Dimorphism of HLA-E and Its Disease Association

 , ,
, ,
Abstract
:1. Introduction
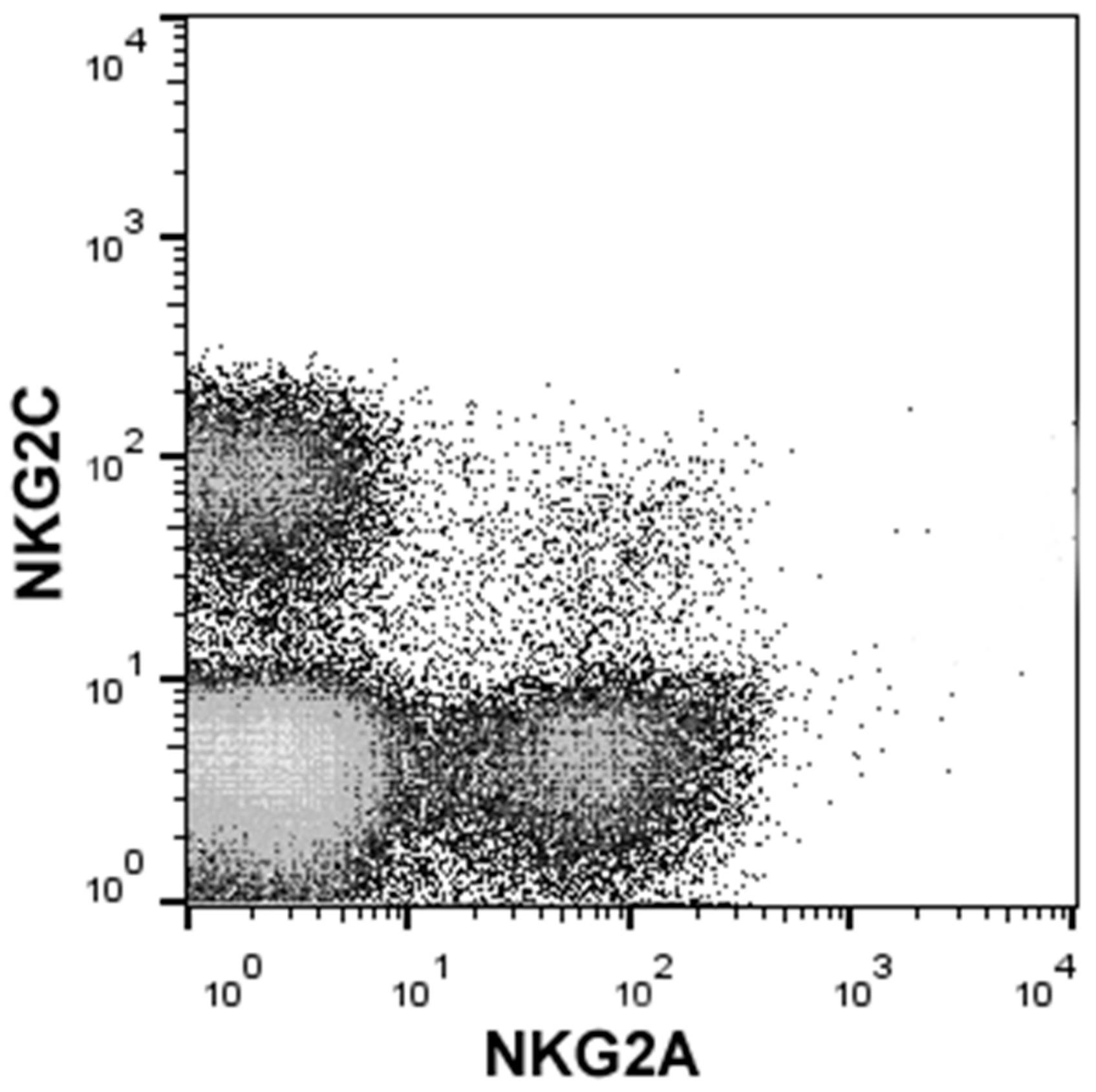
2. HLA-E and NKG2 Receptor Family
3. HLA-E-Peptide Complexes and TCR
4. Evolution and Polymorphism of HLA-E
5. Repertoire of Peptides Presented by HLA-E in Non-Typical Conditions
6. Soluble HLA-E and its Disease Associations
7. HLA-E Dimorphism and Viral Infections
8. HLA-E Dimorphism and Bacterial Infections
9. HLA-E Dimorphism and Cancer
10. HLA-E Dimorphism and Autoimmune Diseases
11. HLA-E Dimorphism and Transplantation
12. HLA-E and Pregnancy Disorders
13. Concluding Remarks
Funding
Conflicts of Interest
Abbreviations
| HCMV | Human Cytomegalovirus |
| HCV | Hepatitis C Virus |
| HIV | Human Immunodeficiency Virus |
| HLA | Human Leukocyte Antigen |
| HSC | Hematopoietic Stem Cell |
| MHC | Major Histocompatibility Complex |
| NKG2 | Natural Killer Group 2 |
References
- Olieslagers, T.I.; Voorter, C.E.M.; Groeneweg, M.; Xu, Y.; Wieten, L.; Tilanus, M.G.J. New insights in HLA-E polymorphism by refined analysis of the full-length gene. Hla 2017, 89, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Braud, V.; Jones, E.Y.; McMichael, A. The human major histocompatibility complex class Ib molecule HLA-E binds signal sequence-derived peptides with primary anchor residues at positions 2 and 9. Eur. J. Immunol. 1997, 27, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, M.P.; Adams, E.; Altman, J.D.; Birnbaum, M.E.; Boggiano, C.; Casorati, G.; Chien, Y.-H.; Conley, A.; Eckle, S.B.G.; Fruh, K.; et al. Casting a wider net: Immunosurveillance by nonclassical MHC molecules. PLoS Pathog. 2019, 15, e1007567. [Google Scholar] [CrossRef] [PubMed]
- Braud, V.M.; Allan, D.S.; Wilson, D.; McMichael, A.J. TAP- and tapasin-dependent HLA-E surface expression correlates with the binding of an MHC class I leader peptide. Curr. Biol. 1998, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.; Goodlett, D.R.; Ishitani, A.; Marquardt, H.; Geraghty, D.E. HLA-E surface expression depends on binding of TAP-dependent peptides derived from certain HLA class I signal sequences. J. Immunol. 1998, 160, 4951–4960. [Google Scholar] [PubMed]
- Bland, F.A.; Lemberg, M.K.; McMichael, A.J.; Martoglio, B.; Braud, V.M. Requirement of the proteasome for the trimming of signal peptide-derived epitopes presented by the nonclassical major histocompatibility complex class I molecule HLA-E. J. Biol. Chem. 2003, 278, 33747–33752. [Google Scholar] [CrossRef] [PubMed]
- Michaelsson, J.; Teixeira de Matos, C.; Achour, A.; Lanier, L.L.; Karre, K.; Soderstrom, K. A signal peptide derived from hsp60 binds HLA-E and interferes with CD94/NKG2A recognition. J. Exp. Med. 2002, 196, 1403–1414. [Google Scholar] [CrossRef]
- van Hall, T.; Oliveira, C.C.; Joosten, S.A.; Ottenhoff, T.H.M. The other Janus face of Qa-1 and HLA-E: Diverse peptide repertoires in times of stress. Microbes Infect. 2010, 12, 910–918. [Google Scholar] [CrossRef]
