The Pivotal Role of TRP Channels in Homeostasis and Diseases throughout the Gastrointestinal Tract



Department of Cellular, Computational and Integrative Biology (CIBIO), University of Trento, Via Sommarive 9, 38123 Povo (Tn), Italy
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(21), 5277; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215277
Submission received: 30 September 2019
/
Revised: 20 October 2019
/
Accepted: 22 October 2019
/
Published: 24 October 2019
(This article belongs to the Special Issue Channels and Transporters in Cells and Tissues)
Abstract
:The transient receptor potential (TRP) channels superfamily are a large group of proteins that play crucial roles in cellular processes. For example, these cation channels act as sensors in the detection and transduction of stimuli of temperature, small molecules, voltage, pH, and mechanical constrains. Over the past decades, different members of the TRP channels have been identified in the human gastrointestinal (GI) tract playing multiple modulatory roles. Noteworthy, TRPs support critical functions related to the taste perception, mechanosensation, and pain. They also participate in the modulation of motility and secretions of the human gut. Last but not least, altered expression or activity and mutations in the TRP genes are often related to a wide range of disorders of the gut epithelium, including inflammatory bowel disease, fibrosis, visceral hyperalgesia, irritable bowel syndrome, and colorectal cancer. TRP channels could therefore be promising drug targets for the treatment of GI malignancies. This review aims at providing a comprehensive picture of the most recent advances highlighting the expression and function of TRP channels in the GI tract, and secondly, the description of the potential roles of TRPs in relevant disorders is discussed reporting our standpoint on GI tract–TRP channels interactions.
1. Introduction
The primary function of the gastrointestinal (GI) tract is the transport, digestion, and absorption of nutrients and liquids. In addition, secretion of enzymes, regulation of homeostatic processes, appetite, bacterial catabolism, defense against toxins and pathogens, and formation of feces are daily carried out as well [1,2]. These regulations of GI functions depends on an appropriate set of several physiologic signals from the luminal contents to nervous, immune, and vascular systems [3,4]. However, the gut lumen does not stand alone, indeed the gut microbiota, composed of trillions of commensal bacteria, colonizes our intestine. These gut microbiota–diet interactions produce a large amount of bioactive metabolites that play crucial roles in the digestion of food, interaction with the gut epithelium, modulation of microorganisms, and development of the immune system [3,4]. The food we eat interacts with the gut epithelium, and, indirectly, with two general types of nerve fibers that broadly innervate the GI tract. The extrinsic afferent nerves originate from the dorsal, nodose and other ganglia, transport signals from the gut to the central nervous system. By contrast, the local nervous system, also known as enteric nervous system (ENS), establishes a widespread neural network enveloped in the wall of the gut. The ENS controls GI tract motility, peristalsis, ion and water secretion and detect chemical, osmolarity, or mechanical changes in the gut, among other functions [5]. As a result, the GI tract possess an elaborate and complex structure of molecular sensors that analyze the composition of food, modulate the secretory activity, and recognize adverse conditions in the lumen and wall of the alimentary canal.
Transient receptor potential (TRP) ion channel family are expressed all along the GI system where they regulate a plethora of functional features acting as molecular sensors and transducers [6,7]. In humans, the TRP family is composed of 28 members separated into six subfamilies (e.g., TRPC1-7, TRPV1-6, TRPM1-8, TRPA1, TRPML1-3, and TRPP1-2), which are mainly permeable to Na+ and Ca2+ and also allow moderate permeability to other physiological and toxic cations [8,9]. Recently, structural studies have revealed the architecture of TRP channels. These channels are homotetramers and each subunit consists of six transmembrane (TM) segments flanked by cytosolic N- and C-terminal domains with an ion permeable pore in the fifth and sixth TM segments [9,10,11,12]. They are extensively disseminated in the central and peripheral nervous systems, mainly in sensory neurons, but also in non-neuronal cells of mammalians. TRP channels act as molecular sensors of multiple and specific stimuli, for example, they can be gated by temperature, either heat (e.g., TRPV1) or cold (e.g., TRPA1 and TRPM8), taste, pH, osmolality, mechanical forces, bioactive substances (e.g., neurotransmitters and hormones), and by several natural and artificial ligands including pungent and cooling agents, fragrant components, dietary ingredients (e.g., chemicals and metabolites), environmental irritants, noxious substances, and inflammatory mediators [13,14,15]. Generally, these exogenous and/or local endogenous stimuli activate channels leading to plasma membrane depolarization and, eventually, to Ca2+ influx which prompts several signal transduction mechanisms (e.g., action potentials) which reach the autonomous and central nervous system, causing reflex responses and sensations [16,17]. Interestingly, a large number of disorders have been associated with TRP channel dysfunctions, such as anomalous expression levels, cellular relocation or mutations indicating their essential role in homeostasis and diseases [15,18,19,20].
This review describes the functions of TRP channel expressed in the GI tract, with special emphasis on their physiological roles. The implications of specific TRP channels on visceral taste, chemical and mechanical stimulation, as well as, their interconnections with gut microbial metabolites produced in the alimentary canal is also discussed. Lastly, we summarize what is known on the involvement of TRPs in GI diseases/disorders and the potential use of pharmacological regulators in the treatment of these malignancies.
2. Expression and Pathophysiological Roles of TRP Channels in the GI Tract
In the vast majority of cases, the TRP family are expressed by primary afferent sensory neurons that arise from ganglia and ENS neurons which, conjunctly, innervate the GI tract [21,22]. However, recent studies have also demonstrated the presence of TRP channels in GI non-neuronal cells, as enterocytes and enteroendocrine cells (including enterochromaffin cells) [7,23]. One of the most studied members of the TRP family is TRPV1, which is well-known as the capsaicin receptor, the irritating compound of hot chili peppers [10,24]. TRPV1 is largely expressed in the GI tract, mainly in primary afferent sensory neurons and in ENS neurons, but also in mucosal epithelial cells and enteroendocrine cells [25,26]. TRPV4, another member of the vanilloid subfamily, has been described to be expressed by primary afferent sensory neurons, epithelial cells, and infiltrated inflammatory cells in the intestine [27,28]. TRPA1 is one of the most promiscuous chemosensors, as it can be activated by a large number of dietary and noxious molecules such as isothiocyanates (horseradish, mustard oil, wasabi, etc.) and allicin (garlic), prostaglandins, exogenous irritants and products of oxidative stress [29]. This channel is expressed by ganglia and enteric primary neurons which project to the gut, and by mucosal cells [21]. Finally, two members of the TRPM subfamily are broadly presents in the GI tract. TRPM5 channels are located on the gut chemosensory cells, and lingual taste buds receptors cells [30,31]. On the other hand, TRPM8 is mostly expressed by primary afferent neurons which operates as “cold chemosensor”, being activated by cold temperatures and several chemicals such as menthol, geraniol, and linalool, to name only a few [32,33,34,35] (Figure 1).
From a cellular point of view, TRP channels display, at least, three functional roles in the GI tract: (i) molecular sensors and primary transductors of chemical and physical stimuli, (ii) effectors of other ion channels and receptors, and (iii) conventional ion channel, enabling the transport of cations across the plasma membrane [36,37]. For example, TRPA1, TRPM5/8, and TRPV1/4 recognize and transduce the signals coming from metabolites and noxious toxins contained in food, playing a key role in the intrinsic control of GI functions, in particular chemesthesis, taste, mechanosensation, visceral thermo- and nociception, and pain [38,39]. In addition, TRP channels are involved in secretory mechanisms of the digestive tract, modulate the alimentary canal motility, contribute in the absorption of Ca2+ and Mg2+ and, finally, are implicated in the control of membrane potential and excitability of neurons and epithelial cells [40,41].
Mutations or changes in the expression or function of TRPs can provoke diseases of the GI system, indicating that the achievement of a threshold of TRP function is critical to preserve normal physiological activity. These dysfunctions include, among others, pain and heartburn, visceral hypersensitivity, irritable bowel syndrome, inflammatory bowel diseases, gastroesophageal reflux disease, intestinal fibrosis, hypomagnesemia, hypocalcemia, and GI cancers. In the next sections, we summarize the current findings regarding the role of TRP proteins in the pathogenesis of some of the most prevalent GI disorders in human (Table 1).
2.1. Visceral Hypersensitivity and Irritable Bowel Syndrome
Sensory signals are dependent on the activities of various ion channels expressed by sensory neurons. The input signals generating in the gut are transported by primary sensory neurons, with their cell body in the dorsal root ganglion, and then, as action potentials, to second-order neurons of the spinal cord. This sensorial information is eventually transmitted to neuronal centers in the thalamus and the brain leading to conscious perception [86,87].
Irritable bowel syndrome (IBS) is the most recurrently diagnosed disorder by gastroenterologists worldwide, affecting more than 10% of the Western population [88,89,90]. IBS is a group of gastrointestinal symptoms characterized by chronic or recurrent abdominal pain and/or discomfort associated with altered defecation pattern without any evidence of an organic damage [90,91]. Frequently, people affected by IBD also present other disorders, such as chronic fatigue syndrome, anxiety, and major depression. The main mediator of the increased perception of pain arises from the gut and experienced by IBS patients is the Visceral Hypersensitivity (VH), which is defined as enhanced perception of noxious stimulus (hyperalgesia) and altered sensation of normal physiological stimuli to the abdominal region (allodynia) [92,93]. The pathogenesis of VH is complex and multifactorial involving neuronal, immune, and endocrine signaling pathways, but also can result from aberrant signaling from the gut to the central nervous system or vice versa [89]. Although the exact pathophysiological mechanism of VH is not fully understood, the upregulation and/or sensitization of nociceptors, including opioid and cannabinoid receptors and, in particular, TRP channels, is recognized to play a pivotal role in altered pain signaling in patients affected by IBS [6,56,94,95,96].
