Pluripotent Cell Models for Gonadal Research


Endocrinology Division, Department of Endocrinology, Metabolism and Cardiovascular System, Section of Medicine, University of Fribourg, 1700 Fribourg, Switzerland
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(21), 5495; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215495
Submission received: 3 October 2019
/
Revised: 30 October 2019
/
Accepted: 1 November 2019
/
Published: 4 November 2019
(This article belongs to the Special Issue Molecular Aspects of Sex Development in Mammals: New Insight)
Abstract
:Sex development is a complex process involving many genes and hormones. Defects in this process lead to Differences of Sex Development (DSD), a group of heterogeneous conditions not as rare as previously thought. Part of the obstacles in proper management of these patients is due to an incomplete understanding of the genetics programs and molecular pathways involved in sex development and DSD. Several challenges delay progress and the lack of a proper model system for the single patient severely hinders advances in understanding these diseases. The revolutionary techniques of cellular reprogramming and guided in vitro differentiation allow us now to exploit the versatility of induced pluripotent stem cells to create alternatives models for DSD, ideally on a patient-specific personalized basis.
1. Introduction
The normal gonadal development in humans requires the participation of two types of cells: germ cells and somatic support cells. The germ cells are the precursors that will originate the corresponding gametes, oocytes in females and sperm cells in males. The progressive transformation of primordial germ cells into gametes cannot be accomplished without the tight control of the somatic support cells that surround them.
In males, this supportive role is taken up by Sertoli cells (SCs). SCs constitute the structural component of the seminiferous tubule, creating the appropriate niche for the germ cells isolating them from the soma thanks to the establishment of the blood–testis barrier (BTB). Yet, the role of SCs is not limited to that. They produce the sex-determining region Y-box 9 (SOX9), a key factor during male sex development, and they also perform a precise control on the cellular transformation of spermatogonial stem cells into mature spermatozoa through the secretion of anti-mullerian hormone (AMH) (Figure 1). It is because of this control over the germ cells that they are often referred as “mother” or “nurse” cells [1]. The steroidogenic production of testosterone is carried out by Leydig cells located in the interstitial tissue between seminiferous tubules [2]. At puberty, SCs mature reducing dramatically the expression of AMH and their proliferation and motility, gaining the ability to support spermatogenesis [3,4].
In females, the supporting role is taken up by the granulosa cells (GCs), also known as follicular cells. In contrast to their male counterpart, one can differentiate between several groups of granulosa cells with specific roles in the control of the oocyte environment. Localized tightly surrounding the primordial oocytes, a group of GCs known as cumulus cells (CCs) control the differentiation and maturation of the oocyte through cell-to-cell interaction (via gap junctions) and paracrine action.
A major player in the development and maintenance of the female phenotype is wnt family member 4 (WNT4): this secreted protein is almost exclusively expressed by GCs during early ovarian development, and also regulates the formation of Müllerian ducts. WNT4 counteracts the effects of fibroblast growth factor 9 (FGF9) and SOX9 in a very delicate equilibrium, with WNT4 turning the balance into ovary development and FGF9 and SOX9 into the testis side. The CCs also produce AMH, whose correlation with follicular aging was well stablished [5].
A second type of GCs known as mural granulosa cells (MCs) rest on a basal lamina and separate the oocyte environment form the vascularized theca interna, creating a blood–follicle barrier. The mural granulosa cell produces a much lower amount of AMH than cumulus cells and its endocrine function is oriented to the production of steroidogenic enzymes like Cytochrome P450 aromatase and the conversion of androgens from the theca cells (androstenedione and testosterone) into estrogens like estradiol [6].
Both populations of GCs show expression of the forkhead box protein L2 (FOXL2) which counteracts the action of SOX9 in female sex development [7,8]. The expression of FOXL2 is indeed observed very early in ovarian development, in the cellular progenitors of GCs. FOXL2 acts over transcriptional targets of various cellular pathways [9]. Studies in mice and goats associated FOXL2 with the repression of SOX9-guided testis determination [10,11]. In a complimentary manner to the FOXL2 signaling pathway, WNT4, β-catenin (CTNNB1) and R-spondin 1 (RSPO1) act during ovarian differentiation to inhibit SOX9 effect on transdifferentiation of pre-granulosa cells (pre-GCs) into SCs after sex determination in mice [12]. Still many things remain unknown about the role of both FOXL2 and RSPO1 in humans but, interestingly, RSPO1 expression had been be detected in human undifferentiated gonads of both sexes but it is upregulated only in the ovary during critical stages of early gonad development (6–9 weeks post fertilization) [13]. Apart from FOXL2, CCs and MCs show marked differences in the expression and regulation of receptors for the luteinizing hormone (LH) and follicle-stimulating hormone (FSH), central elements in the ovarian follicle development [14]. In females, the communication between germ cells and somatic supporters occurs in both directions, having oocyte also control cumulus cells via growth differentiation factor 9 (GDF9), oocyte secreted factors (OSFs) or bone morphogenic protein 15 (BMP15) [15,16].
Alterations of the gonadal germ and/or somatic cells at a genetic or structural level during sexual development often lead to differences/disorders of sex development (DSD) in humans and many different species [17,18,19]. The term DSD involves a group of complex conditions that can affect chromosomal, gonadal, and/or phenotypical sex. These diseases are heterogeneous and not as rare as previously thought: it is estimated that genital anomalies occur in 1:4500–5500 births [20]. They compromise not only internal and external genitalia formation, but also affect fertility to various degrees [21]. AMH expression was observed to be altered in 46,XY DSD conditions like complete and partial gonadal dysgenesis, and ovotesticular dysgenesis among others [22,23]. In fact, AMH levels have become a highly reliable indicator of the existence of testicular tissue in prepubertal patients, while undetectable levels suggest testicular dysgenesis.
Based on results from affected mouse gonads, under loss of FOXL2, SOX9 expression is de-repressed in GCs, which transdifferentiate into SCs. In humans, heterozygous FOXL2 disruption was associated with premature ovarian failure (POF) [24] but homozygous FOXL2 deletion patients have not been reported so far. WNT4 mutations have been associated with Müllerian-duct regression and virilization in 46,XX patients [17] and some cases of 46,XX testicular and ovotesticular DSD are linked to mutations of the RSPO1 gene [25,26]. All these hints suggest that the genes responsible for GCs differentiation also have a high impact on ovarian development.
Consequently, the study of the actions of germ cells and their somatic supporters at a cellular level has been revealed as essential to understand the complex mechanisms underlying the different DSD. In this review, we will analyze the different approaches to modeling DSD gonadal cells and the doors that new induced pluripotent stem cells (iPSCs) technology has recently opened for DSD research.
2. Finding a Perfect Match: The Lack of Right Models for DSD Patients
Due to the high complexity and the great variety of DSDs, the most useful approach would be the study of cells extracted directly from the patient. The first obstacle that this methodology faces is the fact that in some severe cases of DSD with complete gonadal dysgenesis, the patient lacks those types of cells. This limitation cannot be overcome with any of the currently available cellular models (see below), and led to the search of new approaches, ultimately leading to personalized cell modeling. Even when gonadal somatic supporter cells are present, their very limited number significantly complicates the establishment of primary cultures. Animal models and cell lines derived from gonadal tumors have so far been the main alternatives to study the mechanism of disease that lead to DSD. Such models, though, show important differences due to their tumoral origin and the fact that the sex development pathway is not particularly well conserved among species.
2.1. Sertoli Cell Models
Most of the in vitro studies on SCs development were performed in immature rodent testes. The first SC line established, the mouse TM4 cell line, was generated by passing cultures of immature mouse SCs over many generations to generate an immortal line [27]. Other mouse SC-models like 15P-1 cells, where immortalized from testis of transgenic mice expressing large T protein of polyomavirus [28]. Although useful to understand common characteristics of mammalian SCs, murine models have been demonstrated to differ from humans regarding sex determination and development [29,30,31]. Murine SCs have also shown little proliferative ability in vitro, leading to the thought that mammalian SCs do not divide postpuberty. This hypothesis has been recently challenged when studies with SCs isolated from cadaveric testes [32] or donors undergoing bilateral castration [33] demonstrated the ability of human SCs to proliferate when cultured in vitro. Thus, primary human Sertoli cell lines known as HSerCs are now commercially available. Techniques like differential plating seems to improve the viability of isolated SCs [34] but it is very important to remind that, when the focus of the study is the role of SCs in male development, a reliable source of immature SCs from prepubertal patients is essential. Adult patients lack those immature cells and testicular biopsies from very young patients pose important ethical questions.
An alternative that overcomes the problem of in vitro culture of SCs is the use of human cancer cell sources. One of these cell lines is represented by NT2d1 cells derived from pluripotent clonal cells obtained from testicular tumors [35]. NT2d1 cells were extensively used for SCs studies since they express most of genes involved in sex determination [36]. However, we have recently observed a very different transcriptome pattern between NT2d1 cell and primary human SCs when we observed genes related with SC differentiation and maturation [37]. Those findings suggest that their carcinogenic origin partially altered their SC characteristics, and renders these cells less than an ideal model for SCs.
2.2. Granulosa Cell Models
Human GCs are often recovered from women undergoing in vitro fertilization (IVF) after ovarian stimulation and ovulation induction. GCs are isolated from follicle fluid during oocyte retrieval procedures. It is important to remark that, following ovulation, GCs undergo a process of luteinization which involves structural and genomic changes that lead to the terminal differentiation of follicular cells with increased progesterone production [38]. The differentiation of granulosa cells into luteinized cells has effects on intracellular signaling and cell cycle regulation. As mature SCs, luteinized GCs stop their proliferation, which makes the long-term cultivation of GCs primary cells extremely challenging [39].
The initial studies in this direction by Smith et al. [40] and Byong-Lyul et al. [41] confirmed a transformation of the harvested population of GC, observing two different populations in vitro. Some luteinization processes seem to be initiated in these cells but they never achieved the lutein levels observed in situ. It became clear that GCs faith is extremely dependent on its microenvironment. Bruckova et al. [42] described the negative effect of high concentrations of fetal calf serum (FCS) in culture medium, promoting the apparition of a very heterogeneous population that degenerate after a short period (18 days) in culture. The presence of LH acts as a trigger for the luteinization process reducing GCs proliferation while FSH induces a series of mitotic divisions. In a pioneer study the same group [39] highlighted the importance of the follicle fluid in addition to growth factors like fibroblast growth factor basic (bFGF) and epidermal growth factor (EGF) to maintain the proliferative potential of GCs primary cultures. This work shed some light on the preservation and culture of GCs from patients whose gonads have suffered alteration but are not completely absent while it is still unclear if altered GCs will require a different microenvironment to keep their mitotic potential.
In the same fashion as SC, researchers resorted also to cell lines derived from carcinogenic tissues. KGN as well as COV434 cells were obtained from human ovarian granulosa cell tumors [43,44]. KGN cells became the most common cellular model used for the study of human granulosa cells. Although KGN have great potential for steroidogenic studies and their proliferation and immortality make them an easy model to work with in the laboratory, characterization analyses showed that KGN have a mutated form of FOXL2 related to the loss of apoptosis induction mechanism in granulosa tumors. This mutation was not present in any of the other cell models analyzed [45,46]. On the other side, immortalization of human granulosa cells from oocyte retrieval during IVF (HGL5 and HO-23) resulted in cells lacking responsiveness to FSH or human chorionic gonadotropin (hCG) [47,48]. Bayasula et al.’s work on nonluteinized granulosa cell immortalization [49] reports a very similar transcriptome of lentivirus immortalized HGrC1 when compared with primary granulosa cells. Unlike other granulosa cell models, HGrC1 show proliferation and steroidogenesis under gonadotropin regulation, suggesting that this model keeps original granulosa cell functional activity.
