L.; Gates, K.S. 3-Amino-1,2,4-benzotriazine 4-oxide: Characterization of a new metabolite arising from bioreductive processing of the antitumour agent 3-amino-1,2,4-benzotriazine 1,4-dioxide (tirapazamine). J. Org. Chem. 2001, 66, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Hunter, F.W.; Young, R.J.; Shalev, Z.; Vellanki, R.N.; Wang, J.; Gu, Y.; Joshi, N.; Sreebhavan, S.; Weinreb, J.; Goldstein, D.P.; et al. Identification of P450 oxidoreductase as a major determinant of sensitivity to hypoxia-activated prodrugs. Cancer Res. 2015, 75, 4211–4223. [Google Scholar] [CrossRef] [PubMed]
- Delahoussaye, Y.M.; Evans, J.W.; Brown, J.M. Metabolism of tirapazamine by multiple reductases in the nucleus. Biochem. Pharmacol. 2001, 62, 1201–1209. [Google Scholar] [CrossRef]
- Cahill, A.; Jenkins, T.C.; White, I.N.H. Metabolism of 3-amino-1,2,4-benzotriazine-1,4-dioxide (SR 4233) by purified DT-diaphorase under aerobic and anaerobic conditions. Biochem. Pharmacol. 1993, 45, 321–329. [Google Scholar] [CrossRef]
- Elwell, J.H.; Siim, B.G.; Evans, J.W.; Brown, J.M. Adaptation of human tumor cells to tirapazamine under aerobic conditions. Implications of increased antioxidant enzyme activity to mechanism of aerobic cytotoxicity. Biochem. Pharmacol. 1997, 54, 249–257. [Google Scholar] [CrossRef]
- Khan, S.; O’Brien, P.J. Molecular mechanisms of tirapazamine (SR 4233, WIN 59075)-induced hepatocyte toxicity under low oxygen concentrations. Br. J. Cancer 1995, 71, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Walton, M.I.; Workman, P. Enzymology of the reductive bioactivation of SR 4233. A novel benzotriazine di-N-oxide hypoxic cell cytotoxin. Biochem. Pharmacol. 1990, 39, 1735–1742. [Google Scholar] [CrossRef]
- Riley, R.J.; Hemingway, S.A.; Graham, M.A.; Workman, P. Initial characterization of the major mouse cytochrome P450 enzymes involved in the reductive metabolism of the hypoxic cytotoxin 3-amino-1,2,4-benzotriazine-1,4-di-N-oxide (tirapazamine, SR 4233, WIN 59075). Biochem. Pharamacol. 1993, 45, 1065–1077. [Google Scholar] [CrossRef]
- Wang, J.; Biedermann, K.A.; Wolf, C.R.; Brown, J.M. Metabolism of the bioreductive cytotoxin SR 4233 by tumour cells: Enzymatic studies. Br. J. Cancer 1993, 67, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Cahill, A.; White, I.N.H. Reductive metabolism of 3-amino-1,2,4-benzotriazine-1,4-dioxide (SR 4233) and the induction of unscheduled DNA synthesis in rat and human derived cell lines. Carcinogenesis 1990, 11, 1407–1411. [Google Scholar] [CrossRef]
- Silva, J.M.; O’Brien, P.J. Molecular mechanisms of SR 4233-induced hepatocyte toxicity under aerobic versus hypoxic conditions. Br. J. Cancer 1993, 68, 484–491. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, R.V.; Duling, D.R.; Rumyantseva, G.V.; Mason, R.P.; Bridson, P.K. Microsomal reduction of 3-amino-1,2,4-benzotriazine 1,4-dioxide to its free radical. Mol. Pharmacol. 1991, 40, 440–445. [Google Scholar]