- Lorente, E.; Infantes, S.; Abia, D.; Barnea, E.; Beer, I.; Garcia, R.; Lasala, F.; Jimenez, M.; Mir, C.; Morreale, A.; et al. A viral, transporter associated with antigen processing (TAP)-independent, high affinity ligand with alternative interactions endogenously presented by the nonclassical human leukocyte antigen E class I molecule. J. Biol. Chem. 2012, 287, 34895–34903. [Google Scholar] [CrossRef]
- Wieten, L.; Mahaweni, N.M.; Voorter, C.E.M.; Bos, G.M.J.; Tilanus, M.G.J. Clinical and immunological significance of HLA-E in stem cell transplantation and cancer. Tissue Antigens 2014, 84, 523–535. [Google Scholar] [CrossRef]
- Lauterbach, N.; Wieten, L.; Popeijus, H.E.; Voorter, C.E.M.; Tilanus, M.G.J. HLA-E regulates NKG2C+ natural killer cell function through presentation of a restricted peptide repertoire. Hum. Immunol. 2015, 76, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, H.R.; Bowyer, G.; Brackenridge, S.; Lambe, T. HLA-E: Exploiting pathogen-host interactions for vaccine development. Clin. Exp. Immunol. 2019, 196, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Llano, M.; Carretero, M.; Ishitani, A.; Navarro, F.; López-Botet, M.; Geraghty, D.E. HLA-E is a major ligand for the natural killer inhibitory receptor CD94/NKG2A. Proc. Natl. Acad. Sci. 1998, 95, 5199–5204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.; Allan, D.S.; Bowen, M.; Powis, S.J.; Joseph, S.; Verma, S.; Hiby, S.E.; McMichael, A.J.; Loke, Y.W.; Braud, V.M. HLA-E is expressed on trophoblast and interacts with CD94/NKG2 receptors on decidual NK cells. Eur. J. Immunol. 2000, 30, 1623–1631. [Google Scholar] [CrossRef]
- Coupel, S.; Moreau, A.; Hamidou, M.; Horejsi, V.; Soulillou, J.-P.; Charreau, B. Expression and release of soluble HLA-E is an immunoregulatory feature of endothelial cell activation. Blood 2007, 109, 2806–2814. [Google Scholar] [CrossRef]
- Boegel, S.; Löwer, M.; Bukur, T.; Sorn, P.; Castle, J.C.; Sahin, U. HLA and proteasome expression body map. BMC Med. Genom. 2018, 11, 36. [Google Scholar] [CrossRef]
- Lo Monaco, E.; Tremante, E.; Cerboni, C.; Melucci, E.; Sibilio, L.; Zingoni, A.; Nicotra, M.R.; Natali, P.G.; Giacomini, P. Human Leukocyte Antigen E Contributes to Protect Tumor Cells from Lysis by Natural Killer Cells. Neoplasia 2011, 13, 822. [Google Scholar] [CrossRef]
- Pabón, M.A.; Navarro, C.E.; Osorio, J.C.; Gómez, N.; Moreno, J.P.; Donado, A.F.; Pérez, H.C.; Lozano, E. Impact of human leukocyte antigen molecules E, F, and G on the outcome of transplantation. Transpl. Proc. 2014, 46, 2957–2965. [Google Scholar] [CrossRef]
- Durrenberger, P.F.; Webb, L.V.; Sim, M.J.W.; Nicholas, R.S.; Altmann, D.M.; Boyton, R.J. Increased HLA-E expression in white matter lesions in multiple sclerosis. Immunology 2012, 137, 317–325. [Google Scholar] [CrossRef]
- Planas, R.; Carrillo, J.; Sanchez, A.; de Villa, M.C.R.; Nunez, F.; Verdaguer, J.; James, R.F.L.; Pujol-Borrell, R.; Vives-Pi, M. Gene expression profiles for the human pancreas and purified islets in type 1 diabetes: New findings at clinical onset and in long-standing diabetes. Clin. Exp. Immunol. 2010, 159, 23–44. [Google Scholar] [CrossRef]