As discussed for SCs, animals can be also a source of GCs. In the same way as SCs models, species-specific differences in granulosa cell function make these models not ideal for human mechanism studies. As an example of this hypothesis, it is known that mutations of the bone morphogenetic protein 15 (BMP-15) gene cause infertility in sheep and POF in humans while mice show normal follicle development [50,51,52].
2.3. Germ Cell Models
Modelling germ cell lineage can add an extra layer of difficulty considering that they are the only cells transmitted to the next generation. Germ cells are tightly dependent on the interaction of multiple cell types for their development and differentiation [53]. The mechanisms underlying primordial germ cell (PGC) specification are not conserved even across related species. In mammals, recent studies suggest that the specification of mice PGCs differs from those in humans, suggesting that different molecular regulatory mechanisms govern PGC development in both species [54,55,56,57]. Therefore, researchers look at embryonic stem cells (ESCs) for alternatives. Many groups have successfully differentiated germline cells from ESCs in vitro [58,59,60,61,62]. These cells, known as primordial germ cell-like cells (PGCLCs), show a great potential for the study of germ cell differentiation and their interaction with somatic precursors during sexual development. However, embryonic stem cell research depends exclusively on ESCs lines isolated from “discarded” embryos in IVF clinics. This is difficult to reconcile with the need to obtain patient-specific cell sources for the study of DSD.
3. Unleashing the Power of Pluripotency: iPSCs and Guided Differentiation
The availability of specific cell models for the diverse DSD variants might help to better understand the mechanisms involved in human sex development and to improve care for DSD patients. New technologies in the area of cell reprogramming opened a window of opportunity for a revolutionary form of personalized medicine: the reprogramming of terminally differentiated cells into induced pluripotent stem cells (iPSCs) and their guided differentiation into virtually any cell type (Figure 2).
In 2006, Shinya Yamanaka’s group combined the previous knowledge on nuclear transfer [63,64,65], transcription factors [66,67,68] and embryonic stem cells [69,70,71] to induce reprogramming of terminally differentiated fibroblasts into pluripotent stem cells by retroviral transduction [72]. From 24 different factors tested, Yamanaka et al. discovered that the combination of POU class 5 homeobox 1 (OCT3/4), kruppel like factor 4 (Klf4), SRY-box 2 (SOX2), and proto-oncogene C-Myc (C-Myc), all related to undifferentiated ESCs self-renewal and maintenance, played a key role for the generation of the iPSCs. Thanks to this pioneering work, Yamanaka was awarded the Nobel Prize in Physiology or Medicine 2012.
Although iPSCs maintain the essential features of ES cells to propagate in culture indefinitely and to differentiate into each of the three embryonic germ layers, their origin from somatic cells and the reprogramming methods used may lead to significant differences between iPSCs and ESCs [73]. Additionally, the use of retrovirus in the mentioned Yamanaka study to transduce reprogramming factors has certain limitations in personalized medicine due to observed integrations of those factors into the endogenous genome [72]. Furthermore, C-MYC is a known prooncogene and its function as a proliferation enhancer may be achievable by other nononcogenic factors, resulting in less tumorigenic iPSCs [74,75]. Yu et al. [76] refined the reprogramming procedure adding a different cocktail of factors. In this case, OCT4), SOX2, nanog homeobox (NANOG), and lin-28 homolog A (LIN28) were transduced using lentivirus, which increased the efficiency of reprogramming due to the ability to infect non dividing cells. Although of great potential for general cell models, the use of lentivirus transduction is also not very suitable for patient-specific modeling since, despite reducing the carcinogenic potential of c-MYC, the insertional mutagenesis is still a concern [77].
During recent years, many researchers sought alternative ways to generate integration-free iPSCs. Episomal vectors [78], synthesized RNA [79] and Sendai virus, a single chain RNA virus [80], have become of routine use in reprograming experiments. Every system shows advantages and disadvantages that may be differently adapted for every laboratory. Technology based on RNA like synthetized mRNA or Sendai virus completely eliminate the risk of genomic integration intrinsic to all DNA-based approaches but modified mRNA is technically complex and Sendai virus experiments need to be performed under strict biological containment [80]. Conversely, episomal vectors offer an excellent approach due to the simplicity and reproducibility of the method [81]. Modified-mRNAs are lost extremely quickly from host cells and this reprogramming methodology requires daily transfections [82]. Both episomal vectors and Sendai virus remain in the cytoplasm of the host cells but they are lost after several generations [83].
As to the intrinsic efficiency of every iPSCs generation methodology, we have to add the fact that not all cell types are equally receptive to reprogramming [84]. Moreover, scientists have recognized important differences between iPSC lines from diverse origins. These variations are mostly connected to the inherent plasticity of somatic cells [85,86,87]. Novel insights suggest that the original metabolic environment affects the faith of future iPSCs even after reprogramming [88,89]. This principle seems to also affect SC-derived iPSCs [90]. Epigenetic memory was gradually lost in iPSCs after passages, due to either post-reprogramming removal of the epigenetic pattern of somatic cells or selective pressure against iPSC with epigenetic memory [89]. Fibroblasts are the most common cell source used for iPSCs generation thanks to good reprogramming efficiency rates and the simplicity of their culture. Nevertheless, peripheral mononuclear blood cells (PBMCs) [91] or urinary progenitors (UPs) collected from spontaneous urine [92,93] have gained importance in an effort to obtain a high amount of reprogrammable cell sources with a less invasive sampling procedure.
Developing new iPSCs requires exhaustive characterization of new lines in order to define the iPSC populations and to demonstrate their pluripotency. The analysis of the endogenous expression of pluripotency markers like OCT4, stage-specific embryonic antigen 3 (SSEA3), stage-specific embryonic antigen 4 (SSEA4) and TRA-1-81 allows for discrimination between high quality iPSCs and partially reprogrammed colonies. The prolonged culture of iPSCs lines can result in genetic abnormalities causing aneuploidy. Thus, analysis of the karyotype of iPSCs is of vital importance when aiming to provide patient-specific medicine [94]. iPSCs colonies that show genetic abnormalities must be discarded to preserve resemblance to the patient. In addition, high quality iPSCs must demonstrate their ability to differentiate into primordial germ layers of the embryo, i.e., ectoderm, mesoderm and endoderm, in vitro and in vivo [95].
Once stablished, an iPSCs cell line has the potential to differentiate in multiple cellular lines. This process may occur spontaneously, causing the contamination of the culture with undesired populations. A tight control on the environment is necessary to force differentiation of all cells through the same desired path. The first step involves the generation of embryoid bodies, wherein differentiation occurs as a result of the transfer of iPSCs into a suspension culture environment. From this point on, a tight differentiation control can be achieved by the step-wise addition of growth factors, cytokines or inhibitors based on the knowledge of specific differentiation pathways for every desire cell line [96].
4. Modeling the Complexity: Human iPSCs-Derived Models for DSD Research
Our knowledge of human sex differentiation has increased progressively in recent years although advances were not equally represented in both genders. Mechanisms underlying male sexual differentiation have been well studied while many factors regulating female sexual differentiation remain unknown. The current gonadal cell models generated through human iPSCs (hiPSCs) differentiation are summarized in Table 1.
4.1. hiPSCs and Somatic Gonadal Cell Models
In SCs modelling, better understanding of many of the factors involved finally offered us the opportunity to recapitulate somatic support cell differentiation in vitro in a relatively accurate manner. In this respect, original work by Buganim et al. proved the ability of iPSCs to differentiate into SCs [109] in mice. Bucai et al. [97] observed that, when human embrionic stem cells (hESCs) are differentiated into PGCs, the same PGCs seem to guide the differentiation of uncommitted or primitive cells present in the culture into Sertoli-like cells (SLCs), possibly through paracrine factors released in the medium. After obtaining similar results with umbilical cord perivascular cells, Shlush et al. [98] identified the secretion of bone morphogenic protein 4 (BMP4) by undifferentiated cells to be pivotal for the differentiation into SCs.
SOX9, fibroblast growth factor basic (bFGF), fibroblast growth factor 9 (FGF9), prostaglandin 2 (PGD2) and Steroidogenic Factor 1 (SF1) have also been considered as candidate factors due to their role in Sertoli cell differentiation. SOX9 expression is crucial in SCs differentiation and needs to be maintained during the whole process. bFGF and BMP4 are two key regulators in the formation of the urogenital system, via mesoendoderm signaling [110,111]. It is known that both factors are able to stimulate the expression of SOX9 [112]. The expression of SOX9 can also be stimulated in an independent way by PGD2 and FGF9 [113,114]. Remarkably, the action of SF1 has been emphasized as essential for fetal SCs survival. SF1 controls the cell cycle of Sertoli cells during differentiation by regulating the tumor protein P53 (TP53) pathway [115] while up to date, to our knowledge no peer-reviewed articles have proved its effect on iPSCs guided differentiation.
In our study [37], embryoid bodies were treated with a cocktail of growth factors including bFGF, BMP4, PGD2, FGF9 and activin A to promote the endogenous expression of SOX9 until the cells changed morphology and resembled SCs appearance. The elevated expression of SOX9 observed reflected its successful stimulation by the administered growth factors cocktail. The addition of activin A to the cocktail allowed the further expansion of the culture due to its role in SCs proliferation [116].
Characterization with new generation sequencing techniques revealed that this was new and until today the unique human SLCs model is actually closer to HSerCs than the cancer-derived NT2d1 cells. Indeed, SLCs bear a greater resemblance to fetal SCs than to mature cells, a characteristic observed in other iPSCs-derived models [96], but with great interest for the study of SC role during sexual development. These SLCs are still a proof of concept and have some limitations including the use of lentivirus transduction to generate iPSCs and the lack of a long-term study of SLCs characteristics. Nevertheless, it opens the way to patient-specific SC models in DSD research.
Several successes were also achieved in GC modeling. Experiments with murine cells [117,118] demonstrate the effective differentiation of mouse and rat iPSCs into GC-like cells by co-culture with GCs. Those GC-like cells expressed several genes related to granulosa cell function like BMP15, FSHR, AMH and GDF9 and were able to secrete estrogen. These observations highlight the fact that cell-cell connection and factors generated by the differentiated GC in the co-culture can induce the differentiation of stem cells through the granulosa pathway. In line with these results, Liu et al. in 2016 [100] induced the differentiation of human iPSCs into GC-like cells through the addition of a growth factors cocktail to the culture. The cocktail was composed by all-trans-retinoic acid, estradiol, AMH, FSH, Inhibin α, Inhibin β and TGF–β; all crucial for the maintenance of ovarian follicular development and secreted by GCs.
In an independent way, Lipskind et al. [101] demonstrated that native differentiation of human amniocytes-derived iPSCs can produce functional granulosa-like cells without intervention of additional growth factors. Whether this approach is valid for iPSCs derived from different cellular origins has to be elucidated. This growth factors-free differentiation might be taking place thanks to the metabolic memory discussed above, turning the differentiation balance into GCs faith. In line with these thoughts, Anchan et al. [99] have shown how iPSCs derived from GCs were able to redifferentiate into homotypic ovarian steroidogenic cells in vitro. Even though the results of this methodology are inapplicable for most of DSD conditions, it pointed out that the quality of the iPSCs-derived cell model may be dependent on the original cell source.