So far, TRPV1, TRPV4, TRPA1, and TRPM8 channels have been investigated for their implication in visceral pain. Evidence suggests that TRPV1 may be involved in pain and hyperalgesia [42]. In this frame, biopsies from patients affected by IBS reported that nerve fibers expressing TRPV1 in colon was notably increased in number and this increase correlated with pain severity [43,44]. Moreover, TRPV1 expression is improved in preclinical models of VH [45], whereas sensitivity to colorectal pain is decreased by TRPV1 antagonists and reduced in Trpv1 knock-out mice [46,47]. It has been also demonstrated that the upregulation and sensitization of TRPV1 causing hyperalgesia is influenced, at least in part, by inflammatory mediators and endocrine factors produced by the gut microbiota such as histamine, 5-hydroxytryptamine (5-HT) and nerve growth factor (NGF) [45,48,49]. For instance, NGF can raise the levels of TRPV1 and promote its insertion into the plasma membrane of peripheral nerve endings [49]. Strikingly similar to TRPV1, manifold evidences support that TRPV4 activation triggers VH, playing a significant role in hyperalgesia [42,57]. Expressed in the extrinsic primary afferents, TRPV4 potentiate the pro-algesic effects of histamine and 5-HT [56] and co-localize with protease-activated receptor-2 (PAR-2) which mutually cooperate causing mechanical hyperalgesia in the mouse colon [58,59]. The excitatory effect of TRPV4 on colonic afferents is potentiated by PAR-2 agonists [60], while the nociceptor sensitivity is alleviated by application of the TRPV4 antagonist HC067047, further emphasizing the potential role of TRPV4 in VH [61].
Recent studies have recognized that TRPA1 participates in inflammatory responses and the establishment of mechanical and chemical colonic hypersensitivity [21,75]. TRPA1 stimulation induces the Ca2+-mediate secretion of substance P (SP) and calcitonin gene-related peptide (CGRP) and other transmitters from afferent nerve fiber, which eventually results in inflammation and VH [76]. Several murine models of IBS present increased TRPA1 function in sensory ganglia and colon [77]. The same applies to humans, where the upregulation of TRPA1 in biopsies of IBD patients has been observed [78]. Meseguer et al. reported that bacterial lipopolysaccharides (LPS) also may activate TRPA1 channels inducing acute gut inflammation and visceral pain associating microbiome – gut epithelium interactions with the potential role of microorganisms and microbial metabolites [79]. These observations suggest that TRPA1 may be involved in the progression and subsistence of VH [80,81].
Contrary to TRPV1, TRPV4, and TRPA1, TRPM8 activation may attenuate pain through suppression of TRPV1 activity and the inhibition of TRPA1 function [68,69]. The analgesic effects mediated by the activation of TRPM8 can alleviate neuropathic and VH perception or mitigating cold hypersensitivity in inflammatory and nerve-damage [42,70]. Numerous studies have demonstrated that TRP channels are also co-expressed on sensory neurons and frequently have been found upregulated or sensitized in biopsies from IBS patients, suggesting an interconnection among TRPs. As a matter of fact, almost all TRPA1-positive sensory neurons co-express TRPV1 and the gating of TRPA1 can modulate TRPV1 activity [97]. Moreover, intraperitoneal co-injection of a TRPV1 inhibitor and a TRPA1 antagonist in a mouse model of experimental colitis results in a substantial reduction of VH compared to injection of the antagonists separately, suggesting a synergistic effect [97]. These data suggest that the interconnection of TRP channels can assist to pain sensitivity in the GI tract and may serve as novel therapeutic targets, which may potentially be associated with the diet–gut microbiota interactions.
2.2. Inflammatory Bowel Diseases
Trauma or diseases occurring along the GI tract may cause tissue damage leading to the release of inflammatory mediators that activate local neurons and other cells. The secretion of inflammatory molecules is mainly triggered by fibroblasts, epithelial and immune cells surrounding the injured tissue. Thus, sensory neurons provoke inflammatory response, sensing sites of inflammation and promoting protective or dolorous behaviors.
Inflammatory bowel diseases (IBDs) are the most common chronic and relapsing disorders of the GI tract, characterized by severe inflammation resulting from a complex interaction between genetic and environmental factors [98,99]. The major clinical symptoms of IBD comprise abdominal pain, bloody diarrhea, gastrointestinal bleeding, weight loss, fever, and anemia. IBD includes two main subtypes: Crohn’s disease (CD) and ulcerative colitis (UC) [100]. CD affects the whole intestine, as well as the mouth, esophagus, stomach, and the anus, while UC affects only the mucosa of the colon and the rectum. IBDs can be a painful and debilitating diseases with potentially life-threatening complications and poor quality of life for patients [101]. This explains the growing interest in understanding the complex pathophysiological mechanisms of IBD, the identification of the key mediators implicated in the disorders and, eventually, the finding of new therapeutic targets [102]. The involvement of TRP channels has been widely studied in the context of gut inflammation [28,44,76] (Figure 2).
Pro-inflammatory molecules, such as prostaglandins, bradykinin, or ATP, can sensitize TRPV1 and increase the probability of channel gating by heat and agonists (i.e., capsaicin) [103]. Colonic biopsies from patients affected by functional inflammatory disorders (CD and UC) present upregulation of TRPV1 in nociceptive afferent neurons. In this way, it was demonstrated that in the mouse colon TRPV1 enables an upregulated release of the neuropeptides CGRP and SP [44,50]. Because of the fact that TRPV1 mainly operates as an intermediary transducer, interceding in visceral nociception and intestinal inflammation after gut epithelial injury [51,52]. Mounting evidence indicates that TRPV4 exerts a pro-inflammatory role in the GI tract. TRPV4 was found overexpressed in inflamed gut tissue of human CD and, mostly, UC patients and its activation induces somatic and visceral pain [57,62,63]. Further in vitro studies on epithelial expressed TRPV4 have also demonstrated that the channel activation triggers IL-8 production in mucosa cells, suggesting that TRPV4 may play a role in gut inflammation via a non-neuronal mechanism [64]. Analogously to TRPV1, the activation of TRPA1 on visceral sensory neurons induces the secretion of neuropeptides, with consequent vasodilatation, inflammation and hyperalgesia in the gut [81,82]. Likewise, TRPA1 is unequivocally overexpressed in models of colitis in mice and in colon biopsies obtained from human UC and CD patients [78]. However, different lines of evidence have disclosed a protective role of TRPA1 against gut inflammation. TRPA1 may facilitate the restoration of tissue damages in IBD. This helpful effect seems due to the activation of sensory TRPA1 channels but also could involve those expressed in gut epithelial and immune cells, which antagonize the pro-inflammatory activities of sensory TRPV1 channels via neuropeptide release [83]. Although little is known about the TRPM8 involvement in IBD, it has been proposed that the activation of TRPM8 elicit an anti-inflammatory effect via inhibition of neuropeptides release [104].
2.3. Intestinal Fibrosis
Fibrosis is an essential reparative mechanism that occurs after tissue injury or damage. In physiological conditions, fibrosis is an extremely coordinated process, however, when the inflammation persists, myofibroblasts continuously secrete extracellular matrix factors affecting the architecture of the tissue. Therefore, fibrosis is a frequent outgrowth of chronic inflammation related to terminal diseases of many vital organs such as lung, liver, heart, and intestines, being a major mortality risk in developed countries [105].
Intestinal fibrosis is the major complication of IBD [106,107]. The massive stratification of fibrotic tissue increases the consistency of the bowel wall, thus, restraining the elasticity and the function of the affected area. Current anti-inflammatory therapies have had a limited efficacy against fibrosis in CD patients [108]. For this reason, it is highly required to find new treatments taking into account mechanisms of fibrosis [107,109,110].
An intricate set of cytokines, growth factors, and chemokines are involved in the recruitment, differentiation, activation, and regulation of myofibroblasts, therefore, with the intestinal fibrogenesis [111,112]. The main player of this polypeptides’ cocktail is TGF-β, which supports multiple aspects of myofibroblast functions in fibrogenesis such as transformation, proliferation, migration, resistance to apoptosis, stress fiber formation, and collagen synthesis [107,113]. TGF-β and its receptors are up-regulated in the inflamed intestines of IBS patients [114,115,116] and, interestingly, several TGF-β pathways have relevant connections to intracellular Ca2+ homeostasis and dynamics which, in turn, are mediated by some members of the TRP superfamily [107,117,118]. For example, TRPC3, another member of the TRP superfamily, plays a crucial role in the progression of fibroproliferative disorders including renal [119] and myocardial fibrosis [120,121]. On the other hand, it has been described that TRPA1 [84] and TRPC6 [72] channels are involved in human intestinal fibrogenesis. Recently, results indicate that the activation of TRPA1 expressed in myofibroblast, in addition to its anti-inflammatory function in IBD [83], might have significant therapeutic relevance in preventing fibrosis [85]. On the other hand, TRPC6 channels have gained growing importance in the development of fibrosis, especially as mediators of myofibroblast function [73]. It was found that TGF-β stimulation increased TRPC6 expression in fibroblasts and that the inhibition of calcineurin, a downstream mediator of TRPC6-dependent Ca2+ signaling, resulted in reduced myofibroblast differentiation [72]. Similar results have been obtained in vitro treating InMyoFibs human intestinal myofibroblast cells with TGF-β. These cells respond to TGF-β stimulation increasing the production of α-smooth muscle actin (α-SMA) stress fiber along with improved synthesis and functionality of TRPC6 channels [74]. These evidences are consistent with the assumption that increased TRPC6 channel activity is needed for TGF-β-mediated myofibroblast differentiation and fibrosis development. However, in contrast with the above results, the in vitro knockdown of TRPC6 or its pharmacological inhibition also increases the levels of anti-fibrotic IL-10 and IL-11 and the expression of collagen in myofibroblast [74]. These discordant effects of TGF-β related to myofibroblast TRPC6 channel are difficult to elucidate with a single mechanism, suggesting that TRPC6 may be involved in fibrosis in a very elaborate way. Although more exhaustive investigation is necessary to decipher the interconnection between TGF-β and TRPC6, these results contributed to our knowledge about the pathogenesis of intestinal fibrosis and may be beneficial to establish a new therapeutic strategy for chronic IBDs such as CD [71].
3. TRP Channels in Colorectal Cancers
Colorectal cancer (CRC), the most prevalent cancer of the GI tract, represents the third most commonly diagnosed malignancy worldwide in men and women and the second largest cause of death in Europe related to cancer [122,123]. Growth factors, oncogenes and receptors play a critical role in the tumorigenesis, and the propagation of CRC. However, other environmental and genetic factors play significant roles in the development of CRC [124]. Over the last two decades, accumulating evidence has demonstrated that several members of the TRP family channels show altered expression and activity in cancer cells [125,126]. Levels of expression of TRPC, TRPM, and TRPV proteins depend on the cancer stage. Those differences have been recognized mainly in melanoma, glioma and prostate, breast, kidney, and bladder cancers, suggesting an oncogenic role for some TRPs while others may function as tumor suppressors [18,127,128]. TRPs activity implicates many aspects of cancer development or “hallmarks of cancer”, such as tumor migration and invasion, exaggerated cell proliferation, cell survival, angiogenesis, and enhanced resistance to cell death [129,130,131].