- Joosten, S.A.; Sullivan, L.C.; Ottenhoff, T.H.M. Characteristics of HLA-E Restricted T-Cell Responses and Their Role in Infectious Diseases. J. Immunol. Res. 2016, 2016, 2695396. [Google Scholar] [CrossRef] [PubMed]
- Pietra, G.; Romagnani, C.; Moretta, L.; Mingari, M.C. HLA-E and HLA-E-bound peptides: Recognition by subsets of NK and T cells. Curr. Pharm. Des. 2009, 15, 3336–3344. [Google Scholar] [CrossRef] [PubMed]
- Plougastel, B.; Jones, T.; Trowsdale, J. Genomic structure, chromosome location, and alternative splicing of the human NKG2A gene. Immunogenetics 1996, 44, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Braud, V.M.; Allan, D.S.; O’Callaghan, C.A.; Soderstrom, K.; D’Andrea, A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; et al. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 1998, 391, 795–799. [Google Scholar] [CrossRef]
- Della Chiesa, M.; Sivori, S.; Carlomagno, S.; Moretta, L.; Moretta, A. Activating KIRs and NKG2C in Viral Infections: Toward NK Cell Memory? Front. Immunol. 2015, 6, 573. [Google Scholar] [CrossRef]
- Orbelyan, G.A.; Tang, F.; Sally, B.; Solus, J.; Meresse, B.; Ciszewski, C.; Grenier, J.-C.; Barreiro, L.B.; Lanier, L.L.; Jabri, B. Human NKG2E is expressed and forms an intracytoplasmic complex with CD94 and DAP12. J. Immunol. 2014, 193, 610–616. [Google Scholar] [CrossRef]
- Dukovska, D.; Fernandez-Soto, D.; Vales-Gomez, M.; Reyburn, H.T. NKG2H-Expressing T Cells Negatively Regulate Immune Responses. Front. Immunol. 2018, 9, 390. [Google Scholar] [CrossRef] [Green Version]
- Schmiedel, D.; Mandelboim, O. NKG2D Ligands-Critical Targets for Cancer Immune Escape and Therapy. Front. Immunol. 2018, 9, 2040. [Google Scholar] [CrossRef]
- Kim, D.-K.; Kabat, J.; Borrego, F.; Sanni, T.B.; You, C.-H.; Coligan, J.E. Human NKG2F is expressed and can associate with DAP12. Mol. Immunol. 2004, 41, 53–62. [Google Scholar] [CrossRef]
- Guma, M.; Angulo, A.; Vilches, C.; Gomez-Lozano, N.; Malats, N.; Lopez-Botet, M. Imprint of human cytomegalovirus infection on the NK cell receptor repertoire. Blood 2004, 104, 3664–3671. [Google Scholar] [CrossRef] [Green Version]
- López-Botet, M.; Muntasell, A.; Vilches, C. The CD94/NKG2C+ NK-cell subset on the edge of innate and adaptive immunity to human cytomegalovirus infection. Semin. Immunol. 2014, 26, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Q.; Ruckert, T.; Borst, E.M.; Dunst, J.; Haubner, A.; Durek, P.; Heinrich, F.; Gasparoni, G.; Babic, M.; Tomic, A.; et al. Peptide-specific recognition of human cytomegalovirus strains controls adaptive natural killer cells. Nat. Immunol. 2018, 19, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Muntasell, A.; Vilches, C.; Angulo, A.; López-Botet, M. Adaptive reconfiguration of the human NK-cell compartment in response to cytomegalovirus: A different perspective of the host-pathogen interaction. Eur. J. Immunol. 