Rhodococcus as a Versatile Biocatalyst in Organic Synthesis



Department of Biotechnology, Delft University of Technology, Van der Maasweg 9, 2629 HZ Delft, The Netherlands
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(19), 4787; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194787
Submission received: 9 September 2019
/
Revised: 23 September 2019
/
Accepted: 24 September 2019
/
Published: 26 September 2019
(This article belongs to the Special Issue 23rd Anniversary of IJMS: Advances in Biochemistry)
Abstract
:The application of purified enzymes as well as whole-cell biocatalysts in synthetic organic chemistry is becoming more and more popular, and both academia and industry are keen on finding and developing novel enzymes capable of performing otherwise impossible or challenging reactions. The diverse genus Rhodococcus offers a multitude of promising enzymes, which therefore makes it one of the key bacterial hosts in many areas of research. This review focused on the broad utilization potential of the genus Rhodococcus in organic chemistry, thereby particularly highlighting the specific enzyme classes exploited and the reactions they catalyze. Additionally, close attention was paid to the substrate scope that each enzyme class covers. Overall, a comprehensive overview of the applicability of the genus Rhodococcus is provided, which puts this versatile microorganism in the spotlight of further research.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The genus Rhodococcus belongs to the phylum Actinobacteria and its members are aerobic, Gram-positive, non-motile bacteria showing high GC-contents [1]. Numerous members have been isolated from copious sources such as soil, groundwater, marine sediments, internal organs of insects, diseased and healthy animals, or plants-to name a few [2]. While the vast majority is considered harmless, a few species have shown pathogenic properties leading to foal pneumonia (R. equi) or leafy gall disease in plants (R. fascians). Starting in the 1980s, their application as a (whole-cell) biocatalyst came to the fore and since then, numerous reports of successful bioconversions have been published with an increasing trend [3]. The longstanding synthesis of acrylamide on a multi-ton scale run at several production sites worldwide is hereby considered as the most outstanding example of a rhodococcal whole-cell process [4,5,6].
Rhodococcus members are diverse catalysts that degrade a variety of both natural organic and xenobiotic compounds [7]. Amongst others, the Rhodococcus species showed biodegradation potential against short- and long-chain alkanes, halogenated and nitro-substituted aromatic, heterocyclic, and polycyclic compounds [8]. Next to physiological attributes such as a high general tolerance to substrates and solvents [8,9], the metabolic diversity of the genus Rhodococcus can be explained by the (i) presence and mobility of large, linear plasmids; (ii) the multiplicity of catabolic genes; (iii) the high redundancy of biosynthetic pathways; and (iv) sophisticated regulatory networks of their genomes [1,3,10]. This brands the genus Rhodococcus as the strong biocatalytic powerhouse that it is seen as today. The usage of whole biosynthetic pathways from Rhodococcus strains in the bioremediation of organic pollutants derived from petroleum such as o-xylene has been intensively investigated [11,12] and shown to be successful as was, for example, the lignin degradation catalyzed by R. jostii RHA1 [13,14,15]. Here, vanillin, a valuable chemical for food flavoring, was mainly produced [13]. The same strain also displayed biodegradation activity against polychlorinated biphenyls (PCBs) [16]. Additionally, Rhodococcus strains were used in desulfurization reactions able to degrade sulfur-containing compounds produced in the treatment of fossil fuels like benzothiophene (BT) or dibenzothiophene (DBT) [10,17].
As shortly displayed, the implementation of whole pathways to degrade numerous compounds is a promising tool with strong future potential application [7,18]. Nevertheless, this review will only focus on Rhodococcus as a useful tool in organic synthesis, concentrating on defined enzyme reactions catalyzed by purified enzymes or whole-cells primarily leading to enantioenriched products. It should serve as a comprehensive overview of the state-of-the-art biocatalysis that is feasible with Rhodococcus, concentrating on the reaction diversity and the respective substrate scope of each enzyme class described.
2. Promiscuous Redox-Reactions in Rhodococcus
Oxidoreductases (EC 1) catalyze the electron transfer from one molecule (reductant) to another molecule (oxidant), thereby enabling oxidation–reduction reactions often under the requirement of cofactors like the NAD(P)H serving as an electron donor or acceptor. They make one of the biggest groups and as such, an impressive range of redox-reactions is feasible. Additionally, the genus Rhodococcus harbors a large amount of oxidoreductases, some of which are already well-established systems even running at an industrial scale, while others are still being thoroughly investigated and developed.
2.1. Alcohol Dehydrogenases/Keto Reductases
Alcohol dehydrogenases (ADHs, EC 1.1.x) catalyze the transformation of primary and secondary alcohols to aldehydes and ketones. A common feature of an ADH-catalyzed reaction is the requirement for a primary oxidant within the active site of the enzyme, which oxidizes the alcohol. This process generally occurs via a hydride abstraction. Nicotinamide cofactors (NAD+, NADP+), pyrroquinoline quinones, or flavins are typical prosthetic groups acting as hydride acceptors that need to be regenerated during the reaction for the overall process to be a success [19]. A nicotinoprotein from R. erythropolis DSM 1069 showed activity toward aliphatic and aromatic primary and secondary alcohols whereby primary alcohols were the preferred substrates yielding aldehydes [20]. Another application of ADHs in an oxidative sense is the kinetic resolution of secondary alcohols leading to both ketones and enantiopure alcohols. When applying whole-cells of R. ruber DSM 44541, only the (S)-enantiomer of racemic mixtures of alcohols was oxidized leaving behind the unreacted (R)-alcohol. This proof-of-principle was successfully shown for the substrate sulcatol as well as aromatic and aliphatic alcohols [21]. A R. erythropolis mediated kinetic resolution of methyl-nonactate yielded the two enantiomers (+)- and (-)-nonactate in excellent enantiomeric purity [22]. Fascinatingly, the stereoselectivity of the main product was influenced by switching the conditions from aerobic to anaerobic. (+)- and (-)-nonactate are the desired building blocks for the macrotetrolide nonactin.
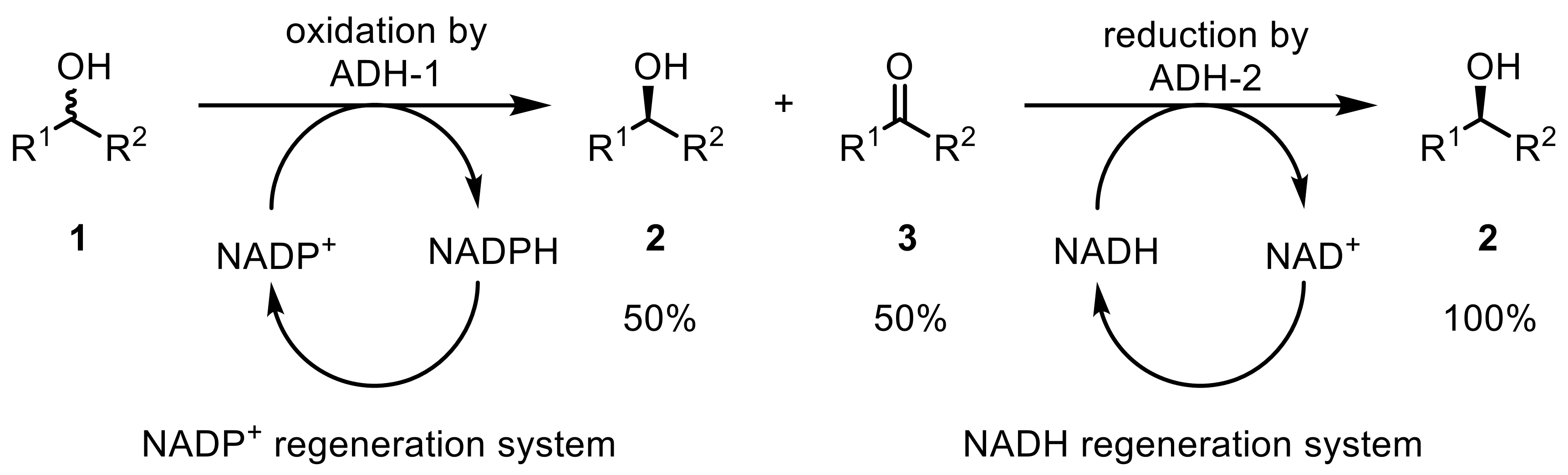
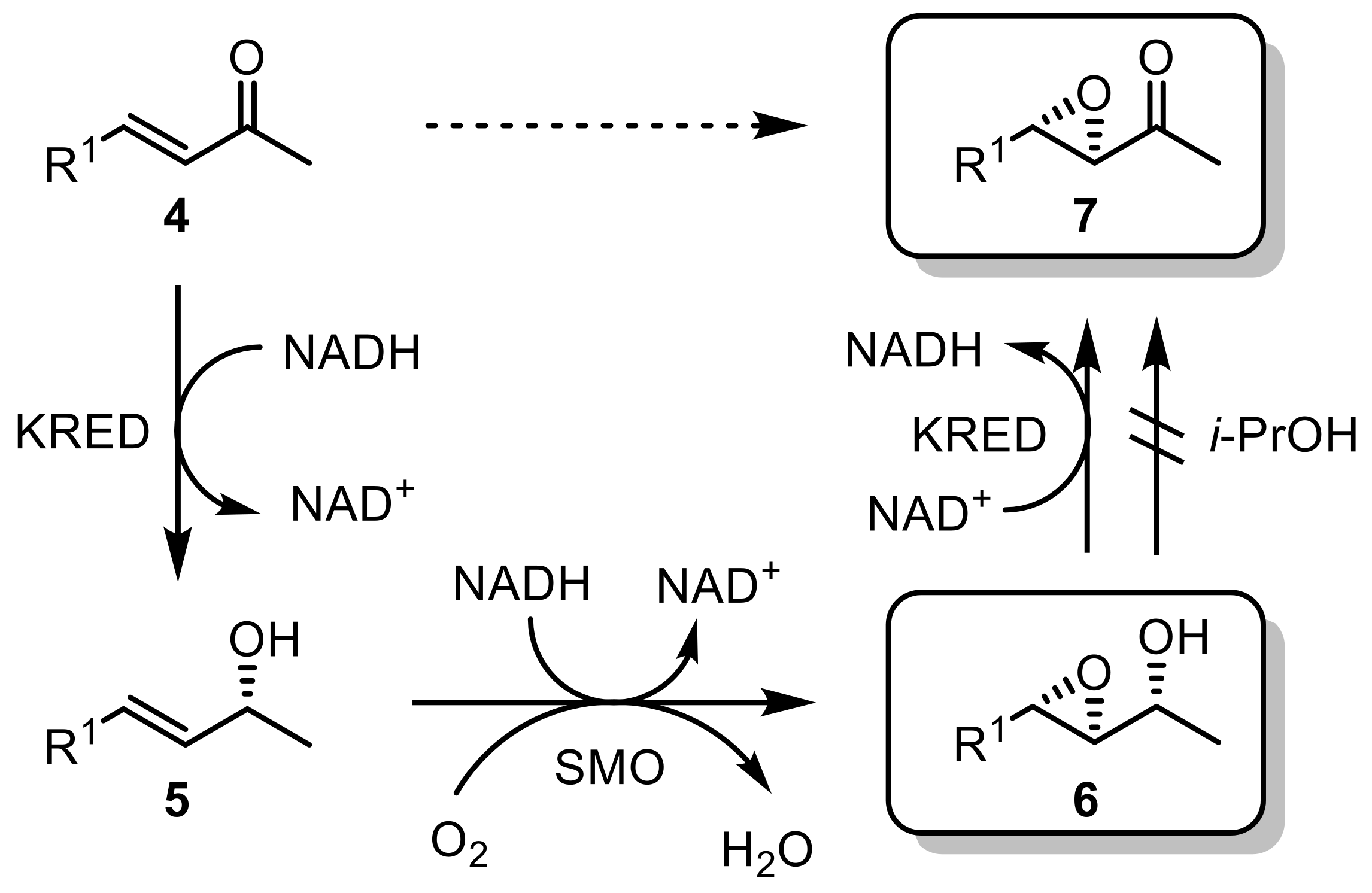
To overcome the limitation of only 50% theoretical yields in kinetic resolutions, a two-ADH system was developed resolving racemic alcohols (1) [23]. First, one ADH (ADH-1) catalyzes the oxidation of one enantiomer to the corresponding ketone (3), leaving one alcohol-enantiomer unreacted (2). A second, stereocomplementary ADH (ADH-2) is afterward applied, which reduces the obtained ketone (3) to the wanted alcohol-enantiomer (2), thereby enabling a theoretical yield of 100% (Scheme 1). Each step is catalyzed by two different ADHs, each requiring another cofactor.
The (S)-selective ADH from R. ruber DSM 44541 (termed ADH-A) was part of this study leading, in combination with (R)-selective ADH from Lactobacillus kefir (LK-ADH), to the formation of (S)-alcohols in excellent yields and ee. A complete stereoinversion of chiral secondary alcohols was also achieved by coupling two stereocomplementary ADHs: while the first ADH oxidized the starting alcohol to the ketone intermediate, a second ADH reduced the ketones to the other alcohol enantiomers, resulting in a complete inversion of the stereochemistry [24].
ADHs from Rhodococcus were also employed in an oxidative manner in whole-cell biotransformations for straightforward one-pot multistep reactions [25]. Two representative examples demonstrate the broad applicability of ADHs from Rhodococcus within biocascades: the double oxidation of c-octane was achieved by a combination of three enzymes, namely a monooxygenase P450 BM3, Lactobacillus brevis ADH (LbADH) and R. erythropolis ADH (ReADH) including a cofactor regeneration system, whereby all enzymes were coexpressed in a single host cell [26]. The monooxygenase first oxyfunctionalizes the non-activated c-octane to c-octanol, which is further oxidized by the two stereocomplementary ADHs. In a similar fashion, four recombinant enzymes (a self-sufficient P450 monooxygenase, two stereocomplementary ADHs (LbADH and ReADH) as well as a ω-transaminase (ATA-117)) were co-expressed in E. coli, catalyzing the chiral amination of benzylic compounds such as (substituted)-ethylbenzene in a one-pot two-step process [27].
ADHs that catalyze the reverse reaction are so-called ketoreductases (KREDs). Through carbonyl reduction, the generation of (chiral) alcohols is thereby enabled. As the introduction of chiral centers is seen as more valuable than the loss of enantiomeric centers, this reverse reaction is more often applied in synthetic chemistry.
The reduction of aldehydes is hereby less often reported due to the fact that this reduction does not invoke a new stereocenter, but leads to primary alcohols. However, a KRED from Rhodococcus sp. was employed in the multi-step chemo-enzymatic synthesis of Guerbet alcohols [28]. It was used to catalyze the final step by reducing an aldehyde to form a primary alcohol. The use of enzymes in this example led to an improvement of the overall reaction conditions, reducing the temperature and applied pressure, and increasing the selectivity compared to alternative routes.
While most of the following research is based on the use of KREDs identified from R. erythropolis (ReADH) and R. ruber (ADH-A), a newly characterized ketoreductase from R. jostii TMP1 was applied in the synthesis of chiral alcohols showing a broad substrate acceptance with increased affinity toward aliphatic 2,3-diketones, butan-3-one-2-yl alkanoates, and acetoin as well as the respective derivatives [29].
The substrate scope of a recombinantly expressed KRED from R. erythropolis was evaluated showing that a multitude of ketones was eligible for KRED-mediated reduction forming the (S)-selective alcohols [30]. Next to mono-, di-, and tri-substituted acetophenones, aliphatic ketones have also been implemented in a biphasic reaction medium using an integrated cofactor-recycling system (formate dehydrogenase system). The synthesis of 1-[3,5-bis(trifluoromethyl)phenyl]ethanol, another di-substituted acetophenone, which acts as a key intermediate in the synthesis of NK-1 receptor antagonists, was achieved with the same enzyme coupled to another cofactor-recycling system (glucose dehydrogenase system) [31]. In a stirred-tank reactor, space-time yields of 260 g/L per day were achieved.
Interestingly, electron-deficient α,β-unsaturated ketones (4) were treated in a tandem reduction–epoxidation–dehydrogenation cascade involving two enzymes, namely an AHD from R. erythropolis DSM 43297 and a styrene monooxygenase both co-expressed in E. coli [32]. While aiming for the synthesis of chiral epoxy ketones (7), it was found that allylic epoxy alcohol intermediates (6) also form valuable synthons due to the presence of three contiguous stereocenters. Through adaption of the reaction conditions (the addition of isopropanol, which caused high concentrations of NADH, thereby blocking the dehydrogenation reaction), the major product could easily be switched from an epoxy ketone (7) to an allylic epoxy alcohol (6), thereby providing two useful synthons with only one reaction set-up (Scheme 2).
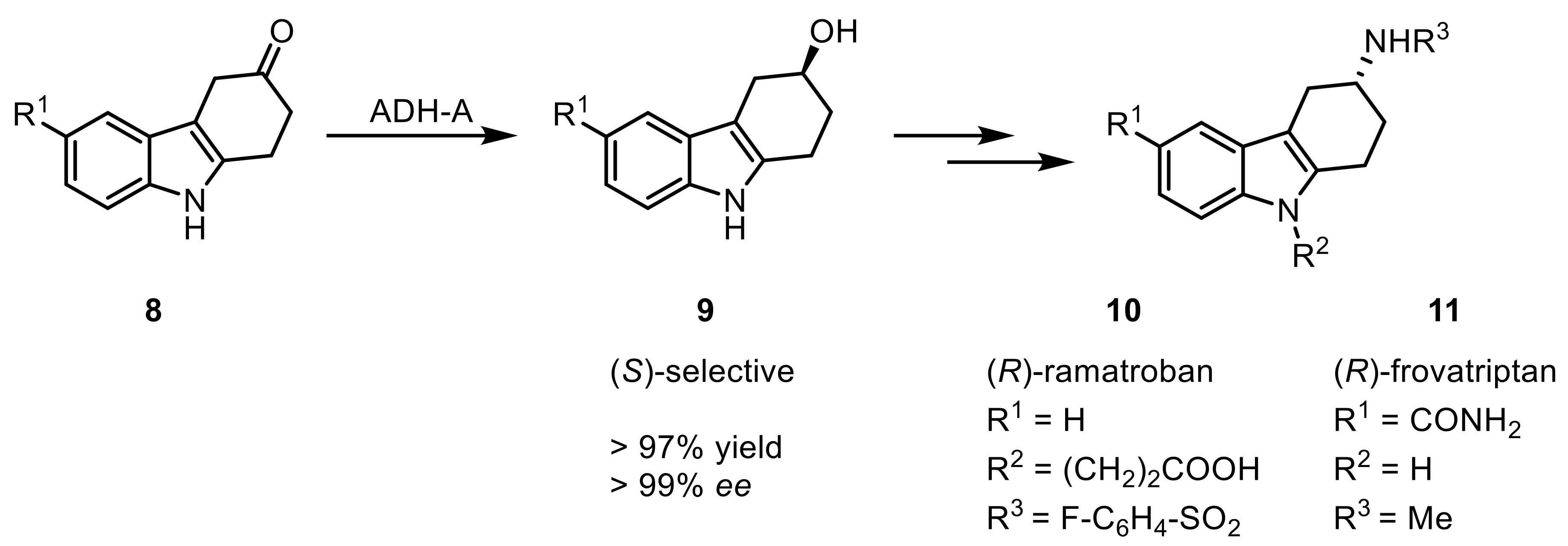
Additionally, trifluoroacetyl-acetophenones lead to chiral fluorinated hydroxyketones when converted by ReADH [33]. The conversion of phenylacyl halides, here especially fluorides, was performed with recombinantly expressed ADH-A yielding fluorohydrins with high yields and enantiomeric purity [34]. ADH-A was also applied in the synthesis of pharmacologically active compounds (R)-Ramatroban (10), a thromboxane receptor and protaflandin DP2 receptor antagonist, and (R)-Frovatriptan (11), a 5-hydroxytryptamine (serotonin) receptor antagonist, treating allergic rhinitis and asthma or migraine headaches, respectively [35,36]. In both total syntheses, the KRED was used to install an (S)-alcohol (9), which was subsequently inverted following a SN2-mechanism to an (R)-amine. Afterward, the selective functionalization of the amine allowed generation of both desired compounds (Scheme 3).
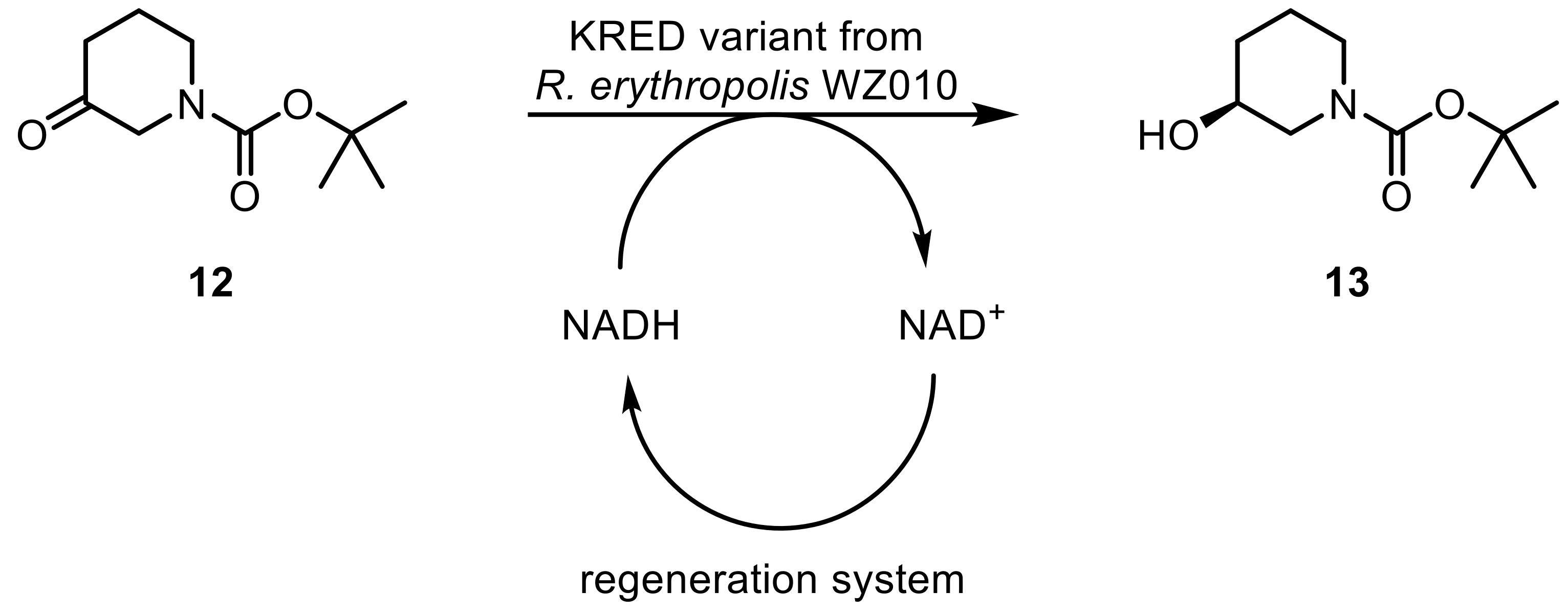
The same enzyme (ADH-A) was applied in the production of syn-α-alkyl-β-hydroxy amides through means of dynamic kinetic resolution. Various acyclic α-alkyl-β-keto amides were reduced, yielding the (S)-selective products’ high yields and excellent ee [37]. Enantiopure 3,4-dihydroisocoumarins were easily accessible via a one-pot dynamic reductive kinetic resolution process catalyzed by ADH-A [38] as was the asymmetric synthesis of (S)-N-Boc-3-hydroxypipridines (13) using a variant (Y54F) of a KRED from R. erythropolis WZ010 [39]. N-Boc-3-piperidone (12) was selectively reduced, thereby forming an important intermediate of ibrutinib, an inhibitor of Bruton’s tyrosine kinase. The enzymatic process enhanced the yield significantly when compared to the chemical route (Scheme 4).
The discussed examples show the broad applicability of ADHs/KREDs in organic synthesis by opening up the possibility of introducing new stereocenters through reduction processes or (dynamic) kinetic resolutions.
2.2. Oxidases
Another class of enzyme, alcohol oxidases, also catalyze the oxidation of alcohols to aldehydes, ketones, and sometimes carboxylic acids. In comparison to alcohol dehydrogenases that catalyze this reaction via an electron-transfer to an organic cofactor, alcohol oxidases transfer electrons to molecular oxygen forming hydrogen peroxide as a side-product. Using oxygen as a cheap and readily available oxidant makes oxidases more interesting than alcohol dehydrogenases as they require expensive cofactors in either stoichiometric amounts or respective cofactor regeneration systems [40].
Vanillyl alcohol oxidase (VAO) from Penicillium simplicissimum is known to catalyze the oxidation of many 4-hydroxybenzylic alcohols. A homologue of this enzyme was found in R. sp. RHA1, which was termed eugenol oxidase (EUGO) [41]. The enzyme was shown to catalyze the oxidation of vanillyl alcohol more efficiently than VAO from P. simplicissimum, but 4-alkylphenols as well as 4-(methoxymethyl)phenol were only poorly converted.
Cholesterol oxidases (ChoX), on the other hand, catalyze the oxidation of the C3 hydroxyl-group of cholesterol and an isomerization reaction, ultimately yielding cholest-4-en-3-one [40]. In addition to cholesterol, a cholesterol oxidase discovered in R. erythropolis also showed activity toward non-steroidal compounds such as smaller cyclic allylic alcohols with good stereo- and enantioselectivity [42]. In an attempt to produce high-quality cholest-4-en-3-one, the use of an aqueous/organic biphasic system was explored which simplified the production process by improving separation and purification [43]. This led to a final product with a purity of 99.78%, which makes this new process design more approachable for industrial applications.
A Rhodococcus oxidase catalyzed reaction already running on industrial scale is a kinetic resolution through the oxidation of racemic iso-propylideneglycerol (14) yielding both (R)-iso-propylideneglyceric acid (15) and (R)-iso-propylideneglycerol (16) [44,45]. The use of this biocatalytic process simplifies the synthesis of desired (R)-iso-propylideneglycerol (16) compared to chemical processes. Whole-cells of R. erythropolis are employed in a fed-batch reactor leading to 50% maximum yield with high ee values. The product is a valuable C3 synthon in the synthesis of β-blockers like (S)-metoprolol (17) (Scheme 5).
2.3. Oxygenases in Rhodococcus
Oxygenases utilize molecular oxygen as both a substrate and electron acceptor, while the (above) discussed dehydrogenases catalyze the oxidation via hydrogen transfer reactions. Oxygenases can be classified into two groups: while monooxygenases introduce only one oxygen-atom, dioxygenases catalyze the introduction of two oxygen atoms from molecular oxygen. This type of reaction is particularly useful in synthetic chemistry as a selective activation of chemically inert C–H bonds is otherwise extremely difficult to achieve using classical chemical procedures.
2.3.1. Monooxygenases
P450 Monooxygenases
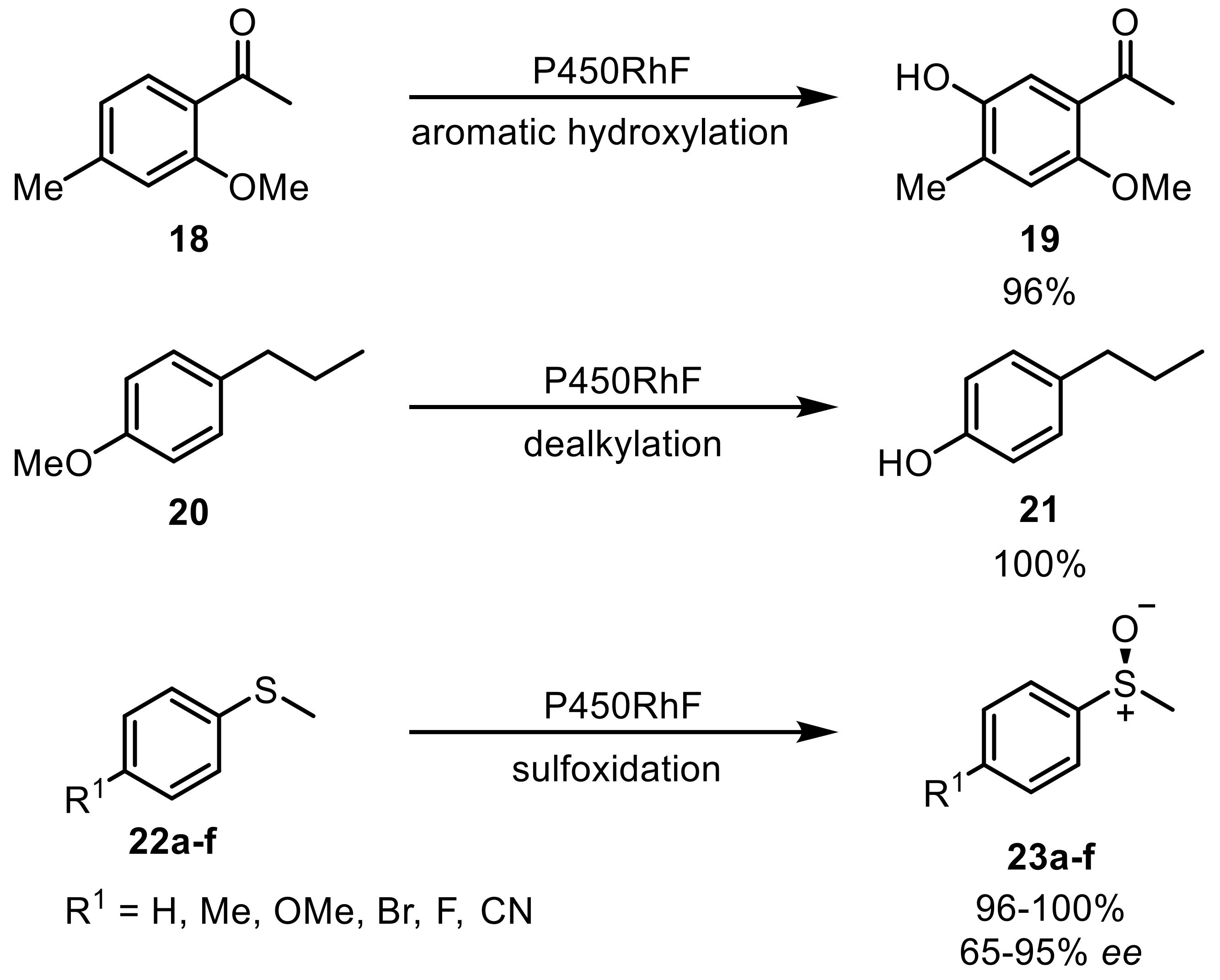
The so-called cytochrome P450s (P450s, CYPs) monooxygenases contain a heme prosthetic group with an Fe(III)-ion embedded in a porphyrin ring with a cysteine sulfur as an axial ligand to the iron. They activate molecular oxygen for the hydroxylation of organic compounds. In this process, the second oxygen is reduced to water. P450s are NAD(P)H dependent enzymes. Therefore, the redox-reaction required electrons are transferred from the cofactor to the heme through one or two ‘electron transport enzymes’ or ‘redox partners’ [46,47]. This makes P450s multicomponent enzymes. The involved components can either be free or directly linked to each other with the latter being called a self-sufficient enzyme. One of these self-sufficient P450s was discovered in R. sp. NCIMB 9784 where the reductase partner (RhFRed) containing the FMN- and NADPH binding motif and a Fe2S2-ferredoxin-like component is directly linked to the oxidase part [48,49]. The natural substrate of this enzyme has not been discovered yet, but it shows a promiscuous substrate scope mediating dealkylation reactions, aromatic hydroxylation, epoxidation, and asymmetric sulfoxidation (Scheme 6) [50].