- Hay, M.P.; Gamage, S.A.; Kovacs, M.S.; Pruijn, F.B.; Anderson, R.F.; Patterson, A.V.; Wilson, W.R.; Brown, J.M.; Denny, W.A. Structure-activity relationships of 1,2,4-benzotriazine 1,4-dioxides as hypoxia-selective analogues of tirapazamine. J. Med. Chem. 2003, 46, 169–182. [Google Scholar] [CrossRef]
- Hicks, K.O.; Siim, B.G.; Jaiswal, J.K.; Pruijn, F.B.; Fraser, A.M.; Patel, R.; Hogg, A.; Liyanage, H.D.S.; Dorie, M.J.; Brown, J.M.; et al. Pharmacokinetic/pharmacodynamic modeling identifies SN30000 and SN29751 as tirapazamine analogues with improved tissue penetration and hypoxic cell killing in tumours. Clin. Cancer Res. 2010, 16, 4946–4957. [Google Scholar] [CrossRef]
- Gu, Y.; Chang, T.T.-A.; Wang, J.; Jaiswal, J.K.; Edwards, D.; Downes, N.J.; Liyanage, H.D.S.; Lynch, C.R.H.; Pruijn, F.B.; Hickey, A.J.R.; et al. Reductive metabolism influences the toxicity and pharmacokinetics of the hypoxia-targeted benzotriazine di-oxide anticancer agent SN30000 in mice. Front. Pharmacol. 2017, 8, 531. [Google Scholar] [CrossRef]
- Zarranz, B.; Jaso, A.; Aldana, I.; Monge, A. Synthesis and anticancer activity evaluation of new 2-alkylcarbonyl and 2-benzoyl-1-trifluoromethyl-quinoxaline-1,4-di-N-oxide derivatives. Bioorg. Med. Chem. 2004, 12, 3711–3721. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, G.; Sarkar, U.; Pullen, S.; Wilson, W.R.; Rajapakse, A.; Fuchs-Knotts, T.; Dates, K.S. DNA strand cleavage by the phenazine di-N-oxide natural product myxin under both aerobic and anaerobic conditions. Chem. Res. Toxicol. 2012, 12, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Vicente, E.; Lima, L.M.; Bongard, E.; Charnaud, S.; Villar, R.; Solano, B.; Burguete, A.; Perez-Silanes, S.; Aldana, I.; Vivas, L.; et al. Synthesis and structure-activity relationship of 3-phenylquinoxaline 1,4-di-N-oxide derivatives as antimalarial agents. Eur. J. Med. Chem. 2008, 43, 1903–1910. [Google Scholar] [CrossRef] [PubMed]
- Barea, C.; Pabon, A.; Perez-Silanes, S.; Galiano, S.; Gonzalez, G.; Monge, A.; Deharo, E.; Aldana, I. New amide derivatives of quinoxaline 1,4-di-N-oxide with leishmanicidal and antiplasmodial activities. Molecules 2013, 18, 4718–4727. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Sa, W.; Cao, C.; Guo, L.; Hao, H.; Liu, Z.; Wang, X.; Yuan, Z. Quinoxaline 1,4-di-N-oxides: Biological activities and mechanisms of action. Front. Pharmacol. 2016, 7, 64. [Google Scholar] [CrossRef]
- Marcus, R.A.; Sutin, N. Electron transfers in chemistry and biology. Biochim. Biophys. Acta 1985, 811, 265–322. [Google Scholar] [CrossRef]