2013, 43, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- López-Botet, M.; Muntasell, A.; Martínez-Rodríguez, J.E.; López-Montañés, M.; Costa-García, M.; Pupuleku, A. Development of the adaptive NK cell response to human cytomegalovirus in the context of aging. Mech. Ageing Dev. 2016, 158, 23–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, Q.; Romagnani, C. About Training and Memory: NK-Cell Adaptation to Viral Infections. Adv. Immunol. 2017, 133, 171–207. [Google Scholar]
- Rolle, A.; Jager, D.; Momburg, F. HLA-E Peptide Repertoire and Dimorphism-Centerpieces in the Adaptive NK Cell Puzzle? Front. Immunol. 2018, 9, 2410. [Google Scholar] [CrossRef] [PubMed]
- Rolle, A.; Meyer, M.; Calderazzo, S.; Jager, D.; Momburg, F. Distinct HLA-E Peptide Complexes Modify Antibody-Driven Effector Functions of Adaptive NK Cells. Cell Rep. 2018, 24, 1967. [Google Scholar] [CrossRef]
- Lin, A.; Yan, W.-H. Heterogeneity of HLA-G Expression in Cancers: Facing the Challenges. Front. Immunol. 2018, 9, 2164. [Google Scholar] [CrossRef]
- Brostjan, C.; Belloón, T.; Sobanov, Y.; López-Botet, M.; Hofer, E. Differential expression of inhibitory and activating CD94/NKG2 receptors on NK cell clones. J. Immunol. Methods 2002, 264, 109–119. [Google Scholar] [CrossRef]
- Kaiser, B.K.; Barahmand-pour, F.; Paulsene, W.; Medley, S.; Geraghty, D.E.; Strong, R.K. Interactions between NKG2x Immunoreceptors and HLA-E Ligands Display Overlapping Affinities and Thermodynamics. J. Immunol. 2014, 174, 2878–2884. [Google Scholar] [CrossRef]
- Pump, W.C.; Kraemer, T.; Huyton, T.; Hò, G.G.T.; Blasczyk, R.; Bade-Doeding, C. Between innate and adaptive immune responses: NKG2A, NKG2C, and CD8+ T cell recognition of HLA-E restricted self-peptides acquired in the absence of HLA-Ia. Int. J. Mol. Sci. 2019, 20, 1454. [Google Scholar] [CrossRef] [PubMed]
- Jouand, N.; Bressollette-Bodin, C.; Gerard, N.; Giral, M.; Guerif, P.; Rodallec, A.; Oger, R.; Parrot, T.; Allard, M.; Cesbron-Gautier, A.; et al. HCMV triggers frequent and persistent UL40-specific unconventional HLA-E-restricted CD8 T-cell responses with potential autologous and allogeneic peptide recognition. PLoS Pathog. 2018, 14, e1007041. [Google Scholar] [CrossRef] [PubMed]
- McMurtrey, C.; Harriff, M.J.; Swarbrick, G.M.; Duncan, A.; Cansler, M.; Null, M.; Bardet, W.; Jackson, K.W.; Lewinsohn, D.A.; Hildebrand, W.; et al. T cell recognition of Mycobacterium tuberculosis peptides presented by HLA-E derived from infected human cells. PLoS ONE 2017, 12, e0188288. [Google Scholar] [CrossRef] [PubMed]
- Bian, Y.; Shang, S.; Siddiqui, S.; Zhao, J.; Joosten, S.A.; Ottenhoff, T.H.M.; Cantor, H.; Wang, C.-R. MHC Ib molecule Qa-1 presents Mycobacterium tuberculosis peptide antigens to CD8+ T cells and contributes to protection against infection. PLoS Pathog. 2017, 13, e1006384. [Google Scholar] [CrossRef] [PubMed]