The reductase unit of this P450 is, however, more often employed. The heme domain of P450RhF can be swapped with other heme domains, thereby fusing the reductase domain RhFRed to a number of different enzymes [47,51]. One successful example is given by the hydroxylation of testosterone by a chimeric fusion protein consisting of the heme-domain from CYP154 from Nocardia farcinca IFM 10152 and the reductase domain RhFRed [51].
Baeyer-Villiger Monooxygenases
The Baeyer-Villiger reaction involves the oxidation of a carbonyl compound, ultimately leading to an ester or lactone. So-called Baeyer-Villiger monooxygenases (BVMOs) catalyze this reaction under milder reaction conditions compared to harsh chemical procedures [52]. Flavin-containing enzymes require electrons from a reduced cofactor. The flavin-cofactor reacts with molecular oxygen, thereby forming a reactive peroxyflavin intermediate, which performs the nucleophilic attack on the carbonyl function. Upon a rearrangement, the respective ester or lactone is formed [52].
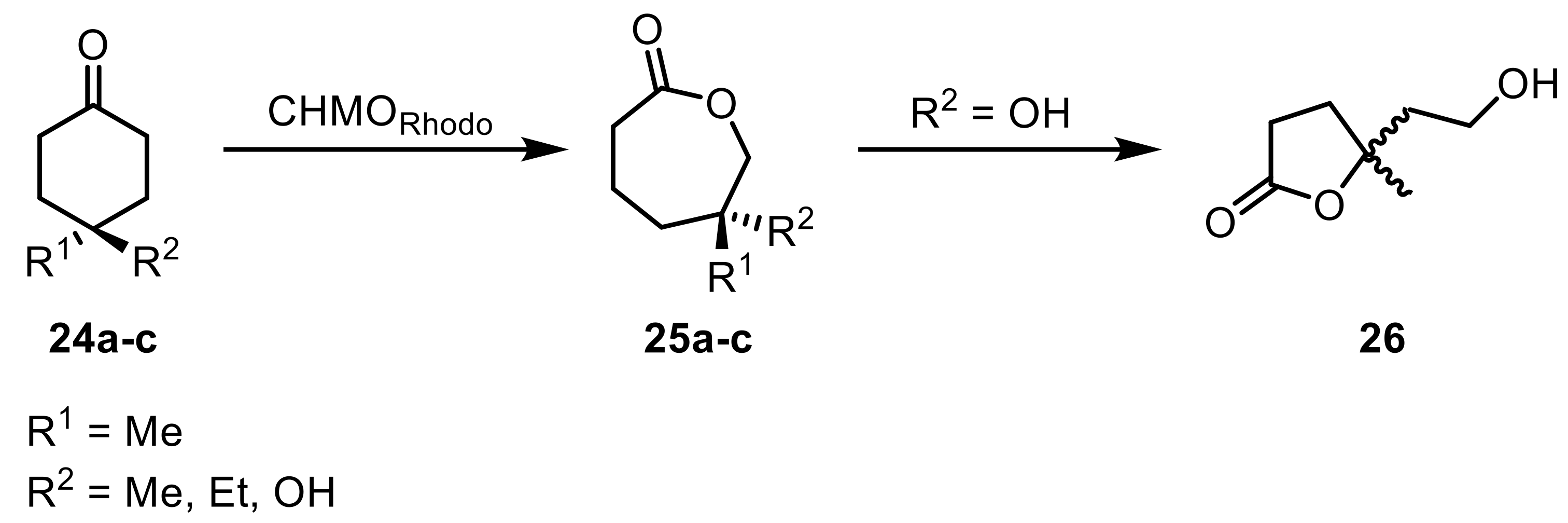
Prochiral c-butanones with alkyl- or aromatic substituents in the 3-position were converted by the two c-hexanone monooxygenases discovered earlier in Rhodococcus [53] (CHMORhodo1 and CHMORhodo2) to yield (S)-selective butyrolactones [54]. Both enzymes displayed especially high activity toward bulky substituent piperonyl and aromatic residues with substituents in the m- and p-position. The same enzymes were employed in an activity screening toward 4-4-disubstituted c-hexanones (24) to obtain the respective caprolactones (25) [55]. In the special case of 4-hydroxy-4-methyl-c-hexanone (24c), a caprolactone (25c) was generated, which spontaneously formed a five-membered ring (26) (Scheme 7).
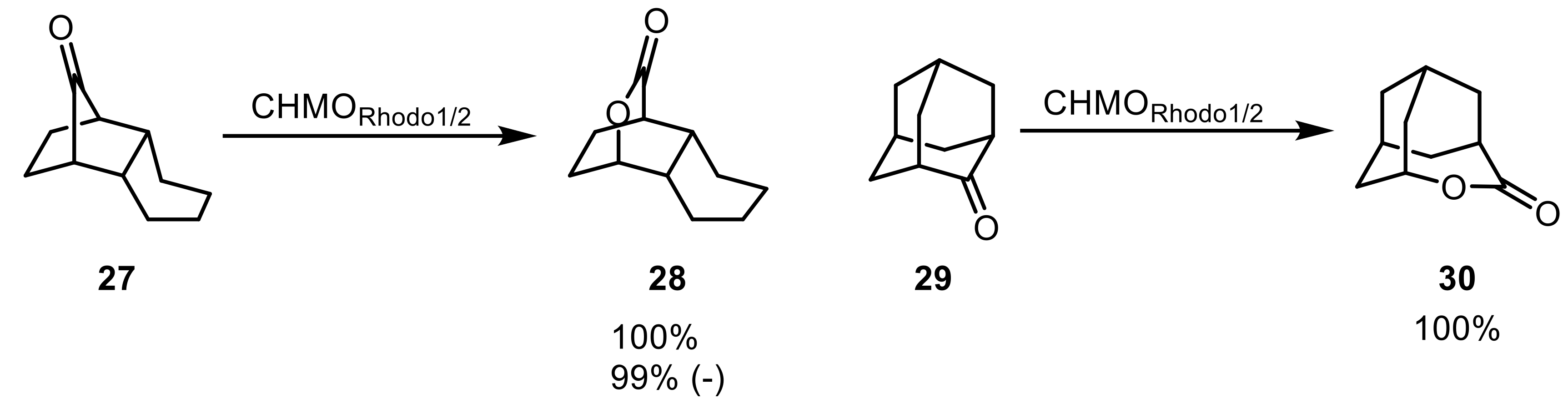
Additionally, the same set of enzymes was used in another screening toward a number of bridged c-ketones [56]. Both tested Rhodococcus enzymes readily converted almost all tested bridged ketones (27,29) with high yields and good to excellent stereoselectivities yielding bicyclic lactones (28,30) (Scheme 8).
The ability to resolve N-protected β-amino ketones was investigated using the two c-hexanone monooxygenases from Rhodococcus amongst other bacterial BVMO [57]. CHMORhodo1+2 both showed the ability to resolve linear(-branched) aliphatic and aryl-aliphatic 4-amino-2-ketones with a strong preference for middle-chain 4-amino-2-ketones. Longer substrates (C12) were, however, not converted. No activity was detected for 5-amino-3-ketones. Next to the more intensively studied c-hexanone monooxygenases from Rhodococcus (CHMORhodo1+2), several novel BVMOs have been identified in various strains. Increasing the number of BVMOs leads to an expansion of the application potential of this useful enzyme class.
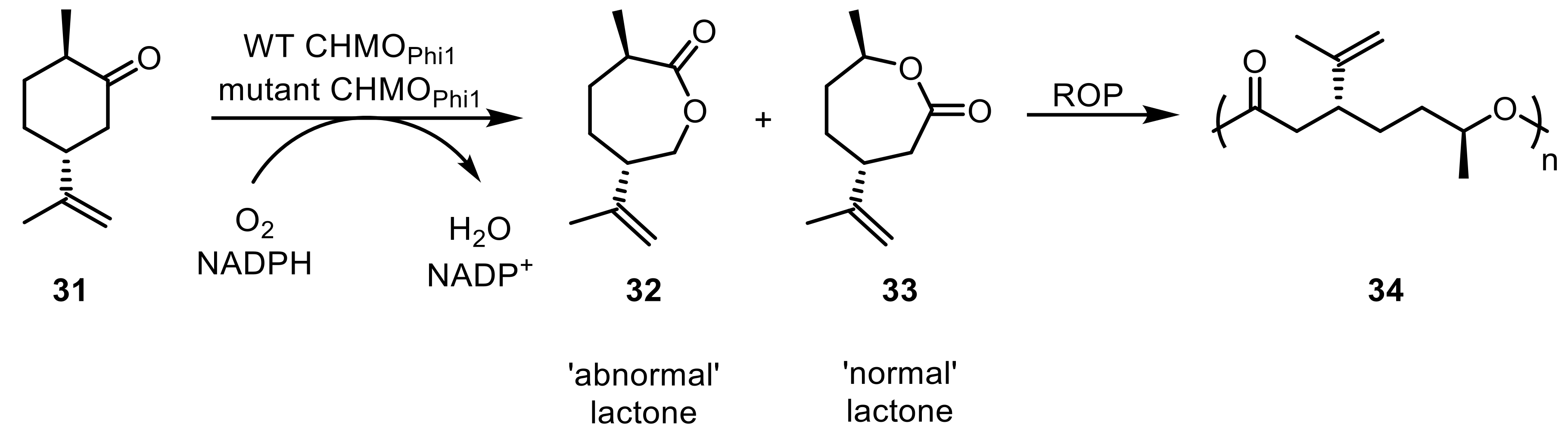
Through a kinetic resolution of 2-(3-penten-1-yl)-c-hexanone catalyzed by a c-dodecanone monooxygenase from R. ruber SC1 (CDMO) [58], the formation of a homologue of the jasmine lactone was achieved with excellent stereoselectivity [59]. Preparation of the jasmine lactone, a desired compound in fragrance industry, was accomplished with a CHMO from Arthrobacter in the same study. The species R. jostii RHA1 is particularly known for a high abundance of oxidative enzymes [60,61], which gave rise to a comprehensive investigation of the presence of BVMOs in this strain [62]. Following a genome mining approach, a total number of 22 novel BVMOs was identified, expressed, and tested on a diverse set of 39 substrates ranging from linear and cyclic aliphatic ketones to aromatic amines, sulfides, and ketones. Here, in particular, six of the identified BVMOs stood out due to their high substrate promiscuity converting at least 10 and up to 29 of the tested substrates. Furthermore, the same microorganism was shown to also host eight new flavin-containing monooxygenases (FMOs), three of which were successfully applied in Baeyer-Villiger oxidations [63]. Interestingly, the novel enzymes did not favor a specific coenzyme (NADH or NADPH) and their potential as useful biocatalysts was shown by successful conversions of both an aromatic and a bicyclic ketone. A novel CHMO from Rhodococcus sp. Phi1 (CHMOPhi1) was used in an attempt to chemo-enzymatically produce the lactone monomer dihydrocarvide (32/33) from monoterpenoid starting materials (31) [64]. A subsequent metal-assisted ring opening polymerization (ROP) led to the generation of polydihydrocarvide (34), a polymer used as a thermoplastic elastomer. Depending on whether the wild-type enzyme or a triple mutant was employed, the synthesis of so-called abnormal (32) and normal lactone (33) were favored, respectively (Scheme 9).
In a similar fashion, a c-pentadecanone monooxygenase from Pseudomonas sp. HI-70 led to the formation of polymenthide in the same study. Finally, a novel BVMO from R. pyridinivorans DSM 44555 with extraordinary resistance toward high substrate loading and good stability was shown to convert a number of linear aliphatic ketones [65]. Both 2- and 3-ketones and their respective derivatives were converted. Particularly interesting was the production of 3-acetoxypropionate from methyl levulinate with a space-time-yield of 5.4 g/L per day, thereby more than doubling the highest STY reported thus far.
Styrene and Indole Monooxygenases
While the BVMOs are built up by a single-component, the styrene (SMO) and indole monooxygenases (IMO) form their own subgroup within the two-component flavin-dependent monooxygenases [66]. Reduced FAD, which binds in the active site of the monooxygenase, is used for the activation of molecular oxygen and is delivered by a NAD(P)H-dependent flavin reductase. While SMOs consist of a monooxygenase (StyA) and a reductase component (StyB), IMOs can either be built up in the same manner (two-component system with monooxygenase ‘IndA’ and reductases ‘IndB’) or as a self-sufficient fusion protein (ImoA2B) associated with an additional monooxygenase (IndA1).
Styrene monooxygenases catalyze the conversion of styrene and its derivatives and also showed activity against aryl alkyl sulfides. The epoxidation reaction as well as the sulfoxidation reaction solely yielded the respective (S)-enantiomers using rhodococcal SMOs with different regeneration systems [67,68,69,70]. Furthermore, the SMO from R. sp. ST-10 was used to convert aliphatic alkenes including terminal, internal, unfunctionalized as well as di- and tri-substituted alkenes, thereby generating (S)-epoxyalkanes [71]. The same gene was overexpressed in Kocuria rhizophila DC2201, a strain with exceptionally high tolerance against organic solvents [72]. This led to an increased conversion yield, thereby making this system a suitable biocatalyst for the environmentally milder production of (S)-epoxyalkanes in high purity.
2.3.2. Dioxygenases
Aromatic dioxygenases carry out the cis-dihydroxylation to arene substrates, thereby generating valuable vicinal cis-dihydrodiols. The aromatic ringhydroxylating dioxygenases are non-heme iron-dependent enzymes containing a mononuclear iron-active site and a Rieske type [FeS] cluster for electron transfer. These enzymes also require a ferredoxin and a flavin containing reductase that reacts with NADH. To date, mainly toluene and naphthalene dioxygenases from Pseudomonas putida (mutants) have been exploited, but next to those, naphthalene dioxygenases [76] and novel o-xylene degrading Rhodococcus strains were also discovered [77].
In particular, an increased interest on the isolation of the strain R. sp. DK17 arose when it was shown to replicate on o-xylene and utilize several aromatic compounds (benzene, alkylbenzene, phenol, phthalate), thereby displaying its high degradation potential [11]. Until now, subsequent studies primarily dealt with the deeper understanding of its unique reaction mechanism including the point of initial attack in the arene substrate [77]. In the course of this in-depth investigation, several substrates have already been shown to be modified by either the wild-type or engineered enzyme [78]. Amongst others, the o-xylene-3,4-dioxygenase from R. sp. DK17 showed, for example, activity against m- and p-xylene [79] as well as larger substrates such as naphthalene, indan, tetralin and indene, whereby in all cases, the respective cis-dihydrodiols were generated [80,81,82].
R. sp. I24 was found to be a strain that oxidizes indene via three different enzyme activities: next to a monooxygenase and a dioxygenase both inducible with naphthalene, a toluene inducible dioxygenase is present [83,84]. Great attention has been paid to the bioconversion of indene to cis-(1S,2R)-indandiol, which is a known precursor for (-)-cis-(1R,2R)-1-aminoindan-2-ol, a key chiral synthon for the HIV protease inhibitor Crixivan. Whole-cell experiments with R. sp. I24 carried out in a batch- or fed-batch manner struggled with low yields due to the numerous side-reactions catalyzed by the other oxygenases present [83,85]. Therefore, the toluene inducible dioxygenase (TID) was heterologously expressed in E. coli and investigated [86]. The desired cis-(1S,2R)-indandiol was produced with an enantiomeric excess of 45.2% over cis-(1R,1R)-indandiol.
Several other Rhodococcus species have been investigated for their degradation potential toward the group of BTEXS (benzene, toluene, ethylbenzene, xylene isomers, styrene) aromatics. For example, the conversion of benzoate was catalyzed by a benzoate dioxygenase with a narrow substrate scope from R. opacus 1CP [87]. 2,3-dihydroxybiphenyl-1,2-dioxygenases from Rhodococcus have been recombinantly expressed, showing activity toward a number of catechols with 2,3-dihydroxybiphenyl being the best accepted substrate [88,89] and several catechol-1,2-dioxygenases were shown to cleave (alkyl-substituted and halogenated) catechols [90,91].
However, to the best of our knowledge, their use as biocatalysts in synthetic chemistry has been limited and not been exploited to its full potential yet.
2.4. Miscellaneous Oxidation Potential
As already mentioned, Rhodococcus strains show impressive degradation behavior toward a multitude of compounds. Several monooxygenases present in these biodegradation pathways have been identified and implemented in biocatalytic applications. To further showcase the hydroxylation potential of enzymes isolated from Rhodococcus as well as whole-cells, several examples are discussed.
With the responsible enzymes staying elusive in some cases, a number of different terpenoids were described to be transformed by whole-cells of Rhodococcus. As an example, d-limonene was oxidized to (+)-trans-carveol by R. opacus PWD4 [92] and β-myrcene to geraniol [93].
A 3-ketosteroid-9α-hydroxylase was identified in R. erythropolis SQ1, which the 9α-hydroxylation of compounds 4-androstene-3,17-dione and 1,4-androstadiene,3-17-dione [94]. R. equi ZMU-LK19 was applied in the asymmetric hydroxylation and diastereoselective oxidation of (+)-2-substituted tetrahydroquinolines generating chiral 2-substituted-1,2,3,4-tetrahydroquinoline-4-ols and chiral 2-substituted-2,3-dihydroquinolin-4(1H)-ones [95].
2.5. C=C–Bond Reductases
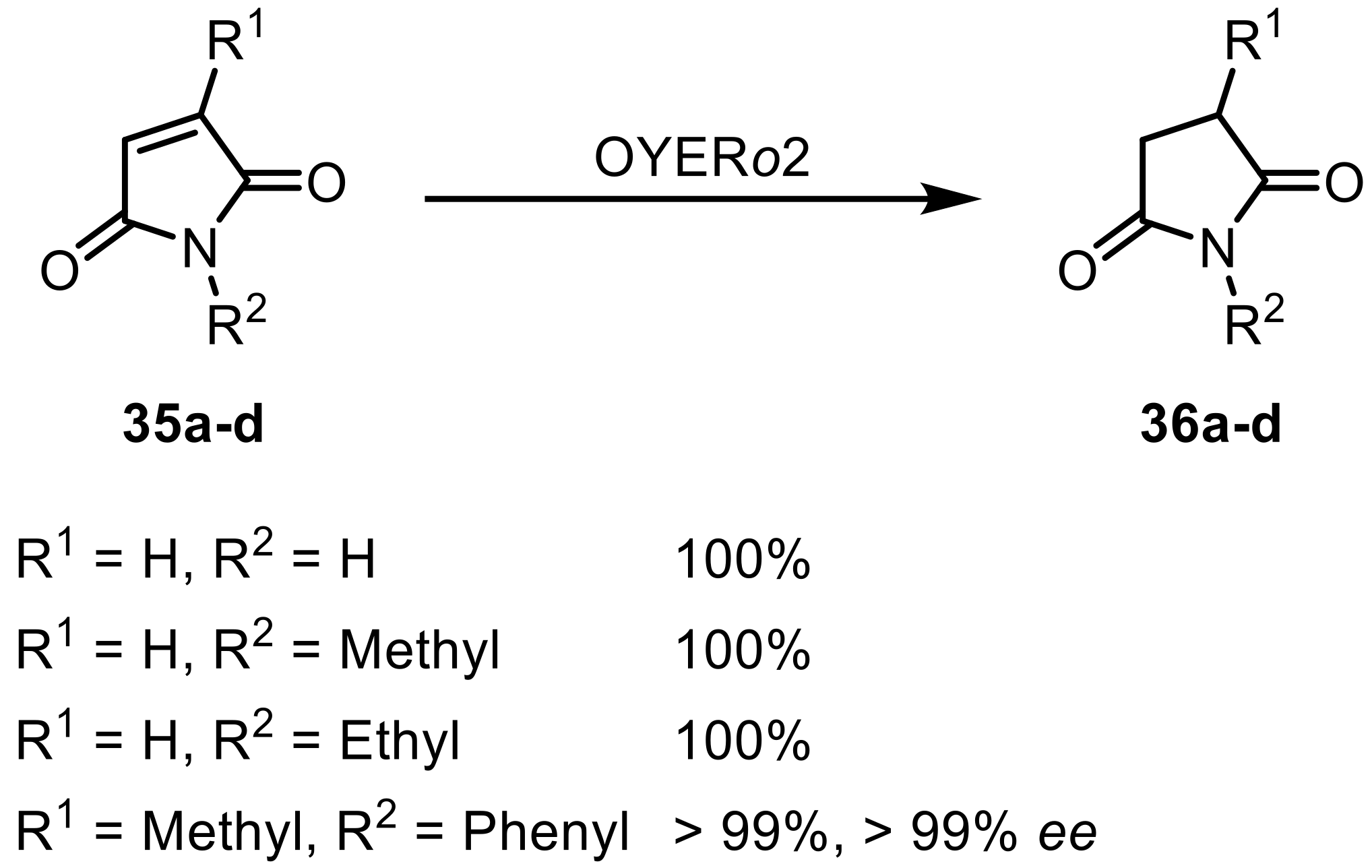
The selective reduction of C=C-double bonds, especially in α,β-unsaturated carbonyl compounds, is seen as a valuable reaction in the production of chiral building blocks and is catalyzed by flavin-dependent ene-reductases (EREDs, EC 1.3.1.31, also called Old Yellow Enzymes (OYE) [96]. Like many other reported organisms, Rhodococcus also showed the reduction-potential toward a diverse number of substrates. The reactivity toward seven chalcone-derivatives was screened for using R. sp. DSM 364 amongst other microorganisms [97]. Whole-cells of R. sp. DSM 364 catalyzed the reduction of all seven substrates including all derivatives with both electron withdrawing and electron donating groups, exclusively to the respective dihydrochalcones while leaving the carbonyl moiety unreacted. Several Rhodococcus strains (R. erythropolis and R. rhodochrous) were used in an investigation into their reduction potential toward activated ketones, an aldehyde, an imide, and nitro-compounds [98]. Based on the conversion of ketoisopherone to levodione, stereoselectivity studies have been performed: all so far have reported that ene-reductases found in plants, yeasts, bacteria, and parasites only gave access to the (R)-configurated product. However, in this study, whole-cell bioconversions of six out of seven strains led to the formation of the (S)-product. The same reaction was carried out with purified ene-reductases, which led to the (R)-product. It was therefore proposed that the whole-cells produce a mixture of both (R)- and (S)-levodione from which only the (R)-enantiomer is further converted by the other enzymes present in the whole-cell mixture leaving the (S)-levodione as main product unreacted [98]. A (R)-selective ‘thermophilic-like’ ene-reductase from R. opacus 1CP obtained by genome mining was heterologously expressed in E. coli and subsequently characterized [99]. Based on sequence similarity, this enzyme was categorized as a member of the ‘thermophilic-like’ (YqjM-like) OYE group, but it only showed a temperature optimum of 37 °C instead of higher temperatures of ≥70 °C, which are usually described for thermophilic enzymes. It showed the highest activities toward (substituted) maleimides (35) leading to the corresponding succinates (36) (Scheme 10).
2.6. Amino Acid and Amine Dehydrogenase
Amino acid (AADH, EC 1.4.1.x) and amine dehydrogenases (AmDH) catalyze the reductive amination of α-keto acids and ketones, yielding α-amino acids and amines, respectively, using NAD(P)H as a cofactor. Ammonia is mostly chosen as the nitrogen source.
R. sp. M4 hosts a phenylalanine dehydrogenase (PheDH), which primarily converts phenylpyruvate to l-phenylalanine via a reductive amination process [100]. Additionally, the enzyme was also shown to accept other (sterically demanding) α-keto acids such as 4-(methylsulfanyl)-2-oxobutanoic acid, 2-oxo-4-phenylbutanoate or 2-oxo-5-phenylpentanoate, making it interesting for its broad substrate tolerance [100,101].
The amine dehydrogenases (AmDH) are a recently developed group of enzymes derived from amino acid dehydrogenases [102]. They act on prochiral ketones, opening up a new synthetic route toward chiral amines. A new (R)-selective AmDH (TM_pheDH) was engineered from the Rhodococcus phenylalanine dehydrogenase by directed evolution [103]. With this new enzyme, it was possible to reduce phenylacetone (37a) and 4-phenyl-2-butanone, leading to (R)-amphetamine (38a) and (R)-1-methyl-3-phenylpropylamine with excellent ee values (>98%). The immobilization of this AmDH on magnetic nanoparticles (MNP) increased the productivity and stability compared to the free enzyme [104]. The enzyme was also found to be active toward o-methoxyphenylacetone derivatives (37b), aliphatic ketones, and so-called ‘bulky-bulky’ ketones such as 1-phenylbutan-2-one or 1-phenylpentan-3-one (Scheme 11) [105].
2.7. Desaturase
A Δ6-desaturase from a R. sp. KSM-B-MT66 mutant was applied in a two-phase system to catalyze a cis-desaturation of low-cost saturated starting material [106]. Next to unsaturated acyl fatty acids, chloroalkanes and simple alkanes were accepted and the dehydrogenation always took place 9-C-atoms away from the terminal methyl group. This reaction was used to generate intermediates for the preparation of substituted fatty acids used in the dermatological pharmacy and was run with a space-time-yield of 16.8 g/L per day. Recently, novel Δ6-desaturase enzymes were identified in R. sp. and their use to produce cis-6-hexadecenoic acid were patented for future applications [107,108].
3. Enzymes from the Aldoxime-Nitrile Pathway
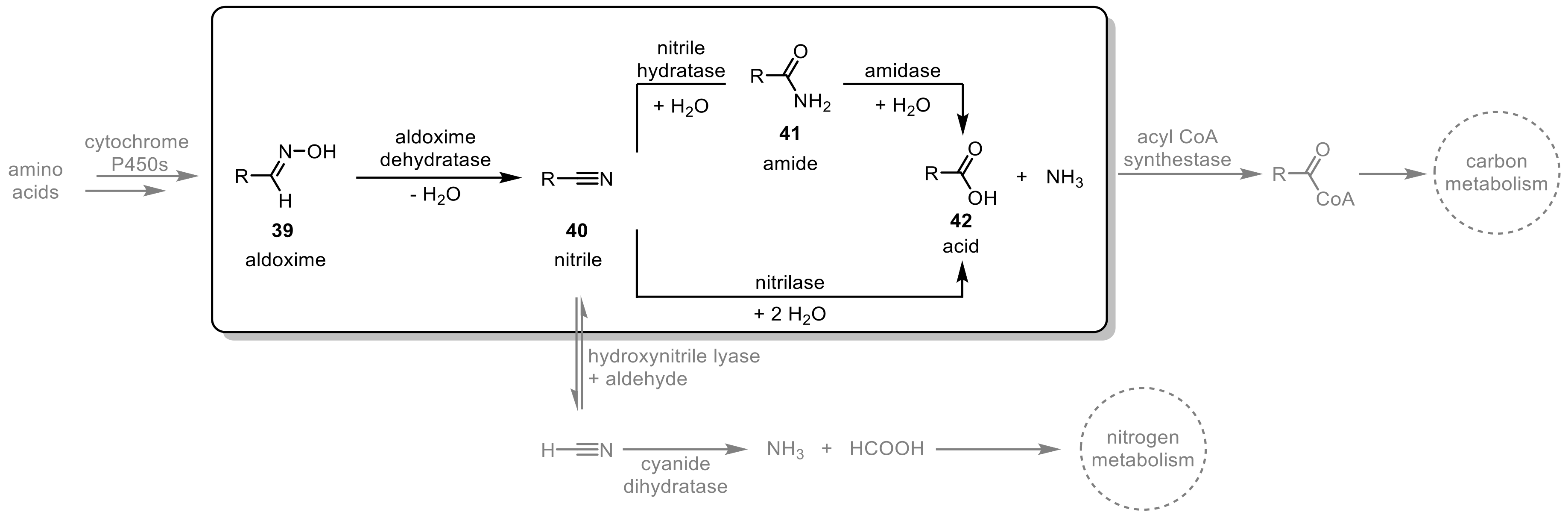
Enzymes present in the aldoxime-nitrile pathway catalyze both the synthesis and decomposition of nitrogen-containing organic compounds, thereby playing a key role in the carbon and nitrogen metabolism of microbes and plants (Scheme 12) [109,110,111]. Through the oxidation and decarboxylation of amino acids, aldoximes (39) are generated [112,113], which are subsequently dehydrated to give nitriles (40). These nitriles can either undergo a hydroxynitrile lyase-catalyzed decomposition reaction yielding hydrogen cyanide and aldehydes or they can be converted to carboxylic acids (42) via two possible routes: a one-step reaction catalyzed by nitrilases or via an amide intermediate (41) catalyzed by a coupled nitrile hydratase and amidase system [109,110]. The resulting acids and ammonia are afterward consumed in the carbon and/or nitrogen metabolism.
Various Rhodococcus species use the aldoxime-nitrile pathway and therefore the respective enzymes (aldoxime dehydratase, nitrilase, nitrile hydratase, and amidase) are present in many strains [111,114]. The versatile use of these enzymes in synthetic organic chemistry from a laboratory ‘proof-of-principle’ to multi-ton scale for industrial applications showcases the strength of Rhodococcus as a biocatalyst in this area.
3.1. Aldoxime Dehydratase