- Anderson, R.F.; Shinde, S.; Hay, M.P.; Denny, W.A. Potentiation of the cytotoxicity of the anticancer agent tirapazamine by benzotriazine-N-oxides: The role of redox equilibria. J. Am. Chem. Soc. 2006, 128, 245–249. [Google Scholar] [CrossRef]
- Čėnas, N.K.; Marcinkevičienė, J.A.; Kulys, J.J.; Usanov, S.A. A negative cooperativity between NADPH and adrenodoxin on binding to NADPH:adrenodoxin reductase. FEBS Lett. 1990, 259, 338–340. [Google Scholar] [CrossRef] [Green Version]
- Marcinkevičienė, J.; Čėnas, N.; Kulys, J.; Usanov, S.A.; Sukhova, N.M.; Selezneva, I.S.; Gryazev, V.F. Nitroreductase reactions of the NADPH: Adrenodoxin reductase and the adrenodoxin complex. Biomed. Biochim. Acta 1990, 49, 167–172. [Google Scholar]
- Wardman, P. Reduction potentials of one-electron couples involving free radicals in aqueous solution. J. Phys. Chem. Ref. Data 1989, 18, 1637–1755. [Google Scholar] [CrossRef]
- Čėnas, N.; Anusevičius, Ž.; Bironaitė, D.; Bachmanova, G.I.; Archakov, A.I.; Öllinger, K. The electron transfer reactions of NADPH:cytochrome P450 reductase with nonphysiological oxidants. Arch. Biochem. Biophys. 1994, 315, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Anusevičius, Ž.; Martinez-Julvez, M.; Genzor, C.G.; Nivinskas, H.; Gomez-Moreno, C.; Nivinskas, H.; Čėnas, N. Electron transfer reactions of Anabaena PCC 7119 ferredoxin: NADP+ reductase with nonphysiological substrates. Biochim. Biophys. Acta 1997, 1320, 247–255. [Google Scholar]
- Lind, J.; Shen, X.; Merenyi, G.; Jonsson, B.Ö. Determination of the rate constant of self-exchange of the O2/O2·− couple in water by 18O/16O isotope marking. J. Am. Chem. Soc. 1989, 111, 7654–7655. [Google Scholar] [CrossRef]
- Anusevičius, Ž.; Šarlauskas, J.; Čėnas, N. Two-electron reduction of quinones by rat liver NAD(P)H: Quinone oxidoreductase: Quantitative structure-activity relationships. Arch. Biochem. Biophys. 2002, 404, 254–262. [Google Scholar] [CrossRef]
- Misevičienė, L.; Anusevičius, Ž.; Šarlauskas, J.; Čėnas, N. Reduction of nitroaromatic compounds by NAD(P)H: Quinone oxidoreductase (NQO1): The role of electron-accepting potency and structural parameters in the substrate specificity. Acta Biochim. Pol. 2006, 53, 569–576. [Google Scholar] [PubMed]
- Mendoza, M.F.; Hollabaugh, N.M.; Hettiarachchi, S.U.; McCarley, R.L. Human NAD(P)H: Quinone oxidoreductase type I (hNQO1) activation of quinone propionic acid trigger groups. Biochemistry 2012, 51, 8014–8026. [Google Scholar] [CrossRef]
- Nemeikaitė-Čėnienė, A.; Jarašienė, R.; Nivinskas, H.; Šarlauskas, J.; Čėnas, N. Cytotoxicity of anticancer aziridinyl-benzoquinones in murine hepatoma MH22a cells: The properties of RH1-resistant subline. Chemija 2015, 26, 46–50. [Google Scholar]