- Prezzemolo, T.; van Meijgaarden, K.E.; Franken, K.L.M.C.; Caccamo, N.; Dieli, F.; Ottenhoff, T.H.M.; Joosten, S.A. Detailed characterization of human Mycobacterium tuberculosis specific HLA-E restricted CD8(+) T cells. Eur. J. Immunol. 2018, 48, 293–305. [Google Scholar] [CrossRef]
- Salerno-Goncalves, R.; Fernandez-Vina, M.; Lewinsohn, D.M.; Sztein, M.B. Identification of a human HLA-E-restricted CD8+ T cell subset in volunteers immunized with Salmonella enterica serovar Typhi strain Ty21a typhoid vaccine. J. Immunol. 2004, 173, 5852–5862. [Google Scholar] [CrossRef] [PubMed]
- Parham, P.; Adams, E.J. Species-specific evolution of MHC class I genes in the higher primates. Immunol. Rev. 2001, 183, 41–64. [Google Scholar]
- Strong, R.K.; Holmes, M.A.; Li, P.; Braun, L.; Lee, N.; Geraghty, D.E. HLA-E allelic variants. Correlating differential expression, peptide affinities, crystal structures, and thermal stabilities. J. Biol. Chem. 2003, 278, 5082–5090. [Google Scholar] [CrossRef]
- Maier, S.; Grzeschik, M.; Weiss, E.H.; Ulbrecht, M. Implications of HLA-E allele expression and different HLA-E ligand diversity for the regulation of NK cells. Hum. Immunol. 2000, 61, 1059–1065. [Google Scholar] [CrossRef]
- Grimsley, C.; Ober, C. Genetic Studies Evidence for Selection of HLA-E. Significance 1997, 8859. [Google Scholar]
- Nattermann, J.; Nischalke, H.D.; Hofmeister, V.; Kupfer, B.; Ahlenstiel, G.; Feldmann, G.; Rockstroh, J.; Weiss, E.H.; Sauerbruch, T.; Spengler, U. HIV-1 infection leads to increased HLA-E expression resulting in impaired function of natural killer cells. Antivir. Ther. 2005, 10, 95–107. [Google Scholar] [PubMed]
- Nattermann, J.; Nischalke, H.D.; Hofmeister, V.; Ahlenstiel, G.; Zimmermann, H.; Leifeld, L.; Weiss, E.H.; Sauerbruch, T.; Spengler, U. The HLA-A2 restricted T cell epitope HCV core 35–44 stabilizes HLA-E expression and inhibits cytolysis mediated by natural killer cells. Am. J. Pathol. 2005, 166, 443–453. [Google Scholar] [CrossRef]
- Lampen, M.H.; Hassan, C.; Sluijter, M.; Geluk, A.; Dijkman, K.; Tjon, J.M.; de Ru, A.H.; van der Burg, S.H.; van Veelen, P.A.; van Hall, T. Alternative peptide repertoire of HLA-E reveals a binding motif that is strikingly similar to HLA-A2. Mol. Immunol. 2013, 53, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, T.; Celik, A.A.; Huyton, T.; Kunze-Schumacher, H.; Blasczyk, R.; Bade-Doding, C. HLA-E: Presentation of a Broader Peptide Repertoire Impacts the Cellular Immune Response-Implications on HSCT Outcome. Stem Cells Int. 2015, 2015, 346714. [Google Scholar] [CrossRef]
- Celik, A.A.; Kraemer, T.; Huyton, T.; Blasczyk, R.; Bade-Döding, C. The diversity of the HLA-E-restricted peptide repertoire explains the immunological impact of the Arg107Gly mismatch. Immunogenetics 2016, 68, 29–41. [Google Scholar] [CrossRef]
- Dong, Y.; Lieskovska, J.; Kedrin, D.; Porcelli, S.; Mandelboim, O.; Bushkin, Y. Soluble nonclassical HLA generated by the metalloproteinase pathway. Hum. Immunol. 2003, 64, 802–810. [Google Scholar] [CrossRef]