Nitriles are valuable starting points in the synthesis of both bulk chemicals and chiral pharmaceuticals [96,112]. Their synthesis, however, either requires high temperatures (ammoxidation) or the use of highly toxic hydrogen cyanide as a reagent. Aldoxime dehydratases (Oxd, EC 4.99.1.5) are a recently discovered group of enzymes found in bacteria [115,116] and offer an environmentally friendly, cyanide-free alternative to producing nitriles that start from easily accessible aldoximes, which can be produced by a condensation reaction of aldehydes with hydroxylamine [117].
Several studies have examined the substrate scope of aldoxime dehydratases from Rhodococcus for non-chiral aldoximes [116,118,119,120]: while arylaliphatic aldoximes were generally better converted by other organisms [121], strain Rhodococcus sp. YH3-3 was the only organism that showed activity against substituted aromatic and furan-derived aldoximes. It also displayed higher activities toward a number of substrates with heteroaromatic moieties compared to other Oxds, thereby showing unique properties [112,116]. Linear and branched aliphatic aldoximes with a chain length of C2 to C6 are accepted and converted to their respective nitrile by a number of ROxd [116,118,119]. A comprehensive study compared the activity of five heterologously expressed aldoxime dehydratases of which two originated from Rhodococcus strains (R. globerolus A-4, OxdRG; R. sp. N-771, OxdRE) toward chiral aldoximes including arylaliphatic, heteroaromatic aliphatic, cyclic aliphatic, and acyclic long-chain aliphatic aldoximes with an particular interest in the stereochemical course of the reaction [120]. Interestingly, in some cases, it was shown that depending on the choice of isomeric structure of the substrate (E- or Z-aldoxime), the enantiopreference in the final nitrile (R- or S-configuration) could be influenced.
A recent example displaying the growing industrial importance of this enzyme class can be seen in a filed patent from BASF, which describes the conversion of a number of terpenes—important odoriferous compounds in the fragrance industry. One example is the biocatalytic production of citronellyl nitrile (44), which is known to have a rose-like fragrance (Scheme 13) [122].
3.2. Nitrile Hydratase
While aldoxime dehydratases form valuable nitriles, nitrile hydratases (NHase, EC 4.2.1.84), which are mononuclear iron- or cobalt-dependent enzymes, belong to the group of nitrile-degrading enzymes yielding amides through hydration reactions. This reaction is often followed by an amidase-catalyzed hydrolytic step when applying Rhodococcus whole-cells, which converts the formed amides to their respective carboxylic acids. This section will summarize both the use of NHase as a single catalyst and systems using both enzymes as a coupled two-step system.
3.2.1. NHase as a Single Biocatalyst
The production of amides from nitriles has become crucial to industry and therefore the commercial interest in nitrile-degrading enzymes has gained immense attention, as many excellent reviews display [109,114,123,124]. Rhodococcus strains are industrially used to prepare amides essential for humankind such as acrylamide (R. rhodochrous J1, >400,000 t/a, Nitto Chemical Industry [5,6]) or nicotinamide (R. rhodochrous J1, >11,500 t/a, Lonza AG [45,125]). In this case, the nitrile-hydratase of industrially important strain R. rhodochrous J1 was shown to be cobalt-dependent [126]. The main products of acrylamide and its polymers are used as coagulators in the leather and textile industry while nicotinamide is one of the two forms of vitamin B3 used in the cosmetics industry and in animal feed supplementation [109,127].
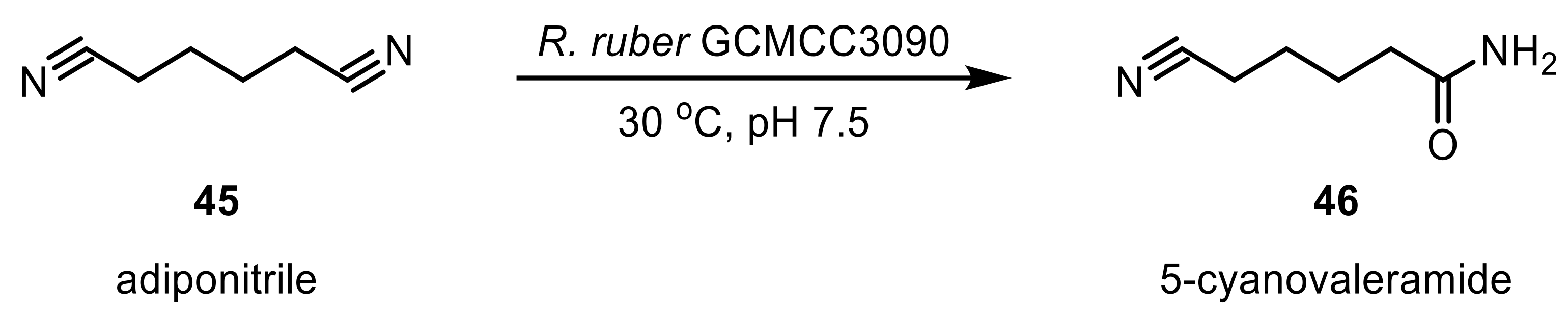
NHases are versatile enzymes that accept a broad range of different nitriles. The hydrolysis of aromatic and arylalkyl nitriles was intensively studied and proven successful for pyridyl-, pyrazinyl-, (substituted) benzyl-, furyl-, and thionyl-moieties [128,129,130,131,132,133] as well as trans-2,3-epoxy-3-aryl-propannitriles [134] or rac-mandelonitrile [135]. R. boritolerans FW815 was shown to have a strong 2,2-dimethyl-c-propanecarbonitrile (DMCPCN) hydratase activity in the absence of amidase activity, leading to an enrichment of 2,2-dimethyl-c-propanecarboxamide (DMCPCA)—an important precursor for the drug cilastatin, which is an inhibitor of a renal peptidase that is involved in the metabolism of other drugs, thereby making these other, combined drugs more effective [136]. Dinitriles are also accepted substrates: whole-cells of Rhodococcus sp. were shown to convert fluorinated aromatic dinitriles [137] and R. rhodochrous IFO 15564 was active toward alicyclic mono- and dinitriles, affording the products in low to moderate yields [138]. Resting cells of R. ruber CGMCC3090 converted the aliphatic adiponitrile to selectively give 5-cyanovaleramide (5-CVAM), which is used in the synthesis of caprolactam, a common precursor for Nylon 6 (Scheme 14) [139].
Prochiral substrates such as α- or β-substituted nitriles have also been used in bioconversions: starting from α-racemic aminonitriles (47), respective (R)-(50) and (S)-selective α-amino acids (51) were produced each using a three-enzyme cascade reaction [140]. In both reaction pathways, a NHase from R. opacus 71D was first applied to give a racemic mixture of α-amino acid amides (48/49). A dynamic kinetic resolution catalyzed by ACL racemase and subsequently either d-aminopeptidase ((R)-amino acid (50)), or l-amino acid amidase ((S)-amino acid (51)) yielded the final products (Scheme 15).
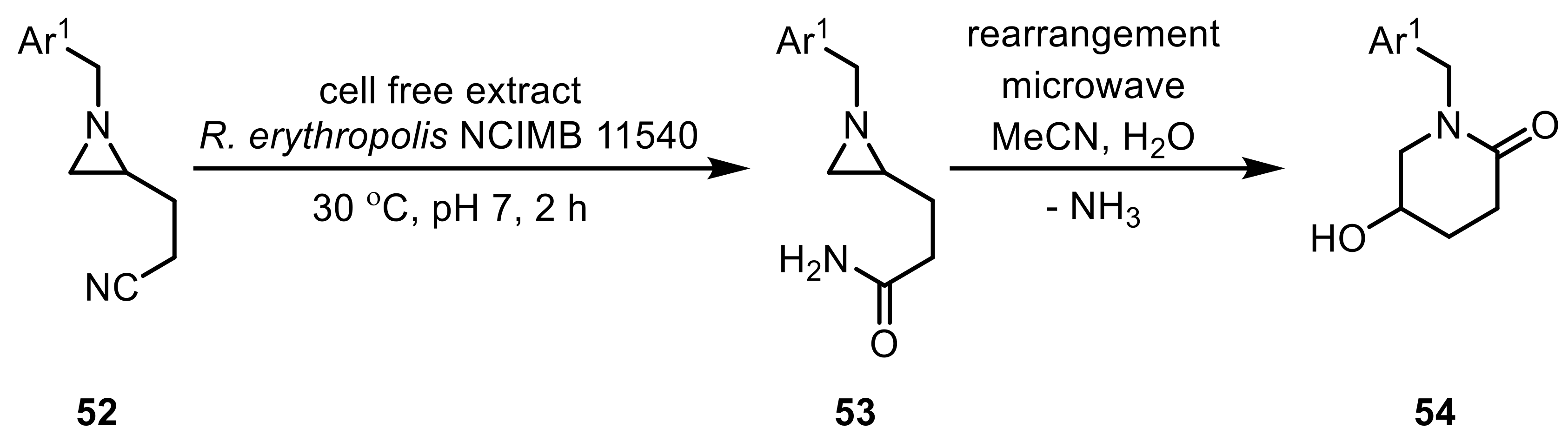
On the other hand, a partially purified NHase was successfully applied in the conversion of β-substituted nitriles such as 3-oxonitriles, 3-hydroxynitriles, and 3-(acyloxy)nitrile, yielding the corresponding amides in moderate to good yields [141]. Whole-cells of R. erythropolis NCIMB 11540 on the other hand were used in the synthesis of piperidin-2-ones (54) starting with a NHase-catalyzed hydration reaction of 1-(arylmethyl)-2-(2-cyanoethyl)aziridines (52) (Scheme 16) [142].
3.2.2. Two-Enzyme Systems
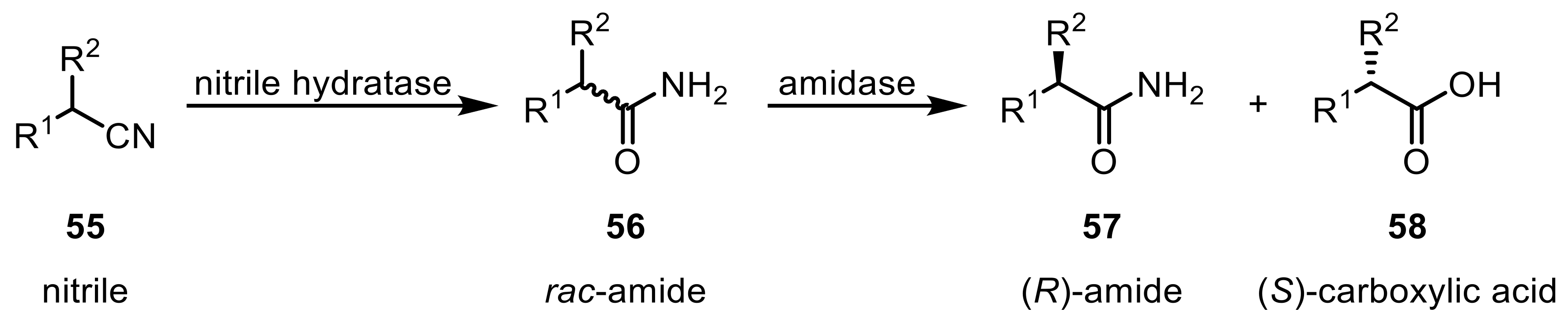
Most of the Rhodococcus strains used in these bioconversions express both NHase and amidase, which can lead to the direct follow-up reaction catalyzed by amidases yielding valuable carboxylic acids. Nitrile hydratases generally show low stereoselectivities. The amidase hydrolysis reaction, however, is usually very stereospecific, leading to the (S) carboxylic acid (58) and leaving the unreacted (R) amide (57) behind (Scheme 17) [143,144]. Many examples have successfully used the presence of both enzymes to achieve the production of both selective amides and carboxylic acids by kinetic resolution.
When starting with prochiral α-substituted nitriles, the synthesis of optically active (R)-amides and (S)-carboxylic acids was achieved. Several 2-aryl-alkylnitriles were accepted substrates [145,146] as was rac-naproxen nitrile, which directly formed the desired and biologically active drug (S)-naproxen [147]. Additionally, enantioenriched 3-aryl-3-hydroxy-2-methylene carboxylic acids and amides were prepared from the respective nitriles using whole-cells of R. rhodochrous AJ270 [148].
Multiple examples of dinitriles were also applied: incubation of R. erythropolis AJ270 cells with meso-c-pentane-1,3-dinitriles yielded optically active amide-products in high yields [149]. Via an amidase-catalyzed kinetic resolution step and following treatment with CH2N2, monocyano amides were resolved into (-)-amide and (-)-ester. Additionally, it was shown that the amidase also catalyzes the desymmetrization of meso-c-pentane-1,3-dicarboxamides, affording enantiopure pentanecarboxylic acids.
Malononitriles are α, α-substituted dinitriles that form malonic diamides and malonic acid monoamides upon incubation with whole-cells of R. rhodochrous IFO 15564 [150]. Next to that, cyanohydrins (α-hydroxy nitriles) were also successfully hydrolyzed, leading to enantiopure α-hydroxy carboxylic acids like the pharmaceutical intermediate (R)-chloromandelic acid on gram-scale [151] as was a large variety of aminonitriles ranging from α-aryl-, α-alkyl-substituted glycine nitriles [152,153,154] to aziridine-2-carbonitriles [155], azetidine-2-carbonitriles, and 4-oxoazetidine-2-carbonitriles [156,157].
Aside from α-substituted nitriles, β-substituted nitriles are also suitable substrates for the NHase/amidase system, yielding interesting building blocks with more remote stereogenic centers. Next to several studies focusing on 3-substituted glutaronitrile derivatives [158,159,160], 3-hydroxy-arylpropanenitriles [161], 3-hydroxy-4-aryloxybutanenitriles [161,162], and 3-(benzoyloxy)pentanenitrile [163] were also converted using Rhodococcus cells. A number of Rhodococcus strains was also shown to be active on 3-benzyl-oxy- and 3-benzoyloxypentanedinitriles [88,159,160,161,164], which opens up a new synthetic route to the drug Atorvastatin (trade name Lipitor by Pfizer), an HMG-CoA reductase inhibitor for lowering blood cholesterol [165].
The above-mentioned examples all successfully exploit the presence of both enzymes to obtain amides and carboxylic acids. However, the amidase also causes unwanted side-product formation in other reactions when the intermediate amide is the desired final product. To avoid the amidase-catalyzed follow-up reaction in the preparation of acrylamide, an amidase-negative mutant was designed using a genetic knock-out technique. This approach increased the acrylamide yield by 25% compared to the wild-type strain [166].
3.3. Amidase
Amidases (EC 3.5.1.4) belong to the group of amidohydrolases and have been widely researched due to the biological relevance of amides in nature. They generally show a broad substrate scope and good enantioselectivity [167]. The examples discussed in the above section already display the versatility of Rhodococcus amidases and their synthetic potential in combination with nitrile hydratases. However, this section highlights specific examples achieved with single-amidase systems.
An amidase from R. sp. MTB5 showed good activity toward a large number of aliphatic, aromatic, and heterocyclic amides [168]. Diamides and specific amino acids were, however, only poorly accepted. α-alkyl phenylacetamides have been kinetically resolved using an amidase present in R. sp. AJ270 [169]. (S)-naproxen can be produced starting from both racemic naproxen nitrile using the two-enzyme system [147] as well as from a racemic naproxen amide using cells of R. erythropolis MP50 [170]. In a similar manner, R. erythropolis AJ270 catalyzed the enantioselective hydrolysis of racemic ibuprofen amide leading to the biologically active compound [171].
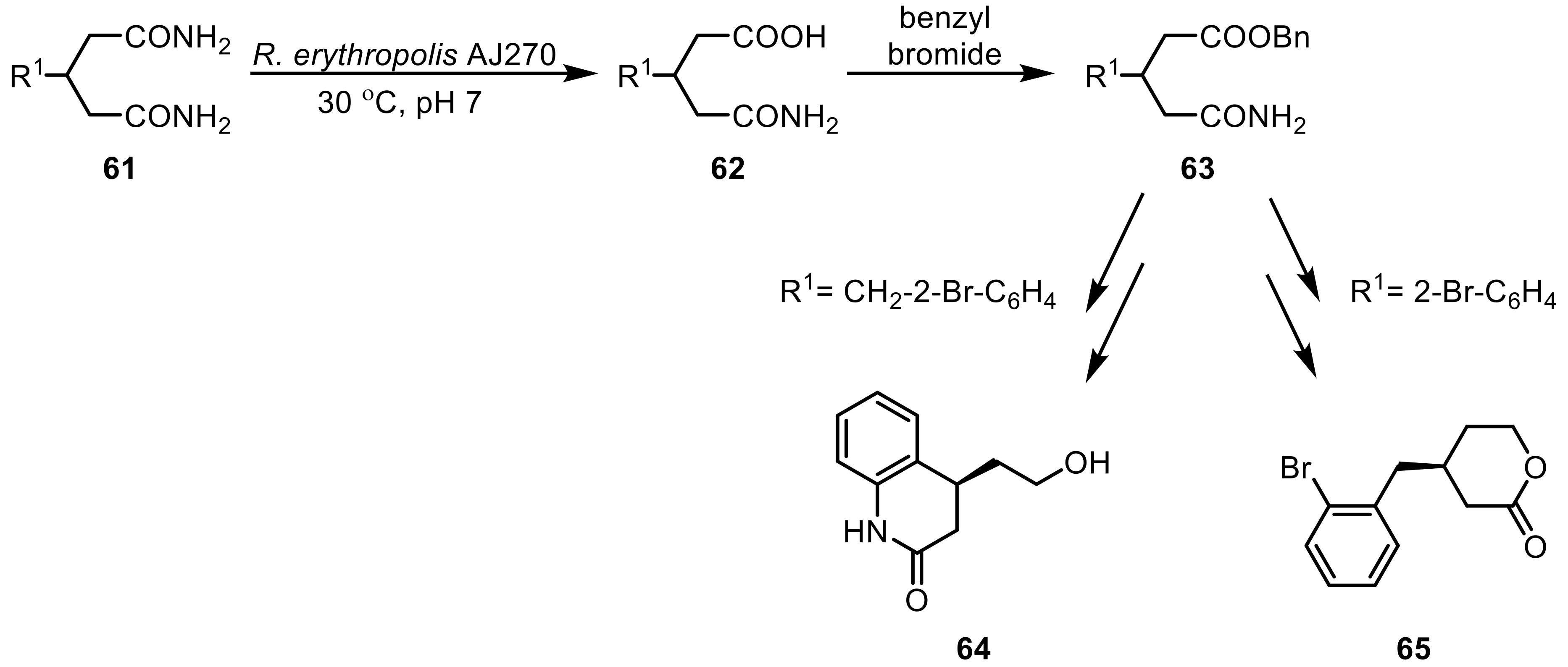
In the production of (R)-malonamic acids, amidases from Rhodococcus have proven to be successful tools. Whole-cells of R. sp. CGMCC 0497 converted aromatic α,α-disubstituted malonamides with substituents in ortho-, meta-, and para-position with high yields and excellent enantioselectivities as well as dialkyl-substituents with slightly lower ee-values [172]. The amidase present in whole-cells of R. erythropolis AJ270 transformed a number of prochiral malonamides yielding a range of carbamoylacetic acid products [173]. α-substituted α-amino-malonamides (59) were transformed to highly functionalized tetrasubstituted α-amino acids (60) with high yields with excellent ee-values by the same organism (Scheme 18) [174].
Interestingly, this organism was also used to catalyze the desymmetrization of both five-membered meso-N-heterocyclic- and meso-c-pentane dicarboxamides to afford functionalized enantiopure derivatives of pyrrolidine, dihydropyrrole, and piperidine as well as c-pentanecarboxylic acids [175,176]. The same amidase was reported to also hydrolyze a palette of prochiral 3-aryl- and 3-arylmethyl glutaramides (61), leading to highly enantioselective glutaric acid monoamide derivatives (62) [177]. This amidase-catalyzed hydrolysis paved the way for a straightforward chemo-enzymatic synthesis of dihydroquinolinone (64) and a δ-lactone (65), thereby showcasing the many synthetic applications of this biocatalyst (Scheme 19).
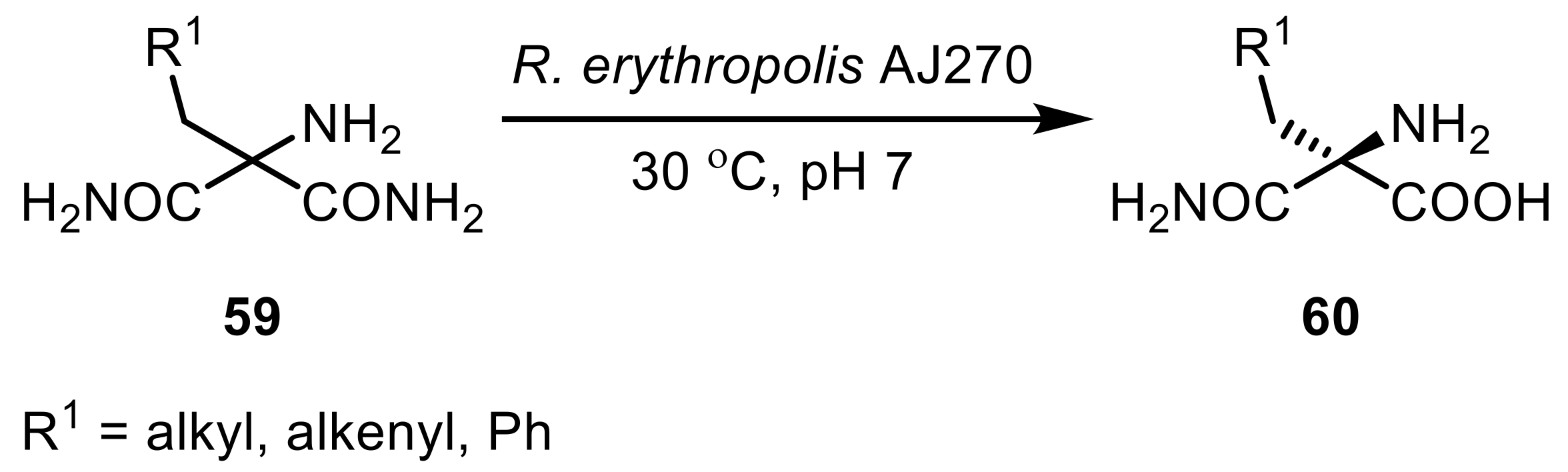
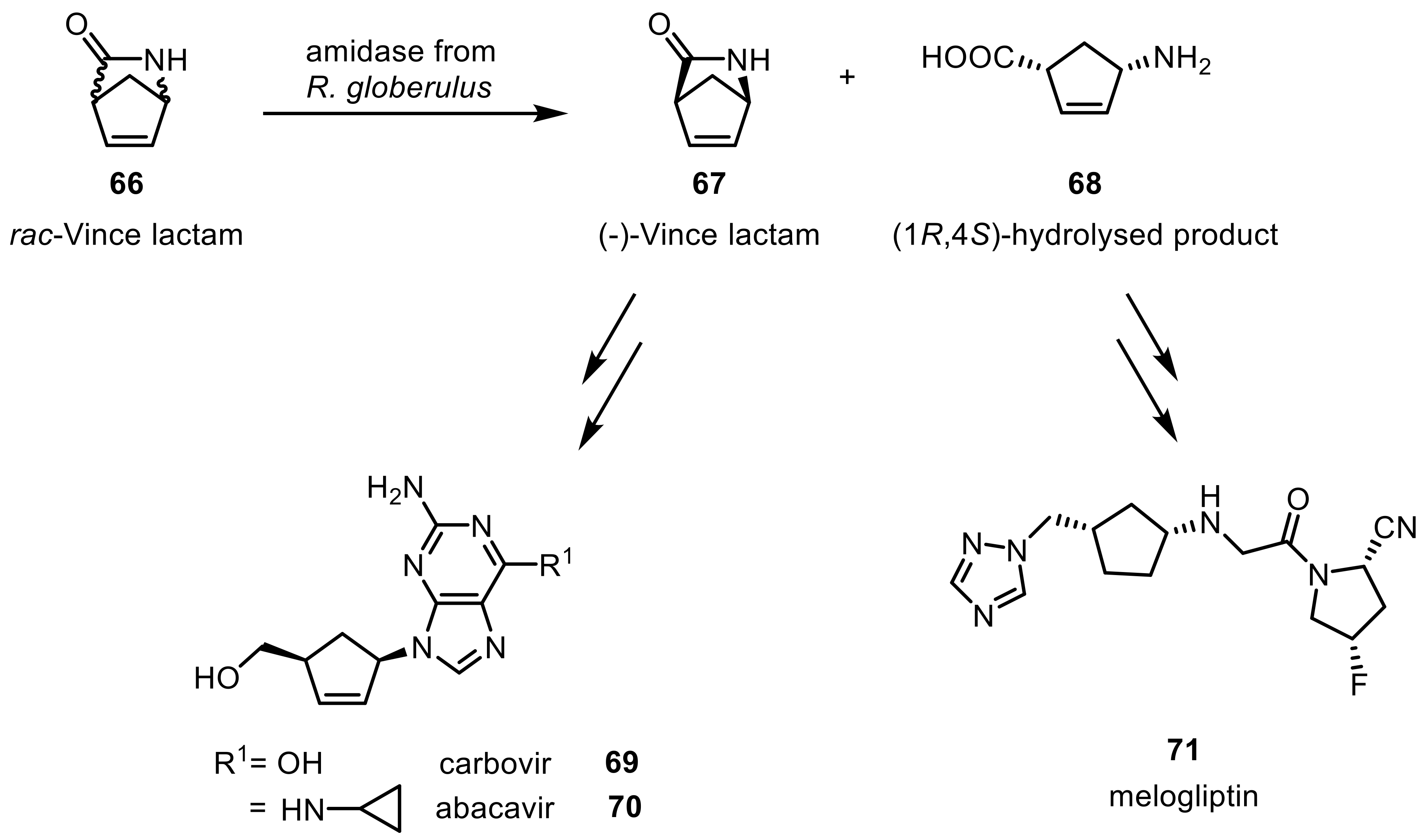
Lactams, cyclic analogues of amides, are generally more stable and their hydrolysis therefore often requires harsher reaction conditions leading to the respective amino acids [167]. Nonetheless, certain amidases (i.e., lactamases from Rhodococcus) were effectively used in the hydrolysis reaction. The hydrolysis of Vince-lactams (66) (2-azabicyclo[2.2.1]hept-5-en-3-one) and their derivatives have received increased attention due to the fact that the two resulting products are valuable synthons in the pharmaceutical industry for the manufacture of antiviral agents carbovir (69) and abacavir (70) or for the antidiabetic melogliptin (71) [178]. R. equi NCBI 40312 showed activity against Vince-lactam [179], and an amidase from R. globerulus presented excellent enantioselectivities for the obtained amino acid products (Scheme 20) [180].
3.4. Nitrilase
While all of the above-mentioned enzymes from Rhodococcus present in the aldoxime-nitrile pathway have been intensively studied and applied in many different processes and reactions, nitrilases (EC 3.5.5.1, or nitrile aminohydrolase) from Rhodococcus are less prominent and other microorganisms (Synechocystis, Alcaligenes, Pseudomonas, etc.) clearly dominate this area of research. However, a small number of reactions have been described using Rhodococcus whole-cells or purified enzymes.
Bioconversions of acrylonitrile with R. rhodochrous J1 yielded acrylic acid with exceptional yields of 390 g/L [183], which were even further improved with a mutant strain of R. rhodochrous tg1-A6 leading to 414.5 g/L [184].
The production of ammonium acrylate by two Rhodococcus isolates was further investigated and compared [185]. While R. erythropolis 3843 expressed a nitrile hydratase/amidase system, R. ruber NCIMB 40757 expressed a nitrilase. In this comparative study, the single-enzyme nitrilase-system was superior to the two-enzyme carboxylic acid production due to favorable reaction kinetics and because the NHase and amidase do not share the same temperature optimum.
R. rhodochrous PA-34 was shown to hydrolyze α-aminonitriles to obtain optically active l-amino acids (amongst others l-leucine, l-valine and l-methionine) as well as d-alanine [186]. Furthermore, strains R. rhodochrous J1 and R. sp. NDB 1165 were used to generate nicotinic acid, also called niacin, starting from 3-cyanopyridine with yields of 172 and 196.8 g/L, respectively [187,188]. Nicotinic acid is, next to nicotinamide, a form of vitamin B3 and its synthesis is therefore commercially interesting.
It is worth mentioning that even though only a small number of nitrilase-catalyzed reactions with Rhodococcus have been described, this nitrile-degrading enzyme class in general is well-studied and applied in a multitude of reactions, which can be followed-up on in several comprehensive reviews and books [114,189,190,191].
4. Hydrolase Activity in Rhodococcus
4.1. Epoxide Hydrolase (EC 3.3.2.x)
Epoxide hydrolases (EHs) catalyze the ring-opening reaction of oxirane rings, leading to vicinal diols and enantiopure epoxides as the final products. Most valuable are EHs that either show high enantioselectivities or a low enantiopreference with high levels of enantioconvergence. Many studies have focused on the asymmetric hydrolysis of geminal (2,2-disubstituted) oxiranes using Rhododoccus as a biocatalyst, whereby all investigated EHs hydrolyzed the (S)-enantiomer forming an (S)-diol and leaving the (R)-epoxide behind [192]. The investigated substrate scope includes linear alkyl-, alkenyl-, alkynyl- or benzylsubstituted epoxides. While R. ruber DSM 43338 acted on 2-methyl-2-(aryl)alkoxyoxiranes [193], R. sp. NCIMB 11216 was shown to convert (±)-2-methyl-2-alkyl-epoxides [194]. Oxiranes bearing an alkene or alkyne function were particularly well transformed using the same strain [195]. Additionally, two Rhodococcus strains isolated from marine sediments were used in the hydrolysis of styrene oxide and its chlorinated derivatives [196].
R. erythropolis DCL14 bears an epoxide hydrolase that holds a particular position within the family of EHs. While most of the described EHs belong to the α,β-hydrolase fold superfamily, the so-called ‘limonene-1,2-epoxide hydrolase’ (LEH) shows a 3-dimensional structure that is dissimilar to the others [197]. Consequently, it follows a different reaction mechanism, which was proposed as a single step ‘push–pull’ mechanism rather than a multiple-step mechanism described for members of the α,β-hydrolase fold superfamily [192]. Initial investigations showed a narrow substrate range for the wild-type enzyme. Only the natural substrate limonene-1,2-epoxide was converted with excellent stereoselectivity while several alicyclic and few 2-methyl-1,2-epoxides showed only reasonable stereoselectivities [198,199]. With an iterative saturation mutagenesis strategy, three LEH variants were developed with an expanded substrate range and opposite stereoselectivities, making this enzyme more attractive for future applications [200].
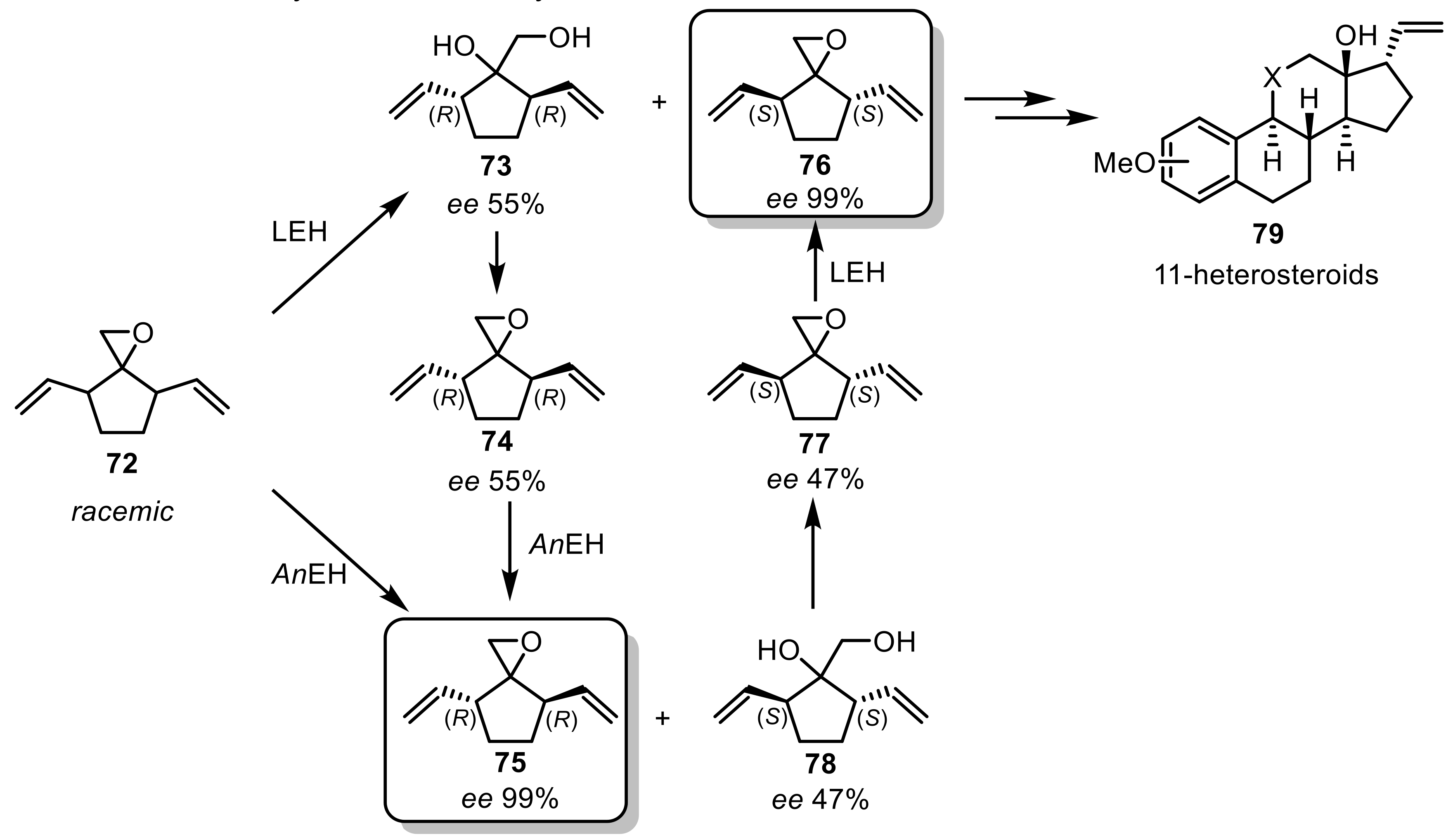