- O’Brien, P.J.; Wong, W.C.; Silva, J.; Khan, S. Toxicity of nitrobenzene compounds toward isolated hepatocytes: Dependence on reduction potential. Xenobiotica 1990, 20, 945–955. [Google Scholar] [CrossRef]
- O’Brien, P.J. Molecular mechanisms of quinone cytotoxicity. Chem. Biol. Interact. 1991, 80, 1–41. [Google Scholar]
- Nemeikaitė-Čėnienė, A.; Šarlauskas, J.; Anusevičius, Ž.; Nivinskas, H.; Čėnas, N. Cytotoxicity of RH1 and related aziridinylbenzoquinones: Involvement of activation by NAD(P)H:quinone oxidoreductase (NQO1) and oxidative stress. Arch. Biochem. Biophys. 2003, 416, 110–118. [Google Scholar] [CrossRef]
- Öllinger, K.; Brunmark, A. Effects of hydroxyl substituent position on 1,4-naphthoquinone toxicity to rat hepatocytes. J. Biol. Chem. 1991, 266, 21496–21503. [Google Scholar] [PubMed]
- Hubig, S.M.; Rathore, R.; Kochi, J.K. Steric control of electron transfer. Changeover from outer-sphere to inner-sphere mechanism in arene/quinone redox pairs. J. Am. Chem. Soc. 1999, 121, 617–626. [Google Scholar] [CrossRef]
- Raabe, J.; Arend, C.; Steinmeier, J.; Dringen, R. Dicumarol inhibits multidrug resistance protein 1-mediated export processes in cultured primary rat astrocytes. Neurochem. Res. 2019, 44, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Buffinton, G.D.; Öllinger, K.; Brunmark, A.; Cadenas, E. DT-diaphorase-catalysed reduction of 1,4-naphthoquinone derivatives and glutathionyl-quinone conjugates. Effects of substituents on autoxidation rates. Biochem. J. 1989, 257, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, Y.; Shiba, D.; Shimamoto, N. Enhancement of DMNQ-induced hepatocyte toxicity by cytochrome P 450 inhibition. Toxicol. Appl. Pharmacol. 2006, 214, 109–117. [Google Scholar] [CrossRef]
- Anusevičius, Ž.; Šarlauskas, J.; Nivinskas, H.; Segura-Aguilar, J.; Čėnas, N. DT-diaphorase catalyzes N-denitration and redox cycling of tetryl. FEBS Lett. 1998, 436, 144–148. [Google Scholar]
- Kurokawa, H.; Ito, H.; Inoue, M.; Tabata, K.; Sato, Y.; Yamagata, K.; Kizaka-Kondoh, S.; Kadonosono, T.; Yano, S.; Inoue, M.; et al. High resolution imaging of intracellular oxygen concentration by phosphorescence lifetime. Sci. Rep. 2015, 5, 10657. [Google Scholar] [CrossRef]
- Zeman, E.M.; Baker, M.A.; Lemmon, M.J.; Pearson, C.I.; Adams, J.A.; Brown, J.M.; Lee, W.W.; Tracy, M. Structure-activity relationships for benzotriazine di-N-oxides. Int. J. Radiat. Oncol. Biol. Phys. 1989, 16, 977–981. [Google Scholar] [CrossRef]
- Long, D.J., II; Iskander, K.; Gaikwad, A.; Arin, M.; Roop, D.R.; Knox, R.; Barrios, R.; Jaiswal, A.K. Disruption of dihydronicotinamide riboside:quinone reductase 2 (NQO2) leads to myeloid hyperplasia of bone marrow and decreased sensitivity to menadione toxicity. J. Biol. Chem. 2002, 247, 46131–46139. [Google Scholar] [CrossRef]