4.2. hiPSCs and Germ Cell Models
The accurate timing of human germ cells specification cannot be determined precisely due to the difficulties and ethical boundaries of using human embryos for research studies. We already mentioned that the mechanisms leading to germ cell specification are not well conserved among different species, making the previous differentiation approaches in mouse and rat iPSCs [119,120] not completely applicable to human cell models [121]. In humans, SOX17 seems to have a key role in embryonic germ cell differentiation and controls the expression of B-lymphocyte-induced maturation protein 1 (BLIMP1), which is the leading gene of germ cell induction in mice. Additionally, SOX2 is essential for mouse germ cell development but is not detected in human embryonic germ cells [55,122]. There are, however, some islands of germ cell differentiation mechanisms that are well conserved across the animal phyla, specially related to mitotic and meiotic phases of germ cells development [123,124]. Those common genes may be used as a foundation stone to build our knowledge of human germ cell development. For instance, genes like nanos homolog 3 (NANOS3) that proved to be crucial during murine germ cell development, seem to play a maintenance role in early germ cell development in humans [125,126].
In the quest to unravel the machinery of germ cell specification and development in both sexes, first attempts from Kee et al. [127] reported the importance of bone morphogenetic protein (BMPs) genes to promote germ cell formation from iPSCs. The same group discovered the action of deleted in azoospermia like (DAZL), deleted in azoospermia (DAZ) and boule homolog (BOULE) promoting germ-cell progression to meiosis and formation of haploid germ cells [102]. They also demonstrated that the expression of vasa homolog (VASA), a germ cell-specific marker, is indeed regulated by DAZL. The DAZ family, located on the Y-chromosome, is one of the conserved clusters among species and has been reported as essential for male fertility. Its deletion leads to azoospermia and oligospermia in humans. These advances allowed for the transformation of iPSCs even from adult human cell sources into advanced meiotic germ cells by overexpression of DAZL, DAZ and BOULE proteins [104,106]. Alternatively, hiPSCs-derived robust meiotic germ cells were obtained by a two-step protocol, removing bFGF at first and subsequently RA, forskolin, leukemia inhibiting factor (LIF) and CYP26 inhibitor [103]. hiPSCs differentiated with mouse spermatogonial stem cell medium expressed markers for post-meiotic spermatocytes and round spermatids [107]. Nonetheless, in our knowledge, complete spermatogenesis in vitro has not yet been accomplished in human.
In human female iPSCs, Eguizabal et al. showed that overexpression of DAZL and BOULE can also lead iPSCs to enter meiosis. Yang et al. [105] used fibroblasts from premature ovarian insufficiency (POI) patients to successfully differentiate them into PGCs that had the potential for meiotic progression in vitro. Mimicking the natural role of the oocyte by stimulation with GDF9 and BMP15 proved further induction of folliculogenesis in differentiated PGCs [108]. Transcriptome analysis of these follicle-like cells (FLCs) resembled in vivo primordial follicles. In the same study, researchers observed that the population of differentiated ESCs organize itself when exposed to the differentiation factors, generating an oocyte-like cell surrounded by a cellular aggregate with Granulosa-like cells at the outer layer. Together with the previous observations discussed above, this new evidence brings increased consistency to the hypothesis of microenvironment guidance over GCs differentiation.
Taken together, great progress has been accomplished in gonadal cell modeling, proving that both male and female iPSCs enclose the potential to differentiate into any gonadal cell type, including germ cells (Figure 3). The use of cellular reprogramming is now leading a pioneering pathway in the generation of fully patient-specific cell models with enormous advantages for their use in DSD research.
5. Future of DSD Models
Our comprehension of the complex processes that drive the mechanism of disease in DSD patients has always been limited by the lack of the proper cells involved in those mechanisms, either by the impossibility to obtain samples from the patient or the absence of orthologous cellular models.
For the last 12 years, the innovative techniques of cellular reprogramming has sparked a revolution in many areas of medicine, including DSD research. Thanks to this powerful tool, we are now able to reprogram virtually any cell form a patient and iPSCs have become an exceptional autologous cell source for cell replacement therapy for a large number of diseases. Looking at DSD patients, we have the opportunity to guide iPSCs re-differentiation into somatic supporters and germ cells, making personalized cell models to reveal the individual complexity underlying such a diverse group of conditions.
Nevertheless, the capacity of differentiation techniques to guide correctly the destiny of the cells is completely dependent on our knowledge of the processes governing the sexual development in humans, which are not totally understood.
Limitations and Uncertainties
Cellular models resembling early development Sertoli and granulosa cells as well as primordial germ cells lacked long-term stability. For example, our preliminary results of long-term culture of SLCs showed a reduction of SOX9 expression after 30 days of culture. This time window suffices to study the mechanism of disease in DSD patients but might be too short for more complex long-term studies, such as therapy or regenerative medicine.
One major challenge that remains is the difficulty to generate terminally differentiated cells from iPSCs. As we discussed through this review, cell differentiation is a highly complex process probably including not only gene transcription changes but also epigenetic and translational regulations that may be essential for late differentiation steps. Taking advantage of the astonishing evolution of new generation techniques, we can now explore the effects that epigenetic and proteomic regulations may have on cellular reprogramming and differentiation. Proteomic study in iPSCs-derived hepatocytes [128] has identified critical regulatory molecules that creates a transcriptional fingerprint during terminal differentiation. We are confident that further studies will provide additional clues to understand the implication of posttranslational regulation in the terminal differentiation of iPSCs-derived cultures, including gonadal cell models.
Novel techniques in gene editing can be also translated into iPSCs-cell modeling research. CRISPR/Cas9 has proven to be a versatile genetic editing tool to correct mutations on iPSCs-derived cell models [129]. Researchers can now study modifications in disease severity by introducing gene deletion or repairing knocked-out expression in isogenic iPSCs-derived cells [130].
A second point to discuss is the selection of cellular sources. As we already mentioned [99,101], the pre-iPSC origin has a strong influence on the faith of (re)differentiated cells. Cellular populations from tissues with ontogenetic similarities to gonadal structures necessitate less complex guided differentiation to generate gonadal somatic supporters. By selecting original cell sources closely related to gonadal cells, we may increase the quality of future cell models. The combination of both a better understanding of ruling factors of sex development and selection of the best cell source available would produce cell models of excellent quality with greater stability under long-term culture.
Another factor that has gained importance is the physical and chemical communication between all the cells of a niche for their correct differentiation and maintenance [39,42,117,118]. This concept is not new: in adult tissues, regeneration is maintained by populations of multipotent progenitor cells that keep proliferation abilities and differentiate when needed. This differentiation is regulated by signals flowing from terminally differentiated cells of the niche to fulfill the demands of the tissue [131]. This idea of the microenvironment as a driving force for differentiation includes not only cell-cell interactions but also soluble factors, extracellular matrix (ECM) proteins and physical conditions like hypoxia, pressure or stiffness of the culture surface [132]. The generation of a synthetic microenvironment that mimics the physiologic conditions where Sertoli, granulosa and germ cells are developed might boost the efficiency of the differentiation process by reducing the chances of spontaneous differentiation and increasing the stability of the whole complex. To this respect, it is now possible to coordinate stem cells into 3D structures, known as organoids, which resemble the native multicellular architecture and functional features of a broad range of organ tissues [133]. Organoids and other 3D-models like organs on a chip (OOC) are platforms to understand the unique mechanisms of human developmental biology and offer an opportunity to model diseases in a more complex way, including those affecting sexual development.
We envision that co-culture of Sertoli-like cells and spermatid-like cells derived from the same cell source within a synthetic matrix that recreates the microtubules environment may not only improve the stability of both cell models but also have a synergistic effect, where SLCs would be able to mature and promote spermatogenesis. This is equally applicable to oocytes, surrounded by granulosa-like cells in a synthetic scaffold as suggested by Laronda et al. [134]. The authors also hypothesized that triggering the initial steps of development and maturation by FSH and LH in oocytes and by RA for Sertoli cells [135], these synthetic ovary or seminiferous tubules would mimic the natural release of all hormones and factors necessary to support reproductive features [134]. Woodruff’s group developed a two-step follicle culture strategy, supplementing medium with E2, FSH and human growth hormone (hGH) followed by the addition of AMH and estradiol, as a recapitulation of the dynamic human follicle growth environment [136]. A distinct advantage of using iPSCs for this purpose lies on their patient-specific origin so that their antigenically match to the acceptor cells, provide a platform for autologous therapies. Applied to a co-culture of hiPSCs-derived cell models, this methodology could improve the (re)generation of personalized synthetics reproductive organs.
The works reviewed are still mainly a proof of concept but lay the groundwork for what would come in the following years. The standardization of both every-time better reprograming methodologies and new generation analyses will aim to boost the quality and robustness of iPSCs-derived models. Together with an increased knowledge of the mechanisms and players involved in gonadal cells differentiation, these advances would open the door for truly patient specific gonadal cell models of priceless value to understand the mechanism of disease in different DSD conditions.
The production of gonadal cells from DSD patients may enable not only the of study in vitro therapies that target the factors affected in DSD but also boost our understanding of the key elements behind normal sex development and gonadal cell survival. This may ultimately contribute to new strategies for the diagnosis and treatment of unexplained infertility, a common health problem that affects approximately 15% to 30% of couples at reproductive age [137].
Author Contributions
Conceptualization, D.R.G. and A.B.-L.; investigation, D.R.G.; resources, A.B.-L.; data curation, D.R.G.; writing—original draft preparation, D.R.G. and A.B.-L.; writing—review & editing, D.R.G. and A.B.-L.; visualization, D.R.G.; supervision, A.B.-L.; project administration, A.B.-L.; funding acquisition, A.B.-L.
Funding
This work was supported by the Swiss National Science Foundation (Grant 320030_160334) and the Research Pool Grant of the University of Fribourg.