A preparative hydrolytic kinetic resolution of rac-trans-spiroepoxides was first achieved with EHs from Aspergillus niger (AnEH) and wild-type LEH from R. erythropolis DCL14. In the kinetic resolution of rac-trans-spiroepoxides (72), the two employed EHs exhibited opposite enantioselectivities, and by applying both enzymes in a combined two-step process, both enantiomers (75,76) were generated with excellent enantiomeric excess values and in high yields: the racemic spiroepoxide (72) was, for example, first hydrolyzed by LEH leading to the (R),(R)-diol (73) and (S),(S)-epoxide (76). Through a chemical recyclization the (R),(R)-diol (73) was transformed into (R),(R)-epoxide (74), keeping the moderate enantiopurity. The spiroepoxide (74) was then subsequently resolved by the enantiocomplementary EH from A. niger. Enantiopure spiroepoxides are seen as valuable synthons in the synthesis of 11-heterosteroids (Scheme 21) [201].
The asymmetric hydrolysis of (±)-2,3-cis- and trans-disubstituted oxiranes was performed by Rhodococcus ruber DSM 43338 with excellent enantioselectivities reaching 95% and 86% ee, respectively [202].
Next to geminal oxiranes, tri-substituted epoxides were also successfully converted. In an enantioconvergent fashion, whole-cells of R. ruber SM 1789 catalyzed the hydrolysis of trialkyl-oxiranes achieving high yields of the respective (R)-configured vicinal diols with high ee values [203]. Expanding the epoxide hydrolase system to substrates bearing olefinic side-chains enabled the straightforward asymmetric synthesis of natural compounds myrcenediol and 7,7-dimethyl-6,8-dioxabicyclo[3.2.1]octane, which is known to be a volatile contributor to the aroma of beer in a chemoenzymatic reaction procedure [204]. In another enantioconvergent asymmetric synthesis using an EH from Rhodococcus, it was possible to synthesize the monoterpenoid coumarin (R)-(+)-marmin with 95% ee under anaerobic reaction conditions [205].
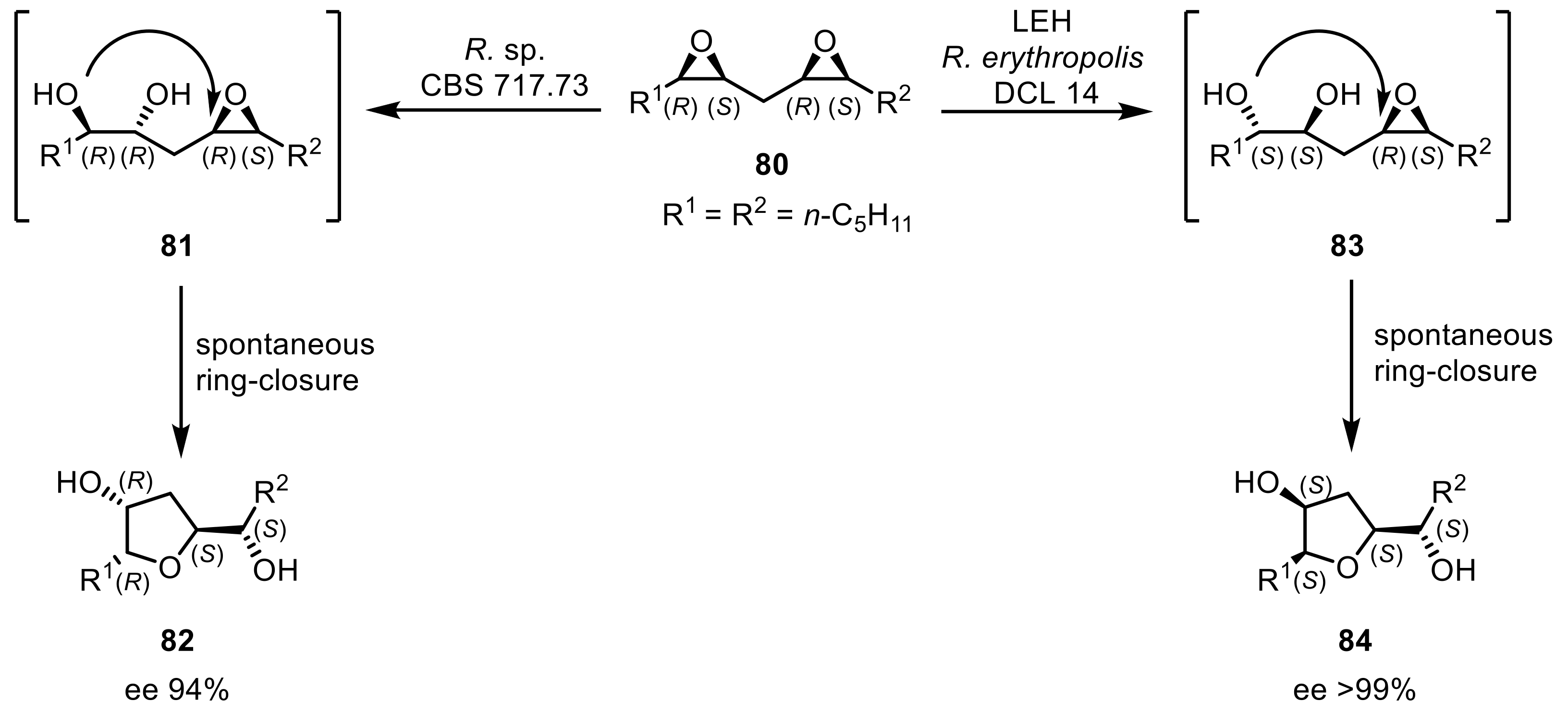
The biocatalytic desymmetrization of meso-epoxides with EHs was the focus of other studies: an EH from R. ruber ML-0004 was expressed in E. coli that showed a selective activity toward cis-epoxysuccinate producing enantiopure l-(+)-tartaric acid [206]. An enzyme-triggered hydrolysis-cyclisation reaction was observed when transforming methylene-interrupted meso-bis-epoxide (80) (6,7:9,10-bis(epoxy)pentadecane) with whole-cells of R. sp. CBS717.73 (Scheme 22). With this reaction, in total, four stereocenters were established in the obtained THF-derived product (82) [207]. Expansion of this study showed that four different THF-derivatives (representative 82,84) each with excellent ee and de values were achieved by using EHs from multiple organisms (e.g., Rhodococcus species, Solanum tuberosum, Aspergillus niger, Mycobacterium tuberculosis). The stereospecificities of the enzymes thereby determined the final stereoselectivities of the products while the cyclization reaction follows Baldwin’s rule without the operation of a cyclase [208].
The three aforementioned variants of LEH obtained by an iterative saturation mutagenesis approach were applied in the hydrolytic desymmetrization of several meso-epoxides and cis-1,2-homodesubstituted meso-epoxides forming both the (R,R)- and (S,S)-diols with very good ee values [200].
The resolution of rac-2-methylglycidyl benzyl ether, a versatile building block, was achieved through a whole-cell bioconversion of R. ruber SM 1789 yielding (R)-vicinal diol [209]. Following this study, several other strains were tested on the same substrate. While most Rhodococcus strains showed (S)-selectivity and acted with the retention of the stereoselectivity, the limonene epoxide hydrolase from R. erythropolis showed an inversion of the substrate, leading to a homochiral mixture [210]. R. ruber CBS 717.73 catalyzed the hydrolysis of 2-benzyloxymethyl-2-methyloxirane, another protected epoxy-alcohol, and was applied in a chemoenzymatic route to produce (S)-chromanemethanol, an important building block in Vitamin E synthesis, in 26% overall yield and 99% ee [211].
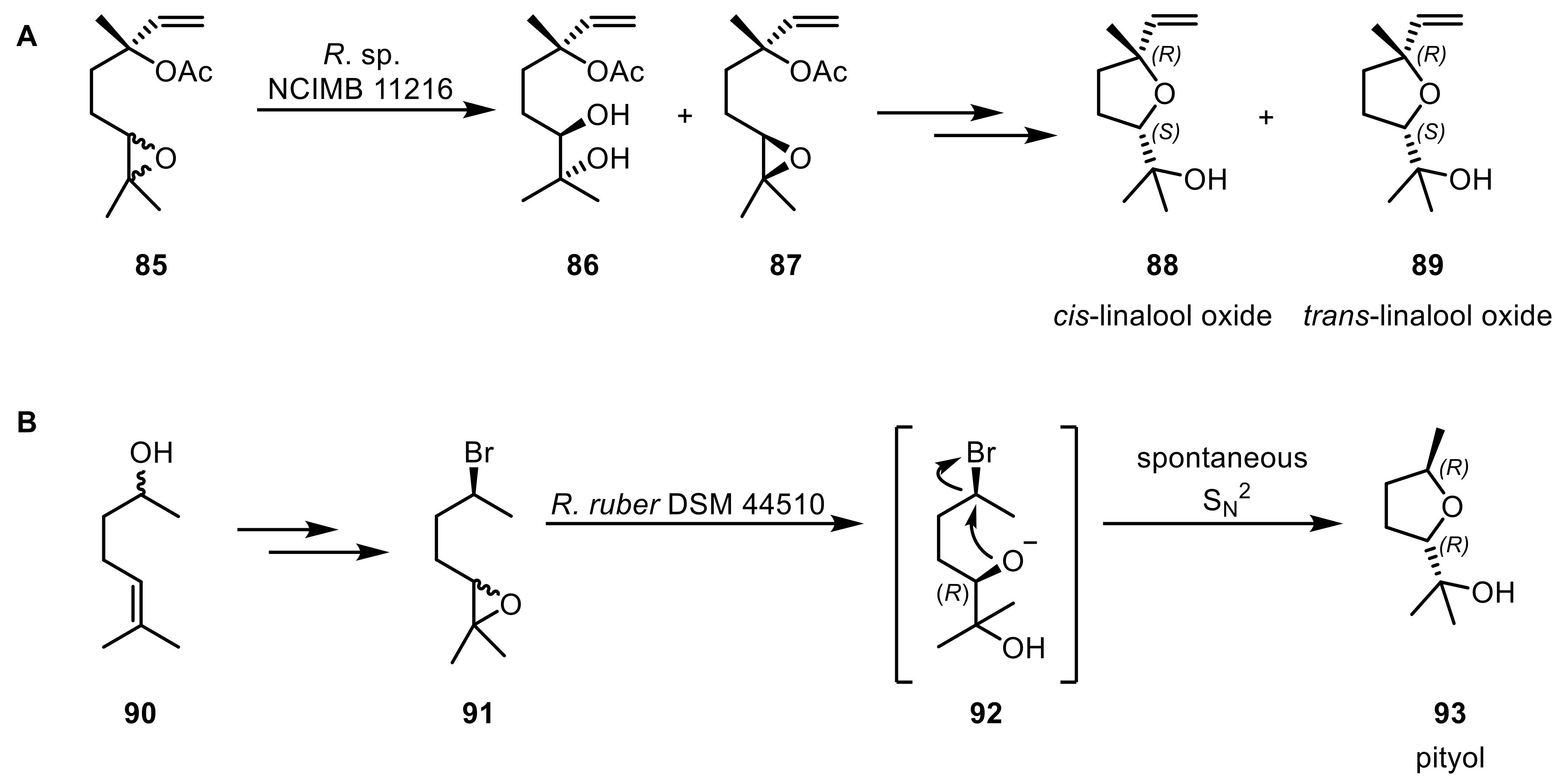
As portrayed, epoxide hydrolases from Rhodococcus are biocatalysts forming valuable synthons that can be used in a multitude of follow-up reactions. Next to the described application potential, Rhodococcus was also utilized in the production of other natural products such as linalool oxides (88,89) or pityol (93). Cis-(88) and trans-linalool (89) oxides were obtained via a chemo-enzymatic route starting from (3RS,6R)-2,3-epoxylinalyl acetate (85) with the R. sp NCIMB 11216 mediated resolution of the diastereomeric mixture of the starting compound as a key step (Scheme 23A) [212]. (2R,5R)-pityol (93), a bark beetle pheromone, was synthesized starting from (±)-sulcatol (90) via an lipase-catalyzed deracemization and a subsequent EH-catalyzed diastereoconvergent hydrolysis of a haloalkyl oxirane (91) (Scheme 23B) [213].
4.2. Esterase Activity
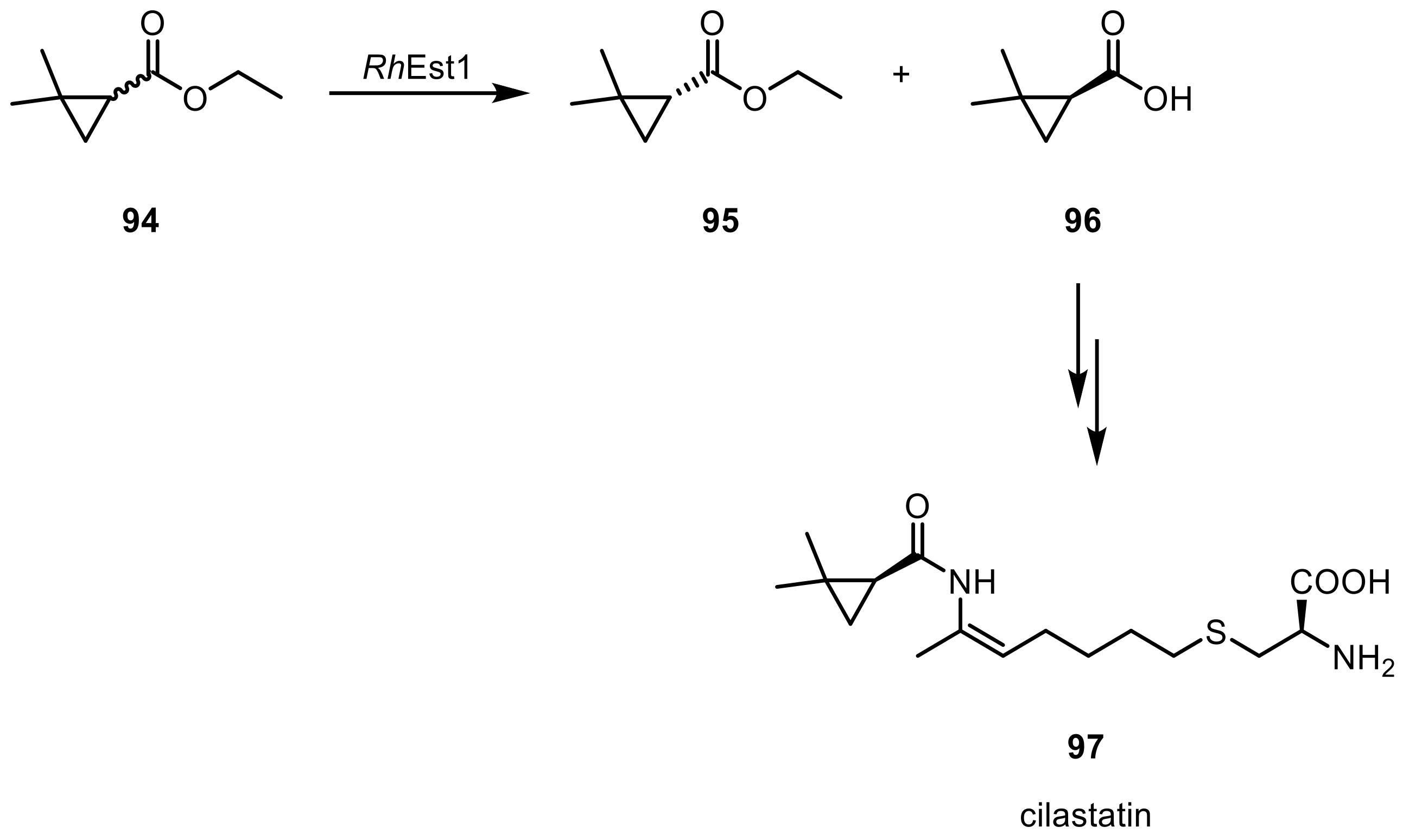
Few reports have concentrated on the application of other hydrolases from Rhodococcus. Amongst them, an esterase from R. sp. ECU1013 (RhEst1) was shown to hydrolyze rac-ethyl-2,2-dimethyl-c-propanecarboxylate (94, rac-DMCPCM) to give (S)-(+)-2,2-dimethyl-c-propylcarboxylic acid (96, (S)-DMCPCA)—a valuable precursor in the synthesis of the drug cilastatin (97) (Scheme 24) [214,215]. This shows an alternative route to cilastatin compared to the previously mentioned nitrile hydratase-catalyzed process starting with 2,2-dimethyl-c-propanecarbonitrile [136].
The production of both (S)- and (R)-linalool was achieved with two (partially) purified enzymes ((S)- and (R)-linalyl acetate hydrolase, respectively) from R. ruber DSM 43338 via the hydrolysis of the corresponding acetate esters [216]. An urethane hydrolase from R. equi TB-60 hydrolyzed a diverse range of compounds including anilides, amides, and esters such as toluene-2,4-dicarbamic acid butyl ester (TDCB) [217].
5. Hydratase Activity
Hydratases catalyze the addition of water to (un)-activated double bonds to form valuable enantiopure alcohols. It can be stated that nitrile hydratases from Rhodococcus have been profoundly investigated and were the center of research regarding hydratase activity. However, several Rhodococcus strains have shown an intriguing hydration potential toward other diverse compounds and their development is on the rise.
5.1. Oleate Hydratase
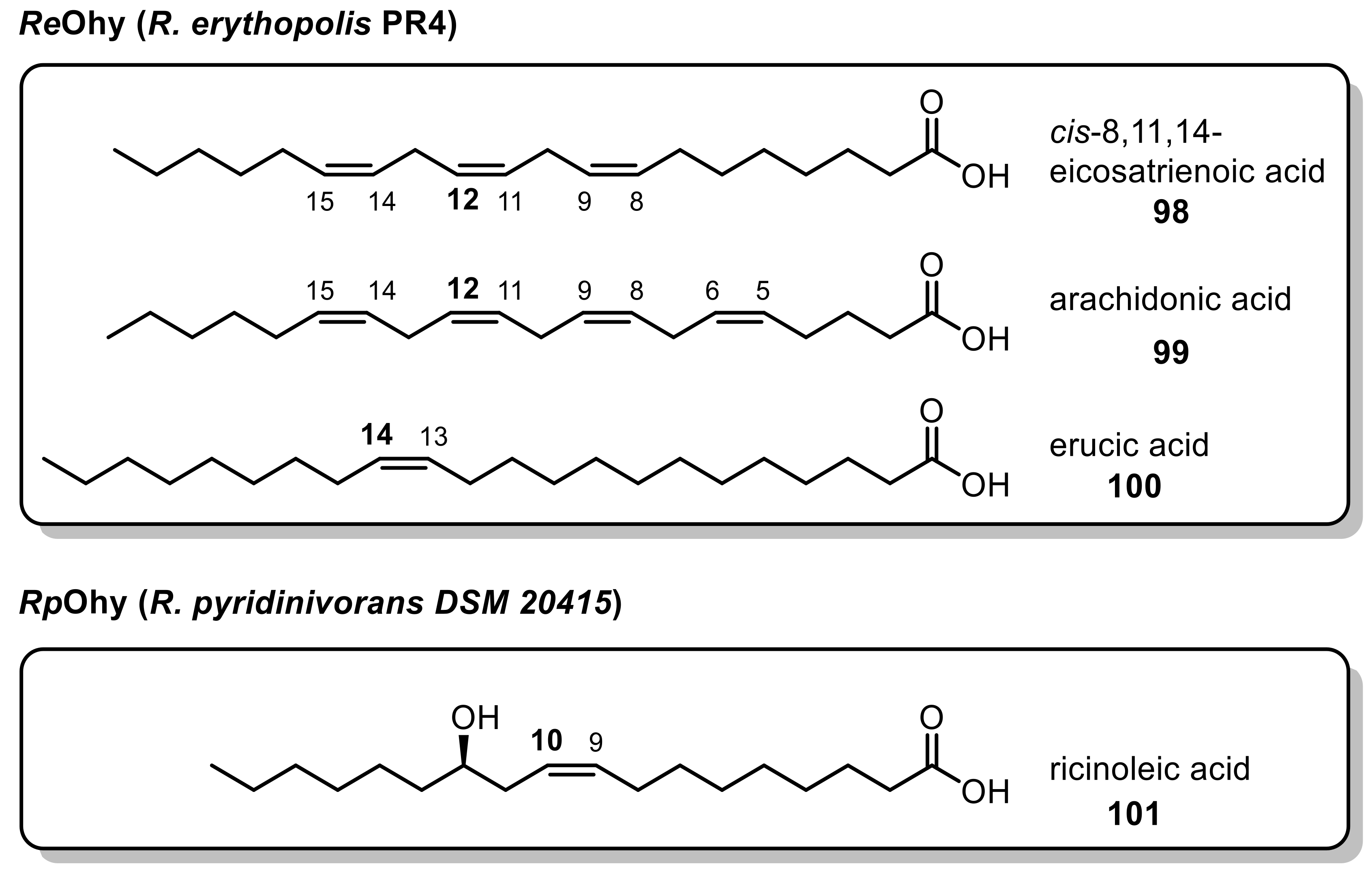
Recently, a novel oleate hydratase (Ohy, EC 4.2.1.53) was isolated from R. erythropolis CCM 2595 and characterized [218]. Structural analysis showed that this Ohy differed structurally from the previously described bacterial Ohys. This enzyme was shown to be an active monomer while all other characterized Ohys showed dimeric structures instead [219,220,221]. The purified Ohy was shown to accept a small range of unsaturated fatty acids, adding water exclusively in the 10-position while more complex lipids were not converted [218]. In-silico analysis of the occurrence and phylogenetic relationship of annotated oleate hydratases in 43 Rhodococcus strains revealed that distinct Rhodococcus clades showed Ohy potential, thereby discovering 19 novel oleate hydratases [222]. Two representatives, Ohys from R. erythropolis PR4 and R. pyridinivorans DSM 20415, sharing a sequence similarity and identity of 46% and 30%, respectively, were heterologously expressed and tested on (multiple) unsaturated fatty acids. While both Ohys converted smaller fatty acids in the same manner, they showed a complementary substrate scope toward sterically demanding and longer fatty acids (Figure 1).
A recently filed patent involves the Ohy-catalyzed conversion of free fatty acids such as oleic acid derived from the renewable feedstock of bio-based oils [223]. This is one example of the successful use of a novel Ohy from Rhodococcus that further improves the potential application of Rhodococcus as a useful biocatalyst.
5.2. Michael Hydratase
Whole-cells of R. rhodochrous ATCC 17896 were shown to catalyze a so-called Michael addition of water to α,β-unsaturated carbonyl compound (E)-4-hydroxy-3-methylbut-2-enoic acid and its respective ethyl-derivative [224]. Upon water addition, an internal nucleophilic attack leads to the formation of useful synthon (R)-4-hydroxy-4-methyldihydrofuran-2(3H)-one. The stereochemical course of the reaction was investigated, showing that the water-addition takes place in syn-fashion [225,226]. Interestingly, this hydratase demonstrated unusual behavior as it requires oxygen for higher activities, but labeling studies with D2O and 18O2 proved a true water addition instead of an oxidative process [226]. Unfortunately, this membrane-associated enzyme has not been isolated and the coding gene remains elusive up to this point.
6. Rhodococcus Acting on Sulfur-Containing Compounds
6.1. Sulfatase
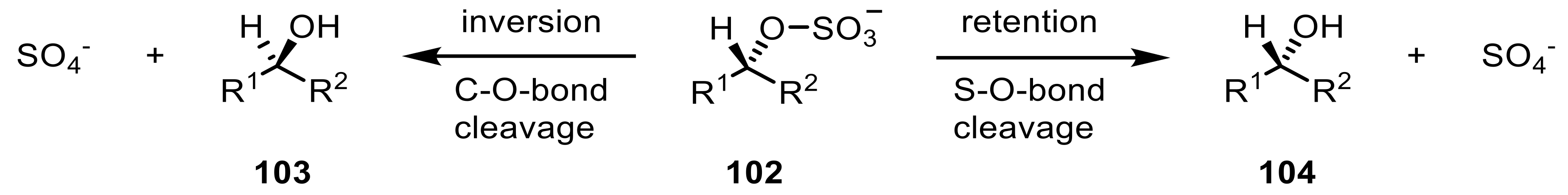
Sulfatases (EC 3.1.6.x) are a group of enzymes that catalyze the hydrolytic cleavage of sulfate esters by releasing inorganic sulfate and the corresponding alcohols [227]. Fascinatingly, sulfatases can achieve an enantio-convergent transformation of racemic sulfate esters (102) into only one stereoisomeric secondary alcohol yielding up to 100% yield theoretically, thereby surpassing traditional kinetic resolution processes. Sulfatases thereby determine not only the enantioselectivity, but also the stereoselectivity with respect to the retention (104) or inversion (103) of the configuration (Scheme 25) [228].
The hydrolysis of an alkyl-sulfate through the breakage of the S-O-bond by nucleophilic attack leads to a retention while the breakage of the C–O-bond on the other hand leads to an inversion of the configuration at the carbon atom bearing the center of chirality [228]. R. ruber DSM 44541 was shown to have a secondary alkyl sulfatase, called RS2, which acts through strict inversion. Methyl- and ethyl-(alkyl)sulfate esters with a varying chain-length were shown to be hydrolyzed [229,230]. The addition of Fe3+-ions led to an increase in activity, which, however, was not suitable for preparative scale reactions [231,232].
While most of the investigations concentrated on secondary alkyl-sulfates, few reports about cyclic sulfates have come forward. Growing cells of R. sp. CCZU10-1, however, were used to transform four cyclic sulfates: 1,3-propanediol cyclic sulfate (1,3-PDS), 1,2-propanediol cyclic sulfate (1,2-PDS), ethylene sulfate, and glycol sulfate [227]. All four cyclic sulfates were hydrolyzed, which makes this the first time that a Rhodococcus species acted on sulfate or sulfite, thereby generating diols.
6.2. Sulfide Monooxygenase
Chiral sulfoxides are key synthons in chiral drug production and are used as versatile auxiliary compounds as chiral ligands or catalysts. The asymmetric oxidation of prochiral sulfides catalyzed by whole-cell biocatalysts is superior to classical chemical routes [233]. Phenyl methyl sulfide (PMS) was oxidized into (S)-phenyl methyl sulfoxide (PMSO) by whole-cells of R. sp. ECU0066 [234,235] and R. sp. CCZU10-1 [233]. Both studies showed the expansion of the substrate scope to a small set of PMS derivatives. A sulfide/sulfoxide flavin-dependent monooxygenase from R. sp. IGTS8 was produced in E. coli and investigated in detail [236]. It was shown that the enzyme oxidizes DBT in two consecutive steps, yielding DBT sulfone.
7. Conclusions
Enzymes present in Rhodococcus strains cover a broad field of diverse reactions ranging from redox-reactions and hydrolysis reactions of epoxides or esters to the hydration of fatty acids or Michael acceptors. The most prominent and best exploited enzymes in Rhodococcus are mostly the enzymes present in the aldoxime-nitrile pathway, with the nitrile hydratase being the prime example of a successful biocatalyst being run on a multi-ton industrial scale process. On the other hand, enzymes that have only recently been discovered like aldoxime dehydratase or enzymes acting on sulfur-containing compounds have the potential for further development as the industrial biocatalysts of the future.
Genome mining as well as the isolation of more Rhodococcus strains especially from rare sites opens up the chance to identify new enzymes with interesting properties such as an extraordinarily broad substrate scope, higher enantio- or stereoselectivities for defined reactions, better overall stabilities at extreme temperatures, or unusual reaction media. Additionally, the engineering of known enzymes to enhance their function also increases the number of possible chemical reactions. One such example is the evolution of amine dehydrogenase from amino acid dehydrogenase, which led to a straightforward synthesis of (R)-selective amines, which had been a major challenge in the past.
Few reactions have been carried out with purified enzymes while the majority of reactions shown were catalyzed by either Rhodococcus whole-cells (wild-type or mutants) or E. coli whole-cell reactions with heterologously expressed enzymes from Rhodococcus. It was shown that enzymes from Rhodococcus are valuable resources in the design of novel biocascades, which will attract only more attention in the near future. Here, the combination of enzymes in well-established expression hosts is theoretically boundless given that the present enzymes share reaction condition ranges (e.g., pH, temperature, solvent) and act on the same desired compounds.
In summary, the genus Rhodococcus truly deserves to be termed as a ‘biocatalytic powerhouse’: its enzymatic diversity and overall robustness make these microorganisms one of the key players in many areas of biocatalysis and will continue to do so.
Author Contributions
Conceptualization, P.L.H. and U.H.; Writing—original draft preparation, H.B.; Writing—review and editing, P.L.H. and U.H.
Funding
This work is part of the Open Technology Program with project number 14170, which is (partly) financed by the Netherlands Organization for Scientific Research (NWO).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Alvarez, H.M. Biology of Rhodococcus, 2nd ed.; Springer: Basel, Switzerland, 2019. [Google Scholar]
- Larkin, M.J.; Kulakov, L.A.; Allen, C.C.R. Biodegradation by Members of the Genus Rhodococcus: Biochemistry, Physiology, and Genetic Adaptation. Adv. Appl. Microbiol. 2006, 59, 1–29. [Google Scholar] [PubMed]
- Zampolli, J.; Zeaiter, Z.; Di Canito, A.; Di Gennaro, P. Genome analysis and -omics approaches provide new insights into the biodegradation potential of Rhodococcus. Appl. Microbiol. Biotechnol. 2019, 103, 1069–1080. [Google Scholar] [CrossRef] [PubMed]
- Abdelraheem, E.M.M.; Busch, H.; Hanefeld, U.; Tonin, F. Biocatalysis explained: From Pharmaceutical to Bulk Chemical Production. React. Chem. Eng. 2019. [Google Scholar] [CrossRef]
- Gröger, H.; Asano, Y.; Bornscheuer, U.T.; Ogawa, J. Development of biocatalytic processes in Japan and Germany: From research synergies to industrial applications. Chem. Asian J. 2012, 7, 1138–1153. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, J.; Shimizu, S. Industrial microbial enzymes: Their discovery by screening and use in large-scale production of useful chemicals in Japan. Curr. Opin. Biotechnol. 2002, 13, 367–375. [Google Scholar] [CrossRef]
- Kim, D.; Choi, K.Y.; Yoo, M.; Zylstra, G.J.; Kim, E. Biotechnological potential of Rhodococcus biodegradative pathways. J. Microbiol. Biotechnol. 2018, 28, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.J.; Kulakov, L.A.; Allen, C.C.R. Biodegradation and Rhodococcus—Masters of catabolic versatility. Curr. Opin. Biotechnol. 2005, 16, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Jiao, S.; Wang, M.; Yu, H.; Shen, Z. Overexpression of epoxide hydrolase in Rhodococcus ruber with high robustness for the synthesis of chiral epichlorohydrin. Process Biochem. 2019, 79, 49–56. [Google Scholar] [CrossRef]
- Van Der Geize, R.; Dijkhuizen, L. Harnessing the catabolic diversity of Rhodococci for environmental and biotechnological applications. Curr. Opin. Microbiol. 2004, 7, 255–261. [Google Scholar] [CrossRef]
- Kim, D.; Kim, Y.S.; Kim, S.K.; Kim, S.W.; Zylstra, G.J.; Kim, Y.M.; Kim, E. Monocyclic aromatic hydrocarbon degradation by Rhodococcus sp. strain DK17. Appl. Environ. Microbiol. 2002, 68, 3270–3278. [Google Scholar] [CrossRef]
- Kim, S.H.; Han, H.Y.; Lee, Y.J.; Kim, C.W.; Yang, J.W. Effect of electrokinetic remediation on indigenous microbial activity and community within diesel contaminated soil. Sci. Total Environ. 2010, 408, 3162–3168. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Roberts, J.N.; Hardiman, E.M.; Singh, R.; Eltis, L.D.; Bugg, T.D.H. Identification of DypB from Rhodococcus jostii RHA1 as a lignin peroxidase. Biochemistry 2011, 50, 5096–5107. [Google Scholar] [CrossRef] [PubMed]
- Mycroft, Z.; Gomis, M.; Mines, P.; Law, P.; Bugg, T.D.H. Biocatalytic conversion of lignin to aromatic dicarboxylic acids in Rhodococcus jostii RHA1 by re-routing aromatic degradation pathways. Green Chem. 2015, 17, 4974–4979. [Google Scholar] [CrossRef]
- Wei, Z.; Wilkinson, R.C.; Rashid, G.M.M.; Brown, D.; Fülöp, V.; Bugg, T.D.H. Characterization of Thiamine Diphosphate-Dependent 4-Hydroxybenzoylformate Decarboxylase Enzymes from Rhodococcus jostii RHA1 and Pseudomonas fluorescens Pf-5 Involved in Degradation of Aryl C-2 Lignin Degradation Fragments. Biochemistry 2019. [Google Scholar] [CrossRef] [PubMed]
- Masai, E.; Yamada, A.; Healy, J.M.; Hatta, T.; Kimbara, K.; Fukuda, M.; Yano, K. Characterization of biphenyl catabolic genes of gram-positive polychlorinated biphenyl degrader Rhodococcus sp. strain RHA1. Appl. Environ. Microbiol. 1995, 61, 2079–2085. [Google Scholar] [PubMed]
- Khairy, H.; Wübbeler, J.H.; Steinbüchel, A. Biodegradation of the Organic Disulfide 4,4′-Dithiodibutyric Acid by Rhodococcus spp. Appl. Environ. Microbiol. 2015, 81, 8294–8306. [Google Scholar] [CrossRef] [PubMed]
- Anteneh, Y.S.; Franco, C.M.M. Whole cell actinobacteria as biocatalysts. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Hollmann, F. Oxidation using Dehydrogenases. In Biocatalysis in Organic Synthesis 3; Faber, K., Fessner, W.-D., Turner, N.J., Eds.; Georg Thieme Verlag KG: Stuttgart, Germany, 2015; pp. 115–138. [Google Scholar]