To date, TRPV1 and TRPV6 have been exclusively recognized as relevant in CRC carcinogenesis. As mentioned above, the TRPV1 channel is also involved in IBD, a chronic disorder strongly correlated with risk of CRC [53]. Animal models have demonstrated a protective role for TRPV1 in the tumorigenesis of colon cancer, indeed TRPV1- Knockout mice exhibited lower expression of anti-inflammatory neuropeptides and greater occurrence and number of CRC compared with controls [54]. In accordance with these data, Sung and colleagues demonstrated that agonist-mediated activation of TRPV1 induced apoptosis of CRC cells [55]. On the other hand, an overexpression of TRPV6 in SW480 colorectal cancer cell lines and colon carcinomas have been associated with an enhanced proliferation of malignant cells [67]. In a recent report, Pérez-Riesgo et al. have assessed differential gene expression of all human TRP channels by Next Generation Sequencing (NGS) to investigate differences in expression of several genes involved in intracellular Ca2+ transport in CRC [65]. The results obtained, comparing human normal colonic cells versus human colon cancer lines, revealed that only a dozen of the TRPs are expressed in colonic cells, either normal or tumor. The data also indicated that TRPV6 was the unique TRP gene significantly overexpressed in CRC cells, resembling the previously reported enhanced levels of TRPV6 in prostate cancer [66]. Notwithstanding, the functional consequences of changes in expression of TRPV1 and TRPV6 channels in CRC remain to be established.
4. Experimental Models and Therapeutic Opportunities
The implication of TRP channels in pathophysiological mechanisms within the GI tract, and the changes in TRPs expression and activity related with VH, IBD, and CRC have attracted enormous interest in the therapeutic exploitation of these proteins [14,132,133]. Growing evidence points out TRP channels as potential targets for novel analgesics effective in GI pathophysiological conditions including VH, IBS, and IBD, thus, over the past years several corporations started research screening to identify TRP modulators. Multiple approaches can be used to counteract dysfunctional TRP channels, such as blocking TRPs with antagonists, desensitizing the channel with agonists or using drugs that prevent channel trafficking and incorporation in the plasma membrane [134].
As discussed earlier, TRPV1 activity is crucial in colorectal mechanosensation and neurogenic inflammation, representing a captivating target for pharmacological treatment of IBS. Several clinical trials have been implemented with selective TRP channel antagonists, including some very promising drugs that have been patented [134,135]. For instance, the TRPV1 antagonist JYL1421 has proved to be effective to counteract VH caused by various stimuli, colitis and the increase in TRPV1 immunoreactivity in DRG [136]. There have been a large number of studies using animal models, mainly mice, to investigate the implication of TRPV1 in IBDs and VH. For example, the 2,4,6-trinitrobenzenesulfonic acid (TNBS)–induced colitis mice model was used to demonstrate that the upregulation of TRPV1 in ganglia neurons triggered chemical and mechanical VH [136]. Moreover, mice lacking TRPV1 failed to develop post-inflammatory VH following acute colitis induced by dextran sulfate sodium (DSS) [137]. Finally, in rat stress-induced models, visceral pain was decreased administrating TRPV1 antagonists [138].
Theoretically, TRPA1 could be an excellent target for the treatment of GI inflammatory disorders and VH, however, studies on TRPA1 antagonists and their efficacy and safety are very low [139,140]. In mice, intracolonic administration of TRPA1 agonists enhanced the visceromotor response [81]. In TNBS-induced colitis mice models, the overexpression of TRPA1 in colonic DRGs led to an increased visceromotor response to colorectal distention. This effect may be prevented treating mice with a TRPA1 small interfering RNA (siRNA) [141] or TRPA1 antagonists [97]. Moreover, in DSS colitis models pretreated with a TRPA1 antagonist, the TRPA1 agonist Allyl isothiocyanate (AITC) induced VH [52]. Interestingly, the combination of a TRPV1 antagonist (BCTC) with a TRPA1 antagonist (TCS-5861528) in a rat model of colitis resulted in a substantial reduction of VH compared to injection of the drugs separately, suggesting a synergistic effect [97].
TRPM8 plays a primary role in the treatment of VH and GI pain as demonstrated by the analgesic effect of its agonists. Menthol, a well-known TRPM8 activator, was used during decades in traditional medicine to alleviate abdominal spasms and visceral pain [142]. In addition, modern clinical trials showed that the ingestion of peppermint-oil in IBS patients significantly decreases abdominal pain perception [143,144] and is effective in the treatment of functional dyspepsia [145]. Preclinical models suggest that the activation of TRPM8 results in a reduced GI pain perception. Transgenic mice lacking TRPM8 showed loss of innocuous cold sensation, diminished responses to noxious cold temperatures, and a reduced response to cooling compounds. For instance, the administration of a mixture of TRPM8 agonists peppermint and caraway oil reduced the colonic hypersensitivity in TNBS-induced models [146]. It has been reported that in clinical trials, the administration of enteric-coated peppermint oil decreased visceral pain and increase the life quality of IBS patients [143,144]. Moreover, TRPM8 activation by its agonist icilin alleviate inflammation in experimental colitis models and reduced the levels of inflammatory cytokines and mucosal injury, probably due to an inhibition of neuropeptide release [104].
The potential use of two TRPV4 antagonists in the treatment of VH and colitis have been suggested in recent studies. First, the TRPV4 antagonist HC067047 has had results beneficial to attenuate the mechanosensitivity perception of VH in ex vivo experiments [61]. Also, the results obtained with the antagonist RN173 injected systemically and administered locally was suggestively encouraging, suggesting that the blockage of TRPV4 may be helpful in attenuation of IBD [62]. Promising results have also been obtained employing live imaging of rectal biopsies [147]. Balemans and colleagues found that the administration of the agonist GSK1016790A triggered an increased Ca2+ response in submucosal neurons of IBS patients probably due to TRPV4 desensitization [147]. The critical role of TRPV4 in VH was analyzed using various preclinical models. For example, the agonist 4α-phorbol 12,13-didecanoate (4α-PDD) induced significant TRPV4-mediated Ca2+ influx in isolated colonic DRG neurons [57]. On the other hand, VH responses of colonic afferent fibers were increased by the agonist 5,6-EET (an endogenous arachidonic acid metabolite) and considerably reduced by TRPV4 antagonists [58]. Finally, in vivo models demonstrated that the administration of supernatants derived from IBS biopsies induced VH in mice, while, conversely, silencing TRPV4 in mouse primary afferent neurons inhibited the hypersensitivity induced by IBS biopsies supernatants [56].
Altogether, these experimental data indicate the crucial role of TRP channels in mechanosensitivity, chemosensitivity, and pain in GI pathological states. However, it should be considered that targeting TRP channels is a challenging therapeutic approach, given their widespread expression and homeostatic roles outside the GI system [51,148]. Accordingly, even though there are some positive reports, the direct inhibition or activation of TRP channels often leads to severe off-target adverse effects. Unfortunately, the direct drugs targeting of TRP channels present some limitations due to their abundant distribution in the body, although other drawbacks must be considered. For example, the activation of TRPV1 channels have a noticeable hyperthermic effect that limits the clinical usefulness of their agonists. Analogously, TRPA1 and TRPM8 activation often are associated to hypothermic side-effects. Moreover, several TRP agonists and inhibitors are not highly specific and cross-reactivity between different TRP channels have been observed. Finally, the expression of specific GI variant and short isoforms further difficult the targeting of these channels.
In recent years, a very promising alternative to the direct inhibition of TRP channels has emerged. Mounting evidence have proved that ω-3 polyunsaturated fatty acid–derived resolvins (Rv) are potent anti-inflammatory agents that preclude the activation of TRPA1, TRPV1, and TRPV4 channels [149,150]. Resolvins are endogenous lipids produced mainly by eosinophils and neutrophils, effective as anti-inflammatory molecules [151]. So far, RvE1, RvD1, and RvD2 are the most studied for their analgesic properties and mounting evidence indicates that Rv strongly interferes indirectly with TRP channels activity [151]. Thus, Rv selectively inhibited acute pain evoked by administration of TRPV1, TRPV4, and TRPA1 agonists and attenuate inflammatory hypersensitivity to various stimuli [152,153]. Although the mechanism by which Rv inhibit TRP channels is not entirely known, these molecules represent a very interesting and promising alternative to resolve VH and GI inflammatory pain [153].
5. Conclusions
Herein, we briefly reviewed these progresses underscoring the important role of TRPs as cellular sensors and transductors of chemical, thermal, and toxic stimuli within the alimentary canal. Our comprehension of the involvement of TRP channels to gastrointestinal taste, sensation, and nociception has considerably increased in the recent past. Some gastrointestinal disorders, including VH, IBS, IBD, and CRC, are characterized by an increased expression of TRP channels and/or change of its function, proof of a direct implication of TRPs in these pathologies. Several studies have demonstrated that these channels are potentially druggable targets in GI dysfunctions and new promising molecules have been discovered. However, future basic research should be focused on deciphering the relationship between the recently disclosed structure-function data and the signaling pathways related to TRP channels for the development of more selective drugs for the treatment of GI pathologies. Alternatively, a comprehensive understanding of gut microbial metabolites in homeostasis and in diseases may also attenuate GI disorders.
Author Contributions
A.A. and J.R. wrote and reviewed the manuscript.
Funding
This work was partially supported by Starting Grants—Young Researchers 2019 from the University of Trento.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Meerveld, B.G.-V.; Johnson, A.C.; Grundy, D. Gastrointestinal Physiology and Function. Actin Cytoskelet. 2017, 239, 1–16. [Google Scholar]
- Fung, T.C.; Olson, C.A.; Hsiao, E.Y. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 2017, 20, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, Y. Gut Microbiota in Health and Disease. Ann. Nutr. Metab. 2018, 71, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M. Physiological underpinnings of irritable bowel syndrome: Neurohormonal mechanisms. J. Physiol. 2014, 592, 2967–2980. [Google Scholar] [CrossRef]
- Boesmans, W.; Owsianik, G.; Tack, J.; Voets, T.; Vanden Berghe, P. TRP channels in neurogastroenterology: Opportunities for therapeutic intervention. Br. J. Pharmacol. 2011, 162, 18–37. [Google Scholar] [CrossRef]
- Bouron, A.; Kiselyov, K.; Oberwinkler, J. Permeation, regulation and control of expression of TRP channels by trace metal ions. Pflug. Arch. Eur. J. Physiol. 2015, 467, 1143–1164. [Google Scholar] [CrossRef]
- Hellmich, U.A.; Gaudet, R. Structural biology of TRP channels. Handb. Exp. Pharmacol. 2014, 223, 963–990. [Google Scholar]
- Liao, M.; Cao, E.; Julius, D.; Cheng, Y. Structure of the TRPV1 ion channel determined by electron cryo-microscopy. Nature 2013, 504, 107–112. [Google Scholar] [CrossRef]
- Zubcevic, L.; Herzik, M.A.; Chung, B.C.; Liu, Z.; Lander, G.C.; Lee, S.-Y. Cryo-electron microscopy structure of the TRPV2 ion channel. Nat. Struct. Mol. Biol. 2016, 23, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Wu, M.; Zubcevic, L.; Borschel, W.F.; Lander, G.C.; Lee, S.Y. Structure of the cold- and menthol-sensing ion channel TRPM8. Science 2018, 359, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B. TRP channels in disease. Biochim. Biophys. Acta Mol. Basis Dis. 2007, 1772, 805–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilius, B.; Flockerzi, V. Mammalian transient receptor potential (TRP) cation channels. Preface. Handb. Exp. Pharmacol. 2014, 223, v–vi. [Google Scholar]
- Gees, M.; Colsoul, B.; Nilius, B. The role of transient receptor potential cation channels in Ca2+ signaling. Cold Spring Harb. Perspect. Biol. 2010, 2, a003962. [Google Scholar] [CrossRef]
- Bourinet, E.; Altier, C.; Hildebrand, M.E.; Trang, T.; Salter, M.W.; Zamponi, G.W. Calcium-Permeable Ion Channels in Pain Signaling. Physiol. Rev. 2014, 94, 81–140. [Google Scholar] [CrossRef]
- Gkika, D.; Prevarskaya, N. Molecular mechanisms of TRP regulation in tumor growth and metastasis. Biochim. Biophys. Acta BBA Bioenerg. 2009, 1793, 953–958. [Google Scholar] [CrossRef] [Green Version]
- Nilius, B.; Owsianik, G. Transient receptor potential channelopathies. Pflug. Arch. Eur. J. Physiol. 2010, 460, 437–450. [Google Scholar] [CrossRef]
- Kondratskyi, A.; Prevarskaya, N. TRP Channels in Cancer. In TRP Channels as Therapeutic Targets: From Basic Science to Clinical Use; Elsevier: Amsterdam, The Netherlands, 2015; pp. 405–418. [Google Scholar]
- Holzer, P. Transient receptor potential (TRP) channels as drug targets for diseases of the digestive system. Pharmacol. Ther. 2011, 131, 142–170. [Google Scholar] [CrossRef] [Green Version]
- Holzer, P.; Izzo, A.A. The pharmacology of TRP channels. Br. J. Pharmacol. 2014, 171, 2469–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellono, N.W.; Bayrer, J.R.; Leitch, D.B.; Castro, J.; Zhang, C.; O’Donnell, T.A.; Brierley, S.M.; Ingraham, H.A.; Julius, D. Enterochromaffin Cells Are Gut Chemosensors that Couple to Sensory Neural Pathways. Cell 2017, 170, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Faussone-Pellegrini, M.-S.; Taddei, A.; Bizzoco, E.; Lazzeri, M.; Vannucchi, M.G.; Bechi, P. Distribution of the vanilloid (capsaicin) receptor type 1 in the human stomach. Histochem. Cell Biol. 2005, 124, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Szallasi, A.; Cortright, D.N.; Blum, C.A.; Eid, S.R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. Nat. Rev. Drug Discov. 2007, 6, 357–372. [Google Scholar] [CrossRef]
- Nilius, B.; Voets, T. The puzzle of TRPV4 channelopathies. EMBO Rep. 2013, 14, 845. [Google Scholar] [CrossRef]
- Vergnolle, N. TRPV4: New therapeutic target for inflammatory bowel diseases. Biochem. Pharmacol. 2014, 89, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Viana, F. TRPA1 channels: Molecular sentinels of cellular stress and tissue damage. J. Physiol. 2016, 594, 4151–4169. [Google Scholar] [CrossRef]
- Bezençon, C.; le Coutre, J.; Damak, S. Taste-signaling proteins are coexpressed in solitary intestinal epithelial cells. Chem. Senses 2007, 32, 41–49. [Google Scholar] [CrossRef]
- Hantute-Ghesquier, A.; Haustrate, A.; Prevarskaya, N.; Lehen’kyi, V. TRPM family channels in cancer. Pharmaceuticals 2018, 11, 58. [Google Scholar] [CrossRef]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with aδ/c-fibers and colocalization with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jones, S.; Brody, K.; Costa, M.; Brookes, S.J.H. Thermosensitive transient receptor potential channels in vagal afferent neurons of the mouse. Am. J. Physiol. Liver Physiol. 2004, 286, G983–G991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Almaraz, L.; Manenschijn, J.A.; de la Peña, E.; Viana, F. TRPM8. Handb. Exp. Pharmacol. 2014, 222, 547–579. [Google Scholar]
- Holzer, P. TRP Channels in the Digestive System. Curr. Pharm. Biotechnol. 2010, 12, 24–34. [Google Scholar] [CrossRef]
- Montell, C. Physiology, Phylogeny, and Functions of the TRP Superfamily of Cation Channels. Sci. STKE 2003, 2001, re1. [Google Scholar] [CrossRef]
- Montell, C.; Birnbaumer, L.; Flockerzi, V. The TRP channels, a remarkably functional family. Cell 2002, 108, 595–598. [Google Scholar] [CrossRef]
- Ramsey, I.S.; Delling, M.; Clapham, D.E. An introduction to TRP channels. Annu. Rev. Physiol. 2006, 68, 619–647. [Google Scholar] [CrossRef]
- Flockerzi, V. An introduction on TRP channels. Handb. Exp. Pharmacol. 2007, 179, 1–19. [Google Scholar]
- Clapham, D.E.; Runnels, L.W.; Strübing, C. The TRP ion channel family. Nat. Rev. Neurosci. 2001, 2, 387–396. [Google Scholar] [CrossRef]
- Balemans, D.; Boeckxstaens, G.E.; Talavera, K.; Wouters, M.M. Transient receptor potential ion channel function in sensory transduction and cellular signaling cascades underlying visceral hypersensitivity. Am. J. Physiol. Liver Physiol. 2017, 312, G635–G648. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.; Yiangou, Y.; Facer, P.; Walters, J.R.F.; Anand, P.; Ghosh, S. Increased capsaicin receptor TRPV1-expressing sensory fibres in irritable bowel syndrome and their correlation with abdominal pain. Gut 2008, 57, 923–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbar, A.; Yiangou, Y.; Facer, P.; Brydon, W.G.; Walters, J.R.F.; Anand, P.; Ghosh, S. Expression of the TRPV1 receptor differs in quiescent inflammatory bowel disease with or without abdominal pain. Gut 2010, 59, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, I.M.; Christianson, J.A. Ion channels, ion channel receptors, and visceral hypersensitivity in irritable bowel syndrome. Neurogastroenterol. Motil. 2016, 28, 1613–1618. [Google Scholar] [CrossRef] [Green Version]
- Winston, J.; Shenoy, M.; Medley, D.; Naniwadekar, A.; Pasricha, P.J. The Vanilloid Receptor Initiates and Maintains Colonic Hypersensitivity Induced by Neonatal Colon Irritation in Rats. Gastroenterology 2007, 132, 615–627. [Google Scholar] [CrossRef]
- Jones, R.C.W.; Xu, L.; Gebhart, G.F.; Jones, R.C.W. The Mechanosensitivity of Mouse Colon Afferent Fibers and Their Sensitization by Inflammatory Mediators Require Transient Receptor Potential Vanilloid 1 and Acid-Sensing Ion Channel 3. J. Neurosci. 2005, 25, 10981–10989. [Google Scholar] [CrossRef]
- Blackshaw, L.A.; Brierley, S.M.; Hughes, P.A. TRP channels: New targets for visceral pain. Gut 2010, 59, 126–135. [Google Scholar] [CrossRef]
- Wouters, M.M.; Balemans, D.; Van Wanrooy, S.; Dooley, J.; Cibert-Goton, V.; Alpizar, Y.A.; Valdez-Morales, E.E.; Nasser, Y.; Van Veldhoven, P.P.; Vanbrabant, W.; et al. Histamine Receptor H1–Mediated Sensitization of TRPV1 Mediates Visceral Hypersensitivity and Symptoms in Patients with Irritable Bowel Syndrome. Gastroenterology 2016, 150, 875–887. [Google Scholar] [CrossRef]
- Engel, M.A.; Khalil, M.; Mueller-Tribbensee, S.M.; Becker, C.; Neuhuber, W.L.; Neurath, M.F.; Reeh, P.W. The proximodistal aggravation of colitis depends on substance P released from TRPV1-expressing sensory neurons. J. Gastroenterol. 2012, 47, 256–265. [Google Scholar] [CrossRef]
- Holzer, P. The pharmacological challenge to tame the transient receptor potential vanilloid-1 (TRPV1) nocisensor. Br. J. Pharmacol. 2008, 155, 1145–1162. [Google Scholar] [CrossRef] [Green Version]
- Mitrovic, M.; Shahbazian, A.; Bock, E.; Pabst, M.A.; Holzer, P. Chemo-nociceptive signalling from the colon is enhanced by mild colitis and blocked by inhibition of transient receptor potential ankyrin 1 channels. Br. J. Pharmacol. 2010, 160, 1430–1442. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, M.; Jarmuż, A.; Wasilewski, A.; Sałaga, M.; Fichna, J. Role of Transient Receptor Potential Channels in Intestinal Inflammation and Visceral Pain. Inflamm. Bowel Dis. 2015, 21, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Vinuesa, A.G.; Sancho, R.; Garcia-Limones, C.; Behrens, A.; Dijke, P.T.; Calzado, M.A.; Munoz, E. Vanilloid Receptor-1 Regulates Neurogenic Inflammation in Colon and Protects Mice from Colon Cancer. Cancer Res. 2012, 72, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Sung, B.; Prasad, S.; Ravindran, J.; Yadav, V.R.; Aggarwal, B.B. Capsazepine, a TRPV1 antagonist, sensitizes colorectal cancer cells to apoptosis by TRAIL through ROS-JNK-CHOP-mediated upregulation of death receptors. Free. Radic. Biol. Med. 2012, 53, 1977–1987. [Google Scholar] [CrossRef] [PubMed]
- Cenac, N.; Bautzova, T.; Le Faouder, P.; Veldhuis, N.A.; Poole, D.P.; Rolland, C.; Bertrand, J.; Liedtke, W.; Dubourdeau, M.; Bertrand-Michel, J.; et al. Quantification and Potential Functions of Endogenous Agonists of Transient Receptor Potential Channels in Patients with Irritable Bowel Syndrome. Gastroenterology 2015, 149, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Cenac, N.; Altier, C.; Chapman, K.; Liedtke, W.; Zamponi, G.; Vergnolle, N. Transient Receptor Potential Vanilloid-4 Has a Major Role in Visceral Hypersensitivity Symptoms. Gastroenterology 2008, 135, 937–946.e2. [Google Scholar] [CrossRef] [Green Version]
- Brierley, S.M.; Page, A.J.; Hughes, P.A.; Adam, B.; Liebregts, T.; Cooper, N.J.; Holtmann, G.; Liedtke, W.; Blackshaw, L.A. Selective role for TRPV4 ion channels in visceral sensory pathways. Gastroenterology 2008, 134, 2059–2069. [Google Scholar] [CrossRef]
- Poole, D.P.; Amadesi, S.; Veldhuis, N.A.; Abogadie, F.C.; Lieu, T.; Darby, W.; Liedtke, W.; Lew, M.J.; McIntyre, P.; Bunnett, N.W. Protease-activated Receptor 2 (PAR2) Protein and Transient Receptor Potential Vanilloid 4 (TRPV4) Protein Coupling Is Required for Sustained Inflammatory Signaling. J. Biol. Chem. 2013, 288, 5790–5802. [Google Scholar] [CrossRef]
- Sipe, W.E.B.; Brierley, S.M.; Martin, C.M.; Phillis, B.D.; Cruz, F.B.; Grady, E.F.; Liedtke, W.; Cohen, D.M.; Vanner, S.; Blackshaw, L.A.; et al. Transient receptor potential vanilloid 4 mediates protease activated receptor 2-induced sensitization of colonic afferent nerves and visceral hyperalgesia. Am. J. Physiol. Liver Physiol. 2008, 294, G1288–G1298. [Google Scholar] [CrossRef]
- McGuire, C.; Boundouki, G.; Hockley, J.R.F.; Reed, D.; Cibert-Goton, V.; Peiris, M.; Kung, V.; Broad, J.; Aziz, Q.; Chan, C.; et al. Ex vivo study of human visceral nociceptors. Gut 2018, 67, 86–96. [Google Scholar] [CrossRef]
- Mokrowiecka, A.; Cygankiewicz, A.I.; Zakrzewski, P.K.; Krajewska, W.M.; Małecka-Panas, E.; Fichna, J.; Janecka, A.; Storr, M.A. Transient Receptor Potential Vanilloid 4 blockade protects against experimental colitis in mice: A new strategy for inflammatory bowel diseases treatment? Neurogastroenterol. Motil. 2012, 24, e557–e560. [Google Scholar]
- D’Aldebert, E.; Cenac, N.; Rousset, P.; Martin, L.; Rolland, C.; Chapman, K.; Selves, J.; Alric, L.; Vinel, J.; Vergnolle, N. Transient Receptor Potential Vanilloid 4 Activated Inflammatory Signals by Intestinal Epithelial Cells and Colitis in Mice. Gastroenterology 2011, 140, 275–285.e3. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Shikano, M.; Kamiya, T.; Joh, T.; Ugawa, S. The TRPV4 channel is a novel regulator of intracellular Ca2+ in human esophageal epithelial cells. Am. J. Physiol. Liver Physiol. 2011, 301, G138–G147. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Riesgo, E.; Gutiérrez, L.G.; Ubierna, D.; Acedo, A.; Moyer, M.P.; Núñez, L.; Villalobos, C. Transcriptomic Analysis of Calcium Remodeling in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 922. [Google Scholar] [CrossRef]
- Stewart, T.A.; Yapa, K.T.; Monteith, G.R. Altered calcium signaling in cancer cells. Biochim. Biophys. Acta BBA Biomembr. 2015, 1848, 2502–2511. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, L.; Peng, J.-B.; Tou, L.; Takanaga, H.; Adam, R.M.; Hediger, M.A.; Freeman, M.R. Calcium-selective ion channel, CaT1, is apically localized in gastrointestinal tract epithelia and is aberrantly expressed in human malignancies. Lab. Investig. 2002, 82, 1755–1764. [Google Scholar] [CrossRef]
- Hosoya, T.; Matsumoto, K.; Tashima, K.; Nakamura, H.; Fujino, H.; Murayama, T.; Horie, S. TRPM8 has a key role in experimental colitis-induced visceral hyperalgesia in mice. Neurogastroenterol. Motil. 2014, 26, 1112–1121. [Google Scholar] [CrossRef]
- Harrington, A.M.; Hughes, P.A.; Martin, C.M.; Yang, J.; Castro, J.; Isaacs, N.J.; Blackshaw, A.L.; Brierley, S.M.; Blackshaw, L.A. A novel role for TRPM8 in visceral afferent function. Pain 2011, 152, 1459–1468. [Google Scholar] [CrossRef]
- Knowlton, W.M.; Daniels, R.L.; Palkar, R.; McCoy, D.D.; McKemy, D.D. Pharmacological Blockade of TRPM8 Ion Channels Alters Cold and Cold Pain Responses in Mice. PLoS ONE 2011, 6, e25894. [Google Scholar] [CrossRef]
- Kurahara, L.H.; Hiraishi, K.; Sumiyoshi, M.; Doi, M.; Hu, Y.; Aoyagi, K.; Jian, Y.; Inoue, R. Significant contribution of TRPC6 channelmediated Ca2+ influx to the pathogenesis of Crohn’s disease fibrotic stenosis. J. Smooth Muscle Res. 2016, 52, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Burr, A.R.; Davis, G.F.; Birnbaumer, L.; Molkentin, J.D. A TRPC6-dependent pathway for myofibroblast transdifferentiation and wound healing in vivo. Dev. Cell 2012, 23, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Inoue, R.; Jensen, L.J.; Shi, J.; Morita, H.; Nishida, M.; Honda, A.; Ito, Y. Transient Receptor Potential Channels in Cardiovascular Function and Disease. Circ. Res. 2006, 99, 119–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurahara, L.H.; Sumiyoshi, M.; Aoyagi, K.; Hiraishi, K.; Nakajima, K.; Nakagawa, M.; Hu, Y.; Inoue, R. Intestinal myofibroblast TRPC6 channel may contribute to stenotic fibrosis in Crohn’s disease. Inflamm. Bowel Dis. 2015, 21, 496–506. [Google Scholar] [CrossRef]
- Lapointe, T.K.; Altier, C. The role of TRPA1 in visceral inflammation and pain. Channels 2011, 5, 525–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, M.A.; Leffler, A.; Niedermirtl, F.; Babes, A.; Zimmermann, K.; Filipović, M.R.; Izydorczyk, I.; Eberhardt, M.; Kichko, T.I.; Mueller–Tribbensee, S.M.; et al. TRPA1 and Substance P Mediate Colitis in Mice. Gastroenterology 2011, 141, 1346–1358. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guo, C.-H.; Chowdhury, M.A.; Dai, T.-L.; Han, W. TRPA1 in the spinal dorsal horn is involved in post-inflammatory visceral hypersensitivity: In vivo study using TNBS-treated rat model. J. Pain Res. 2016, 9, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Kun, J.; Szitter, I.; Kemeny, A.; Perkecz, A.; Kereskai, L.; Pohoczky, K.; Vincze, Á.; Godi, S.; Szabó, I.; Szolcsányi, J.; et al. Upregulation of the Transient Receptor Potential Ankyrin 1 Ion Channel in the Inflamed Human and Mouse Colon and Its Protective Roles. PLoS ONE 2014, 9, e108164. [Google Scholar] [CrossRef]
- Meseguer, V.; Alpizar, Y.A.; Luis, E.; Tajada, S.; Denlinger, B.; Fajardo, O.; Manenschijn, J.-A.; Fernandez-Pena, C.; Talavera, A.; Kichko, T.; et al. TRPA1 channels mediate acute neurogenic inflammation and pain produced by bacterial endotoxins. Nat. Commun. 2014, 5, 3125. [Google Scholar] [CrossRef] [Green Version]