- Allard, M.; Oger, R.; Vignard, V.; Percier, J.-M.; Fregni, G.; Perier, A.; Caignard, A.; Charreau, B.; Bernardeau, K.; Khammari, A.; et al. Serum soluble HLA-E in melanoma: A new potential immune-related marker in cancer. PLoS ONE 2011, 6, e21118. [Google Scholar] [CrossRef]
- Morandi, F.; Cangemi, G.; Barco, S.; Amoroso, L.; Giuliano, M.; Gigliotti, A.R.; Pistoia, V.; Corrias, M.V. Plasma levels of soluble HLA-E and HLA-F at diagnosis may predict overall survival of neuroblastoma patients. Biomed. Res. Int. 2013, 2013, 956878. [Google Scholar] [CrossRef]
- Wagner, B.; da Silva Nardi, F.; Schramm, S.; Kraemer, T.; Celik, A.A.; Durig, J.; Horn, P.A.; Duhrsen, U.; Nuckel, H.; Rebmann, V. HLA-E allelic genotype correlates with HLA-E plasma levels and predicts early progression in chronic lymphocytic leukemia. Cancer 2017, 123, 814–823. [Google Scholar] [CrossRef]
- Goel, R.; Kabeerdoss, J.; Mohan, H.; Danda, S.; Jayaseelan, V.; Kumar, T.S.; Jude, J.; Bacon, P.; Joseph, G.; Danda, D. Soluble-HLA-E: A follow up biomarker in Takayasu arteritis, independent of HLA-E genotype. Int. J. Rheum. Dis. 2018, 21, 532–540. [Google Scholar] [CrossRef]
- Schulte, D.; Vogel, M.; Langhans, B.; Kramer, B.; Korner, C.; Nischalke, H.D.; Steinberg, V.; Michalk, M.; Berg, T.; Rockstroh, J.K.; et al. The HLA-E(R)/HLA-E(R) genotype affects the natural course of hepatitis C virus (HCV) infection and is associated with HLA-E-restricted recognition of an HCV-derived peptide by interferon-gamma-secreting human CD8(+) T cells. J. Infect. Dis. 2009, 200, 1397–1401. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Fulgencio, M.; Berenguer, J.; Rallon, N.; Fernandez-Rodriguez, A.; Miralles, P.; Soriano, V.; Jimenez-Sousa, M.A.; Cosin, J.; Medrano, J.; Garcia-Alvarez, M.; et al. HLA-E variants are associated with sustained virological response in HIV/hepatitis C virus-coinfected patients on hepatitis C virus therapy. Aids 2013, 27, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Crux, N.B.; Elahi, S. Human Leukocyte Antigen (HLA) and Immune Regulation: How Do Classical and Non-Classical HLA Alleles Modulate Immune Response to Human Immunodeficiency Virus and Hepatitis C Virus Infections? Front. Immunol. 2017, 8, 832. [Google Scholar] [CrossRef] [PubMed]
- Rohn, H.; Michita, R.T.; Schramm, S.; Dolff, S.; Gackler, A.; Korth, J.; Heinemann, F.M.; Wilde, B.; Trilling, M.; Horn, P.A.; et al. HLA-E Polymorphism Determines Susceptibility to BK Virus Nephropathy after Living-Donor Kidney Transplant. Cells 2019, 8, 847. [Google Scholar] [CrossRef]
- Prasetyo, A.A.; Dharmawan, R.; Raharjo, I. Hudiyono Human Leukocyte Antigen-E Alleles are Associated with Hepatitis C Virus, Torque Teno Virus, and Toxoplasma Co-infections but are not Associated with Hepatitis B Virus, Hepatitis D Virus, and GB Virus C Co-infections in Human Immunodeficiency Virus Patient. J. Glob. Infect. Dis. 2016, 8, 75–81. [Google Scholar]