- Kirkland, D.; Marzin, D. An assessment of the genotoxicity of 2-hydroxy-1,4-naphthoquinone, the natural dye ingredient of Henna. Mutat. Res. 2003, 537, 183–199. [Google Scholar] [CrossRef]
- Pechurskaja, T.A.; Harnastai, I.N.; Grabovec, I.P.; Gilep, A.A.; Usanov, S.A. Adrenodoxin supports reactions catalyzed by microsomal steroidgenetic cytochrome P450s. Biochem. Biophys. Res. Commun. 2007, 353, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Prochaska, H.J. Purification and crystallization of rat liver NAD(P)H:quinone-acceptor oxidoreductase by cibacron blue affinity chromatography: Identification of a new and potent inhibitor. Arch. Biochem. Biophys. 1988, 267, 529–538. [Google Scholar] [CrossRef]
- Ito, M. Microassay for studying anticellular effects of human interferons. J. Interferon Res. 1984, 4, 603–608. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | E17 (V) a | kcat/Km (M−1·s−1) | |
|---|---|---|---|---|
| P-450R | ADR/ADX | |||
| Aromatic N-oxides | ||||
| 1 | 1,2,4-Benzotriazine-1,4-dioxide | −0.318 | 4.3 ± 0.4 × 105 | 4.0 ± 0.3 × 105 |
| 2 | 7-CF3-tirapazamine | −0.345 | 8.7 ± 0.7 × 104 | 1.9 ± 0.2 × 105 |
| 3 | 7-Cl-tirapazamine | −0.400 | 6.9 ± 0.7 × 104 | 7.9 ± 0.6 × 105 |
| 4 | 7-F-tirapazamine | −0.400 | 3.4 ± 0.3 × 104 | 5.2 ± 0.6 × 105 |
| 5 | Tirapazamine | −0.455 | 1.1 ± 0.1 × 104 | 1.0 ± 0.1 × 105 |
| 6 | 7-CH3-tirapazamine | −0.474 | 8.6 ± 0.7 × 103 | 1.2 ± 0.1 × 105 |
| 7 | 7-C2H5O-tirapazamine | −0.494 | 4.5 ± 0.5 × 103 | 1.5 ± 0.1 × 105 |
| 8 | 3-Amino-1,2,4-benzotriazine-1-oxide | −0.568 | 2.8 ± 0.2 × 103 | 7.2 ± 0.8 × 103 |
| 9 | Quinoxaline-1,4-dioxide | −0.575 | 3.3 ± 0.2 × 103 | 1.9 ± 0.2 × 104 |
| Quinones | ||||
| 10 | 5-Hydroxy-1,4-naphthoquinone | −0.090 | 1.5 ± 0.1 × 107 | 5.6 ± 0.6 × 106 |
| 11 | 5,8-Dihydroxy-1,4-naphthoquinone | −0.110 | 1.8 ± 0.1 × 107 | 1.1 ± 0.1 × 107 |
| 12 | 9,10-Phenanthrene quinone | −0.120 | 1.5 ± 0.2 × 107 | 6.1 ± 0.5 × 106 |
| 13 | 1,4-Naphthoquinone | −0.150 | 1.1 ± 0.1 × 107 | 4.0 ± 0.3 × 106 |
| 14 | 2-Methyl-1,4-naphthoquinone | −0.200 | 4.2 ± 0.4 × 106 | 3.6 ± 0.3 × 106 |
| 15 | Tetramethyl-1,4-benzoquinone | −0.260 | 8.6 ± 0.6 × 105 | 9.3 ± 0.7 × 105 |
| 16 | 1,8-Dihydroxy-9,10-anthraquinone | −0.325 | 1.2 ± 0.3 × 105 | 6.0 ± 0.7 × 105 |
| 17 | 2-Hydroxy-1,4-naphthoquinone | −0.410 | 1.3 ± 0.2 × 104 | 2.0 ± 0.3 × 105 |
| No. | Compound | E17 (V) | kcat (s−1) | kcat/Km (M−1·s−1) |
|---|---|---|---|---|
| 1 | 1,2,4-Benzotriazine-1,4-dioxide | −0.318 | 1.0 ± 0.10 | 5.1 ± 0.7 × 103 |
| 2 | 7-CF3-tirapazamine | −0.345 | 1.3 ± 0.10 | 3.7 ± 0.4 × 103 |
| 3 | 7-Cl-tirapazamine | −0.400 | 1.0 ± 0.10 | 4.3 ± 0.4 × 103 |
| 4 | 7-F-tirapazamine | −0.400 | 0.7 ± 0.10 | 4.9 ± 0.6 × 103 |
| 5 | Tirapazamine | −0.455 | 0.2 ± 0.03 | 2.4 ± 0.2 × 103 |
| 6 | 7-CH3-tirapazamine | −0.474 | 0.3 ± 0.05 | 1.7 ± 0.1 × 103 |