Acknowledgments
We would like to thank all those researchers who have dedicated their professional careers to understand and improve the lives of DSD patients.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Franca, L.R.; Hess, R.A.; Dufour, J.M.; Hofmann, M.C.; Griswold, M.D. The Sertoli cell: One hundred fifty years of beauty and plasticity. Andrology 2016, 4, 189–212. [Google Scholar] [CrossRef] [PubMed]
- Zirkin, B.R.; Papadopoulos, V. Leydig cells: Formation, function, and regulation. Biol. Reprod. 2018, 99, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Lucas, T.F.; Nascimento, A.R.; Pisolato, R.; Pimenta, M.T.; Lazari, M.F.; Porto, C.S. Receptors and signaling pathways involved in proliferation and differentiation of Sertoli cells. Spermatogenesis 2014, 4, e28138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crisostomo, L.; Alves, M.G.; Gorga, A.; Sousa, M.; Riera, M.F.; Galardo, M.N.; Meroni, S.B.; Oliveira, P.F. Molecular Mechanisms and Signaling Pathways Involved in the Nutritional Support of Spermatogenesis by Sertoli Cells. Methods Mol. Biol 2018, 1748, 129–155. [Google Scholar] [CrossRef]
- Kedem, A.; Yung, Y.; Yerushalmi, G.M.; Haas, J.; Maman, E.; Hanochi, M.; Hemi, R.; Orvieto, R.; Dor, J.; Hourvitz, A. Anti Mullerian Hormone (AMH) level and expression in mural and cumulus cells in relation to age. J. Ovarian Res. 2014, 7, 113. [Google Scholar] [CrossRef]
- Stocco, C. Aromatase expression in the ovary: Hormonal and molecular regulation. Steroids 2008, 73, 473–487. [Google Scholar] [CrossRef] [Green Version]
- Cocquet, J.; De Baere, E.; Gareil, M.; Pannetier, M.; Xia, X.; Fellous, M.; Veitia, R.A. Structure, evolution and expression of the FOXL2 transcription unit. Cytogenet. Genome Res. 2003, 101, 206–211. [Google Scholar] [CrossRef]
- Cocquet, J.; Pailhoux, E.; Jaubert, F.; Servel, N.; Xia, X.; Pannetier, M.; De Baere, E.; Messiaen, L.; Cotinot, C.; Fellous, M.; et al. Evolution and expression of FOXL2. J. Med. Genet. 2002, 39, 916–921. [Google Scholar] [CrossRef]
- Caburet, S.; Georges, A.; L’Hote, D.; Todeschini, A.L.; Benayoun, B.A.; Veitia, R.A. The transcription factor FOXL2: At the crossroads of ovarian physiology and pathology. Mol. Cell Endocrinol. 2012, 356, 55–64. [Google Scholar] [CrossRef]
- Pailhoux, E.; Vigier, B.; Chaffaux, S.; Servel, N.; Taourit, S.; Furet, J.P.; Fellous, M.; Grosclaude, F.; Cribiu, E.P.; Cotinot, C.; et al. A 11.7-kb deletion triggers intersexuality and polledness in goats. Nat. Genet. 2001, 29, 453–458. [Google Scholar] [CrossRef]
- Uhlenhaut, N.H.; Jakob, S.; Anlag, K.; Eisenberger, T.; Sekido, R.; Kress, J.; Treier, A.C.; Klugmann, C.; Klasen, C.; Holter, N.I.; et al. Somatic Sex Reprogramming of Adult Ovaries to Testes by FOXL2 Ablation. Cell 2009, 139, 1130–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maatouk, D.M.; Mork, L.; Chassot, A.A.; Chaboissier, M.C.; Capel, B. Disruption of mitotic arrest precedes precocious differentiation and transdifferentiation of pregranulosa cells in the perinatal Wnt4 mutant ovary. Dev. Biol. 2013, 383, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Tomaselli, S.; Megiorni, F.; Lin, L.; Mazzilli, M.C.; Gerrelli, D.; Majore, S.; Grammatico, P.; Achermann, J.C. Human RSPO1/R-spondin1 Is Expressed during Early Ovary Development and Augments beta-Catenin Signaling. PLoS ONE 2011, 6, e16366. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, J.V.; Kristensen, S.G.; Nielsen, M.E.; Humaidan, P.; Dal Canto, M.; Fadini, R.; Schmidt, K.T.; Ernst, E.; Yding Andersen, C. LH-receptor gene expression in human granulosa and cumulus cells from antral and preovulatory follicles. J. Clin. Endocrinol. Metab. 2012, 97, E1524–E1531. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Lane, M.; Thompson, J.G. Oocyte-secreted factors: Regulators of cumulus cell function and oocyte quality. Hum. Reprod. Update 2008, 14, 159–177. [Google Scholar] [CrossRef]
- Huang, Z.; Wells, D. The human oocyte and cumulus cells relationship: New insights from the cumulus cell transcriptome. Mol. Hum. Reprod. 2010, 16, 715–725. [Google Scholar] [CrossRef]
- Biason-Lauber, A.; Konrad, D.; Navratil, F.; Schoenle, E.J. A WNT4 mutation associated with Mullerian-duct regression and virilization in a 46,XX woman. N. Engl. J. Med. 2004, 351, 792–798. [Google Scholar] [CrossRef]
- Meyers-Wallen, V.N. Gonadal and Sex Differentiation Abnormalities of Dogs and Cats. Sex. Dev. 2012, 6, 46–60. [Google Scholar] [CrossRef]
- Parma, P.; Veyrunes, F.; Pailhoux, E. Sex Reversal in Non-Human Placental Mammals. Sex. Dev. 2016, 10, 326–344. [Google Scholar] [CrossRef]
- Lee, P.A.; Nordenström, A.; Houk, C.P.; Ahmed, S.F.; Auchus, R.; Baratz, A.; Baratz Dalke, K.; Liao, L.M.; Lin-Su, K.; Looijenga 3rd, L.H.J.; et al. Global disorders of sex development update since 2006: Perceptions, approach and care. Horm. Res. Paediat. 2016, 86, 158–180. [Google Scholar] [CrossRef]
- Eid, W.; Biason-Lauber, A. Why boys will be boys and girls will be girls: Human sex development and its defects. Birth Defects Res. C 2016, 108, 365–379. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Achermann, J.C.; Arlt, W.; Balen, A.; Conway, G.; Edwards, Z.; Elford, S.; Hughes, I.A.; Izatt, L.; Krone, N.; et al. Society for Endocrinology UK guidance on the initial evaluation of an infant or an adolescent with a suspected disorder of sex development (Revised 2015). Clin. Endocrinol. 2016, 84, 771–788. [Google Scholar] [CrossRef] [PubMed]
- Edelsztein, N.Y.; Grinspon, R.P.; Schteingart, H.F.; Rey, R.A. Anti-Mullerian hormone as a marker of steroid and gonadotropin action in the testis of children and adolescents with disorders of the gonadal axis. Int. J. Pediatr. Endocrinol. 2016, 2016, 20. [Google Scholar] [CrossRef]
- Crisponi, L.; Deiana, M.; Loi, A.; Chiappe, F.; Uda, M.; Amati, P.; Bisceglia, L.; Zelante, L.; Nagaraja, R.; Porcu, S.; et al. The putative forkhead transcription factor FOXL2 is mutated in blepharophimosis/ptosis/epicanthus inversus syndrome. Nat. Genet. 2001, 27, 159–166. [Google Scholar] [CrossRef]
- Parma, P.; Radi, O.; Vidal, V.; Chaboissier, M.C.; Dellambra, E.; Valentini, S.; Guerra, L.; Schedl, A.; Camerino, G. R-spondin1 is essential in sex determination, skin differentiation and malignancy. Nat. Genet. 2006, 38, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Tomaselli, S.; Megiorni, F.; De Bernardo, C.; Felici, A.; Marrocco, G.; Maggiulli, G.; Grammatico, B.; Remotti, D.; Saccucci, P.; Valentini, F.; et al. Syndromic true hermaphroditism due to an R-spondin1 (RSPO1) homozygous mutation. Hum. Mutat. 2008, 29, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Mather, J.P. Establishment and characterization of two distinct mouse testicular epithelial cell lines. Biol. Reprod. 1980, 23, 243–252. [Google Scholar] [CrossRef]
- Paquis-Flucklinger, V.; Michiels, J.F.; Vidal, F.; Alquier, C.; Pointis, G.; Bourdon, V.; Cuzin, F.; Rassoulzadegan, M. Expression in transgenic mice of the large T antigen of polyomavirus induces Sertoli cell tumours and allows the establishment of differentiated cell lines. Oncogene 1993, 8, 2087–2094. [Google Scholar]
- Larney, C.; Bailey, T.L.; Koopman, P. Switching on sex: Transcriptional regulation of the testis-determining gene Sry. Development 2014, 141, 2195–2205. [Google Scholar] [CrossRef]
- McClelland, K.; Bowles, J.; Koopman, P. Male sex determination: Insights into molecular mechanisms. Asian J. Androl. 2012, 14, 164–171. [Google Scholar] [CrossRef]
- Warr, N.; Bogani, D.; Siggers, P.; Brixey, R.; Tateossian, H.; Dopplapudi, A.; Wells, S.; Cheeseman, M.; Xia, Y.; Ostrer, H.; et al. Minor abnormalities of testis development in mice lacking the gene encoding the MAPK signalling component, MAP3K1. PLoS ONE 2011, 6, e19572. [Google Scholar] [CrossRef] [PubMed]
- Chui, K.; Trivedi, A.; Cheng, C.Y.; Cherbavaz, D.B.; Dazin, P.F.; Ai, L.T.H.; Mitchell, J.B.; Rabinovich, G.A.; Noble-Haeusslein, L.J.; John, C.M. Characterization and Functionality of Proliferative Human Sertoli Cells. Cell Transplant. 2011, 20, 619–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, E.A.; Barten-van Rijbroek, A.D.; Kal, H.B.; Sadri-Ardekani, H.; Mizrak, S.C.; van Pelt, A.M.; de Rooij, D.G. Proliferative activity in vitro and DNA repair indicate that adult mouse and human Sertoli cells are not terminally differentiated, quiescent cells. Biol. Reprod. 2009, 80, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Hai, Y.; Yao, C.; Chen, Z.; Hou, J.; Li, Z.; He, Z. Long-term culture and significant expansion of human Sertoli cells whilst maintaining stable global phenotype and AKT and SMAD1/5 activation. Cell Commun. Signal. 2015, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Andrews, P.W.; Damjanov, I.; Simon, D.; Banting, G.S.; Carlin, C.; Dracopoli, N.C.; Fogh, J. Pluripotent embryonal carcinoma clones derived from the human teratocarcinoma cell line Tera-2. Differentiation in vivo and in vitro. Lab. Invest. 1984, 50, 147–162. [Google Scholar] [PubMed]
- De Santa Barbara, P.; Bonneaud, N.; Boizet, B.; Desclozeaux, M.; Moniot, B.; Sudbeck, P.; Scherer, G.; Poulat, F.; Berta, P. Direct interaction of SRY-related protein SOX9 and steroidogenic factor 1 regulates transcription of the human anti-Mullerian hormone gene. Mol. Cell Biol. 1998, 18, 6653–6665. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Gutierrez, D.; Eid, W.; Biason-Lauber, A. A Human Gonadal Cell Model From Induced Pluripotent Stem Cells. Front. Genet. 2018, 9, 498. [Google Scholar] [CrossRef]
- Alexopoulos, E.; Shahid, J.; Ongley, H.Z.; Richardson, M.C. Luteinized human granulosa cells are associated with endogenous basement membrane-like components in culture. Mol. Hum. Reprod. 2000, 6, 324–330. [Google Scholar] [CrossRef]
- Bruckova, L.; Soukup, T.; Visek, B.; Moos, J.; Moosova, M.; Pavelkova, J.; Rezabek, K.; Kucerova, L.; Micuda, S.; Brcakova, E.; et al. Proliferative potential and phenotypic analysis of long-term cultivated human granulosa cells initiated by addition of follicular fluid. J. Assist. Reprod. Genet. 2011, 28, 939–950. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.L.; Kendall, J.Z.; Dandekar, P.V.; Quigley, M.M.; Schmidt, K.L. Characterization of long-term monolayer cultures of human granulosa cells from follicles of different size and exposed in vivo to clomiphene citrate and hCG. J. Reprod. Fertil. 1984, 71, 279–287. [Google Scholar] [CrossRef]
- Lie, B.L.; Leung, E.; Leung, P.C.; Auersperg, N. Long-term growth and steroidogenic potential of human granulosa-lutein cells immortalized with SV40 large T antigen. Mol. Cell Endocrinol. 1996, 120, 169–176. [Google Scholar] [CrossRef]
- Bruckova, L.; Soukup, T.; Moos, J.; Moosova, M.; Pavelkova, J.; Rezabek, K.; Visek, B.; Mokry, J. The cultivation of human granulosa cells. Acta Med. (Hradec Kral.) 2008, 51, 165–172. [Google Scholar] [CrossRef]
- Nishi, Y.; Yanase, T.; Mu, Y.; Oba, K.; Ichino, I.; Saito, M.; Nomura, M.; Mukasa, C.; Okabe, T.; Goto, K.; et al. Establishment and characterization of a steroidogenic human granulosa-like tumor cell line, KGN, that expresses functional follicle-stimulating hormone receptor. Endocrinology 2001, 142, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg-Bakker, C.A.; Hagemeijer, A.; Franken-Postma, E.M.; Smit, V.T.; Kuppen, P.J.; van Ravenswaay Claasen, H.H.; Cornelisse, C.J.; Schrier, P.I. Establishment and characterization of 7 ovarian carcinoma cell lines and one granulosa tumor cell line: Growth features and cytogenetics. Int. J. Cancer 1993, 53, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Klausen, C.; Leung, P.C. Overexpression of wild-type but not C134W mutant FOXL2 enhances GnRH-induced cell apoptosis by increasing GnRH receptor expression in human granulosa cell tumors. PLoS ONE 2013, 8, e55099. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, S.; Butzow, R.; Andersson, N.; Alexiadis, M.; Unkila-Kallio, L.; Heikinheimo, M.; Fuller, P.J.; Anttonen, M. The FOXL2 C134W mutation is characteristic of adult granulosa cell tumors of the ovary. Mod. Pathol. 2010, 23, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, K.; Dantes, A.; Schere-Levy, C.; Barash, A.; Yoshida, Y.; Kotsuji, F.; Vlodavsky, I.; Amsterdam, A. Induction of Ad4BP/SF-1, steroidogenic acute regulatory protein, and cytochrome P450scc enzyme system expression in newly established human granulosa cell lines. Endocrinology 1998, 139, 4679–4687. [Google Scholar] [CrossRef]
- Rainey, W.H.; Sawetawan, C.; Shay, J.W.; Michael, M.D.; Mathis, J.M.; Kutteh, W.; Byrd, W.; Carr, B.R. Transformation of Human Granulosa-Cells with the E6 and E7 Regions of Human Papillomavirus. J. Clin. Endocr. Metab. 1994, 78, 705–710. [Google Scholar] [CrossRef]
- Iwase, A.; Kiyono, T.; Takikawa, S.; Goto, M.; Nakamura, T.; Nagatomo, Y.; Nakahara, T.; Kotani, T.; Kobayashi, H.; Kondo, M.; et al. Establishment of a human nonluteinized granulosa cell line that transitions from the gonadotropin-independent to the gonadotropin-dependent status. Endocrinology 2012, 153, 2851–2860. [Google Scholar] [CrossRef]
- Di Pasquale, E.; Rossetti, R.; Marozzi, A.; Bodega, B.; Borgato, S.; Cavallo, L.; Einaudi, S.; Radetti, G.; Russo, G.; Sacco, M.; et al. Identification of new variants of human BMP15 gene in a large cohort of women with premature ovarian failure. J. Clin. Endocrinol. Metab. 2006, 91, 1976–1979. [Google Scholar] [CrossRef]
- Galloway, S.M.; McNatty, K.P.; Cambridge, L.M.; Laitinen, M.P.; Juengel, J.L.; Jokiranta, T.S.; McLaren, R.J.; Luiro, K.; Dodds, K.G.; Montgomery, G.W.; et al. Mutations in an oocyte-derived growth factor gene (BMP15) cause increased ovulation rate and infertility in a dosage-sensitive manner. Nat. Genet. 2000, 25, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Wang, P.; DeMayo, J.; DeMayo, F.J.; Elvin, J.A.; Carino, C.; Prasad, S.V.; Skinner, S.S.; Dunbar, B.S.; Dube, J.L.; et al. Synergistic roles of bone morphogenetic protein 15 and growth differentiation factor 9 in ovarian function. Mol. Endocrinol. 2001, 15, 854–866. [Google Scholar] [CrossRef] [PubMed]
- Lesch, B.J.; Page, D.C. Genetics of germ cell development. Nat. Rev. Genet. 2012, 13, 781–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irie, N.; Tang, W.W.; Azim Surani, M. Germ cell specification and pluripotency in mammals: A perspective from early embryogenesis. Reprod. Med. Biol. 2014, 13, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Irie, N.; Weinberger, L.; Tang, W.W.C.; Kobayashi, T.; Viukov, S.; Manor, Y.S.; Dietmann, S.; Hanna, J.H.; Surani, M.A. SOX17 Is a Critical Specifier of Human Primordial Germ Cell Fate. Cell 2015, 160, 253–268. [Google Scholar] [CrossRef]
- Saitou, M.; Yamaji, M. Germ cell specification in mice: Signaling, transcription regulation, and epigenetic consequences. Reproduction 2010, 139, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, M.; Seki, Y.; Kurimoto, K.; Yabuta, Y.; Yuasa, M.; Shigeta, M.; Yamanaka, K.; Ohinata, Y.; Saitou, M. Critical function of Prdm14 for the establishment of the germ cell lineage in mice. Nat. Genet. 2008, 40, 1016–1022. [Google Scholar] [CrossRef]
- Geijsen, N.; Horoschak, M.; Kim, K.; Gribnau, J.; Eggan, K.; Daley, G.Q. Derivation of embryonic germ cells and male gametes from embryonic stem cells. Nature 2004, 427, 148–154. [Google Scholar] [CrossRef]
- Ko, K.; Huebner, K.; Mueller-Keuker, J.; Schoeler, H.R. In vitro derivation of germ cells from embryonic stem cells. Front. Biosci. 2010, 15, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, E.; Forabosco, A.; Schlessinger, D. Germ cell formation from embryonic stem cells and the use of somatic cell nuclei in oocytes. Ann. N. Y. Acad. Sci. 2011, 1221, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Toyooka, Y.; Tsunekawa, N.; Akasu, R.; Noce, T. Embryonic stem cells can form germ cells in vitro. Proc. Natl. Acad Sci. USA 2003, 100, 11457–11462. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.B.; Meng, Q.G.; Li, N. In Vitro Derivation of Germ Cells From Embryonic Stem Cells in Mammals. Mol. Reprod. Dev. 2010, 77, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Gurdon, J.B. The developmental capacity of nuclei taken from intestinal epithelium cells of feeding tadpoles. J. Embryol. Exp. Morphol. 1962, 10, 622–640. [Google Scholar] [PubMed]
- Tada, M.; Takahama, Y.; Abe, K.; Nakatsuji, N.; Tada, T. Nuclear reprogramming of somatic cells by in vitro hybridization with ES cells. Curr. Biol. 2001, 11, 1553–1558. [Google Scholar] [CrossRef] [Green Version]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810–813. [Google Scholar] [CrossRef]
- Davis, R.L.; Weintraub, H.; Lassar, A.B. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell 1987, 51, 987–1000. [Google Scholar] [CrossRef]
- Schneuwly, S.; Klemenz, R.; Gehring, W.J. Redesigning the body plan of Drosophila by ectopic expression of the homoeotic gene Antennapedia. Nature 1987, 325, 816–818. [Google Scholar] [CrossRef]
- Yamanaka, S.; Blau, H.M. Nuclear reprogramming to a pluripotent state by three approaches. Nature 2010, 465, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef]
- Smith, A.G.; Heath, J.K.; Donaldson, D.D.; Wong, G.G.; Moreau, J.; Stahl, M.; Rogers, D. Inhibition of pluripotential embryonic stem cell differentiation by purified polypeptides. Nature 1988, 336, 688–690. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Puri, M.C.; Nagy, A. Concise review: Embryonic stem cells versus induced pluripotent stem cells: The game is on. Stem Cells 2012, 30, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Aoi, T.; Yae, K.; Nakagawa, M.; Ichisaka, T.; Okita, K.; Takahashi, K.; Chiba, T.; Yamanaka, S. Generation of pluripotent stem cells from adult mouse liver and stomach cells. Science 2008, 321, 699–702. [Google Scholar] [CrossRef] [PubMed]
- Wernig, M.; Meissner, A.; Cassady, J.P.; Jaenisch, R. c-Myc is dispensable for direct reprogramming of mouse fibroblasts. Cell Stem Cell 2008, 2, 10–12. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Kim, T.K.; Eberwine, J.H. Mammalian cell transfection: The present and the future. Anal. Bioanal. Chem. 2010, 397, 3173–3178. [Google Scholar] [CrossRef]
- Yu, J.Y.; Hu, K.J.; Smuga-Otto, K.; Tian, S.L.; Stewart, R.; Slukvin, I.I.; Thomson, J.A. Human Induced Pluripotent Stem Cells Free of Vector and Transgene Sequences. Science 2009, 324, 797–801. [Google Scholar] [CrossRef] [Green Version]
- Warren, L.; Manos, P.D.; Ahfeldt, T.; Loh, Y.H.; Li, H.; Lau, F.; Ebina, W.; Mandal, P.K.; Smith, Z.D.; Meissner, A.; et al. Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. Cell Stem Cell 2010, 7, 618–630. [Google Scholar] [CrossRef]
- Fusaki, N.; Ban, H.; Nishiyama, A.; Saeki, K.; Hasegawa, M. Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome. Proc. Jpn. Acad. Jpn. Ser. B 2009, 85, 348–362. [Google Scholar] [CrossRef] [Green Version]
- Bang, J.S.; Choi, N.Y.; Lee, M.; Ko, K.; Lee, H.J.; Park, Y.S.; Jeong, D.; Chung, H.M.; Ko, K. Optimization of episomal reprogramming for generation of human induced pluripotent stem cells from fibroblasts. Anim. Cells Syst. 2018, 22, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, P.K.; Rossi, D.J. Reprogramming human fibroblasts to pluripotency using modified mRNA. Nat. Protoc. 2013, 8, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Wang, Q. Efficient Generation of Non-Integration and Feeder-Free Induced Pluripotent Stem Cells from Human Peripheral Blood Cells by Sendai Virus. Cell Physiol. Biochem. 2018, 50, 1318–1331. [Google Scholar] [CrossRef] [PubMed]
- Nefzger, C.M.; Rossello, F.J.; Chen, J.; Liu, X.D.; Knaupp, A.S.; Firas, J.; Paynter, J.M.; Pflueger, J.; Buckberry, S.; Lim, S.M.; et al. Cell Type of Origin Dictates the Route to Pluripotency. Cell Rep. 2017, 21, 2649–2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, A. DNA methylation patterns and epigenetic memory. Gene Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, Z.; Wilson, K.D.; Wu, Y.; Hu, S.J.; Quertermous, T.; Wu, J.C. Persistent Donor Cell Gene Expression among Human Induced Pluripotent Stem Cells Contributes to Differences with Human Embryonic Stem Cells. PLoS ONE 2010, 5, e8975. [Google Scholar] [CrossRef] [PubMed]
- Polo, J.M.; Liu, S.; Figueroa, M.E.; Kulalert, W.; Eminli, S.; Tan, K.Y.; Apostolou, E.; Stadtfeld, M.; Li, Y.; Shioda, T.; et al. Cell type of origin influences the molecular and functional properties of mouse induced pluripotent stem cells. Nat. Biotechnol. 2010, 28, 848–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panopoulos, A.D.; Yanes, O.; Ruiz, S.; Kida, Y.S.; Diep, D.; Tautenhahn, R.; Herrerias, A.; Batchelder, E.M.; Plongthongkum, N.; Lutz, M.; et al. The metabolome of induced pluripotent stem cells reveals metabolic changes occurring in somatic cell reprogramming. Cell Res. 2012, 22, 168–177. [Google Scholar] [CrossRef]
- Spyrou, J.; Gardner, D.K.; Harvey, A.J. Metabolism Is a Key Regulator of Induced Pluripotent Stem Cell Reprogramming. Stem Cells Int. 2019, 2019. [Google Scholar] [CrossRef]
- Setthawong, P.; Phakdeedindan, P.; Tiptanavattana, N.; Rungarunlert, S.; Techakumphu, M.; Tharasanit, T. Generation of porcine induced-pluripotent stem cells from Sertoli cells. Theriogenology 2019, 127, 32–40. [Google Scholar] [CrossRef]
- Loh, Y.H.; Hartung, O.; Li, H.; Guo, C.G.; Sahalie, J.M.; Manos, P.D.; Urbach, A.; Heffner, G.C.; Grskovic, M.; Vigneault, F.; et al. Reprogramming of T Cells from Human Peripheral Blood. Cell Stem Cell 2010, 7, 15–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Babot, G.; Balyura, M.; Hadjidemetriou, I.; Ajodha, S.J.; Taylor, D.R.; Ghataore, L.; Taylor, N.F.; Schubert, U.; Ziegler, C.G.; Storr, H.L.; et al. Modeling Congenital Adrenal Hyperplasia and Testing Interventions for Adrenal Insufficiency Using Donor-Specific Reprogrammed Cells. Cell Rep. 2018, 22, 1236–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Benda, C.; Duzinger, S.; Huang, Y.H.; Li, X.Y.; Li, Y.H.; Guo, X.P.; Cao, G.K.; Chen, S.; Hao, L.L.; et al. Generation of Induced Pluripotent Stem Cells from Urine. J. Am. Soc. Nephrol. 2011, 22, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Marti, M.; Mulero, L.; Pardo, C.; Morera, C.; Carrio, M.; Laricchia-Robbio, L.; Esteban, C.R.; Izpisua Belmonte, J.C. Characterization of pluripotent stem cells. Nat. Protoc. 2013, 8, 223–253. [Google Scholar] [CrossRef]
- Sekine, K.; Takebe, T.; Suzuki, Y.; Kamiya, A.; Nakauchi, H.; Taniguchi, H. Highly Efficient Generation of Definitive Endoderm Lineage from Human Induced Pluripotent Stem Cells. Transpl. Proc. 2012, 44, 1127–1129. [Google Scholar] [CrossRef]
- Terryn, J.; Tricot, T.; Gajjar, M.; Verfaillie, C. Recent advances in lineage differentiation from stem cells: Hurdles and opportunities? F1000Res 2018, 7, 220. [Google Scholar] [CrossRef]
- Bucay, N.; Yebra, M.; Cirulli, V.; Afrikanova, I.; Kaido, T.; Hayek, A.; Montgomery, A.M.P. A Novel Approach for the Derivation of Putative Primordial Germ Cells and Sertoli Cells from Human Embryonic Stem Cells. Stem Cells 2009, 27, 68–77. [Google Scholar] [CrossRef]
- Shlush, E.; Maghen, L.; Swanson, S.; Kenigsberg, S.; Moskovtsev, S.; Barretto, T.; Gauthier-Fisher, A.; Librach, C.L. In vitro generation of Sertoli-like and haploid spermatid-like cells from human umbilical cord perivascular cells. Stem Cell Res. Ther. 2017, 8, 37. [Google Scholar] [CrossRef]
- Anchan, R.; Gerami-Naini, B.; Lindsey, J.S.; Ho, J.W.K.; Kiezun, A.; Lipskind, S.; Ng, N.; LiCausi, J.A.; Kim, C.S.; Brezina, P.; et al. Efficient Differentiation of Steroidogenic and Germ-Like Cells from Epigenetically-Related iPSCs Derived from Ovarian Granulosa Cells. PLoS ONE 2015, 10, e0119275. [Google Scholar] [CrossRef]
- Liu, T.; Li, Q.; Wang, S.; Chen, C.; Zheng, J. Transplantation of ovarian granulosalike cells derived from human induced pluripotent stem cells for the treatment of murine premature ovarian failure. Mol. Med. Rep. 2016, 13, 5053–5058. [Google Scholar] [CrossRef]
- Lipskind, S.; Lindsey, J.S.; Gerami-Naini, B.; Eaton, J.L.; O’Connell, D.; Kiezun, A.; Ho, J.W.K.; Ng, N.; Parasar, P.; Ng, M.; et al. An Embryonic and Induced Pluripotent Stem Cell Model for Ovarian Granulosa Cell Development and Steroidogenesis. Reprod. Sci. 2018, 25, 712–726. [Google Scholar] [CrossRef] [PubMed]
- Kee, K.; Angeles, V.T.; Flores, M.; Nguyen, H.N.; Pera, R.A.R. Human DAZL, DAZ and BOULE genes modulate primordial germ-cell and haploid gamete formation. Nature 2009, 462, U222–U295. [Google Scholar] [CrossRef] [PubMed]
- Eguizabal, C.; Montserrat, N.; Vassena, R.; Barragan, M.; Garreta, E.; Garcia-Quevedo, L.; Vidal, F.; Giorgetti, A.; Veiga, A.; Belmonte, J.C.I. Complete Meiosis from Human Induced Pluripotent Stem Cells. Stem Cells 2011, 29, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Medrano, J.V.; Ramathal, C.; Nguyen, H.N.; Simon, C.; Pera, R.A.R. Divergent RNA-Binding Proteins, DAZL and VASA, Induce Meiotic Progression in Human Germ Cells Derived In Vitro. Stem Cells 2012, 30, 441–451. [Google Scholar] [CrossRef]
- Yang, S.; Ding, S.F.; He, S.W.; He, L.X.; Gao, K.F.; Peng, S.P.; Shuai, C.J. Differentiation of primordial germ cells from premature ovarian insufficiency-derived induced pluripotent stem cells. Stem Cell Res. Ther. 2019, 10, 156. [Google Scholar] [CrossRef]
- Panula, S.; Medrano, J.V.; Kee, K.; Bergstrom, R.; Ha, N.N.; Byers, B.; Wilson, K.D.; Wu, J.C.; Simon, C.; Hovatta, O.; et al. Human germ cell differentiation from fetal- and adult-derived induced pluripotent stem cells. Hum. Mol. Genet. 2011, 20, 752–762. [Google Scholar] [CrossRef]
- Easley, C.A.; Phillips, B.T.; McGuire, M.M.; Barringer, J.M.; Valli, H.; Hermann, B.P.; Simerly, C.R.; Rajkovic, A.; Miki, T.; Orwig, K.E.; et al. Direct Differentiation of Human Pluripotent Stem Cells into Haploid Spermatogenic Cells. Cell Rep. 2012, 2, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Jung, D.J.; Xiong, J.; Ye, M.; Qin, X.S.; Li, L.; Cheng, S.F.; Luo, M.Y.; Peng, J.; Dong, J.; Tang, F.C.; et al. In vitro differentiation of human embryonic stem cells into ovarian follicle-like cells. Nat. Commun. 2017, 8, 15680. [Google Scholar] [CrossRef]
- Buganim, Y.; Itskovich, E.; Hu, Y.C.; Cheng, A.W.; Ganz, K.; Sarkar, S.; Fu, D.D.; Welstead, G.G.; Page, D.C.; Jaenisch, R. Direct Reprogramming of Fibroblasts into Embryonic Sertoli-like Cells by Defined Factors. Cell Stem Cell 2012, 11, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Evseenko, D.; Zhu, Y.H.; Schenke-Layland, K.; Kuo, J.; Latour, B.; Ge, S.D.; Scholes, J.; Dravid, G.; Li, X.M.; MacLellan, W.R.; et al. Mapping the first stages of mesoderm commitment during differentiation of human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2010, 107, 13742–13747. [Google Scholar] [CrossRef] [Green Version]
- Lawson, K.A.; Dunn, N.R.; Roelen, B.A.J.; Zeinstra, L.M.; Davis, A.M.; Wright, C.V.E.; Korving, J.P.W.F.M.; Hogan, B.L.M. Bmp4 is required for the generation of primordial germ cells in the mouse embryo. Gene Dev. 1999, 13, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Kan, M.; McKeehan, W.L.; de Crombrugghe, B. Up-regulation of the chondrogenic Sox9 gene by fibroblast growth factors is mediated by the mitogen-activated protein kinase pathway. Proc. Natl. Acad. Sci. USA 2000, 97, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kobayashi, A.; Sekido, R.; DiNapoli, L.; Brennan, J.; Chaboissier, M.C.; Poulat, F.; Behringer, R.R.; Lovell-Badge, R.; Capel, B. Fgf9 and Wnt4 act as antagonistic signals to regulate mammalian sex determination. PLoS Biol. 2006, 4, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Moniot, B.; Declosmenil, F.; Barrionuevo, F.; Scherer, G.; Aritake, K.; Malki, S.; Marzi, L.; Cohen-Solal, A.; Georg, I.; Klattig, J.; et al. The PGD2 pathway, independently of FGF9, amplifies SOX9 activity in Sertoli cells during male sexual differentiation. Development 2009, 136, 1813–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anamthathmakula, P.; Miryala, C.S.J.; Moreci, R.S.; Kyathanahalli, C.; Hassan, S.S.; Condon, J.C.; Jeyasuria, P. Steroidogenic Factor 1 (Nr5a1) is Required for Sertoli Cell Survival Post Sex Determination. Sci. Rep. 2019, 9, 4452. [Google Scholar] [CrossRef]
- Nicholls, P.K.; Stanton, P.G.; Chen, J.L.; Olcorn, J.S.; Haverfield, J.T.; Qian, H.; Walton, K.L.; Gregorevic, P.; Harrison, C.A. Activin signaling regulates Sertoli cell differentiation and function. Endocrinology 2012, 153, 6065–6077. [Google Scholar] [CrossRef]
- Kang, Y.; Cheng, M.J.; Xu, C.J. Secretion of oestrogen from murine-induced pluripotent stem cells co-cultured with ovarian granulosa cells in vitro. Cell Biol Int 2011, 35, 871–874. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Wu, Z.; Tan, X.J.; Liu, F.Y.; Huang, X.H.; Fang, X.L. Differentiation of rat iPS cells and ES cells into granulosa cell-like cells in vitro. Acta. Bioch. Bioph. Sin. 2013, 45, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Hubner, K.; Fuhrmann, G.; Christenson, L.K.; Kehler, J.; Reinbold, R.; De La Fuente, R.; Wood, J.; Strauss, J.F.; Boiani, M.; Scholer, H.R. Derivation of oocytes from mouse embryonic stem cells. Science 2003, 300, 1251–1256. [Google Scholar] [CrossRef]
- Nakaki, F.; Hayashi, K.; Ohta, H.; Kurimoto, K.; Yabuta, Y.; Saitou, M. Induction of mouse germ-cell fate by transcription factors in vitro. Nature 2013, 501, 222. [Google Scholar] [CrossRef]
- Tang, W.W.C.; Dietmann, S.; Irie, N.; Leitch, H.G.; Floros, V.I.; Bradshaw, C.R.; Hackett, J.A.; Chinnery, P.F.; Surani, M.A. A Unique Gene Regulatory Network Resets the Human Germline Epigenome for Development. Cell 2015, 161, 1453–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M.; Kawaguchi, T.; Durcova-Hills, G.; Imai, H. Generation of germ cells from pluripotent stem cells in mammals. Reprod. Med. Biol. 2018, 17, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Eirin-Lopez, J.M.; Ausio, J. Boule and the Evolutionary Origin of Metazoan Gametogenesis: A Grandpa’s Tale. Int. J. Evol. Biol. 2011, 2011, 972457. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.Y.; Moore, F.L.; Pera, R.A.R. A gene family required for human germ cell development evolved from an ancient meiotic gene conserved in metazoans. Proc. Natl. Acad. Sci. USA 2001, 98, 7414–7419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panula, S.; Reda, A.; Stukenborg, J.B.; Ramathal, C.; Sukhwani, M.; Albalushi, H.; Edsgard, D.; Nakamura, M.; Soder, O.; Orwig, K.E.; et al. Over Expression of NANOS3 and DAZL in Human Embryonic Stem Cells. PLoS ONE 2016, 11, e0165268. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Ye, S.C.; Liang, D.L.; Wang, P.X.; Fu, J.; Ma, Q.; Kong, R.J.; Shi, L.H.; Gong, X.P.; Chen, W.; et al. In Vitro Modeling of Human Germ Cell Development Using Pluripotent Stem Cells. Stem Cell Rep. 2018, 10, 509–523. [Google Scholar] [CrossRef] [Green Version]
- Kee, K.; Gonsalves, J.M.; Clark, A.T.; Pera, R.A.R. Bone morphogenetic proteins induce germ cell differentiation from human embryonic stem cells. Stem Cells Dev. 2006, 15, 831–837. [Google Scholar] [CrossRef]
- Hurrell, T.; Segeritz, C.P.; Vallier, L.; Lilley, K.S.; Cromarty, A.D. A proteomic time course through the differentiation of human induced pluripotent stem cells into hepatocyte-like cells. Sci. Rep. 2019, 9, 3270. [Google Scholar] [CrossRef]
- Ben Jehuda, R.; Shemer, Y.; Binah, O. Genome Editing in Induced Pluripotent Stem Cells using CRISPR/Cas9. Stem Cell Rev. Rep. 2018, 14, 323–336. [Google Scholar] [CrossRef]
- Chang, C.Y.; Ting, H.C.; Su, H.L.; Jeng, J.R. Combining Induced Pluripotent Stem Cells and Genome Editing Technologies for Clinical Applications. Cell Transplant. 2018, 27, 379–392. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, E.; Chen, T. A matter of life and death: Self-renewal in stem cells. Embo. Rep. 2013, 14, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Pennarossa, G.; Santoro, R.; Manzoni, E.F.M.; Pesce, M.; Gandolfi, F.; Brevini, T.A.L. Epigenetic Erasing and Pancreatic Differentiation of Dermal Fibroblasts into Insulin-Producing Cells are Boosted by the Use of Low-Stiffness Substrate. Stem Cell Rev. Rep. 2018, 14, 398–411. [Google Scholar] [CrossRef] [PubMed]
- McCauley, H.A.; Wells, J.M. Pluripotent stem cell-derived organoids: Using principles of developmental biology to grow human tissues in a dish. Development 2017, 144, 958–962. [Google Scholar] [CrossRef] [PubMed]
- Laronda, M.M.; Burdette, J.E.; Kim, J.J.; Woodruff, T.K. Recreating the female reproductive tract in vitro using iPSC technology in a linked microfluidics environment. Stem Cell Res. Ther. 2013, 4, S13. [Google Scholar] [CrossRef] [PubMed]
- Meroni, S.B.; Galardo, M.N.; Rindone, G.; Gorga, A.; Riera, M.F.; Cigorraga, S.B. Molecular Mechanisms and Signaling Pathways Involved in Sertoli Cell Proliferation. Front. Endocrinol. 2019, 10, 224. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, J.Y.; Romero, M.M.; Smith, K.N.; Shea, L.D.; Woodruff, T.K. In vitro follicle growth supports human oocyte meiotic maturation. Sci Rep. 2015, 5, 17323. [Google Scholar] [CrossRef]
- Quaas, A.; Dokras, A. Diagnosis and treatment of unexplained infertility. Rev. Obs. Gynecol. 2008, 1, 69–76. [Google Scholar]
Figure 1.
Human sex determination and differentiation: Gonadal somatic supporters, Sertoli cells and granulosa cells, play a central role in sexual development, orchestrating the differentiation of the other gonadal cells and secreting SOX9 and forkhead box L2 (FOXL2). Both factors counteract each other in a delicate equilibrium and its expression must be maintained through different stages of development to keep the final sexual faith into males or females respectively.
Figure 1.
Human sex determination and differentiation: Gonadal somatic supporters, Sertoli cells and granulosa cells, play a central role in sexual development, orchestrating the differentiation of the other gonadal cells and secreting SOX9 and forkhead box L2 (FOXL2). Both factors counteract each other in a delicate equilibrium and its expression must be maintained through different stages of development to keep the final sexual faith into males or females respectively.

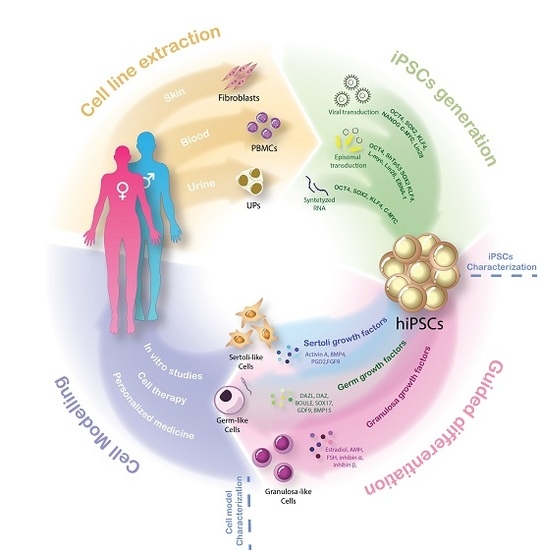
Figure 2.
Cellular reprograming and guided differentiation of hiPSCs: The scheme represents the four stages of the iPSCs-derived gonadal cell model generation for DSD patients. Isolation of fibroblasts, peripheral blood mononuclear cells or even urinary precursors directly from the patient have been considered as easy to obtain and patient-friendly cell sources. Up to date, three cellular reprogramming techniques are commonly used to generate hiPSCs from diverse cell sources: viral transduction of Yamanaka and Thomson reprograming factors; episomal transduction adding a EBNA1 factor and silencing expression of the TP53 gene; and synthetized RNA. Guided differentiation by expression specific factors can force transformation of cell into Sertoli-like (BMP4, Activin A, PDG2 and FGF9) Granulosa-like (Estradiol, AMH, FSH, inhibin α and inhibin β) or germ-like cells (DAZL, DAZ, BOULE, GDF9 and BMP15). Addition of both those factors into the culture medium and forced overexpression proved to induce differentiation of hiPSCs. Gonadal cell-like models directly derived from DSD patients suppose unique tools to perform patient-specific in vitro studies and open the door for more advances in cell therapy and personalized medicine for DSD patients. hiPSCs: human induced pluripotent stem cells, PBMCS: peripheral blood mononuclear cells, UPS: urinary progenitors.
Figure 2.
Cellular reprograming and guided differentiation of hiPSCs: The scheme represents the four stages of the iPSCs-derived gonadal cell model generation for DSD patients. Isolation of fibroblasts, peripheral blood mononuclear cells or even urinary precursors directly from the patient have been considered as easy to obtain and patient-friendly cell sources. Up to date, three cellular reprogramming techniques are commonly used to generate hiPSCs from diverse cell sources: viral transduction of Yamanaka and Thomson reprograming factors; episomal transduction adding a EBNA1 factor and silencing expression of the TP53 gene; and synthetized RNA. Guided differentiation by expression specific factors can force transformation of cell into Sertoli-like (BMP4, Activin A, PDG2 and FGF9) Granulosa-like (Estradiol, AMH, FSH, inhibin α and inhibin β) or germ-like cells (DAZL, DAZ, BOULE, GDF9 and BMP15). Addition of both those factors into the culture medium and forced overexpression proved to induce differentiation of hiPSCs. Gonadal cell-like models directly derived from DSD patients suppose unique tools to perform patient-specific in vitro studies and open the door for more advances in cell therapy and personalized medicine for DSD patients. hiPSCs: human induced pluripotent stem cells, PBMCS: peripheral blood mononuclear cells, UPS: urinary progenitors.

Figure 3.
Known factors involved in guided differentiation of hiPSCs into gonadal cells: Expression of growth factors can guide the differentiation of embryoids bodies into Sertoli, Granulosa or germ cell-like cells. Bone morphogenic protein 4 (BMP4), Activin A, prostaglandin 2 (PDG2) and fibroblast growth factor 9 (FGF9) and retinoid acid (RA) successfully differentiated into Sertoli-like cells are able to endogenously express SOX9, anti-Mullerian hormone (AMH), claudin-11 (CLDN11) and other Sertoli markers [37,98]. Granulosa-like cell differentiation has shown to be stimulated by estradiol, AMH, follicle stimulating hormone (FSH), inhibin α and inhibin β [109,110,111]. BMPs proteins together with GDNF, DAZL and BOULE are known to initiate differentiation into a meiotic primordial germ cell-like (PGCL) phenotype [55,102,103,104]. While complete spermatogenesis in human cells was not achieved yet, PGCs differentiated with SOX17, GDF9 and BMP15 factors showed the ability to induce folliculogenesis [108].
Figure 3.
Known factors involved in guided differentiation of hiPSCs into gonadal cells: Expression of growth factors can guide the differentiation of embryoids bodies into Sertoli, Granulosa or germ cell-like cells. Bone morphogenic protein 4 (BMP4), Activin A, prostaglandin 2 (PDG2) and fibroblast growth factor 9 (FGF9) and retinoid acid (RA) successfully differentiated into Sertoli-like cells are able to endogenously express SOX9, anti-Mullerian hormone (AMH), claudin-11 (CLDN11) and other Sertoli markers [37,98]. Granulosa-like cell differentiation has shown to be stimulated by estradiol, AMH, follicle stimulating hormone (FSH), inhibin α and inhibin β [109,110,111]. BMPs proteins together with GDNF, DAZL and BOULE are known to initiate differentiation into a meiotic primordial germ cell-like (PGCL) phenotype [55,102,103,104]. While complete spermatogenesis in human cells was not achieved yet, PGCs differentiated with SOX17, GDF9 and BMP15 factors showed the ability to induce folliculogenesis [108].


{kind=link}
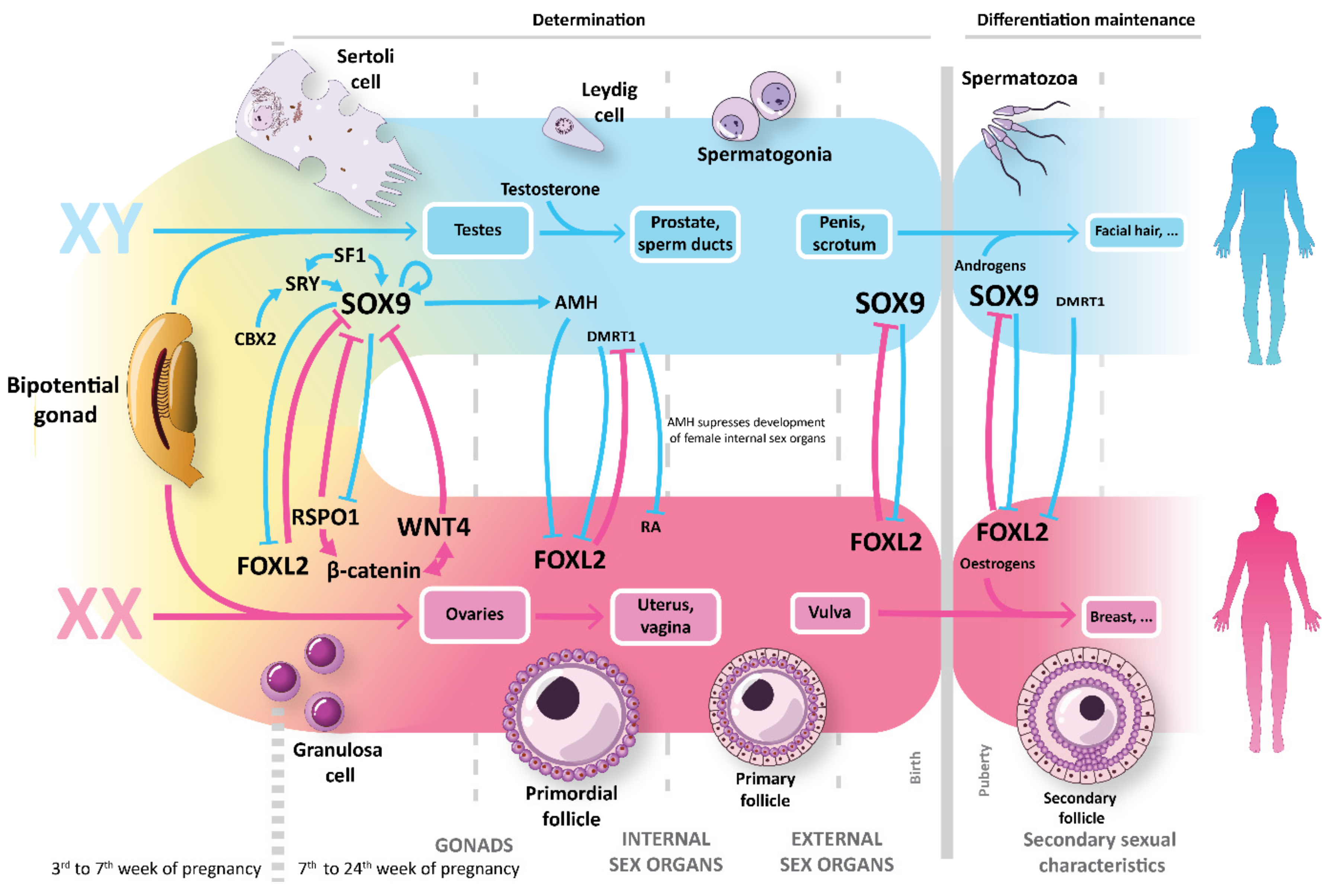{kind=link}
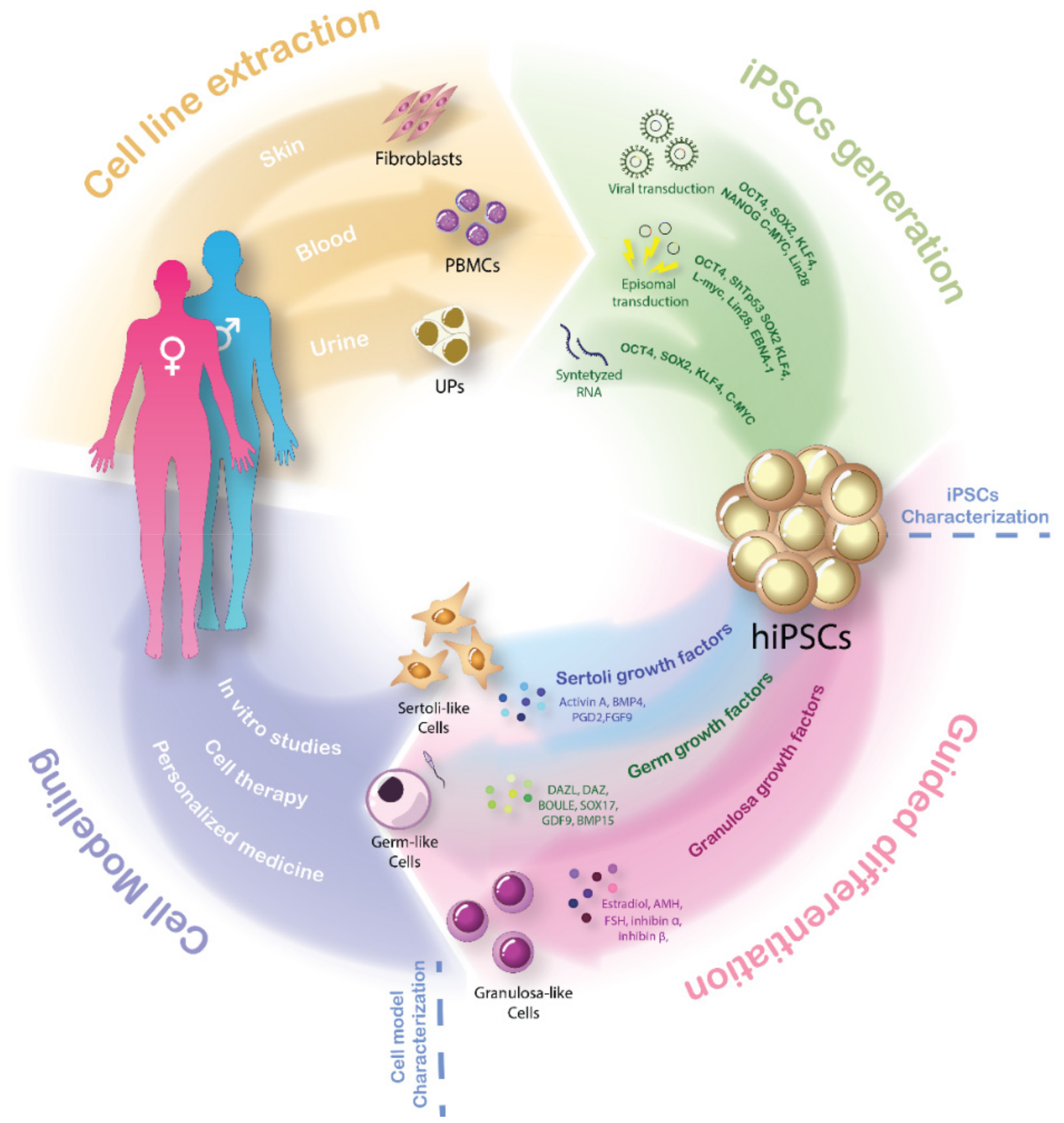{kind=link}
{kind=link}
Table 1.
Human PSCs-Derived Gonadal Cell Models.
| Cell Type | Cell Model | Differentiation Mechanism | Remarks | Reference |
|---|---|---|---|---|
| Sertoli cell | SLC | Unguided differentiation in co-culture with PGCs | Possible paracrine action of PGCs on differentiation of SLCs | Bucay et al., 2009 [97] |
| SLC | 5-step differentiation protocol, including RA, LIF, GDNF, putrescine, testosterone and FSH | BMP4 secretion by undifferentiated cells | Shlush et al., 2017 [98] | |
| SLC | Addition of BMP4, PGDS, bFGF, FGF9 and Activin-A to growth medium | Transcriptomic landscape resemble differentiating Sertoli cells | Rodríguez Gutiérrez et al., 2018 [37] | |
| Granulosa cell | Ovarian steroidogenic cells | Redifferentiation of iPSCs derived from GCs into homotypic ovarian steroidogenic cells | Cellular origin have strong effect on final differentiation faith | Anchan et al., 2015 [99] |
| GLC | Addition of all-trans-retinoic acid, estradiol, AMH, FSH, Inhibin α, Inhibin β and TGF –β to growth medium | GLCs can rescue ovarian failure when transplanted in POF mice. | Liu et al., 2016 [100] | |
| GLC | Differentiation from human amniocytes without intervention of additional growth factors. | GLC able to synthetize E2 | Lipskind et al., 2018 [101] | |
| Primordial germ cell | hPGCLCs | Differentiation induction by recombinant BMPs | BMPs induce differentiation of germ cells from hES cells | Kee et al., 2009 [102] |
| Haploid Gamete-like Cells | Two step protocol: Culture in bFGF-depleted ES cell media followed by addition of RA | Epigenetic memory of the reprogrammed somatic cells | Eguizabal et al.,2011 [103] | |
| Meiotic GCs | Overexpression of VASA and/or DAZL following differentiation on matrigel-coated plates | Differentiation of germ cells is dependent on post-translational regulation by RNA-binding proteins like VASA | Medrano et al., 2011 [104] | |
| hPGCLCs | Addition of BMP2 or BMP4, LIF, SCF, EGF and ROCK inhibitor to growth medium | SOX17 is the key regulator of hPGCLC specification | Irie et al., 2015 [55] | |
| hPGCLCs | (1) in vitro: same protocol as Irie et al., 2015 (2) in vivo: xenotransplantation into NOD/SCID mice | hPGCLCs had the potential for meiotic progression in vitro | Yang et al., 2019 [105] | |
| Spermatid | SpLC | Differentiation induction by BMPs followed by overexpression of DAZ, DAZL, and BOULE | hiPSC lines can differentiate to haploid cells with characteristic staining of ACROSIN for spermatid. | Panula et al., 2011 [106] |
| SSC | Direct differentiation using mouse spermatogonial stem cell (SSC) medium | hPSCs differentiate into spermatogonia, spermatocytes and haploid spermatids | Easley et al., 2012 [107] | |
| Oocyte | FLCs | Overexpression of DAZL and BOULE with recombinant human GDF9 and BMP15 | GDF9 and BMP15 induce ovarian follicle formation in hESCs | Jung et al.,2017 [108] |
FLCs: follicle-like cells, GLC: Granulosa-like cells, hPGCLCs: human primordial germ cell-like cells, SLC: Sertoli-like cells, SpLC: spermatid-like cells, SSC: spermatogenic stem cells.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rodríguez Gutiérrez, D.; Biason-Lauber, A. Pluripotent Cell Models for Gonadal Research. Int. J. Mol. Sci. 2019, 20, 5495. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215495
AMA Style
Rodríguez Gutiérrez D, Biason-Lauber A. Pluripotent Cell Models for Gonadal Research. International Journal of Molecular Sciences. 2019; 20(21):5495. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215495
Chicago/Turabian StyleRodríguez Gutiérrez, Daniel, and Anna Biason-Lauber. 2019. "Pluripotent Cell Models for Gonadal Research" International Journal of Molecular Sciences 20, no. 21: 5495. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215495
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.