- Schenkels, P.; Duine, J.A. Nicotinoprotein (NADH-containing) alcohol dehydrogenase from Rhodococcus erythropolis DSM 1069: An efficient catalyst for coenzyme-independent oxidation of a broad spectrum of alcohols and the interconversion of alcohols and aldehydes. Microbiology 2000, 146, 775–785. [Google Scholar] [CrossRef]
- Stampfer, W.; Kosjek, B.; Moitzi, C.; Kroutil, W.; Faber, K. Biocatalytic asymmetric hydrogen transfer. Angew. Chem. Int. Ed. 2002, 41, 1014–1017. [Google Scholar] [CrossRef]
- Nikodinovic, J.; Dinges, J.M.; Bergmeier, S.C.; McMills, M.C.; Wright, D.L.; Priestley, N.D. Resolution of methyl nonactate by Rhodococcus erythropolis under aerobic and anaerobic conditions. Org. Lett. 2006, 8, 443–445. [Google Scholar] [CrossRef]
- Voss, C.V.; Gruber, C.C.; Faber, K.; Knaus, T.; Macheroux, P.; Kroutil, W. Orchestration of concurrent oxidation and reduction cycles for stereoinversion and deracemisation of sec-alcohols. J. Am. Chem. Soc. 2008, 130, 13969–13972. [Google Scholar] [CrossRef] [PubMed]
- Voss, C.V.; Gruber, C.C.; Kroutil, W. Deracemization of secondary alcohols through a concurrent tandem biocatalytic oxidation and reduction. Angew. Chem. Int. Ed. 2008, 47, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Li, Z. Whole-Cell Cascade Biotransformations for One-Pot Multistep Organic Synthesis. ChemCatChem 2018, 10, 2164–2178. [Google Scholar] [CrossRef]
- Müller, C.A.; Weingartner, A.M.; Dennig, A.; Ruff, A.J.; Gröger, H.; Schwaneberg, U. A whole cell biocatalyst for double oxidation of cyclooctane. J. Ind. Microbiol. Biotechnol. 2016, 43, 1641–1646. [Google Scholar] [CrossRef] [PubMed]
- Both, P.; Busch, H.; Kelly, P.P.; Mutti, F.G.; Turner, N.J.; Flitsch, S.L. Whole-Cell Biocatalysts for Stereoselective C-H Amination Reactions. Angew. Chem. Int. Ed. 2016, 55, 1511–1513. [Google Scholar] [CrossRef] [PubMed]
- Biermann, M.; Gruß, H.; Hummel, W.; Gröger, H. Guerbet Alcohols: From Processes under Harsh Conditions to Synthesis at Room Temperature under Ambient Pressure. ChemCatChem 2016, 8, 895–899. [Google Scholar] [CrossRef]
- Stankevičiūtė, J.; Kutanovas, S.; Rutkienė, R.; Tauraitė, D.; Striela, R.; Meškys, R. Ketoreductase TpdE from Rhodococcus jostii TMP1: Characterization and application in the synthesis of chiral alcohols. PeerJ 2015, 3, e1387. [Google Scholar] [CrossRef]
- Gröger, H.; Hummel, W.; Rollmann, C.; Chamouleau, F.; Hüsken, H.; Werner, H.; Wunderlich, C.; Abokitse, K.; Drauz, K.; Buchholz, S. Preparative asymmetric reduction of ketones in a biphasic medium with an (S)-alcohol dehydrogenase under in situ-cofactor-recycling with a formate dehydrogenase. Tetrahedron 2004, 60, 633–640. [Google Scholar] [CrossRef]
- Pollard, D.; Truppo, M.; Pollard, J.; Chen, C.-Y.; Moore, J. Effective synthesis of (S)-3,5-bistrifluoromethylphenyl ethanol by asymmetric enzymatic reduction. Tetrahedron Asymmetry 2006, 17, 554–559. [Google Scholar] [CrossRef]
- Liu, Y.C.; Wu, Z.L. Switchable asymmetric bio-epoxidation of α,β-unsaturated ketones. Chem. Commun. 2016, 52, 1158–1161. [Google Scholar] [CrossRef]
- Grau, B.T.; Devine, P.N.; DiMichele, L.N.; Kosjek, B. Chemo- and enantioselective routes to chiral fluorinated hydroxyketones using ketoreductases. Org. Lett. 2007, 9, 4951–4954. [Google Scholar] [CrossRef]
- Borzecka, W.; Lavandera, I.; Gotor, V. Synthesis of enantiopure fluorohydrins using alcohol dehydrogenases at high substrate concentrations. J. Org. Chem. 2013, 78, 7312–7317. [Google Scholar] [CrossRef]
- Busto, E.; Gotor-Fernández, V.; Gotor, V. Asymmetric chemoenzymatic synthesis of ramatroban using lipases and oxidoreductases. J. Org. Chem. 2012, 77, 4842–4848. [Google Scholar] [CrossRef]
- Busto, E.; Martínez-Montero, L.; Gotor, V.; Gotor-Fernández, V. Chemoenzymatic asymmetric synthesis of serotonin receptor agonist (R)-frovatriptan. Eur. J. Org. Chem. 2013, 2013, 4057–4064. [Google Scholar] [CrossRef]
- Méndez-Sánchez, D.; Mourelle-Insua, Á.; Gotor-Fernández, V.; Lavandera, I. Synthesis of α-Alkyl-β-Hydroxy Amides through Biocatalytic Dynamic Kinetic Resolution Employing Alcohol Dehydrogenases. Adv. Synth. Catal. 2019, 361, 2706–2712. [Google Scholar] [CrossRef]
- Mangas-Sánchez, J.; Busto, E.; Gotor, V.; Gotor-Fernández, V. One-pot synthesis of enantiopure 3,4-dihydroisocoumarins through dynamic reductive kinetic resolution processes. Org. Lett. 2013, 15, 3872–3875. [Google Scholar] [CrossRef]
- Ying, X.; Zhang, J.; Wang, C.; Huang, M.; Ji, Y.; Cheng, F.; Yu, M.; Wang, Z.; Ying, M. Characterization of a carbonyl reductase from Rhodococcus erythropolis WZ010 and its variant Y54F for asymmetric synthesis of (S)-N-Boc-3-hydroxypiperidine. Molecules 2018, 23, 3117. [Google Scholar] [CrossRef]
- Ewing, T.A.; Fraaije, M.W.; van Berkel, W.J.H. Oxidation using Alcohol oxidases. In Biocatalysis in Organic Synthesis 3; Faber, K., Fessner, W.-D., Turner, N.J., Eds.; Georg Thieme Verlag KG: Stuttgart, Germany, 2015; pp. 157–186. [Google Scholar]
- Jin, J.; Mazon, H.; Van Den Heuvel, R.H.H.; Janssen, D.B.; Fraaije, M.W. Discovery of a eugenol oxidase from Rhodococcus sp. strain RHA1. FEBS J. 2007, 274, 2311–2321. [Google Scholar] [CrossRef]
- Dieth, S.; Tritsch, D.; Biellmann, J.-F. Resolution of allylic alcohols by cholesterol oxidase isolated from Rhodococcus erythropolis. Tetrahedron Lett. 1995, 36, 2243–2246. [Google Scholar] [CrossRef]
- Wu, K.; Li, W.; Song, J.; Li, T. Production, Purification, and Identification of Cholest-4-en-3-one Produced by Cholesterol Oxidase from Rhodococcus sp. in Aqueous/Organic Biphasic System. Biochem. Insights 2015, 8, S1–S8. [Google Scholar] [CrossRef]
- Bertola, M.A.; Koger, H.S.; Phillips, G.T.; Marx, A.F.; Claassen, V.P. A process for the preparation of (R)- and (S)-2,2-R1,R2-1,3-dioxolane-4-methanol. E.P. 0244912 A1, 5 May 1987. [Google Scholar]
- Liese, A.; Seelbach, K.; Buchholz, A.; Haberland, J. Processes. In Industrial Biotransformations; Liese, A., Seelbach, K., Wandrey, C., Eds.; Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 2006; pp. 147–513. [Google Scholar]
- Nolte, J.C.; Urlacher, V.B. Cytochrome P450 in the oxidation of alkenes. In Biocatalysis in Organic Synthesis 3; Faber, K., Fessner, W.-D., Turner, N.J., Eds.; Georg Thieme Verlag KG: Stuttgart, Germany, 2015; pp. 21–63. [Google Scholar]
- Roper, L.; Grogan, G. Biocatalysis for Organic Chemists: Hydroxylations. In Organic Synthesis Using Biocatalysis; Goswami, A., Stewart, J.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 213–241. [Google Scholar]
- Roberts, G.A.; Celik, A.; Hunter, D.J.B.; Ost, T.W.B.; White, J.H.; Chapman, S.K.; Turner, N.J.; Flitsch, S.L. A self-sufficient cytochrome p450 with a primary structural organization that includes a flavin domain and a [2Fe-2S] redox center. J. Biol. Chem. 2003, 278, 48914–48920. [Google Scholar] [CrossRef]
- Roberts, G.A.; Grogan, G.; Greter, A.; Flitsch, S.L.; Turner, N.J. Identification of a new class of cytochrome P450 from a Rhodococcus sp. J. Bacteriol. 2002, 184, 3898–3908. [Google Scholar] [CrossRef]
- O’Reilly, E.; Corbett, M.; Hussain, S.; Kelly, P.P.; Richardson, D.; Flitsch, S.L.; Turner, N.J. Substrate promiscuity of cytochrome P450 RhF. Catal. Sci. Technol. 2013, 3, 1490–1492. [Google Scholar] [CrossRef]
- Sabbadin, F.; Grogan, G.; Bruce, N.C. LICRED: A versatile drop-in vector for rapid generation of redox-self-sufficient cytochromes P450. Methods Mol. Biol. 2010, 11, 987–994. [Google Scholar] [CrossRef]
- De Gonzalo, G.; van Berkel, W.J.H.; Fraaije, M.W. Baeyer-Villiger Oxidation. In Biocatalysis in Organic Synthesis 3; Faber, K., Fessner, W.-D., Turner, N.J., Eds.; Georg Thieme Verlag KG: Stuttgart, Germany, 2015; pp. 187–234. [Google Scholar]
- Brzostowicz, P.C.; Walters, D.M.; Thomas, S.M.; Nagarajan, V.; Rouvière, P.E. mRNA differential display in a microbial enrichment culture: Simultaneous identification of three cyclohexanone monooxygenases from three species. Appl. Environ. Microbiol. 2003, 69, 334–342. [Google Scholar] [CrossRef]
- Rudroff, F.; Rydz, J.; Ogink, F.H.; Fink, M.; Mihovilovic, M.D. Comparing the stereoselective biooxidation of cyclobutanones by recombinant strains expressing bacterial baeyer—Villiger monooxygenases. Adv. Synth. Catal. 2007, 349, 1436–1444. [Google Scholar] [CrossRef]
- Mihovilovic, M.D.; Snajdrova, R.; Grötzl, B. Microbial Baeyer-Villiger oxidation of 4,4-disubstituted cyclohexan- and cyclohexenones by recombinant whole-cells expressing monooxygenases of bacterial origin. J. Mol. Catal. B Enzym. 2006, 39, 135–140. [Google Scholar] [CrossRef]
- Snajdrova, R.; Braun, I.; Bach, T.; Mereiter, K.; Mihovilovic, M.D. Biooxidation of bridged cycloketones using Baeyer-Villiger monooxygenases of various bacterial origin. J. Org. Chem. 2007, 72, 9597–9603. [Google Scholar] [CrossRef]
- Rehdorf, J.; Mihovilovic, M.D.; Fraaije, M.W.; Bornscheuer, U.T. Enzymatic synthesis of enantiomerically pure β-amino ketones, β-amino esters, and β-amino alcohols with Baeyer-Villiger monooxygenases. Chem. A Eur. J. 2010, 16, 9525–9535. [Google Scholar] [CrossRef]
- Kostichka, K.; Thomas, S.M.; Gibson, K.J.; Nagarajan, V.; Cheng, Q. Cloning and characterization of a gene cluster for cyclododecanone oxidation in Rhodococcus ruber SC1. J. Bacteriol. 2001, 183, 6478–6486. [Google Scholar] [CrossRef]
- Fink, M.J.; Rudroff, F.; Mihovilovic, M.D. Baeyer-Villiger monooxygenases in aroma compound synthesis. Bioorg. Med. Chem. Lett. 2011, 21, 6135–6138. [Google Scholar] [CrossRef]
- Montersino, S.; van Berkel, W.J.H. Functional annotation and characterization of 3-hydroxybenzoate 6-hydroxylase from Rhodococcus jostii RHA1. Biochim. Biophys. Acta Proteins Proteom. 2012, 1824, 433–442. [Google Scholar] [CrossRef]
- McLoed, M.P.; Warren, R.L.; Hsiao, W.W.L.; Araki, N.; Myhre, M.; Fernandes, C.; Miyazawa, D.; Wong, W.; Lillquist, A.L.; Wang, D.; et al. The complete genome of Rhodococcus sp. RHA1 provides insights into a catabolic powerhouse. PNAS 2006, 103, 15582–15587. [Google Scholar] [CrossRef]
- Riebel, A.; Dudek, H.M.; De Gonzalo, G.; Stepniak, P.; Rychlewski, L.; Fraaije, M.W. Expanding the set of rhodococcal Baeyer-Villiger monooxygenases by high-throughput cloning, expression and substrate screening. Appl. Microbiol. Biotechnol. 2012, 95, 1479–1489. [Google Scholar] [CrossRef]
- Riebel, A.; De Gonzalo, G.; Fraaije, M.W. Expanding the biocatalytic toolbox of flavoprotein monooxygenases from Rhodococcus jostii RHA1. J. Mol. Catal. B Enzym. 2013, 88, 20–25. [Google Scholar] [CrossRef]
- Messiha, H.L.; Ahmed, S.T.; Karuppiah, V.; Suardíaz, R.; Ascue Avalos, G.A.; Fey, N.; Yeates, S.; Toogood, H.S.; Mulholland, A.J.; Scrutton, N.S. Biocatalytic Routes to Lactone Monomers for Polymer Production. Biochemistry 2018, 57, 1997–2008. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Li, C.-X.; Xu, J.-H.; Zheng, G.-W. Efficient Synthesis of Methyl 3-Acetoxypropionate by a newly identified Baeyer-Villiger Monooxygenase. Appl. Environ. Microbiol. 2019, 85, 1–10. [Google Scholar] [CrossRef]
- Heine, T.; van Berkel, W.J.H.; Gassner, G.; van Pée, K.H.; Tischler, D. Two-component FAD-dependent monooxygenases: Current knowledge and biotechnological opportunities. Biology 2018, 7, 42. [Google Scholar] [CrossRef]
- Paul, C.E.; Tischler, D.; Riedel, A.; Heine, T.; Itoh, N.; Hollmann, F. Nonenzymatic Regeneration of Styrene Monooxygenase for Catalysis. ACS Catal. 2015, 5, 2961–2965. [Google Scholar] [CrossRef]
- Riedel, A.; Heine, T.; Westphal, A.H.; Conrad, C.; Rathsack, P.; van Berkel, W.J.H.; Tischler, D. Catalytic and hydrodynamic properties of styrene monooxygenases from Rhodococcus opacus 1CP are modulated by cofactor binding. AMB Express 2015, 5, 1–11. [Google Scholar] [CrossRef]
- Toda, H.; Imae, R.; Komio, T.; Itoh, N. Expression and characterization of styrene monooxygenases of Rhodococcus sp. ST-5 and ST-10 for synthesizing enantiopure (S)-epoxides. Appl. Microbiol. Biotechnol. 2012, 96, 407–418. [Google Scholar] [CrossRef]
- Toda, H.; Imae, R.; Itoh, N. Efficient biocatalysis for the production of enantiopure (S)-epoxides using a styrene monooxygenase (SMO) and Leifsonia alcohol dehydrogenase (LSADH) system. Tetrahedron Asymmetry 2012, 23, 1542–1549. [Google Scholar] [CrossRef]
- Toda, H.; Imae, R.; Itoh, N. Bioproduction of chiral epoxyalkanes using styrene monooxygenase from Rhodococcus sp. ST-10 (RhSMO). Adv. Synth. Catal. 2014, 356, 3443–3450. [Google Scholar] [CrossRef]
- Toda, H.; Ohuchi, T.; Imae, R.; Itoh, N. Microbial production of aliphatic (S)-epoxyalkanes by using Rhodococcus sp. strain ST-10 styrene monooxygenase expressed in organic-solvent-tolerant Kocuria rhizophila DC2201. Appl. Environ. Microbiol. 2015, 81, 1919–1925. [Google Scholar] [CrossRef]
- Tischler, D.; Eulberg, D.; Lakner, S.; Kaschabek, S.R.; Van Berkel, W.J.H.; Schlömann, M. Identification of a novel self-sufficient styrene monooxygenase from Rhodococcus opacus 1CP. J. Bacteriol. 2009, 191, 4996–5009. [Google Scholar] [CrossRef]
- Tischler, D.; Kermer, R.; Gröning, J.A.D.; Kaschabek, S.R.; Van Berkel, W.J.H.; Schlömann, M. StyA1 and StyA2B from Rhodococcus opacus 1CP: A multifunctional styrene monooxygenase system. J. Bacteriol. 2010, 192, 5220–5227. [Google Scholar] [CrossRef]
- Heine, T.; Grossmann, C.; Hofmann, S.; Tischler, D. Indigoid dyes by group E monooxygenases: Mechanism and biocatalysis. Biol. Chem. 2019, 400, 939–950. [Google Scholar] [CrossRef]
- Gakhar, L.; Malik, Z.A.; Allen, C.C.R.; Lipscomb, D.A.; Larkin, M.J.; Ramaswamy, S. Structure and increased thermostability of Rhodococcus sp. naphthalene 1,2-dioxygenase. J. Bacteriol. 2005, 187, 7222–7231. [Google Scholar] [CrossRef]
- Kim, D.; Yoo, M.; Choi, K.Y.; Kang, B.S.; Kim, E. Characterization and engineering of an o-xylene dioxygenase for biocatalytic applications. Bioresour. Technol. 2013, 145, 123–127. [Google Scholar] [CrossRef]
- Kim, D.; Choi, K.Y.; Yoo, M.; Choi, J.N.; Lee, C.H.; Zylstra, G.J.; Kang, B.S.; Kim, E. Benzylic and aryl hydroxylations of m-xylene by o-xylene dioxygenase from Rhodococcus sp. strain DK17. Appl. Microbiol. Biotechnol. 2010, 86, 1841–1847. [Google Scholar] [CrossRef]
- Kim, D.; Kim, Y.S.; Jung, J.W.; Zylstra, G.J.; Kim, Y.M.; Kim, S.K.; Kim, E. Regioselective oxidation of xylene isomers by Rhodococcus sp. strain DK17. FEMS Microbiol. Lett. 2003, 223, 211–214. [Google Scholar] [CrossRef]
- Kim, D.; Lee, J.S.; Choi, K.Y.; Kim, Y.S.; Choi, J.N.; Kim, S.K.; Chae, J.C.; Zylstra, G.J.; Lee, C.H.; Kim, E. Effect of functional groups on the regioselectivity of a novel o-xylene dioxygenase from Rhodococcus sp. strain DK17. Enzym. Microb. Technol. 2007, 41, 221–225. [Google Scholar] [CrossRef]
- Kim, D.; Yoo, M.; Choi, K.Y.; Kang, B.S.; Kim, T.K.; Hong, S.G.; Zylstra, G.J.; Kim, E. Differential degradation of bicyclics with aromatic and alicyclic rings by Rhodococcus sp. strain DK17. Appl. Environ. Microbiol. 2011, 77, 8280–8287. [Google Scholar] [CrossRef]
- Kim, D.; Lee, C.H.; Choi, J.N.; Choi, K.Y.; Zylstra, G.J.; Kim, E. Aromatic hydroxylation of indan by o-xylene-degrading Rhodococcus sp. strain DK17. Appl. Environ. Microbiol. 2010, 76, 375–377. [Google Scholar] [CrossRef]
- Chartrain, M.; Jackey, B.; Taylor, C.; Sandford, V.; Gbewonyo, K.; Lister, L.; Dimichele, L.; Hirsch, C.; Heimbuch, B.; Maxwell, C.; et al. Bioconversion of indene to cis (1S,2R) indandiol and trans (1R,2R) indandiol by Rhodococcus species. J. Ferment. Bioeng. 1998, 86, 550–558. [Google Scholar] [CrossRef]
- Stafford, D.E.; Yanagimachi, K.S.; Lessard, P.A.; Rijhwani, S.K.; Sinskey, A.J.; Stephanopoulos, G. Optimizing bioconversion pathways through systems analysis and metabolic engineering. Proc. Natl. Acad. Sci. USA 2002, 99, 1801–1806. [Google Scholar] [CrossRef] [Green Version]
- Amanullah, A.; Hewitt, C.J.; Nienow, A.W.; Lee, C.; Chartrain, M.; Buckland, B.C.; Drew, S.W.; Woodley, J.M. Fed-batch bioconversion of indene to cis-indandiol. Enzym. Microb. Technol. 2002, 31, 954–967. [Google Scholar] [CrossRef]
- Priefert, H.; O’Brien, X.M.; Lessard, P.A.; Dexter, A.F.; Choi, E.E.; Tomic, S.; Nagpal, G.; Cho, J.J.; Agosto, M.; Yang, L.; et al. Indene bioconversion by a toluene inducible dioxygenase of Rhodococcus sp. I24. Appl. Microbiol. Biotechnol. 2004, 65, 168–176. [Google Scholar] [CrossRef]
- Solyanikova, I.P.; Emelyanova, E.V.; Borzova, O.V.; Golovleva, L.A. Benzoate degradation by Rhodococcus opacus 1CP after dormancy: Characterization of dioxygenases involved in the process. J. Environ. Sci. Health Part B 2016, 51, 182–191. [Google Scholar] [CrossRef]
- Yang, X.; Xie, F.; Zhang, G.; Shi, Y.; Qian, S. Purification, characterization, and substrate specificity of two 2,3-dihydroxybiphenyl 1,2-dioxygenase from Rhodococcus sp. R04, showing their distinct stability at various temperature. Biochimie 2008, 90, 1530–1538. [Google Scholar] [CrossRef]
- Xiong, F.; Shuai, J.J.; Peng, R.H.; Tian, Y.S.; Zhao, W.; Yao, Q.H.; Xiong, A.S. Expression, purification and functional characterization of a recombinant 2,3-dihydroxybiphenyl-1,2-dioxygenase from Rhodococcus rhodochrous. Mol. Biol. Rep. 2011, 38, 4303–4308. [Google Scholar] [CrossRef]
- Cha, C.J. Metabolizing Alkyl-Substituted Catechols N75 Capable of metabolising alkyl-substituted catechols. J. Microbiol. Biotechnol. 2006, 16, 778–785. [Google Scholar]
- Matsumura, E.; Ooi, S.; Murakami, S.; Takenaka, S.; Aoki, K. Constitutive synthesis, purification, and characterization of catechol 1,2-dioxygenase from the aniline-assimilating bacterium Rhodococcus sp. AN-22. J. Biosci. Bioeng. 2004, 98, 71–76. [Google Scholar] [CrossRef]
- Deutz, W.A.; Fjaellman, A.H.M.; Ren, S.; Jourdat, C.; Witholt, B. Biotransformation of d-limonene to (+) trans carveol by toluene-grown Rhodococcus opacus PWD4 cells. Appl. Environ. Microbiol. 2001, 67, 2829–2832. [Google Scholar] [CrossRef]
- Thompson, M.L.; Marriott, R.; Dowle, A.; Grogan, G. Biotransformation of β-myrcene to geraniol by a strain of Rhodococcus erythropolis isolated by selective enrichment from hop plants. Appl. Microbiol. Biotechnol. 2010, 85, 721–730. [Google Scholar] [CrossRef]
- Van Der Geize, R.; Hessels, G.I.; Van Gerwen, R.; Van Der Meijden, P.; Dijkhuizen, L. Molecular and functional characterization of kshA and kshB, encoding two components of 3-ketosteroid 9α-hydroxylase, a class IA monooxygenase, in Rhodococcus erythropolis strain SQ1. Mol. Microbiol. 2002, 45, 1007–1018. [Google Scholar] [CrossRef]
- Li, K.; Wang, J.; Wu, K.; Zheng, D.; Zhou, X.; Han, W.; Wan, N.; Cui, B.; Chen, Y. Enantioselective synthesis of 1,2,3,4-tetrahydroquinoline-4-ols and 2,3-dihydroquinolin-4(1:H)-ones via a sequential asymmetric hydroxylation/diastereoselective oxidation process using Rhodococcus equi ZMU-LK19. Org. Biomol. Chem. 2017, 15, 3580–3584. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Brady, D. Broadening the Scope of Biocatalysis in Sustainable Organic Synthesis. ChemSusChem 2019, 12, 2859–2881. [Google Scholar] [CrossRef]
- Stompor, M.; Kałużny, M.; Żarowska, B. Biotechnological methods for chalcone reduction using whole cells of Lactobacillus, Rhodococcus and Rhodotorula strains as a way to produce new derivatives. Appl. Microbiol. Biotechnol. 2016, 100, 8371–8384. [Google Scholar] [CrossRef]
- Chen, B.-S.; Medici, R.; van der Helm, M.P.; van Zwet, Y.; Gjonaj, L.; van der Geest, R.; Otten, L.G.; Hanefeld, U. Rhodococcus strains as source for ene-reductase activity. Appl. Microbiol. Biotechnol. 2018, 102, 5545–5556. [Google Scholar] [CrossRef]
- Riedel, A.; Mehnert, M.; Paul, C.E.; Westphal, A.H.; van Berkel, W.J.H.; Tischler, D. Functional characterization and stability improvement of a “thermophilic-like” ene-reductase from Rhodococcus opacus 1CP. Front. Microbiol. 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Hummel, W.; Schütte, H.; Schmidt, E.; Wandrey, C.; Kula, M.R. Isolation of l-phenylalanine dehydrogenase from Rhodococcus sp. M4 and its application for the production of l-phenylalanine. Appl. Microbiol. Biotechnol. 1987, 26, 409–416. [Google Scholar] [CrossRef]
- Bradshaw, C.W.; Wong, C.H.; Hummel, W.; Kula, M.R. Enzyme-catalyzed asymmetric synthesis of (S)-2-amino-4-phenylbutanoic acid and (R)-2-hydroxy-4-phenylbutanoic acid. Bioorg. Chem. 1991, 19, 29–39. [Google Scholar] [CrossRef]
- Sharma, M.; Mangas-Sanchez, J.; Turner, N.J.; Grogan, G. NAD(P)H-Dependent Dehydrogenases for the Asymmetric Reductive Amination of Ketones: Structure, Mechanism, Evolution and Application. Adv. Synth. Catal. 2017, 359, 2011–2025. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.J.; Toh, H.H.; Yang, Y.; Adams, J.P.; Snajdrova, R.; Li, Z. Engineering of amine dehydrogenase for asymmetric reductive amination of ketone by evolving Rhodococcus phenylalanine dehydrogenase. ACS Catal. 2015, 5, 1119–1122. [Google Scholar] [CrossRef]
- Liu, J.; Pang, B.Q.W.; Adams, J.P.; Snajdrova, R.; Li, Z. Coupled Immobilized Amine Dehydrogenase and Glucose Dehydrogenase for Asymmetric Synthesis of Amines by Reductive Amination with Cofactor Recycling. ChemCatChem 2017, 9, 425–431. [Google Scholar] [CrossRef]
- Knaus, T.; Böhmer, W.; Mutti, F.G. Amine dehydrogenases: Efficient biocatalysts for the reductive amination of carbonyl compounds. Green Chem. 2017, 19, 453–463. [Google Scholar] [CrossRef]
- Takeuchi, K.; Koike, K.; Ito, S. Production of cis-unsaturated hydrocarbons by a strain of Rhodococcus in repeated batch culture with a phase-inversion, hollow-fiber system. J. Biotechnol. 1990, 14, 179–186. [Google Scholar] [CrossRef]
- Araki, H.; Hagihara, H.; Takigawa, H.; Tsujino, Y.; Ozaki, K. Novel Genes Encoding Hexadecanoic Acid Δ6-Desaturase Activity in a Rhodococcus sp. Curr. Microbiol. 2016, 73, 646–651. [Google Scholar] [CrossRef]
- Araki, H.; Hagiwara, H.; Kotani, S.; Yukiharu, T.; Takikawa, H. Rhodococcus Fatty Acid Δ6-Desaturase Genes Useful for Production of cis-6-Hexadecenoic Acid. J.P. 2,004,283,168, 14 October 2004. [Google Scholar]
- Bhalla, T.C.; Kumar, V.; Kumar, V. Enzymes of aldoxime-nitrile pathway for organic synthesis. Rev. Environ. Sci. Biotechnol. 2018, 17, 229–239. [Google Scholar] [CrossRef]
- Bhalla, T.C.; Kumar, V.; Kumar, V.; Thakur, N. Savitri Nitrile Metabolizing Enzymes in Biocatalysis and Biotransformation. Appl. Biochem. Biotechnol. 2018, 185, 925–946. [Google Scholar] [CrossRef]
- Kato, Y.; Ryoko, O.; Asano, Y. Distribution of aldoxime dehydratase in microorganisms. Appl. Environ. Microbiol. 2000, 66, 2290–2296. [Google Scholar] [CrossRef]
- Betke, T.; Higuchi, J.; Rommelmann, P.; Oike, K.; Nomura, T.; Kato, Y.; Asano, Y.; Gröger, H. Biocatalytic Synthesis of Nitriles through Dehydration of Aldoximes: The Substrate Scope of Aldoxime Dehydratases. ChemBioChem 2018, 19, 768–779. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Yamamoto, K.; Asano, Y. Identification and characterization of CYP79D16 and CYP71AN24 catalyzing the first and second steps in l-phenylalanine-derived cyanogenic glycoside biosynthesis in the Japanese apricot, Prunus mume Sieb. et Zucc. Plant Mol. Biol. 2014, 86, 215–223. [Google Scholar] [CrossRef]
- Gong, J.S.; Shi, J.S.; Lu, Z.M.; Li, H.; Zhou, Z.M.; Xu, Z.H. Nitrile-converting enzymes as a tool to improve biocatalysis in organic synthesis: Recent insights and promises. Crit. Rev. Biotechnol. 2017, 37, 69–81. [Google Scholar] [CrossRef]
- Asano, Y.; Kato, Y. Z-phenylacetaldoxime degradation by a novel aldoxime dehydratase from Bacillus sp. strain OxB-1. FEMS Microbiol. Lett. 1998, 158, 185–190. [Google Scholar] [CrossRef]
- Kato, Y.; Tsuda, T.; Asano, Y. Nitrile hydratase involved in aldoxime metabolism from Rhodococcus sp. strain YH3-3. J. Mol. Catal. B Enzym. 1999, 6, 249–256. [Google Scholar] [CrossRef]
- Metzner, R.; Okazaki, S.; Asano, Y.; Gröger, H. Cyanide-free enantioselective synthesis of nitriles: Synthetic proof of a biocatalytic concept and mechanistic insights. ChemCatChem 2014, 6, 3105–3109. [Google Scholar] [CrossRef]
- Kato, Y.; Yoshida, S.; Xie, S.-X.; Asano, Y. Aldoxime Dehydratase Co-existing with Nitrile Hydratase and Amidase in the Iron-Type Nitrile Hydratase-Producer Rhodococcus sp. N-771. J. Biosci. Bioeng. 2004, 97, 250–259. [Google Scholar] [CrossRef]
- Xie, S.X.; Kato, Y.; Komeda, H.; Yoshida, S.; Asano, Y. A Gene Cluster Responsible for Alkylaldoxime Metabolism Coexisting with Nitrile Hydratase and Amidase in Rhodococcus globerulus A-4. Biochemistry 2003, 42, 12056–12066. [Google Scholar] [CrossRef]
- Betke, T.; Rommelmann, P.; Oike, K.; Asano, Y.; Gröger, H. Cyanide-Free and Broadly Applicable Enantioselective Synthetic Platform for Chiral Nitriles through a Biocatalytic Approach. Angew. Chem. Int. Ed. 2017, 56, 12361–12366. [Google Scholar] [CrossRef]
- Kato, Y.; Nakamura, K.; Sakiyama, H.; Mayhew, S.G.; Asano, Y. Novel heme-containing lyase, phenylacetaldoxime dehydratase from Bacillus sp. strain OxB-1: Purification, characterization, and molecular cloning of the gene. Biochemistry 2000, 39, 800–809. [Google Scholar] [CrossRef]
- BASF, S.E. Method for Biocatalytic Production of Nitriles from Oximes and Oxime Dehydratases Usable Therein. U.S. 9,080,191, 14 July 2015. [Google Scholar]
- Asano, Y. Nitrile and amide hydrolysis and enzymatic synthesis of amides and peptides. In Biocatalysis in Organic Synthesis 1; Faber, K., Fessner, W.-D., Turner, N.J., Eds.; Georg Thieme Verlag KG: Stuttgart, Germany, 2015; pp. 255–276. [Google Scholar]
- Prasad, S.; Bhalla, T.C. Nitrile hydratases (NHases): At the interface of academia and industry. Biotechnol. Adv. 2010, 28, 725–741. [Google Scholar] [CrossRef]
- Nagasawa, T.; Shimizu, H.; Yamada, H. The superiority of the third-generation catalyst, Rhodococcus rhodochrous J1 nitrile hydratase, for industrial production of acrylamide. Appl. Microb. Biotechnol. 1993, 40, 189–195. [Google Scholar] [CrossRef]
- Nagasawa, T.; Takeuchi, K.; Nardi-Dei, V.; Mihara, Y.; Yamada, H. Optimum culture conditions for the production of cobalt-containing nitrile hydratase by Rhodococcus rhodochrous J1. Appl. Microbiol. Biotechnol. 1991, 34, 783–788. [Google Scholar] [CrossRef]
- Li, B.; Su, J.; Tao, J. Enzyme and process development for production of nicotinamide. Org. Process Res. Dev. 2011, 15, 291–293. [Google Scholar] [CrossRef]
- Nagasawa, T.; Mathew, C.D.; Mauger, J.; Yamada, H. Nitrile Hydratase-Catalyzed Production of Nicotinamide from 3-Cyanopyridine in Rhodococcus rhodochrous J1. Appl. Environ. Microbiol. 1988, 54, 1766–1779. [Google Scholar]
- Mauger, J.; Nagasawa, T.; Yamada, H. Nitrile hydratase-catalyzed production of isonicotinamide, picolinamide and pyrazinamide from 4-cyanopyridine, 2-cyanopyridine and cyanopyrazine in Rhodococcus rhodochrous J1. J. Biotechnol. 1988, 8, 87–95. [Google Scholar] [CrossRef]
- Meth-Cohn, O.; Wang, M.X. An in-depth study of the biotransformation of nitriles into amides and/or acids using Rhodococcus rhodochrous AJ270. J. Chem. Soc. Perkin Trans. 1 1997, 1099–1104. [Google Scholar] [CrossRef]
- Kashiwagi, M.; Fuhshuku, K.I.; Sugai, T. Control of the nitrile-hydrolyzing enzyme activity in Rhodococcus rhodochrous IFO 15564: Preferential action of nitrile hydratase and amidase depending on the reaction condition factors and its application to the one-pot preparation of amides from aldehyd. J. Mol. Catal. B Enzym. 2004, 29, 249–258. [Google Scholar] [CrossRef]
- Holtze, M.S.; Sørensen, J.; Hansen, H.C.B.; Aamand, J. Transformation of the herbicide 2,6-dichlorobenzonitrile to the persistent metabolite 2,6-dichlorobenzamide (BAM) by soil bacteria known to harbour nitrile hydratase or nitrilase. Biodegradation 2006, 17, 503–510. [Google Scholar] [CrossRef]
- Tang, R.; Shen, Y.; Wang, M.; Zhai, Y.; Gao, Q. Highly chemoselective and efficient production of 2,6-difluorobenzamide using Rhodococcus ruber CGMCC3090 resting cells. J. Biosci. Bioeng. 2017, 124, 641–646. [Google Scholar] [CrossRef]
- Wang, M.X.; Lin, S.J.; Liu, C.S.; Zheng, Q.Y.; Li, J.S. Nitrile biotransformations for highly efficient and enantioselective syntheses of electrophilic oxiranecarboxamides. J. Org. Chem. 2003, 68, 4570–4573. [Google Scholar] [CrossRef]
- Pawar, S.V.; Yadav, G.D. Enantioselective enzymatic hydrolysis of rac-mandelonitrile to (R)-mandelamide by nitrile hydratase immobilized on poly(vinyl alcohol)/chitosan-glutaraldehyde support. Ind. Eng. Chem. Res. 2014, 53, 7986–7991. [Google Scholar] [CrossRef]
- Wang, Y.J.; Xue, Y.P.; Liu, Z.Q.; Zheng, Y.G.; Zheng, R.C. Screening, cultivation, and biocatalytic performance of Rhodococcus boritolerans FW815 with strong 2,2-dimethylcyclopropanecarbonitrile hydratase activity. J. Ind. Microbiol. Biotechnol. 2012, 39, 409–417. [Google Scholar] [CrossRef]
- Crosby, J.; Moilliet, J.; Parratt, J.S.; Turner, N.J. Regioselective hydrolysis of aromatic dinitriles using a whole cell catalyst. J. Chem. Soc. Perkin Trans. 1 1994, 3, 1679–1687. [Google Scholar] [CrossRef]
- Matoishi, K.; Sano, A.; Imai, N.; Yamazaki, T.; Yokoyama, M.; Sugai, T.; Ohta, H. Rhodococcus rhodochrous IFO 15564-mediated hydrolysis of alicyclic nitriles and amides: Stereoselectivity and use for kinetic resolution and asymmetrization. Tetrahedron Asymmetry 1998, 9, 1097–1102. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, M.; Li, X.; Zhang, J.; Sun, H.; Luo, J. Highly efficient synthesis of 5-cyanovaleramide by Rhodococcus ruber CGMCC3090 resting cells. J. Chem. Technol. Biotechnol. 2012, 87, 1396–1400. [Google Scholar] [CrossRef]
- Yasukawa, K.; Hasemi, R.; Asano, Y. Dynamic kinetic resolution of α-aminonitriles to form chiral α-amino acids. Adv. Synth. Catal. 2011, 353, 2328–2332. [Google Scholar] [CrossRef]
- Kubáč, D.; Kaplan, O.; Elišáková, V.; Pátek, M.; Vejvoda, V.; Slámová, K.; Tóthová, A.; Lemaire, M.; Gallienne, E.; Lutz-Wahl, S.; et al. Biotransformation of nitriles to amides using soluble and immobilized nitrile hydratase from Rhodococcus erythropolis A4. J. Mol. Catal. B Enzym. 2008, 50, 107–113. [Google Scholar] [CrossRef]
- Vervisch, K.; D’Hooghe, M.; Rutjes, F.P.J.T.; De Kimpe, N. Chemical and enzymatic synthesis of 2-(2-carbamoylethyl)- and 2-(2-carboxyethyl)aziridines and their conversion into δ-lactams and γ-lactones. Org. Lett. 2012, 14, 106–109. [Google Scholar] [CrossRef]
- Ohta, H. Stereochemistry of enzymatic hydrolysis of nitriles. Chimia (Aarau) 1996, 50, 434–436. [Google Scholar]
- Sugai, T.; Yamazaki, T.; Yokoyama, M.; Ohta, H. Biocatalysis in Organic Synthesis: The use of nitrile- and amide-hydrolyzing microorganisms. Biosci. Biotech. Biochem. 1997, 61, 1419–1427. [Google Scholar] [CrossRef]
- Kakeya, H.; Sakai, N.; Sugai, T.; Ohta, H. Microbial hydrolysis as a potent method for the preparation of optically active nitriles, amides and carboxylic acids. Tetrahedron Lett. 1991, 32, 1343–1346. [Google Scholar] [CrossRef]
- Beard, T.; Cohen, M.A.; Parratt, J.S.; Turner, N.J.; Crosby, J.; Moilliet, J. Stereoselective hydrolysis of nitriles and amides under mild conditions using a whole cell catalyst. Tetrahedron Asymmetry 1993, 4, 1085–1104. [Google Scholar] [CrossRef]
- Layh, N.; Stolz, A.; Böhme, J.; Effenberger, F.; Knackmuss, H.J. Enantioselective hydrolysis of racemic naproxen nitrile and naproxen amide to S-naproxen by new bacterial isolates. J. Biotechnol. 1994, 33, 175–182. [Google Scholar] [CrossRef]
- Wang, M.X.; Wu, Y. Nitrile biotransformations for the synthesis of enantiomerically enriched Baylis-Hillman adducts. Org. Biomol. Chem. 2003, 1, 535–540. [Google Scholar] [CrossRef]
- Ao, Y.F.; Wang, D.X.; Zhao, L.; Wang, M.X. Synthesis of quaternary-carbon-containing and functionalized enantiopure pentanecarboxylic acids from biocatalytic desymmetrization of meso-cyclopentane-1,3-dicarboxamides. Chem. Asian J. 2015, 10, 938–947. [Google Scholar] [CrossRef]
- Yokoyama, M.; Kashiwagi, M.; Iwasaki, M.; Fuhshuku, K.I.; Ohta, H.; Sugai, T. Realization of the synthesis of α,α-disubstituted carbamylacetates and cyanoacetates by either enzymatic or chemical functional group transformation, depending upon the substrate specificity of Rhodococcus amidase. Tetrahedron Asymmetry 2004, 15, 2817–2820. [Google Scholar] [CrossRef]
- Osprian, I.; Fechter, M.H.; Griengl, H. Biocatalytic hydrolysis of cyanohydrins: An efficient approach to enantiopure α-hydroxy carboxylic acids. J. Mol. Catal. B Enzym. 2003, 24–25, 89–98. [Google Scholar] [CrossRef]
- Wegman, M.A.; Heinemann, U.; Van Rantwijk, F.; Stolz, A.; Sheldon, R.A. Hydrolysis of d,l-phenylglycine nitrile by new bacterial cultures. J. Mol. Catal. B Enzym. 2001, 11, 249–253. [Google Scholar] [CrossRef]
- Lin, Z.J.; Zheng, R.C.; Zheng, Y.G.; Shen, Y.C. Biosynthesis of 2-amino-2,3-dimethylbutyramide by nitrile hydratase from a newly isolated cyanide-resistant strain of Rhodococcus qingshengii. Biotechnol. Lett. 2011, 33, 1809–1813. [Google Scholar] [CrossRef]
- Wang, M.X.; Lin, S.J. Practical and convenient enzymatic synthesis of enantiopure α-amino acids and amides. J. Org. Chem. 2002, 67, 6542–6545. [Google Scholar] [CrossRef]
- Morán-Ramallal, R.; Liz, R.; Gotor, V. Bacterial preparation of enantiopure unactivated aziridine-2-carboxamides and their transformation into enantiopure nonnatural amino acids and vic-diamines. Org. Lett. 2007, 9, 521–524. [Google Scholar] [CrossRef]
- Leng, D.H.; Wang, D.X.; Huang, Z.T.; Wang, M.X. Highly efficient and enantioselective biotransformations of β-lactam carbonitriles and carboxamides and their synthetic applications. Org. Biomol. Chem. 2010, 8, 4736–4743. [Google Scholar] [CrossRef]
- Leng, D.H.; Wang, D.X.; Pan, J.; Huang, Z.T.; Wang, M.X. Highly efficient and enantioselective biotransformations of racemic azetidine-2-carbonitriles and their synthetic applications. J. Org. Chem. 2009, 74, 6077–6082. [Google Scholar] [CrossRef]
- Kakeya, H.; Sakai, N.; Sano, A.; Yokoyama, M.; Sugai, T.; Ohta, H. Microbiol hydrolysis of 3-substituted glutaronitriles. Chem. Lett. 1991, 20, 1823–1824. [Google Scholar] [CrossRef]
- Crosby, J.A.; Parrattt, J.S.; Turner, N.J. Enzymatic hydrolysis of prochiral dinitriles. Tetrahedron Asymmetry 1992, 3, 1547–1550. [Google Scholar] [CrossRef]
- Maddrell, S.J.; Turner, N.J.; Kerridge, A.; Willetts, A.J.; Crosby, J. Nitrile Hydratase Enzymes in Organic Synthesis. Tetrahedron Lett. 1996, 37, 6001–6004. [Google Scholar] [CrossRef]
- Kinfe, H.H.; Chhiba, V.; Frederick, J.; Bode, M.L.; Mathiba, K.; Steenkamp, P.A.; Brady, D. Enantioselective hydrolysis of β-hydroxy nitriles using the whole cell biocatalyst Rhodococcus rhodochrous ATCC BAA-870. J. Mol. Catal. B Enzym. 2009, 59, 231–236. [Google Scholar] [CrossRef]
- Wu, Z.L.; Li, Z.Y. Biocatalytic asymmetric hydrolysis of (±)-β-hydroxy nitriles by Rhodococcus sp. CGMCC 0497. J. Mol. Catal. B Enzym. 2003, 22, 105–112. [Google Scholar] [CrossRef]
- Yokoyama, M.; Imai, N.; Sugai, T.; Ohta, H. Preparation of both enantiomers of methyl 3-benzoyloxypentanoate by enzyme-catalysed hydrolysis of corresponding racemic nitrile and amide. J. Mol. Catal. B Enzym. 1996, 1, 135–141. [Google Scholar] [CrossRef]
- Vink, M.K.S.; Schortinghuis, C.A.; Luten, J.; Van Maarseveen, J.H.; Schoemaker, H.E.; Hiemstra, H.; Rutjes, F.P.J.T. A stereodivergent approach to substituted 4-hydroxypiperidines. J. Org. Chem. 2002, 67, 7869–7871. [Google Scholar] [CrossRef]
- Méndez-Sánchez, D.; López-Iglesias, M.; Gotor-Fernández, V. Hydrolases in Organic Chemistry. Recent Achievements in the Synthesis of Pharmaceuticals. Curr. Org. Chem. 2016, 20, 1186–1203. [Google Scholar] [CrossRef]
- Ma, Y.; Yu, H.; Pan, W.; Liu, C.; Zhang, S.; Shen, Z. Identification of nitrile hydratase-producing Rhodococcus ruber TH and characterization of an amiE-negative mutant. Bioresour. Technol. 2010, 101, 285–291. [Google Scholar] [CrossRef]
- Hall, M.; Faber, K.; Tasnadi, G. Hydrolysis of Amides. In Biocatalysis in Organic Synthesis 1; Faber, K., Fessner, W.-D., Turner, N.J., Eds.; Georg Thieme Verlag KG: Stuttgart, Germany, 2015; pp. 303–329. [Google Scholar]
- Ismailsab, M.; Monisha, T.R.; Reddy, P.V.; Santoshkumar, M.; Nayak, A.S.; Karegoudar, T.B. Biotransformation of aromatic and heterocyclic amides by amidase of whole cells of Rhodococcus sp. MTB5: Biocatalytic characterization and substrate specificity. Biocatal. Biotransform. 2017, 35, 74–85. [Google Scholar] [CrossRef]
- Wang, M.X.; Lu, G.; Ji, G.J.; Huang, Z.T.; Meth-Cohn, O.; Colby, J. Enantioselective biotransformations of racemic α-substituted phenylacetonitriles and phenylacetamides using Rhodococcus sp. AJ270. Tetrahedron Asymmetry 2000, 11, 1123–1135. [Google Scholar] [CrossRef]
- Effenberger, F.; Graef, B.W.; Oßwald, S. Preparation of (S)-naproxen by enantioselective hydrolysis of racemic naproxen amide with resting cells of Rhodococcus erythropolis MP50 in organic solvents. Tetrahedron Asymmetry 1997, 8, 2749–2755. [Google Scholar] [CrossRef]
- Snell, D.; Colby, J. Enantioselective hydrolysis of racemic ibuprofen amide to S-(+)-ibuprofen by Rhodococcus AJ270. Enzym. Microb. Technol. 1999, 24, 160–163. [Google Scholar] [CrossRef]
- Wu, Z.L.; Li, Z.Y. Practical synthesis of optically active α,α-disubstituted malonamic acids through asymmetric hydrolysis of malonamide derivatives with Rhodococcus sp. CGMCC 0497. J. Org. Chem. 2003, 68, 2479–2482. [Google Scholar] [CrossRef]
- Zhang, L.B.; Wang, D.X.; Wang, M.X. Microbial whole cell-catalyzed desymmetrization of prochiral malonamides: Practical synthesis of enantioenriched functionalized carbamoylacetates and their application in the preparation of unusual α-amino acids. Tetrahedron 2011, 67, 5604–5609. [Google Scholar] [CrossRef]
- Zhang, L.B.; Wang, D.X.; Zhao, L.; Wang, M.X. Synthesis and application of enantioenriched functionalized α-tetrasubstituted α-amino acids from biocatalytic desymmetrization of prochiral α-aminomalonamides. J. Org. Chem. 2012, 77, 5584–5591. [Google Scholar] [CrossRef]
- Chen, P.; Gao, M.; Wang, D.X.; Zhao, L.; Wang, M.X. Enantioselective biotransformations of racemic and meso pyrrolidine-2,5-dicarboxamides and their application in organic synthesis. J. Org. Chem. 2012, 77, 4063–4072. [Google Scholar] [CrossRef]
- Chen, P.; Gao, M.; Wang, D.X.; Zhao, L.; Wang, M.X. Practical biocatalytic desymmetrization of meso-N-heterocyclic dicarboxamides and their application in the construction of aza-sugar containing nucleoside analogs. Chem. Commun. 2012, 48, 3482–3484. [Google Scholar] [CrossRef]
- Ao, Y.F.; Zhang, L.-B.; Wang, Q.Q.; Wang, D.X.; Wang, M.X. Biocatalytic Desymmetrization of Prochiral 3-Aryl and 3-Arylmethyl Glutaramides: Different Remote Substituent Effect on Catalytic Efficiency and Enantioselectivity. Adv. Synth. Catal. 2018, 360, 4594–4603. [Google Scholar] [CrossRef]
- Venhoff, N.; Setzer, B.; Melkaoui, K.; Walker, U.A. Mitochondrial toxicity of tenofovir, emtricitabine and abacavir alone and in combination with additional nucleoside reverse transcriptase inhibitors. Antivir. Ther. 2007, 12, 1075–1085. [Google Scholar]
- Taylor, S.J.C.; Sutherland, A.G.; Lee, C.; Wisdom, R.; Thomas, S.; Roberts, S.M.; Evans, C. Chemoenzymatic Synthesis of (-)-carbovir utilising a whole cell catalysed resolution of 2-azabicyclo[2.2.1]hept-5-en-3-one. J. Chem. Soc. Chem. Commun. 1990, 16, 1120–1121. [Google Scholar] [CrossRef]
- Assaf, Z.; Eger, E.; Vitnik, Z.; Fabian, W.M.F.; Ribitsch, D.; Guebitz, G.M.; Faber, K.; Hall, M. Identification and application of enantiocomplementary lactamases for Vince lactam derivatives. ChemCatChem 2014, 6, 2517–2521. [Google Scholar] [CrossRef]
- Fukuta, Y.; Koizumi, S.; Komeda, H.; Asano, Y. A new aryl acylamidase from Rhodococcus sp. strain Oct1 acting on ω-lactams: Its characterization and gene expression in Escherichia coli. Enzym. Microb. Technol. 2010, 46, 237–245. [Google Scholar] [CrossRef]
- Asano, Y.; Fukuta, Y.; Yoshida, Y.; Komeda, H. The screening, characterization, and use of ω-laurolactam hydrolase: A new enzymatic synthesis of 12-aminolauric acid. Biosci. Biotechnol. Biochem. 2008, 72, 2141–2150. [Google Scholar] [CrossRef]
- Nagasawa, T.; Nakamura, T.; Yamada, H. Production of acrylic acid and methacrylic acid using Rhodococcus rhodochrous J1 nitrilase. Appl. Microbiol. Biotechnol. 1990, 34, 322–324. [Google Scholar] [CrossRef]
- Luo, H.; Wang, T.; Yu, H.; Yang, H.; Shen, Z. Expression and catalyzing process of the nirilase in Rhodococcus rhodochrous tg1-A6. Mod. Chem. Ind. 2006, 26, 109–111. [Google Scholar]
- Webster, N.A.; Ramsden, D.K.; Hughes, J. Comparative characterisation of two Rhodococcus species as potential biocatalysts for ammonium acrylate production. Biotechnol. Lett. 2001, 23, 95–101. [Google Scholar] [CrossRef]
- Bhalla, T.C.; Miura, A.; Wakamoto, A.; Ohba, Y.; Furuhashi, K. Asymmetric hydrolysis of α-aminonitriles to optically active amino acids by a nitrilase of Rhodococcus rhodochrous PA-34. Appl. Microbiol. Biotechnol. 1992, 37, 184–190. [Google Scholar] [CrossRef]
- Mathew, C.D.; Nagasawa, T.; Kobayashi, M.; Yamada, H. Nitrilase-Catalyzed Production of Nicotinic Acid from 3-Cyanopyridine in Rhodococcus rhodochrous J1. Appl. Environ. Microbiol. 1988, 54, 1030–1032. [Google Scholar]
- Prasad, S.; Misra, A.; Jangir, V.P.; Awasthi, A.; Raj, J.; Bhalla, T.C. A propionitrile-induced nitrilase of Rhodococcus sp. NDB 1165 and its application in nicotinic acid synthesis. World J. Microbiol. Biotechnol. 2007, 23, 345–353. [Google Scholar] [CrossRef]
- Martínková, L.; Vesela, A.B. Hydrolysis of nitriles to carboxylic acids. In Biocatalysis in Organic Synthesis 1; Faber, K., Fessner, W.-D., Turner, N.J., Eds.; Georg Thieme Verlag KG: Stuttgart, Germany, 2015; pp. 277–302. [Google Scholar]
- Gong, J.S.; Lu, Z.M.; Li, H.; Shi, J.S.; Zhou, Z.M.; Xu, Z.H. Nitrilases in nitrile biocatalysis: Recent progress and forthcoming research. Microb. Cell Fact. 2012, 11, 1–18. [Google Scholar] [CrossRef]
- Nigam, V.K.; Arfi, T.; Kumar, V.; Shukla, P. Bioengineering of Nitrilases towards Its Use as Green Catalyst: Applications and Perspectives. Indian J. Microbiol. 2017, 57, 131–138. [Google Scholar] [CrossRef]
- Archelas, A.; Iacazio, G.; Kotik, M. Epoxide hydrolase and its application in organic synthesis. In Green Biocatalysis; Patel, R.M., Ed.; John Wiley and Sons Inc.: Hoboken, NJ, USA, 2016; pp. 179–230. [Google Scholar]
- Osprian, I.; Kroutil, W.; Mischitz, M.; Faber, K. Biocatalytic resolution of 2-methyl-2-(aryl)alkyloxiranes using novel bacterial epoxide hydrolases. Tetrahedron Asymmetry 1997, 8, 65–71. [Google Scholar] [CrossRef]
- Wandel, U.; Mischitz, M.; Kroutil, W.; Faber, K. Highly selective asymmetric hydrolysis of 2,2-disubstituted epoxides using lyophilized cells of Rhodococcus sp. NCIMB 11216. J. Chem. Soc. Perkin Trans. 1 1995, 735–736. [Google Scholar] [CrossRef]
- Osprian, I.; Stampfer, W.; Faber, K. Selectivity enhancement of epoxide hydrolase catalyzed resolution of 2,2-disubstituted oxiranes by substrate modification. J. Chem. Soc. Perkin Trans. 1 2000, 3779–3785. [Google Scholar] [CrossRef]
- Woo, J.H.; Kang, K.M.; Kwon, T.H.; Park, N.H.; Lee, E.Y. Isolation, identification and characterization of marine bacteria exhibiting complementary enantioselective epoxide hydrolase activity for preparing chiral chlorinated styrene oxide derivatives. J. Ind. Eng. Chem. 2015, 28, 225–228. [Google Scholar] [CrossRef]
- Arand, M.; Hallberg, B.M.; Zou, J.; Bergfors, T.; Oesch, F.; Van der Werf, M.J.; De Bont, J.A.M.; Jones, T.A.; Mowbray, S.L. Structure of Rhodococcus erythropolis limonene-1,2-epoxide hydrolase reveals a novel active site. EMBO J. 2003, 22, 2583–2592. [Google Scholar] [CrossRef]
- Van Der Werf, M.J.; Orru, R.V.A.; Overkamp, K.M.; Swarts, H.J.; Osprian, I.; Steinreiber, A.; De Bont, J.A.M.; Faber, K. Substrate specificity and stereospecificity of limonene-1,2-epoxide hydrolase from Rhodococcus erythropolis DCL14; an enzyme showing sequential and enantioconvergent substrate conversion. Appl. Microbiol. Biotechnol. 1999, 52, 380–385. [Google Scholar] [CrossRef]
- Van der Werf, M.; Overkamp, K.; de Bont, J. DCL14 Belongs to a Novel Class of Epoxide Hydrolases. J. Bacteriol. 1998, 180, 5052–5057. [Google Scholar]
- Zheng, H.; Reetz, M.T. Manipulating the stereoselectivity of limonene epoxide hydrolase by directed evolution based in iterative saturation mutagenesis. J. Am. Chem. Soc. 2010, 132, 15744–15751. [Google Scholar] [CrossRef]
- Bottalla, A.L.; Ibrahim-Ouali, M.; Santelli, M.; Furstoss, R.; Archelas, A. Epoxide hydrolase-catalysed kinetic resolution of a spiroepoxide, a key building block of various 11-heterosteroids. Adv. Synth. Catal. 2007, 349, 1102–1110. [Google Scholar] [CrossRef]
- Kroutil, W.; Mischitz, M.; Faber, K. Derazemisation of (±)-2,3-disubstituted oxiranes via biocatalytic hydrolysis Using Bacterial Epoxide Hydrolases: Kinetics of an Enantioconvergent Process. J. Chem. Soc. Perkin Trans. 1 1997, 3629–3636. [Google Scholar] [CrossRef]
- Steinreiber, A.; Mayer, S.F.; Saf, R.; Faber, K. Biocatalytic asymmetric and enantioconvergent hydrolysis of trisubstituted oxiranes. Tetrahedron Asymmetry 2001, 12, 1519–1528. [Google Scholar] [CrossRef]
- Steinreiber, A.; Mayer, S.F.; Faber, K. Asymmetric total synthesis of a beer-aroma constituent based on enantioconvergent biocatalytic hydrolysis of trisubstituted epoxides. Synthesis (Stuttg) 2001, 13, 2035–2039. [Google Scholar] [CrossRef]
- Edegger, K.; Mayer, S.F.; Steinreiber, A.; Faber, K. Chemo-enzymatic enantio-convergent asymmetric synthesis of (R)-(+)-Marmin. Tetrahedron 2004, 60, 583–588. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Xu, Y.; Ping, L.; Zheng, Y. Cloning, sequencing, and expression of a novel epoxide hydrolase gene from Rhodococcus opacus in Escherichia coli and characterization of enzyme. Appl. Microbiol. Biotechnol. 2007, 74, 99–106. [Google Scholar] [CrossRef]
- Glueck, S.M.; Fabian, W.M.F.; Faber, K.; Mayer, S.F. Biocatalytic asymmetric rearrangement of a methylene-interrupted bis-epoxide: Simultaneous control of four asymmetric centers through a biomimetic reaction cascade. Chem. A Eur. J. 2004, 10, 3467–3478. [Google Scholar] [CrossRef]
- Ueberbacher, B.T.; Oberdorfer, G.; Gruber, K.; Faber, K. Epoxide-hydrolase-initiated hydrolysis/rearrangement cascade of a methylene-interrupted bis-epoxide yields chiral THF moieties without involvement of a “Cyclase”. ChemBioChem 2009, 10, 1697–1704. [Google Scholar] [CrossRef]
- Hellström, H.; Steinreiber, A.; Mayer, S.F.; Faber, K. Bacterial epoxide hydrolase-catalyzed resolution of a 2,2-disubstituted oxirane: Optimization and upscaling. Biotechnol. Lett. 2001, 23, 169–173. [Google Scholar] [CrossRef]
- Simeó, Y.; Faber, K. Selectivity enhancement of enantio- and stereo-complementary epoxide hydrolases and chemo-enzymatic deracemization of (±)-2-methylglycidyl benzyl ether. Tetrahedron Asymmetry 2006, 17, 402–409. [Google Scholar] [CrossRef]
- Fuchs, M.; Simeo, Y.; Ueberbacher, B.T.; Mautner, B.; Netscher, T.; Faber, K. Enantiocomplementary chemoenzymatic asymmetric synthesis of (R)- And (S)-chromanemethanol. Eur. J. Org. Chem. 2009, 2009, 833–840. [Google Scholar] [CrossRef]
- Mischitz, M.; Faber, K. Chemo-enzymatic synthesis of linalool oxide. Synlett 1996, 978–980. [Google Scholar] [CrossRef]
- Steinreiber, A.; Edegger, K.; Mayer, S.F.; Faber, K. (2R,5S)-Pityol through enzyme-triggered ring closure. Tetrahedron Asymmetry 2001, 12, 2067–2071. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, J.; Luan, Z.J.; Xu, G.C.; Park, S.; Xu, J.H. Cloning and characterization of a novel esterase from Rhodococcus sp. for highly enantioselective synthesis of a chiral cilastatin precursor. Appl. Environ. Microbiol. 2014, 80, 7348–7355. [Google Scholar] [CrossRef]
- Luan, Z.J.; Yu, H.L.; Ma, B.-D.; Qi, Y.K.; Chen, Q.; Xu, J.H. Dramatically improved performance of an esterase for cilastatin synthesis by cap domain engineering. Ind. Eng. Chem. Res. 2016, 55, 12167–12172. [Google Scholar] [CrossRef]
- Pogorevc, M.; Strauss, U.T.; Hayn, M.; Faber, K. Novel carboxyl esterase preparations for the resolution of linalyl acetate. Mon. Chem. 2000, 131, 639–644. [Google Scholar] [CrossRef]
- Akutsu-Shigeno, Y.; Adachi, Y.; Yamada, C.; Toyoshima, K.; Nomura, N.; Uchiyama, H.; Nakajima-Kambe, T. Isolation of a bacterium that degrades urethane compounds and characterization of its urethane hydrolase. Appl. Microbiol. Biotechnol. 2006, 70, 422–429. [Google Scholar] [CrossRef]
- Lorenzen, J.; Janke, R.; Waldow, A.; Qoura, F.; Loll, B.; Brück, T. Rhodococcus erythropolis Oleate Hydratase: A New Member in the Oleate Hydratase Family Tree—Biochemical and Structural Studies. ChemCatChem 2017, 10, 407–414. [Google Scholar] [CrossRef]
- Engleder, M.; Pavkov-Keller, T.; Emmerstorfer, A.; Hromic, A.; Schrempf, S.; Steinkellner, G.; Wriessnegger, T.; Leitner, E.; Strohmeier, G.A.; Kaluzna, I.; et al. Structure-Based Mechanism of Oleate Hydratase from Elizabethkingia meningoseptica. ChemBioChem 2015, 16, 1730–1734. [Google Scholar] [CrossRef]
- Park, A.K.; Lee, G.H.; Kim, D.W.; Jang, E.H.; Kwon, H.T.; Chi, Y.M. Crystal structure of oleate hydratase from Stenotrophomonas sp. KCTC 12332 reveals conformational plasticity surrounding the FAD binding site. Biochem. Biophys. Res. Commun. 2018, 499, 772–776. [Google Scholar] [CrossRef]
- Volkov, A.; Khoshnevis, S.; Neumann, P.; Herrfurth, C.; Wohlwend, D.; Ficner, R.; Feussner, I. Crystal structure analysis of a fatty acid double-bond hydratase from Lactobacillus acidophilus. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 648–657. [Google Scholar] [CrossRef]
- Busch, H.; Tonin, F.; Alvarenga, N.; van den Broek, M.; Lu, S.; Daran, J.-M.; Hanefeld, U.; Hagedoorn, P.-L. Exploring the abundance of oleate hydratases in the genus Rhodococcus—Discovery of novel enzymes with complementary substrate scope. Manuscript in preparation.
- Brueck, T.; Lorenzen, J. A Process for the Cell-Free Enzymtaic Production of 10-Hydroxystearic Acid (10-HSA) from Bio-Based Oils for Lubricant Formulation. WO 2,019,063,718, 27 September 2018. [Google Scholar]
- Holland, H.L.; Gu, J.X. Preparation of (R)-3-hydroxy-3-alkylbutanolides by biocatalytic hydration of 3-alkyl-2-butenolides using Rhodococcus rhodochrous. Biotechnol. Lett. 1998, 20, 1125–1126. [Google Scholar] [CrossRef]
- Chen, B.-S.; Resch, V.; Otten, L.G.; Hanefeld, U. Enantioselective Michael Addition of Water. Chem. A Eur. J. 2015, 21, 3020–3030. [Google Scholar] [CrossRef]
- Busch, H.; Alvarenga, N.; Abdelraheem, E.; Hoek, M.; Hagedoorn, P.-L.; Hanefeld, U. Re-investigation of hydration potential of Rhodococcus whole-cell biocatalysts towards Michael acceptors. ChemCatChem 2019. [Google Scholar] [CrossRef]
- He, Y.C.; Tao, Z.C.; Zhang, D.P.; Yang, Z.X.; Gao, S.; Ma, C.L. Biotransformation of 1,3-propanediol cyclic sulfate and its derivatives to diols by Rhodococcus sp. Biotechnol. Lett. 2015, 37, 183–188. [Google Scholar] [CrossRef]
- Gadler, P.; Faber, K. New enzymes for biotransformations: Microbial alkyl sulfatases displaying stereo- and enantioselectivity. Trends Biotechnol. 2007, 25, 83–88. [Google Scholar] [CrossRef]
- Pogorevc, M.; Kroutil, W.; Wallner, S.R.; Faber, K. Enantioselective stereoinversion in the kinetic resolution of rac-sec-alkyl sulfate esters by hydrolysis with an alkylsulfatase from Rhodococcus ruber DSM 44541 furnishes homochiral products. Angew. Chem. Int. Ed. 2002, 41, 4052–4054. [Google Scholar] [CrossRef]
- Pogorevc, M.; Faber, K. Purification and characterization of an inverting stereo- and enantioselective sec-alkylsulfatase from the gram-positive bacterium Rhodococcus ruber DSM 44541. Appl. Environ. Microbiol. 2003, 69, 2810–2815. [Google Scholar] [CrossRef]
- Pogorevc, M.; Faber, K. Enantioselective stereoinversion of sec-alkyl sulfates by an alkylsulfatase from Rhodococcus ruber DSM 44541. Tetrahedron Asymmetry 2002, 13, 1435–1441. [Google Scholar] [CrossRef]
- Pogorevc, M.; Strauss, U.T.; Riermeier, T.; Faber, K. Selectivity-enhancement in enantioselective hydrolysis of sec-alkyl sulfates by an alkylsulfatase from Rhodococcus ruber DSM 44541. Tetrahedron Asymmetry 2002, 13, 1443–1447. [Google Scholar] [CrossRef]
- He, Y.C.; Ma, C.L.; Yang, Z.X.; Zhou, M.; Xing, Z.; Ma, J.T.; Yu, H.L. Highly enantioselective oxidation of phenyl methyl sulfide and its derivatives into optically pure (S)-sulfoxides with Rhodococcus sp. CCZU10-1 in an n-octane-water biphasic system. Appl. Microbiol. Biotechnol. 2013, 97, 10329–10337. [Google Scholar] [CrossRef]
- Li, A.T.; Zhang, J.D.; Xu, J.H.; Lu, W.Y.; Lin, G.Q. Isolation of Rhodococcus sp. strain ECU0066, a new sulfide monooxygenase-producing strain for asymmetric sulfoxidation. Appl. Environ. Microbiol. 2009, 75, 551–556. [Google Scholar] [CrossRef]
- Li, A.T.; Zhang, J.D.; Yu, H.L.; Pan, J.; Xu, J.H. Significantly improved asymmetric oxidation of sulfide with resting cells of Rhodococcus sp. in a biphasic system. Process Biochem. 2011, 46, 689–694. [Google Scholar] [CrossRef]
- Lei, B.; Shiao-Chun, T.U. Gene overexpression, purification, and identification of a desulfurization enzyme from Rhodococcus sp. strain IGTS8 as a sulfide/sulfoxide monooxygenase. J. Bacteriol. 1996, 178, 5699–5705. [Google Scholar] [CrossRef]
Scheme 1.
Deracemization of racemic alcohols using a two-enzyme system with stereocomplementary ADHs [23].
Scheme 1.
Deracemization of racemic alcohols using a two-enzyme system with stereocomplementary ADHs [23].

Scheme 2.
Enzymatic cascade for selective synthesis of epoxy ketones and allylic epoxy alcohols [32].
Scheme 2.
Enzymatic cascade for selective synthesis of epoxy ketones and allylic epoxy alcohols [32].

Scheme 3.
KRED catalyzed ketone reduction in a multistep chemo-enzymatic reaction sequence to pharmacologically active compounds (R)-ramatroban and (R)-frovatriptan [35,36].

Scheme 4.
Asymmetric bioreduction of N-Boc-3-piperidone (12) catalyzed by KRED [39].
Scheme 4.
Asymmetric bioreduction of N-Boc-3-piperidone (12) catalyzed by KRED [39].

Scheme 5.
R. erythropolis oxidase catalyzed process to produce (R)-iso-propylideneglycerol (16), which is used as C3-synthon for (S)-metoprolol (17) [44].
Scheme 5.
R. erythropolis oxidase catalyzed process to produce (R)-iso-propylideneglycerol (16), which is used as C3-synthon for (S)-metoprolol (17) [44].

Scheme 6.
Representative bioconversion catalyzed by P450RhF monooxygenase showing aromatic hydroxylation, dealkylation, and sulfoxidation potential [50].
Scheme 6.
Representative bioconversion catalyzed by P450RhF monooxygenase showing aromatic hydroxylation, dealkylation, and sulfoxidation potential [50].

Scheme 7.
Rhodococcus Baeyer-Villiger monooxygenase catalyzed oxidation of 4,4-disubstituted c-hexanones (24) [55].
Scheme 7.
Rhodococcus Baeyer-Villiger monooxygenase catalyzed oxidation of 4,4-disubstituted c-hexanones (24) [55].

Scheme 8.
Representative Baeyer-Villiger oxidations of bridged c-ketones (27,29) catalyzed by CHMOs from Rhodococcus [56].
Scheme 8.
Representative Baeyer-Villiger oxidations of bridged c-ketones (27,29) catalyzed by CHMOs from Rhodococcus [56].

Scheme 9.
‘Semisynthetic’ production of polydihydrocarvide (32/33) using a triple mutant of CHMOPhi1 [64].
Scheme 9.
‘Semisynthetic’ production of polydihydrocarvide (32/33) using a triple mutant of CHMOPhi1 [64].

Scheme 10.
Ene-reductase catalyzed reduction of α,β-unsaturated maleimides (35) [99].
Scheme 10.
Ene-reductase catalyzed reduction of α,β-unsaturated maleimides (35) [99].

Scheme 11.
Reductive amination of representative ketones catalyzed by AmDH engineered from R. sp. M4 [103,105].

Scheme 12.
General aldoxime-nitrile pathway in microbes with synthetically used enzymes from Rhodococcus highlighted [109].
Scheme 12.
General aldoxime-nitrile pathway in microbes with synthetically used enzymes from Rhodococcus highlighted [109].

Scheme 13.
Patented biocatalytic citronellyl nitrile (44) production by dehydration of citronellal oxime (43) catalyzed by aldoxime dehydratase [122].
Scheme 13.
Patented biocatalytic citronellyl nitrile (44) production by dehydration of citronellal oxime (43) catalyzed by aldoxime dehydratase [122].

Scheme 14.
Nitrile hydratase catalyzed bioconversion of adiponitrile (45) yielding 5-cyanovaleramide (46) using whole-cells of R. ruber GCMCC3090 [139].
Scheme 14.
Nitrile hydratase catalyzed bioconversion of adiponitrile (45) yielding 5-cyanovaleramide (46) using whole-cells of R. ruber GCMCC3090 [139].

Scheme 15.
Dynamic kinetic resolution of rac-aminonitriles to produce chiral α-amino acids (50,51). The first step is catalyzed by a nitrile hydratase from R. opacus 71D [140].
Scheme 15.
Dynamic kinetic resolution of rac-aminonitriles to produce chiral α-amino acids (50,51). The first step is catalyzed by a nitrile hydratase from R. opacus 71D [140].

Scheme 16.
Chemo-enzymatic cascade reaction to synthesize piperidin-2-ones (54). The enzymatically catalyzed step was performed by the soluble nitrile hydratase from R. erythropolis NCIMB 11540 [142].
Scheme 16.
Chemo-enzymatic cascade reaction to synthesize piperidin-2-ones (54). The enzymatically catalyzed step was performed by the soluble nitrile hydratase from R. erythropolis NCIMB 11540 [142].

Scheme 17.
General scheme of the NHase/amidase catalyzed reaction.

Scheme 18.
Synthesis of enantioenriched functionalized α-tetrasubstituted α-amino acids (60) through amidase-catalyzed hydrolysis [174].
Scheme 18.
Synthesis of enantioenriched functionalized α-tetrasubstituted α-amino acids (60) through amidase-catalyzed hydrolysis [174].

Scheme 19.
Chemo-enzymatic syntheses of dihydroquinolinone (64) and δ-lactone (65) [177].
Scheme 19.
Chemo-enzymatic syntheses of dihydroquinolinone (64) and δ-lactone (65) [177].

Scheme 20.
Kinetic resolution of Vince-lactam using an amidase from R. globerulus with subsequent follow-up chemistry of enantiopure products leading to antiviral compounds carbovir (69) and abacavir (70) and antidiabetic melogliptin (71) [180].
Scheme 20.
Kinetic resolution of Vince-lactam using an amidase from R. globerulus with subsequent follow-up chemistry of enantiopure products leading to antiviral compounds carbovir (69) and abacavir (70) and antidiabetic melogliptin (71) [180].

Scheme 21.
Optimized two-enzyme system for hydrolytic kinetic resolution of trans-spiroepoxide (72) using AnEH and LEH. Enantiopure spiroepoxides (75,76) can be used in the synthesis of enantiopure 11-heterosteroids (79) [201].
Scheme 21.
Optimized two-enzyme system for hydrolytic kinetic resolution of trans-spiroepoxide (72) using AnEH and LEH. Enantiopure spiroepoxides (75,76) can be used in the synthesis of enantiopure 11-heterosteroids (79) [201].

Scheme 22.
Stereochemical course of hydrolysis/cyclisation reaction catalyzed by two Rhodococcus representatives.
Scheme 22.
Stereochemical course of hydrolysis/cyclisation reaction catalyzed by two Rhodococcus representatives.

Scheme 23.
Natural product synthesis of (A) cis-(88) and trans-linalool oxides (89) and (B) pityol (93) using Rhodococcus whole-cell biocatalysts.
Scheme 23.
Natural product synthesis of (A) cis-(88) and trans-linalool oxides (89) and (B) pityol (93) using Rhodococcus whole-cell biocatalysts.

Scheme 24.
Enantioselective resolution of a racemic mixture of DMCPCM (94) to yield (S)-DMCPCA (96)—a precursor for the drug cilastatin (97) [214].
Scheme 24.
Enantioselective resolution of a racemic mixture of DMCPCM (94) to yield (S)-DMCPCA (96)—a precursor for the drug cilastatin (97) [214].

Figure 1.
Exemplary display of complementary substrate scope of two Ohy from Rhodococcus. The bold number indicates the location of the hydroxy-group in the final hydrated product.
Figure 1.
Exemplary display of complementary substrate scope of two Ohy from Rhodococcus. The bold number indicates the location of the hydroxy-group in the final hydrated product.

Scheme 25.
Retention or conversion during enzymatic sulfate ester hydrolysis catalyzed by sulfatase.
Scheme 25.
Retention or conversion during enzymatic sulfate ester hydrolysis catalyzed by sulfatase.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Busch, H.; Hagedoorn, P.-L.; Hanefeld, U. Rhodococcus as a Versatile Biocatalyst in Organic Synthesis. Int. J. Mol. Sci. 2019, 20, 4787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194787
AMA Style
Busch H, Hagedoorn P-L, Hanefeld U. Rhodococcus as a Versatile Biocatalyst in Organic Synthesis. International Journal of Molecular Sciences. 2019; 20(19):4787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194787
Chicago/Turabian StyleBusch, Hanna, Peter-Leon Hagedoorn, and Ulf Hanefeld. 2019. "Rhodococcus as a Versatile Biocatalyst in Organic Synthesis" International Journal of Molecular Sciences 20, no. 19: 4787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194787
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.