- Zhang, J.; Pan, L.; Chen, L.; Feng, X.; Zhou, L.; Zheng, S. Non-classical MHC-Ι genes in chronic hepatitis B and hepatocellular carcinoma. Immunogenetics 2012, 64, 251–258. [Google Scholar] [CrossRef]
- Lajoie, J.; Hargrove, J.; Zijenah, L.S.; Humphrey, J.H.; Ward, B.J.; Roger, M. Genetic Variants in Nonclassical Major Histocompatibility Complex Class I Human Leukocyte Antigen (HLA)–E and HLA-G Molecules Are Associated with Susceptibility to Heterosexual Acquisition of HIV-1. J. Infect. Dis. 2005, 193, 298–301. [Google Scholar] [CrossRef]
- Guberina, H.; da Silva Nardi, F.; Michita, R.T.; Dolff, S.; Bienholz, A.; Heinemann, F.M.; Wilde, B.; Trilling, M.; Horn, P.A.; Kribben, A.; et al. Susceptibility of HLA-E*01:03 Allele Carriers to Develop Cytomegalovirus Replication After Living-Donor Kidney Transplantation. J. Infect. Dis. 2018, 217, 1918–1922. [Google Scholar] [CrossRef]
- Ferguson, R.; Ramanakumar, A.V.; Richardson, H.; Tellier, P.-P.; Coutlee, F.; Franco, E.L.; Roger, M. Human leukocyte antigen (HLA)-E and HLA-G polymorphisms in human papillomavirus infection susceptibility and persistence. Hum. Immunol. 2011, 72, 337–341. [Google Scholar] [CrossRef]
- Tamouza, R.; Busson, M.; Fortier, C.; Diagne, I.; Diallo, D.; Sloma, I.; Contouris, H.; Krishnamoorthy, R.; Labie, D.; Girot, R.; et al. HLA-E*0101 allele in homozygous state favors severe bacterial infections in sickle cell anemia. Hum. Immunol. 2007, 68, 849–853. [Google Scholar] [CrossRef]
- Tamouza, R.; Rocha, V.; Busson, M.; Fortier, C.; El Sherbini, S.M.; Esperou, H.; Filion, A.; Socie, G.; Dulphy, N.; Krishnamoorthy, R.; et al. Association of HLA-E polymorphism with severe bacterial infection and early transplant-related mortality in matched unrelated bone marrow transplantation. Transplantation 2005, 80, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Lu, R.; Xie, S.; Wen, X.; Wang, H.; Gao, X.; Guo, L. Human leukocyte antigen-E alleles and expression in patients with serous ovarian cancer. Cancer Sci. 2015, 106, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wieten, L.; Wang, S.; Cai, Y.; Olieslagers, T.; He, L.; Tilanus, M.G.J.; Hong, W. Clinical significance of HLA-E genotype and surface/soluble expression levels between healthy individuals and patients with acute leukemia. Leuk. Lymphoma 2018, 60, 208–251. [Google Scholar] [CrossRef] [PubMed]
- Hirankarn, N.; Kimkong, I.; Mutirangura, A. HLA-E polymorphism in patients with nasopharyngeal carcinoma. Tissue Antigens 2004, 64, 588–592. [Google Scholar] [CrossRef]
- Douik, H.; Romdhane, N.A.; Guemira, F. Are HLA-E*0103 alleles predictive markers for nasopharyngeal cancer risk? Pathol. Res. Pract. 2016, 212, 345–349. [Google Scholar] [CrossRef]
- Martin, P.; Krsnik, I.; Navarro, B.; Provencio, M.; Garcia, J.F.; Bellas, C.; Vilches, C.; Gomez-Lozano, N. HLA Allele E*01:01 Is Associated with a Reduced Risk of EBV-Related Classical Hodgkin Lymphoma Independently of HLA-A*01/*02. PLoS ONE 2015, 10, e0135512. [Google Scholar] [CrossRef]
- Hassen, E.; Ghedira, R.; Ghandri, N.; Farhat, K.; Gabbouj, S.; Bouaouina, N.; Abdelaziz, H.; Nouri, A.; Chouchane, L. Lack of association between human leukocyte antigen-E alleles and nasopharyngeal carcinoma in Tunisians. DNA Cell Biol. 2011, 30, 603–609. [Google Scholar] [CrossRef]
- Veiga-Castelli, L.C.; de Paula Cruz, A.S.; Inacio, M.M.; Mendes-Junior, C.T.; Vianello-Brondani, R.; Moreau, P.; Castelli, E.C.; Donadi, E.A. Lack of association between HLA-E polymorphisms and transitional cell carcinoma of the bladder. Tissue Antigens 2013, 82, 197–200. [Google Scholar] [CrossRef]
- Moya-Quiles, M.R.; Martinez-Escribano, J.; Guerra-Perez, N.; Muro, M.; Marin, L.; Campillo, J.A.; Montes-Ares, O.; Frias, J.; Minguela, A.; Garcia-Alonso, A.M.; et al. Lack of association between HLA-E polymorphism and primary cutaneous melanoma in Spanish patients. J. Derm. Sci. 2005, 40, 62–64. [Google Scholar] [CrossRef]
- Zhen, Z.-J.; Ling, J.-Y.; Cai, Y.; Luo, W.-B.; He, Y.-J. Impact of HLA-E gene polymorphism on HLA-E expression in tumor cells and prognosis in patients with stage III colorectal cancer. Med. Oncol. 2013, 30, 482. [Google Scholar] [CrossRef]
- Tremante, E.; Ginebri, A.; Lo Monaco, E.; Benassi, B.; Frascione, P.; Grammatico, P.; Cappellacci, S.; Catricala, C.; Arcelli, D.; Natali, P.G.; et al. A melanoma immune response signature including Human Leukocyte Antigen-E. Pigment. Cell Melanoma Res. 2014, 27, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhang, D.; Li, F.; Xiao, Z.; Wu, M.; Shi, D.; Xiang, P.; Bao, Z. Loss of Fas expression and high expression of HLA-E promoting the immune escape of early colorectal cancer cells. Oncol. Lett. 2017, 13, 3379–3386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozgul Ozdemir, R.B.; Ozdemir, A.T.; Oltulu, F.; Kurt, K.; Yigitturk, G.; Kirmaz, C. A comparison of cancer stem cell markers and nonclassical major histocompatibility complex antigens in colorectal tumor and noncancerous tissues. Ann. Diagn. Pathol. 2016, 25, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.-Y.; Lv, Y.-G.; Wang, L.; Shi, S.-J.; Yang, F.; Zheng, G.-X.; Wen, W.-H.; Yang, A.-G. Predictive value of HLA-G and HLA-E in the prognosis of colorectal cancer patients. Cell. Immunol. 2015, 293, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Bossard, C.; Bezieau, S.; Matysiak-Budnik, T.; Volteau, C.; Laboisse, C.L.; Jotereau, F.; Mosnier, J.-F. HLA-E/beta2 microglobulin overexpression in colorectal cancer is associated with recruitment of inhibitory immune cells and tumor progression. Int. J. Cancer 2012, 131, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Talebian Yazdi, M.; van Riet, S.; van Schadewijk, A.; Fiocco, M.; van Hall, T.; Taube, C.; Hiemstra, P.S.; van der Burg, S.H. The positive prognostic effect of stromal CD8+ tumor-infiltrating T cells is restrained by the expression of HLA-E in non-small cell lung carcinoma. Oncotarget 2016, 7, 3477–3488. [Google Scholar] [PubMed]
- Andersson, E.; Poschke, I.; Villabona, L.; