Protein Abundance of Clinically Relevant Drug Transporters in The Human Kidneys

 ,
, 
Abstract
:1. Introduction
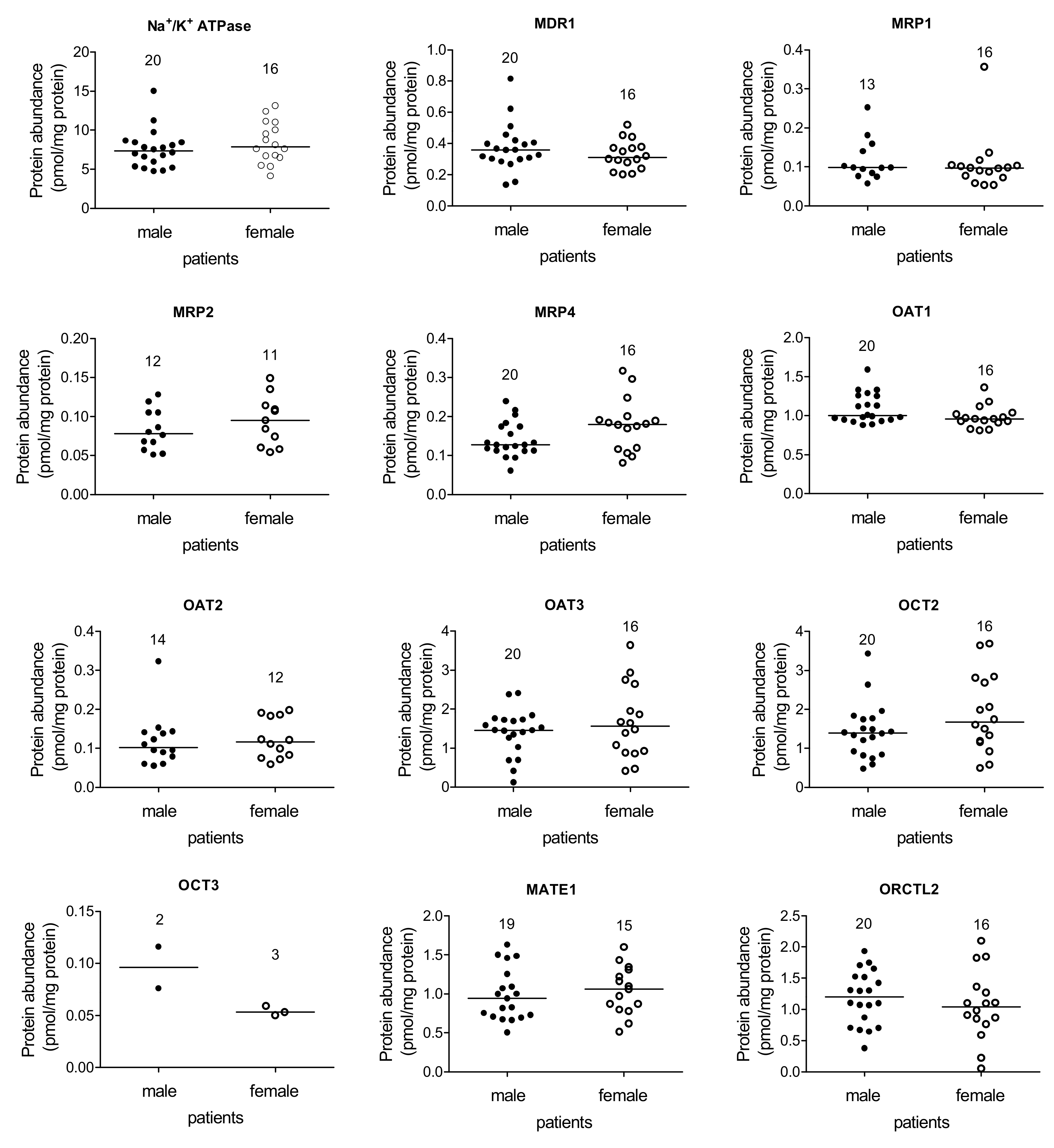
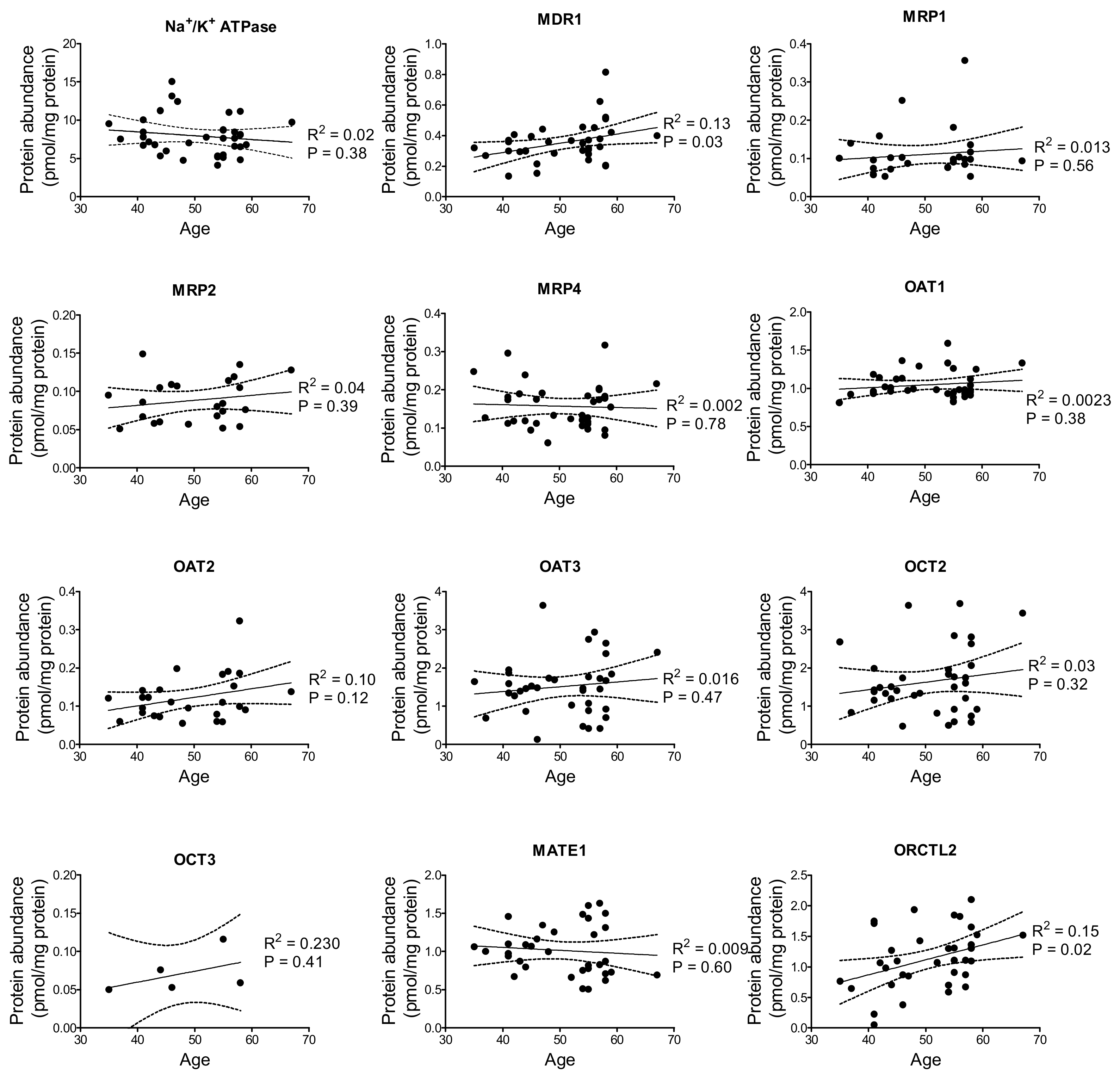
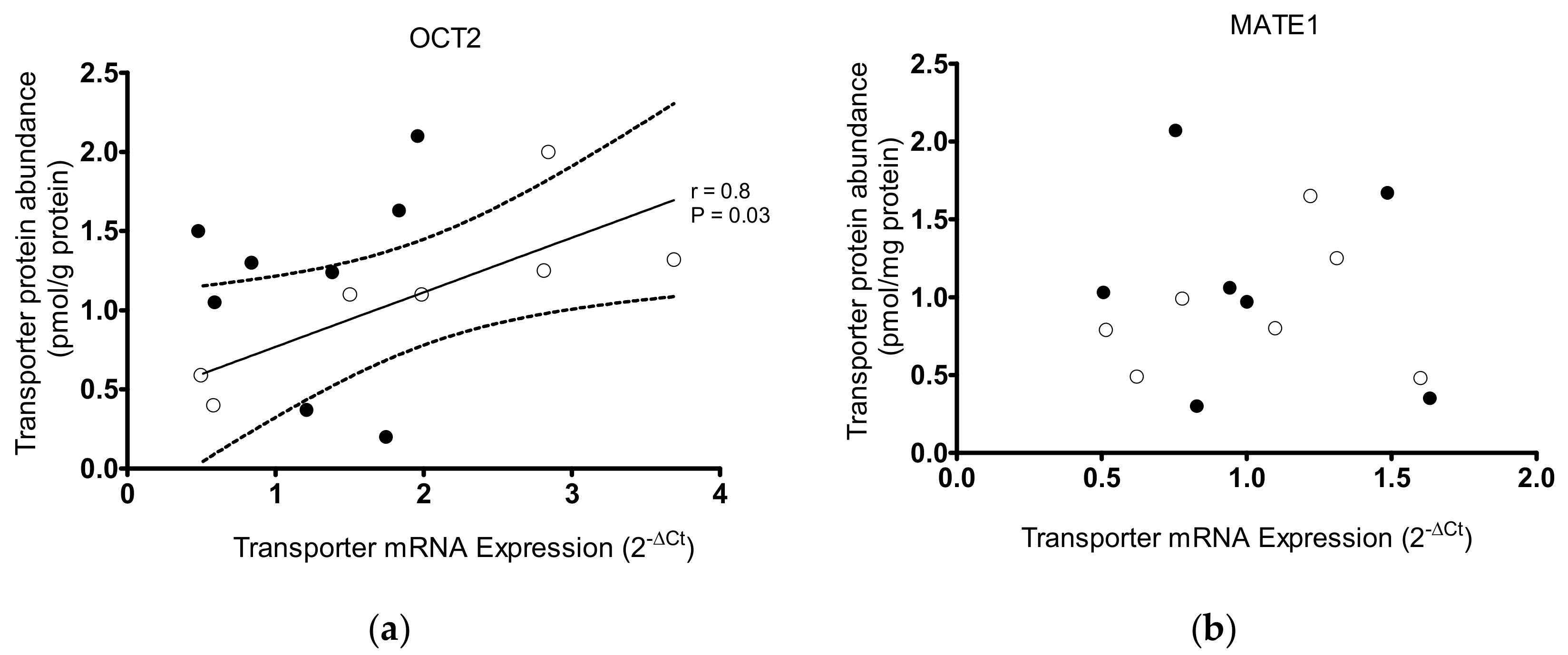
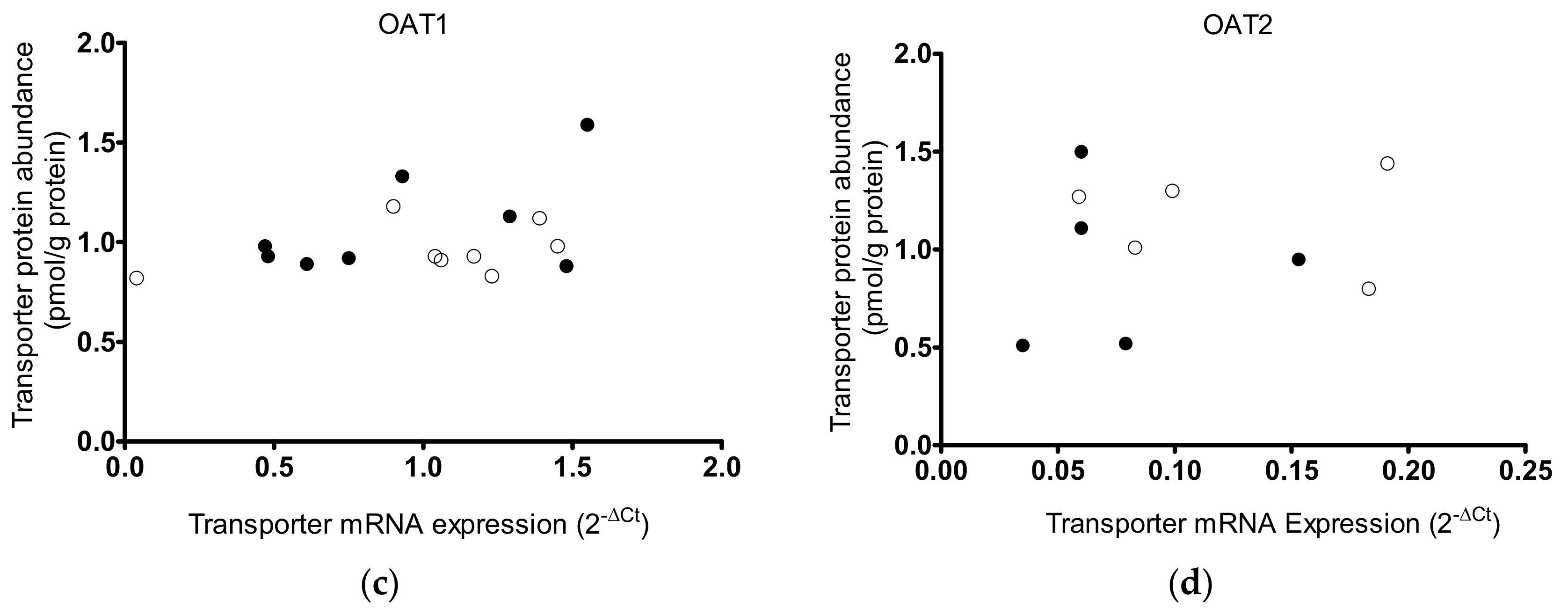
2. Results
- Na+/K+ ATPase correlated directly with MRP2/MRP4/OAT2/OCT2 abundance;
- MDR1 with MRP2/OAT2/OAT3/OCT2;
- MRP2 with Na+/K+ ATPase/MDR1/MRP4/OAT3/OCT2;
- MRP4 with Na+/K+ ATPase/MRP2/OAT3/OCT2;
- OAT2 with Na+/K+ ATPase/MDR1/OAT3/OCT2/MATE1;
- OAT3 with MDR1/MRP2/MRP4/OAT2/OCT2/ORCTL2/MATE1;
- OCT2 with Na+/K+ ATPase/MDR1/MRP2/MRP4/OAT2/OAT3/ORCTL2/MATE1;
- OCT3 with MATE1 (inversely correlated);
- ORCTL2 with OAT3/OCT2;
- MATE1 with OAT2/OAT3/OCT2/OCT3
3. Discussion
4. Materials and Methods
4.1. Human Kidneys
4.2. Protein Quantification by LC-MS/MS
4.3. Real-Time PCR Analysis
4.4. Immunochemistry
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gattineni, J.; Baum, M. Developmental changes in renal tubular transport-an overview. Pediatr. Nephrol. 2015, 30, 2085–2098. [Google Scholar] [CrossRef] [PubMed]
- McDonough, A.A. Mechanisms of proximal tubule sodium transport regulation that link extracellular fluid volume and blood pressure. Am. J. Physiol. Integr. Comp. Physiol. 2010, 298, R851–R861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Wijst, J.; Belge, H.; Bindels, R.J.M.; Devuyst, O. Learning physiology from inherited kidney disorders. Physiol. Rev. 2019, 99, 1575–1653. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.K.; Wu, W.; Bush, K.T.; Hoenig, M.P.; Blantz, R.C.; Bhatnagar, V. Handling of drugs, metabolites, and uremic toxins by kidney proximal tubule drug transporters. Clin. J. Am. Soc. Nephrol. 2015, 10, 2039–2049. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, K.M.; Stocker, S.L.; Wittwer, M.B.; Xu, L.; Giacomini, K.M. Renal transporters in drug development. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 503–529. [Google Scholar] [CrossRef] [PubMed]
- Sabolić, I.; Asif, A.R.; Budach, W.E.; Wanke, C.; Bahn, A.; Burckhardt, G. Gender differences in kidney function. Pflugers Arch. 2007, 455, 397–429. [Google Scholar] [CrossRef]
- Ljubojevic, M.; Balen, D.; Breljak, D.; Kušan, M.; Anzai, N.; Bahn, A.; Burckhardt, G.; Sabolić, I. Renal expression of organic anion transporter OAT2 in rats and mice is regulated by sex hormones. Am. J. Physiol. Renal. Physiol. 2007, 292, F361–F372. [Google Scholar] [CrossRef]
- Buist, S.C.N.; Klaassen, C.D. Rat and mouse differences in gender-predominant expression of organic anion transporter (Oat1-3; Slc22a6-8) mRNA levels. Drug. Metab. Dispos. 2004, 32, 620–625. [Google Scholar] [CrossRef]
- Koepsell, H. Polyspecific organic cation transporters and their biomedical relevance in kidney. Curr. Opin. Nephrol. Hypertens. 2013, 22, 533–538. [Google Scholar] [CrossRef]
- Reece, M.; Prawitt, D.; Landers, J.; Kast, C.; Gros, P.; Housman, D.; Zabel, B.U.; Pelletier, J. Functional characterization of ORCTL2 - an organic cation transporter expressed in the renal proximal tubules. FEBS Lett. 1998, 433, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Launay-Vacher, V.; Izzedine, H.; Karie, S.; Hulot, J.S.; Baumelou, A.; Deray, G. Renal tubular drug transporters. Nephron. Physiol. 2006, 103, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.H.; Dantzler, W.H. Molecular and cellular physiology of renal organic cation and anion transport. Physiol. Rev. 2004, 84, 987–1049. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, M.; Sugawara, I.; Hamada, H.; Tsuruo, T.; Kato, T.; Sato, K.; Hikage, T.; Watanuki, T.; Mori, S. Elevated expression of P-Glycoprotein in kidney and urinary bladder cancers. Tohoku J. Exp. Med. 1991, 164, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Slot, A.J.; Molinski, S.V.; Cole, S.P. Mammalian multidrug-resistance proteins (MRPs). Essays Biochem. 2011, 50, 179–207. [Google Scholar] [CrossRef] [Green Version]
- Schaub, T.P.; Kartenbeck, J.; König, J.; Spring, H.; Dörsam, J.; Staehler, G.; Störkel, S.; Thon, W.F.; Keppler, D. Expression of the MRP2 gene-encoded conjugate export pump in human kidney proximal tubules and in renal cell carcinoma. J. Am. Soc. Nephrol. 1999, 10, 1159–1169. [Google Scholar]
- Scheffer, G.L.; Kool, M.; de Haas, M.; de Vree, J.M.L.; Pijnenborg, A.C.; Bosman, D.K.; Elferink, R.P.; van der Valk, P.; Borst, P.; Scheper, R.J. Tissue distribution and induction of human multidrug resistant protein 3. Lab. Investig. 2002, 82, 193–201. [Google Scholar] [CrossRef]
- Van Aubel, R.A.; Smeets, P.H.; Peters, J.G.; Bindels, R.J.; Russel, F.G. The MRP4/ABCC4 gene encodes a novel apical organic anion transporter in human kidney proximal tubules: putative efflux pump for urinary cAMP and cGMP. J. Am. Soc. Nephrol. 2002, 13, 595–603. [Google Scholar]
- Huls, M.; Brown, C.D.; Windass, A.S.; Sayer, R.; van den Heuvel, J.J.; Heemskerk, S.; Russel, F.G.; Masereeuw, R. The breast cancer resistance protein transporter ABCG2 is expressed in the human kidney proximal tubule apical membrane. Kidney. Int. 2008, 73, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Breljak, D.; Ljubojevic, M.; Hagos, Y.; Micek, V.; Eror, D.B.; Madunić, I.V.; Brzica, H.; Karaica, D.; Radovic, N.; Kraus, O.; et al. Distribution of organic anion transporters NaDC3 and OAT1-3 along the human nephron. Am. J. Physiol. Physiol. 2016, 311, F227–F238. [Google Scholar] [CrossRef] [Green Version]
- Atilano-Roque, A.; Roda, G.; Fogueri, U.; Kiser, J.J.; Joy, M.S.; Arya, V. Effect of disease pathologies on transporter expression and function. J. Clin. Pharmacol. 2016, 56, S205–S221. [Google Scholar] [CrossRef]
- Motohashi, H.; Sakurai, Y.; Saito, H.; Masuda, S.; Urakami, Y.; Goto, M.; Fukatsu, A.; Ogawa, O.; Inui, K.-I. Gene expression levels and immunolocalization of organic ion transporters in the human kidney. J. Am. Soc. Nephrol. 2002, 13, 866–874. [Google Scholar] [PubMed]
- Otsuka, M.; Matsumoto, T.; Morimoto, R.; Arioka, S.; Omote, H.; Moriyama, Y. From the Cover: A human transporter protein that mediates the final excretion step for toxic organic cations. Proc. Natl. Acad. Sci. USA 2005, 102, 17923–17928. [Google Scholar] [CrossRef] [PubMed]
- Pappireddi, N.; Martin, L.; Wühr, M. A Review on Quantitative Multiplexed Proteomics. Chembiochem. 2019, 20, 1210–1224. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Hosey-Cojocari, C.; Basit, A.; Unadkat, J.D.; Leeder, J.S.; Prasad, B. Optimized Renal Transporter Quantification by Using Aquaporin 1 and Aquaporin 2 as Anatomical Markers: Application in Characterizing the Ontogeny of Renal Transporters and Its Correlation with Hepatic Transporters in Paired Human Samples. AAPS J. 2019, 21, 88. [Google Scholar] [CrossRef] [PubMed]
- Jaitovich, A.A.; Bertorello, A.M. Na+, K+ -ATPase: An indispensable ion pumping-signaling mechanism across mammalian cell membranes. Semin. Nephrol. 2006, 26, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Basit, A.; Radi, Z.; Vaidya, V.S.; Karasu, M.; Prasad, B. Kidney cortical transporter expression across species using quantitative proteomics. Drug Metab. Dispos. 2019, 47, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.; Johnson, K.; Billington, S.; Lee, C.; Chung, G.W.; Brown, C.D.; Kelly, E.J.; Himmelfarb, J.; Unadkat, J.D. Abundance of drug transporters in the human kidney cortex as quantified by quantitative targeted proteomics. Drug Metab. Dispos. 2016, 44, 1920–1924. [Google Scholar] [CrossRef]
- Cheung, K.W.K.; van Groen, B.D.; Spaans, E.; van Borselen, M.D.; de Bruijn, A.C.J.M.; Simons-Oosterhuis, Y.; Tibboel, D.; Samsom, J.N.; Verdijk, R.M.; Smeets, B.; et al. A comprehensive analysis of ontogeny of renal drug transporters: mRNA analyses, quantitative proteomics, and localization. Clin. Pharmacol. Ther. 2019, 106, 1083–1092. [Google Scholar] [CrossRef]
- König, J.; Zolk, O.; Singer, K.; Hoffmann, C.; Fromm, M. Double-transfected MDCK cells expressing human OCT1/MATE1 or OCT2/MATE1: determinants of uptake and transcellular translocation of organic cations. Br. J. Pharmacol. 2011, 163, 546–555. [Google Scholar] [CrossRef]
- Deutsch, B.; Neumeister, C.; Schwantes, U.; Fromm, M.F.; König, J. Interplay of the organic cation transporters OCT1 and OCT2 with the apically localized export protein MATE1 for the polarized transport of trospium. Mol. Pharm. 2019, 16, 510–517. [Google Scholar] [CrossRef]
- Marquez, B.; Van Bambeke, F. ABC multidrug transporters: target for modulation of drug pharmacokinetics and drug-drug interactions. Curr. Drug Targets 2011, 12, 600–620. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Zhong, X.-B. Epigenetic regulation of drug metabolism and transport. Acta Pharm. Sin. B 2015, 5, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gröer, C.; Brück, S.; Lai, Y.; Paulick, A.; Busemann, A.; Heidecke, C.; Siegmund, W.; Oswald, S. LC–MS/MS-based quantification of clinically relevant intestinal uptake and efflux transporter proteins. J. Pharm. Biomed. Anal. 2013, 85, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Joseph, S.; Nicolson, T.J.; Hammons, G.; Word, B.; Green-Knox, B.; Lyn-Cook, B. Expression of drug transporters in human kidney: impact of sex, age, and ethnicity. Biol. Sex Differ. 2015, 6, 4. [Google Scholar] [CrossRef]
- Biermann, J.; Lang, D.; Gorboulev, V.; Koepsell, H.; Sindic, A.; Schröter, R.; Zvirbliene, A.; Pavenstädt, H.; Schlatter, E.; Ciarimboli, G. Characterization of regulatory mechanisms and states of human organic cation transporter 2. Am. J. Physiol. Cell Physiol. 2006, 290, C1521–C1531. [Google Scholar] [CrossRef]
- Engel, U.; Breborowicz, D.; Bøg-Hansen, T.; Francis, D. Lectin staining of renal tubules in normal kidney. APMIS 1997, 105, 31–34. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Protein Name | UniProtKB ID | Plasma Membrane Domain |
|---|---|---|---|
| ABCB1 | MDR1 (P-gp) | P08183 | apical [13] |
| ABCC1 | MRP1 | P33527 | basolateral [14] |
| ABCC2 | MRP2 | Q92887 | apical [15] |
| ABCC3 | MRP3 | O15438 | basolateral [16] |
| ABCC4 | MRP4 | O15439 | apical [17] |
| ABCG2 | BCRP | Q9UNQ0 | apical [18] |
| ATP1A1 | Na+/K+ ATPase 1 | P05023 | basolateral [19] |
| SLC15A1 | PEPT1 | P46059 | apical [20] |
| SLC15A2 | PEPT2 | Q16348 | apical [20] |
| SLC22A2 | OCT2 | O15244 | basolateral [21] |
| SLC22A3 | OCT3 | O75751 | basolateral (presumed) |
| SLC22A6 | OAT1 | Q4U2R8 | basolateral [19] |
| SLC22A7 | OAT2 | Q9Y694 | basolateral [19] |
| SLC22A8 | OAT3 | Q8TCC7 | basolateral [21] |
| SLC22A18 | ORCTL2 | Q96BI1 | apical [10] |
| SLC47A1 | MATE1 | Q96FL8 | apical [22] |
| Primer | Sequences (5′ → 3′) | |
|---|---|---|
| OCT2 | Forward | GGA ATA GCA TGG TTG AGG ACC A |
| Reverse | GGG GCT ATG ATT CCC CCA AAA | |
| MATE1 | Forward | AAG CTG GAG CTG GAT GCA GTC |
| Reverse | CAG CAG AGG AGC AGG ACG AGC | |
| OAT1 | Forward | CAG ACA GCT GTG TCA GGG AC |
| Reverse | GAA TGG GCA TCC ACT CCA CA | |
| OAT2 | Forward | CTG GTT GGG TAC CTG ATA CGG |
| Reverse | CAA GTA CCT GTG GGC CTC TTT | |
| GAPDH | Forward | CAA GCT CAT TTC CTG GTA TGA C |
| Reverse | GTG TGG TGG GGG ACT GAG TGT GG | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oswald, S.; Müller, J.; Neugebauer, U.; Schröter, R.; Herrmann, E.; Pavenstädt, H.; Ciarimboli, G. Protein Abundance of Clinically Relevant Drug Transporters in The Human Kidneys. Int. J. Mol. Sci. 2019, 20, 5303. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215303
Oswald S, Müller J, Neugebauer U, Schröter R, Herrmann E, Pavenstädt H, Ciarimboli G. Protein Abundance of Clinically Relevant Drug Transporters in The Human Kidneys. International Journal of Molecular Sciences. 2019; 20(21):5303. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215303
Chicago/Turabian StyleOswald, Stefan, Janett Müller, Ute Neugebauer, Rita Schröter, Edwin Herrmann, Hermann Pavenstädt, and Giuliano Ciarimboli. 2019. "Protein Abundance of Clinically Relevant Drug Transporters in The Human Kidneys" International Journal of Molecular Sciences 20, no. 21: 5303. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215303