- Christianson, J.A.; Bielefeldt, K.; Malin, S.A.; Davis, B.M. Neonatal colon insult alters growth factor expression and TRPA1 responses in adult mice. Pain 2010, 151, 540–549. [Google Scholar] [CrossRef] [Green Version]
- Brierley, S.M.; Hughes, P.A.; Page, A.J.; Kwan, K.Y.; Martin, C.M.; O’Donnell, T.A.; Cooper, N.J.; Harrington, A.M.; Adam, B.; Liebregts, T.; et al. The Ion Channel TRPA1 Is Required for Normal Mechanosensation and Is Modulated by Algesic Stimuli. Gastroenterology 2009, 137, 2084–2095. [Google Scholar] [CrossRef]
- Kondo, T.; Obata, K.; Miyoshi, K.; Sakurai, J.; Tanaka, J.; Miwa, H.; Noguchi, K. Transient receptor potential A1 mediates gastric distention-induced visceral pain in rats. Gut 2009, 58, 1342–1352. [Google Scholar] [CrossRef] [PubMed]
- Utsumi, D.; Matsumoto, K.; Tsukahara, T.; Amagase, K.; Tominaga, M.; Kato, S. Transient receptor potential vanilloid 1 and transient receptor potential ankyrin 1 contribute to the progression of colonic inflammation in dextran sulfate sodium-induced colitis in mice: Links to calcitonin gene-related peptide and substance P. J. Pharmacol. Sci. 2018, 136, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Kono, T.; Kaneko, A.; Omiya, Y.; Ohbuchi, K.; Ohno, N.; Yamamoto, M. Epithelial transient receptor potential ankyrin 1 (TRPA1)-dependent adrenomedullin upregulates blood flow in rat small intestine. Am. J. Physiol. Liver Physiol. 2012, 304, G428–G436. [Google Scholar] [CrossRef] [PubMed]
- Kurahara, L.H.; Hiraishi, K.; Hu, Y.; Koga, K.; Onitsuka, M.; Doi, M.; Aoyagi, K.; Takedatsu, H.; Kojima, D.; Fujihara, Y.; et al. Activation of Myofibroblast TRPA1 by Steroids and Pirfenidone Ameliorates Fibrosis in Experimental Crohn’s Disease. Cell. Mol. Gastroenterol. Hepatol. 2018, 5, 299–318. [Google Scholar] [CrossRef]
- Christianson, J.A.; Bielefeldt, K.; Altier, C.; Cenac, N.; Davis, B.M.; Gebhart, G.F.; High, K.W.; Kollarik, M.; Randich, A.; Undem, B.; et al. Development, plasticity and modulation of visceral afferents. Brain Res. Rev. 2009, 60, 171–186. [Google Scholar] [CrossRef] [Green Version]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Lovell, R.M.; Ford, A.C. Global Prevalence of and Risk Factors for Irritable Bowel Syndrome: A Meta-analysis. Clin. Gastroenterol. Hepatol. 2012, 10, 712–721. [Google Scholar] [CrossRef]
- Farmer, A.D.; Ruffle, J.K. Irritable bowel syndrome. Hamdan Med. J. 2019, 47, 350–353. [Google Scholar] [CrossRef] [Green Version]
- Chey, W.D.; Kurlander, J.; Eswaran, S. Irritable bowel syndrome: A clinical review. JAMA 2015, 313, 949–958. [Google Scholar] [CrossRef]
- Lacy, B.E.; Mearin, F.; Chang, L.; Chey, W.D.; Lembo, A.J.; Simren, M.; Spiller, R. Bowel disorders. Gastroenterology 2016, 150, 1393–1407. [Google Scholar] [CrossRef]
- Farmer, A.D.; Aziz, Q. Gut pain & visceral hypersensitivity. Br. J. Pain 2013, 7, 39–47. [Google Scholar] [PubMed] [Green Version]
- De Winter, B.Y.; Deiteren, A.; De Man, J.G.; De Winter, B. Novel nervous system mechanisms in visceral pain. Neurogastroenterol. Motil. 2016, 28, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P.; Hassan, A.M.; Jain, P.; Reichmann, F.; Farzi, A. Neuroimmune pharmacological approaches. Curr. Opin. Pharmacol. 2015, 25, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Nociceptive TRP Channels: Sensory Detectors and Transducers in Multiple Pain Pathologies. Pharmaceuticals 2016, 9, 72. [Google Scholar] [CrossRef]
- McMahon, S.B.; La Russa, F.; Bennett, D.L.H. Crosstalk between the nociceptive and immune systems in host defence and disease. Nat. Rev. Neurosci. 2015, 16, 389–402. [Google Scholar] [CrossRef]
- Vermeulen, W.; De Man, J.G.; De Schepper, H.U.; Bult, H.; Moreels, T.G.; Pelckmans, P.A.; De Winter, B.Y. Role of TRPV1 and TRPA1 in visceral hypersensitivity to colorectal distension during experimental colitis in rats. Eur. J. Pharmacol. 2013, 698, 404–412. [Google Scholar] [CrossRef]
- Miyoshi, J.; Chang, E.B. The gut microbiota and inflammatory bowel diseases. Transl. Res. 2017, 179, 38–48. [Google Scholar] [CrossRef]
- Oligschlaeger, Y.; Yadati, T.; Houben, T.; Oliván, C.M.C.; Shiri-Sverdlov, R.; Oliván, C.C. Inflammatory Bowel Disease: A Stressed “Gut/Feeling”. Cells 2019, 8, 659. [Google Scholar] [CrossRef]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Nathan, C.; Ding, A. Nonresolving Inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Rubin, D.C.; Shaker, A.; Levin, M.S. Chronic intestinal inflammation: Inflammatory bowel disease and colitis-associated colon cancer. Front. Immunol. 2012, 3, 107. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Julius, D. The vanilloid receptor: A molecular gateway to the pain pathway. Annu. Rev. Neurosci. 2001, 24, 487–517. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R.; Hyun, E.; Zhao, L.; Lapointe, T.K.; Chapman, K.; Hirota, C.L.; Ghosh, S.; McKemy, D.D.; Vergnolle, N.; Beck, P.L.; et al. TRPM8 activation attenuates inflammatory responses in mouse models of colitis. Proc. Natl. Acad. Sci. USA 2013, 110, 7476–7481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, A.; Correale, C.; Szabo, H.; Montorsi, M. Intestinal Fibrosis in Crohns Disease: Medical Treatment or Surgery? Curr. Drug Targets 2010, 11, 242–248. [Google Scholar] [CrossRef]
- Inoue, R.; Kurahara, L.H.; Hiraishi, K. TRP channels in cardiac and intestinal fibrosis. Semin. Cell Dev. Biol. 2019, 94, 40–49. [Google Scholar] [CrossRef]
- Li, C.; Kuemmerle, J.F. Mechanisms that mediate the development of fibrosis in patients with Crohn’s disease. Inflamm. Bowel Dis. 2014, 20, 1250–1258. [Google Scholar] [CrossRef]
- Latella, G.; Di Gregorio, J.; Flati, V.; Rieder, F.; Lawrance, I.C. Mechanisms of initiation and progression of intestinal fibrosis in IBD. Scand. J. Gastroenterol. 2014, 50, 53–65. [Google Scholar] [CrossRef]
- Rosenbloom, J.; Macarak, E.; Piera-Velazquez, S.; Jimenez, S.A. Human fibrotic diseases: Current challenges in fibrosis research. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2017; Volume 1627, pp. 1–23. [Google Scholar]
- Meier, J.K.H.; Scharl, M.; Miller, S.N.; Brenmoehl, J.; Hausmann, M.; Kellermeier, S.; Schölmerich, J.; Rogler, G. Specific differences in migratory function of myofibroblasts isolated from Crohn’s disease fistulae and strictures. Inflamm. Bowel Dis. 2011, 17, 202–212. [Google Scholar] [CrossRef]
- Valentich, J.D.; Popov, V.; Saada, J.I.; Powell, D.W. Phenotypic characterization of an intestinal subepithelial myofibroblast cell line. Am. J. Physiol. Physiol. 2017, 272, C1513–C1524. [Google Scholar] [CrossRef]
- Yun, S.-M.; Kim, S.-H.; Kim, E.-H. The Molecular Mechanism of Transforming Growth Factor-β Signaling for Intestinal Fibrosis: A Mini-Review. Front. Pharmacol. 2019, 10, 162. [Google Scholar] [CrossRef] [PubMed]
- Stempien-Otero, A.; Kim, D.-H.; Davis, J. Molecular networks underlying myofibroblast fate and fibrosis. J. Mol. Cell. Cardiol. 2016, 97, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Prunotto, M.; Desmoulière, A.; Varga, J.; De Wever, O.; Mareel, M.; Gabbiani, G. Recent developments in myofibroblast biology: Paradigms for connective tissue remodeling. Am. J. Pathol. 2012, 180, 1340–1355. [Google Scholar] [CrossRef] [PubMed]
- Babyatsky, M.W.; Rossiter, G.; Podolsky, D.K. Expression of transforming growth factors α and β in colonic mucosa in inflammatory bowel disease. Gastroenterology 1996, 110, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Janssen, L.J.; Mukherjee, S.; Ask, K. Calcium Homeostasis and Ionic Mechanisms in Pulmonary Fibroblasts. Am. J. Respir. Cell Mol. Biol. 2015, 53, 135–148. [Google Scholar] [CrossRef]
- Giorgi, C.; Danese, A.; Missiroli, S.; Patergnani, S.; Pinton, P. Calcium Dynamics as a Machine for Decoding Signals. Trends Cell Biol. 2018, 28, 258–273. [Google Scholar] [CrossRef]
- Saliba, Y.; Karam, R.; Smayra, V.; Aftimos, G.; Abramowitz, J.; Birnbaumer, L.; Farès, N. Evidence of a role for fibroblast transient receptor potential canonical 3 Ca2+ channel in renal fibrosis. J. Am. Soc. Nephrol. 2015, 26, 1855–1876. [Google Scholar] [CrossRef]
- Seo, K.; Rainer, P.P.; Hahn, V.S.; Lee, D.-I.; Jo, S.-H.; Andersen, A.; Liu, T.; Xu, X.; Willette, R.N.; Lepore, J.J.; et al. Combined TRPC3 and TRPC6 blockade by selective small-molecule or genetic deletion inhibits pathological cardiac hypertrophy. Proc. Natl. Acad. Sci. USA 2014, 111, 1551–1556. [Google Scholar] [CrossRef] [Green Version]
- Saliba, Y.; Jebara, V.; Hajal, J.; Maroun, R.; Chacar, S.; Smayra, V.; Abramowitz, J.; Birnbaumer, L.; Fares, N. Transient Receptor Potential Canonical 3 and Nuclear Factor of Activated T Cells C3 Signaling Pathway Critically Regulates Myocardial Fibrosis. Antioxid. Redox Signal. 2019, 30, 1851–1879. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef]
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Inamura, K. Colorectal Cancers: An Update on Their Molecular Pathology. Cancers 2018, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Santoni, G.; Farfariello, V. TRP channels and cancer: New targets for diagnosis and chemotherapy. Endocr. Metab. Immune Disord. Drug Targets 2011, 11, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Shapovalov, G.; Ritaine, A.; Skryma, R.; Prevarskaya, N. Role of TRP ion channels in cancer and tumorigenesis. Semin. Immunopathol. 2016, 38, 357–369. [Google Scholar] [CrossRef]
- Gkika, D.; Prevarskaya, N. TRP channels in prostate cancer: The good, the bad and the ugly? Asian J. Androl. 2011, 13, 673–676. [Google Scholar] [CrossRef]
- Liberati, S.; Morelli, M.B.; Nabissi, M.; Santoni, M.; Santoni, G. Oncogenic and anti-oncogenic effects of transient receptor potential channels. Curr. Top. Med. Chem. 2013, 13, 344–366. [Google Scholar] [CrossRef]
- Shapovalov, G.; Lehen’Kyi, V.; Skryma, R.; Prevarskaya, N. TRP channels in cell survival and cell death in normal and transformed cells. Cell Calcium 2011, 50, 295–302. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Pla, A.F.; Gkika, D. Emerging role of TRP channels in cell migration: From tumor vascularization to metastasis. Front. Physiol. 2013, 4, 311. [Google Scholar]
- Moran, M.M.; McAlexander, M.A.; Bíró, T.; Szallasi, A. Transient receptor potential channels as therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 601–620. [Google Scholar] [CrossRef]
- Kaneko, Y.; Szallasi, A. Transient receptor potential (TRP) channels: A clinical perspective. Br. J. Pharmacol. 2014, 171, 2474–2507. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A. Transient receptor potential channels in pain and inflammation: Therapeutic opportunities. Pain Pract. 2010, 10, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Hong, S.; Cui, M.; Sharma, P.K.; Lee, J.; Choi, S. Transient receptor potential vanilloid type 1 antagonists: A patent review (2011–2014). Expert Opin. Ther. Pat. 2015, 25, 291–318. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.; Nordstrom, E.; Mannem, A.; Smith, C.; Banerjee, B.; Sengupta, J.N. The role of transient receptor potential vanilloid 1 in mechanical and chemical visceral hyperalgesia following experimental colitis. Neuroscience 2007, 148, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Lapointe, T.K.; Basso, L.; Iftinca, M.C.; Flynn, R.; Chapman, K.; Dietrich, G.; Vergnolle, N.; Altier, C. TRPV1 sensitization mediates postinflammatory visceral pain following acute colitis. Am. J. Physiol. Liver Physiol. 2015, 309, G87–G99. [Google Scholar] [CrossRef] [Green Version]
- Wijngaard, R.M.V.D.; Klooker, T.K.; Welting, O.; Stanisor, O.I.; Wouters, M.M.; Van Der Coelen, D.; Bulmer, D.C.; Peeters, P.J.; Aerssens, J.; De Hoogt, R.; et al. Essential role for TRPV1 in stress-induced (mast cell-dependent) colonic hypersensitivity in maternally separated rats. Neurogastroenterol. Motil. 2009, 21, 1107-e94. [Google Scholar] [CrossRef]
- Preti, D.; Saponaro, G.; Szallasi, A. Transient receptor potential ankyrin 1 (TRPA1) antagonists. Pharm. Pat. Anal. 2015, 4, 75–94. [Google Scholar] [CrossRef]
- Chen, J.; Hackos, D.H. TRPA1 as a drug target--promise and challenges. Naunyn Schmiedebergs Arch. Pharmacol. 2015, 388, 451–463. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Zuo, X.; Zhen, Y.; Yu, Y.; Gao, L. Transient receptor potential ankyrin-1 participates in visceral hyperalgesia following experimental colitis. Neurosci. Lett. 2008, 440, 237–241. [Google Scholar] [CrossRef]
- Hicks, G.A. TRP channels as therapeutic targets: Hot property, or time to cool down? Neurogastroenterol. Motil. 2006, 18, 590–594. [Google Scholar] [CrossRef]
- Cappello, G.; Spezzaferro, M.; Grossi, L.; Manzoli, L.; Marzio, L. Peppermint oil (Mintoil®) in the treatment of irritable bowel syndrome: A prospective double blind placebo-controlled randomized trial. Dig. Liver Dis. 2007, 39, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Merat, S.; Khalili, S.; Mostajabi, P.; Ghorbani, A.; Ansari, R.; Malekzadeh, R. The effect of enteric-coated, delayed-release peppermint oil on irritable bowel syndrome. Dig. Dis. Sci. 2010, 55, 1385–1390. [Google Scholar] [CrossRef] [PubMed]
- Rösch, W.; Liebregts, T.; Gundermann, K.-J.; Vinson, B.; Holtmann, G. Phytotherapy for functional dyspepsia: A review of the clinical evidence for the herbal preparation STW 5. Phytomedicine 2006, 13, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Adam, B.; Liebregts, T.; Best, J.; Bechmann, L.; Lackner, C.; Neumann, J.; Koehler, S.; Holtmann, G. A combination of peppermint oil and caraway oil attenuates the post-inflammatory visceral hyperalgesia in a rat model. Scand. J. Gastroenterol. 2006, 41, 155–160. [Google Scholar] [CrossRef]
- Balemans, D.; Aguilera-Lizarraga, J.; Florens, M.V.; Jain, P.; Denadai-Souza, A.; Viola, M.F.; Aguiar Alpizar, Y.; Van Der Merwe, S.; Vanden Berghe, P.; Talavera, K.; et al. Histamine-mediated potentiation of TRPA1 and TRPV4 signaling in submucosal neurons in IBS patients. Am. J. Physiol. Liver Physiol. 2019, 316, G338–G349. [Google Scholar]
- Patapoutian, A.; Tate, S.; Woolf, C.J. Transient receptor potential channels: Targeting pain at the source. Nat. Rev. Drug Discov. 2009, 8, 55–68. [Google Scholar] [CrossRef]
- Ji, R.-R.; Xu, Z.-Z.; Strichartz, G.; Serhan, C.N. Emerging roles of resolvins in the resolution of inflammation and pain. Trends Neurosci. 2011, 34, 599–609. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.-Z.; Zhang, L.; Liu, T.; Park, J.Y.; Berta, T.; Yang, R.; Serhan, C.N.; Ji, R.-R. Resolvins RvE1 and RvD1 attenuate inflammatory pain via central and peripheral actions. Nat. Med. 2010, 16, 592–597. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Bang, S.; Yoo, S.; Yang, T.; Cho, H.; Kim, Y.; Hwang, S. Resolvin D1 attenuates activation of sensory transient receptor potential channels leading to multiple anti-nociception. Br. J. Pharmacol. 2010, 161, 707–720. [Google Scholar] [CrossRef] [Green Version]
- Park, C.-K.; Xu, Z.-Z.; Liu, T.; Lü, N.; Serhan, C.N.; Ji, R.-R. Resolvin D2 is a potent endogenous inhibitor for transient receptor potential subtype V1/A1, inflammatory pain, and spinal cord synaptic plasticity in mice: Distinct roles of resolvin D1, D2, and E1. J. Neurosci. 2011, 31, 18433–18438. [Google Scholar] [CrossRef] [PubMed]
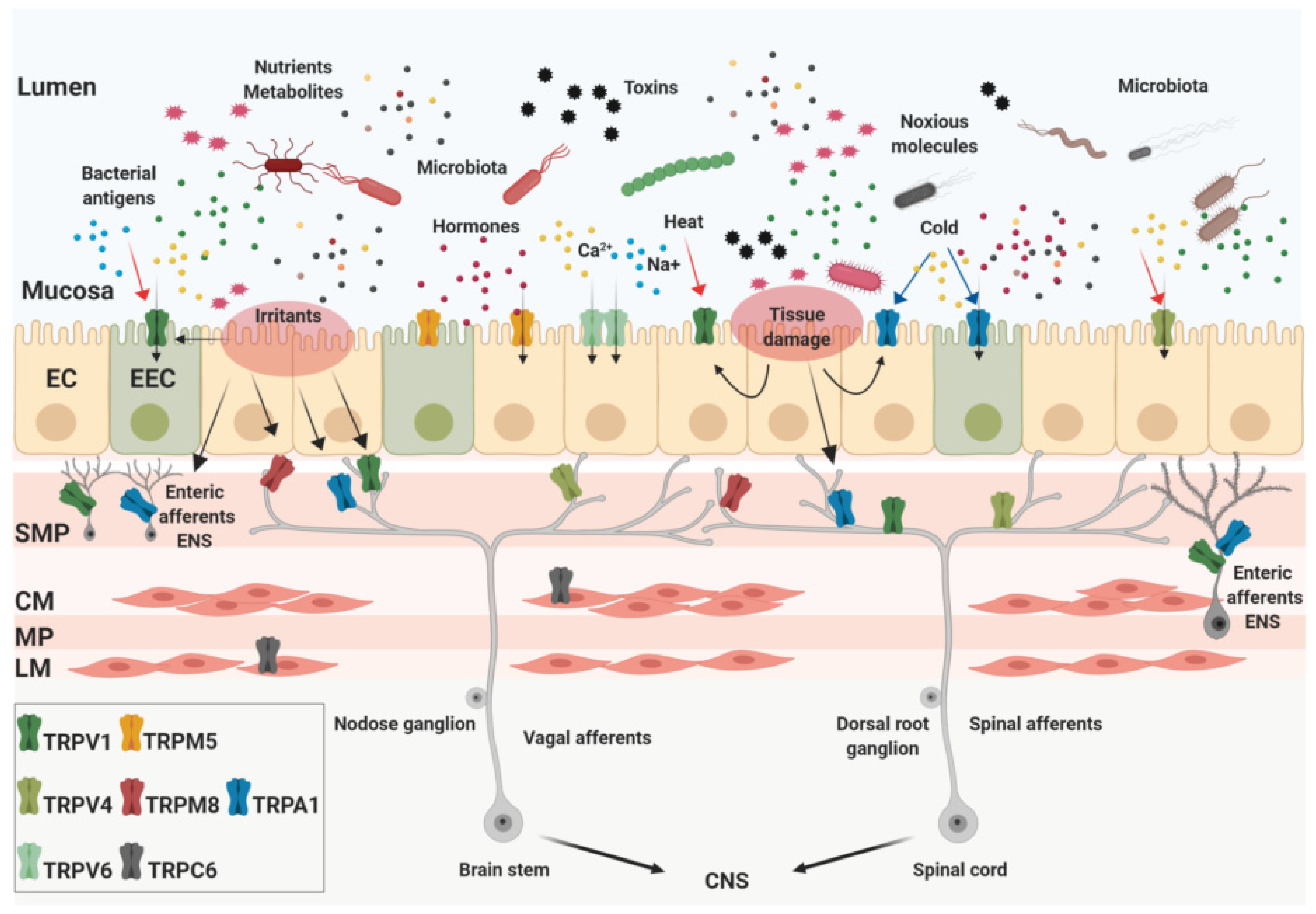
Figure 1.
Expression of the transient receptor potential (TRP) channels in the gut. The intestinal lumen is densely populated by nutrients, gut microbial metabolites, chemicals, and toxins that can be present in the food ingested, released by the intestinal tissue or produced by bacteria. The gastrointestinal (GI) tract mucosa is mainly composed by epithelial cells (enterocytes, EC) and enteroendocrine cells (for instance enterochromaffin cells, EEC) whose apical membranes express several TRP channels. They act as sensors of specific stimuli such as temperature, pH, osmolality, mechanical stress, and recognize bioactive molecules. In addition to the mucosa, TRPs are located in almost all layers of the intestinal wall. Some of them are expressed by afferent neurons in the myenteric (MP) and submucosal plexus (SMP) as part of the intrinsic enteric nervous system (ENS). Two layers of muscle tissue, the circular (CM) and the longitudinal (LM) also express TRP channels. Finally, TRP channels expressed by extrinsic afferent nerves are implicated in the critical mechanism of visceral perception, transporting signals from the gut to the central nervous system (CNS). Created with BioRender.com.