| 7 | 7-C2H5O-tirapazamine | −0.494 | 0.3 ± 0.04 | 1.0 ± 0.1 × 103 |
| 8 | 3-Amino-1,2,4-benzotriazine-1-oxide | −0.568 | 0.1 ± 0.02 | 1.6 ± 0.2 × 103 |
| 9 | Quinoxaline-1,4-dioxide | −0.575 | 0.6 ± 0.10 | 7.9 ± 0.9 × 103 |
| No. | Compound | E17 (V) | log D | cL50 (µM) MH22a | GI50 (µM) HCT-116 |
|---|---|---|---|---|---|
| Aromatic N-oxides | |||||
| 1 | 1,2,4-Benzotriazine-1,4-dioxide | −0.318 | −0.70 | 11 ± 1.5 | n.d. |
| 2 | 7-CF3-tirapazamine | −0.345 | 0.76 | 3.4 ± 0.4 | 6.0 ± 1.0 |
| 3 | 7-Cl-tirapazamine | −0.400 | 0.49 | 3.1 ± 0.5 | 13 ± 1.5 |
| 4 | 7-F-tirapazamine | −0.400 | 0.03 | 7.2 ± 1.0 | n.d. |
| 5 | Tirapazamine | −0.455 | 0.11 | 31 ± 5.5 | 75 ± 7.0 |
| 6 | 7-CH3-tirapazamine | −0.474 | 0.40 | 83 ± 10 | 50 ± 6.0 |
| 7 | 7-C2H5O-tirapazamine | −0.494 | 0.08 | 64 ± 10 | 60 ± 7.0 |
| 8 | 3-Amino-1,2,4-benzotriazine-1-oxide | −0.568 | 0.30 | ≥ 600 | ≥ 600 |
| 9 | Quinoxaline-1,4-dioxide | −0.575 | −0.90 | 325 ± 40 | n.d. |
| Quinones | |||||
| 10 | 5-Hydroxy-1,4-naphthoquinone | −0.090 | 1.82 | 2.5 ± 0.04 a | 0.30 ± 0.05 |
| 11 | 5,8-Dihydroxy-1,4-naphthoquinone | −0.110 | 2.19 | 0.58 ± 0.05 a | 0.25 ± 0.04 |
| 12 | 9,10-Phenanthrene quinone | −0.120 | 2.92 | 4.6 ±0.3 a 3.9 ± 0.4 b | 1.0 ± 0.1 |
| 13 | 1,4-Naphthoquinone | −0.150 | 1.49 | 3.1 ±0.05 a | 1.0 ± 0.1 |
| 14 | 2-Methyl-1,4-naphthoquinone | −0.200 | 1.89 | 18 ± 1.3 a | 7.0 ± 1.2 |
| 15 | Tetramethyl-1,4-benzoquinone | −0.260 | 2.61 | 59 ± 5.0 a 50 ± 5.0 b | 45 ± 6.5 |
| 16 | 1,8-Dihydroxy-9,10-anthraquinone | −0.325 | 3.56 | 120 ± 15 a | n.d. |
| 17 | 2-Hydroxy-1,4-naphthoquinone | −0.410 | −0.52 | 500 ± 80 a 430 ± 60 b | 300 ± 28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nemeikaitė-Čėnienė, A.; Šarlauskas, J.; Jonušienė, V.; Marozienė, A.; Misevičienė, L.; Yantsevich, A.V.; Čėnas, N. Kinetics of Flavoenzyme-Catalyzed Reduction of Tirapazamine Derivatives: Implications for Their Prooxidant Cytotoxicity. Int. J. Mol. Sci. 2019, 20, 4602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184602
Nemeikaitė-Čėnienė A, Šarlauskas J, Jonušienė V, Marozienė A, Misevičienė L, Yantsevich AV, Čėnas N. Kinetics of Flavoenzyme-Catalyzed Reduction of Tirapazamine Derivatives: Implications for Their Prooxidant Cytotoxicity. International Journal of Molecular Sciences. 2019; 20(18):4602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184602
Chicago/Turabian StyleNemeikaitė-Čėnienė, Aušra, Jonas Šarlauskas, Violeta Jonušienė, Audronė Marozienė, Lina Misevičienė, Aliaksei V. Yantsevich, and Narimantas Čėnas. 2019. "Kinetics of Flavoenzyme-Catalyzed Reduction of Tirapazamine Derivatives: Implications for Their Prooxidant Cytotoxicity" International Journal of Molecular Sciences 20, no. 18: 4602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184602