Figure 1.
Expression of the transient receptor potential (TRP) channels in the gut. The intestinal lumen is densely populated by nutrients, gut microbial metabolites, chemicals, and toxins that can be present in the food ingested, released by the intestinal tissue or produced by bacteria. The gastrointestinal (GI) tract mucosa is mainly composed by epithelial cells (enterocytes, EC) and enteroendocrine cells (for instance enterochromaffin cells, EEC) whose apical membranes express several TRP channels. They act as sensors of specific stimuli such as temperature, pH, osmolality, mechanical stress, and recognize bioactive molecules. In addition to the mucosa, TRPs are located in almost all layers of the intestinal wall. Some of them are expressed by afferent neurons in the myenteric (MP) and submucosal plexus (SMP) as part of the intrinsic enteric nervous system (ENS). Two layers of muscle tissue, the circular (CM) and the longitudinal (LM) also express TRP channels. Finally, TRP channels expressed by extrinsic afferent nerves are implicated in the critical mechanism of visceral perception, transporting signals from the gut to the central nervous system (CNS). Created with BioRender.com.

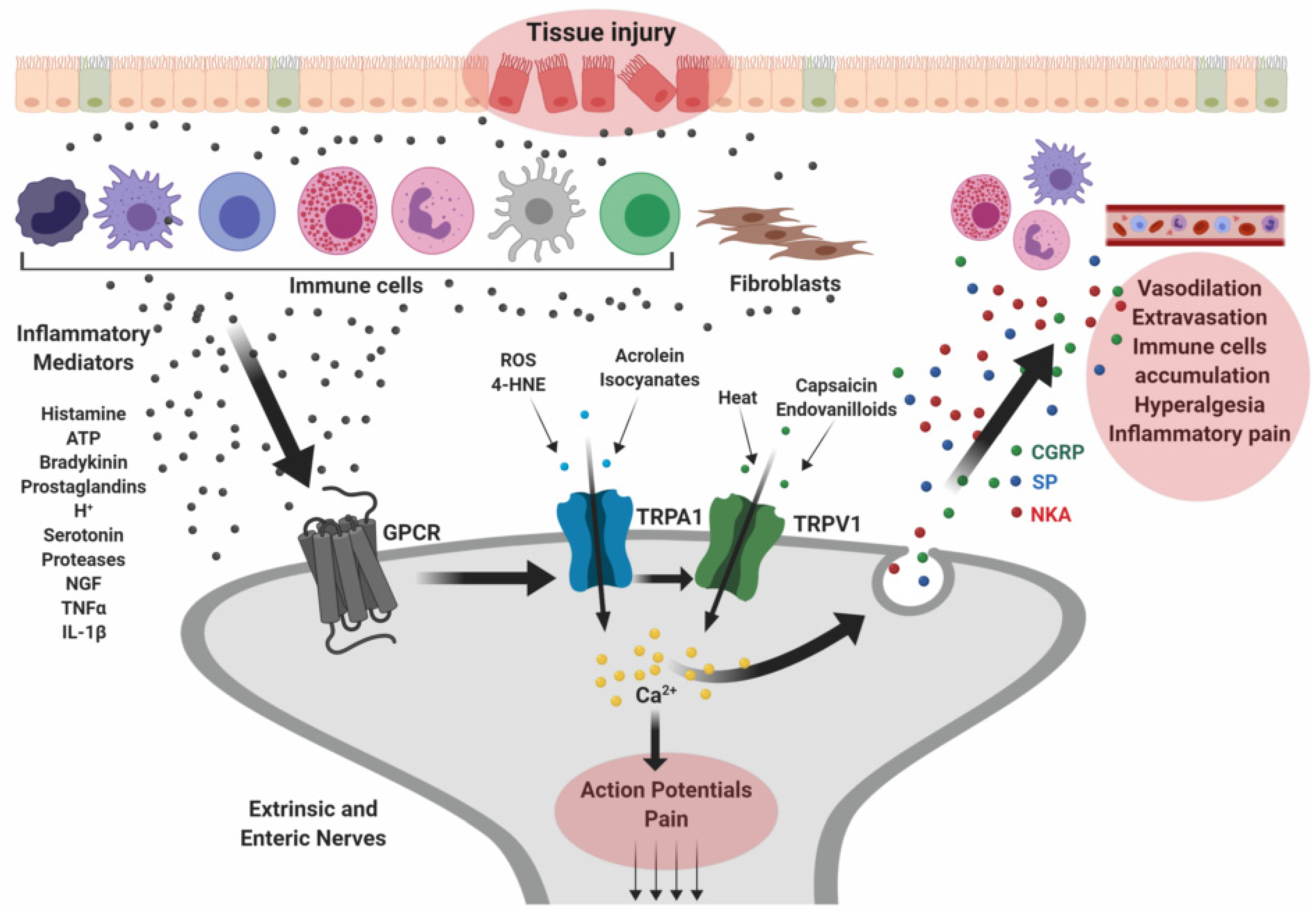
Figure 2.
Implications of TRP in the pathophysiology of inflammatory bowel diseases. Tissue damage occurring along the GI mucosa may cause release of inflammatory mediators that activate principally immune cells and fibroblasts surrounding the injured tissue. These cells release more inflammatory molecules, including ATP, histamine, tumor necrosis factor α (TNF-α), interleukin-1β (IL-1β), nerve growth factor (NGF), and proteases, which bind specific G protein-coupled receptors (GPCRs) expressed in primary afferent sensory neurons. The consequent activation of TRP channels trigger the release of neuropeptides such as substance P (SP), calcitonin gene-related peptide (CGRP), and neurokinin A (NKA) which promote neutrophil accumulation, vasodilation, pain and other inflammatory responses. Created with BioRender.com.
Figure 2.
Implications of TRP in the pathophysiology of inflammatory bowel diseases. Tissue damage occurring along the GI mucosa may cause release of inflammatory mediators that activate principally immune cells and fibroblasts surrounding the injured tissue. These cells release more inflammatory molecules, including ATP, histamine, tumor necrosis factor α (TNF-α), interleukin-1β (IL-1β), nerve growth factor (NGF), and proteases, which bind specific G protein-coupled receptors (GPCRs) expressed in primary afferent sensory neurons. The consequent activation of TRP channels trigger the release of neuropeptides such as substance P (SP), calcitonin gene-related peptide (CGRP), and neurokinin A (NKA) which promote neutrophil accumulation, vasodilation, pain and other inflammatory responses. Created with BioRender.com.


{kind=link}
{kind=link}
Table 1.
TRP channels in the alimentary canal.
| Ligands | Physical Stimuli | Endogenous Stimuli | GI Tract Expression | GI Related Disorders | |
|---|---|---|---|---|---|
| TRPV1 | Capsaicin (red pepper) Piperine (black pepper) Gingerol (ginger) Resiniferatoxin (Euphorbia poissonii) | T ≥ 43 °C Voltage Distension pH | Cannabinoids Anandamide Eicosanoids Acid Bradykinin Serotonin Histamine Proteases | Sensory neurons Enteric neurons Epithelial cells Enteroendocrine cells [25,26] | IBS VH [42,43,44,45,46,47,48,49] IBD [44,50,51,52] CRC [53,54,55] |
| TRPV4 | 4-α Phorbol (C. tiglium) Bisandrographolide A (Andrographis paniculata) | T ≥ 25 °C Mechanical Osmolarity Distension pH | Anandamide Eicosanoids Bradykinin Citrate Arachidonic acids Histamine Proteases | Sensory neurons Epithelial cells [27,28] | IBS VH [42,56,57,58,59,60,61] IBD [57,62,63,64] |
| TRPV6 | Voltage | Epithelial cells [65,66] | CRC [65,66,67] | ||
| TRPM5 | Steviol glycosides (Stevia rebaudiana) Rutamarin (Ruta graveolens) | Voltage pH | Intracellular Calcium Arachidonic acid Phospholipase C Sugars Acid | Epithelial cells Enteroendocrine cells Taste receptor cells [30,31] | |
| TRPM8 | Menthol (mint) Linalool (laurels, cinnamon, rosewood) Geraniol (geranium, rose oil) Eucalyptol (Eucalyptus) | T ≤ 25 °C Voltage | Cannabinoids Anandamide Lysophospholipids Polyunsaturated fatty acids Bradykinin Prostaglandins Serotonin | Sensory neurons [32,33,34,35] | IBS VH [42,68,69,70] |
| TRPC6 | Hyperforin and Adhyperforin (Hypericum perforatum) | Mechanical | Arachidonic acid Lysophospholipids Eicosanoids | Smooth muscle cells [71] | IF [71,72,73,74] |
| TRPA1 | Allicin (garlic) Carvacrol (oregano) Cinnamaldehyde (cinnamon) Diallyl disulfide (garlic) Gingerol (ginger) Ally isothiocyanate (mustard horseradish, wasabi) | T ≤ 18 °C Mechanical Distension pH | Cannabinoids Bradykinin Nicotine Prostaglandins Histamine Proteases ROS 4-HNE | Sensory neurons Enteric neurons Epithelial cells Enteroendocrine cells [21] | IBS VH [21,75,76,77,78,79,80,81] IBD [78,81,82,83] IF [84,85] |
Abbreviations: IBS—Irritable Bowel Syndrome; VH—Visceral Hypersensitivity; IBD—Inflammatory Bowel Diseases (Crohn’s disease and ulcerative colitis); IF—Intestinal Fibrosis; CR—Colorectal Cancer; ROS—Reactive oxygen species; 4-HNE—4-Hydroxynonenal.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alaimo, A.; Rubert, J. The Pivotal Role of TRP Channels in Homeostasis and Diseases throughout the Gastrointestinal Tract. Int. J. Mol. Sci. 2019, 20, 5277. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215277
AMA Style
Alaimo A, Rubert J. The Pivotal Role of TRP Channels in Homeostasis and Diseases throughout the Gastrointestinal Tract. International Journal of Molecular Sciences. 2019; 20(21):5277. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215277
Chicago/Turabian StyleAlaimo, Alessandro, and Josep Rubert. 2019. "The Pivotal Role of TRP Channels in Homeostasis and Diseases throughout the Gastrointestinal Tract" International Journal of Molecular Sciences 20, no. 21: 5277. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215277
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.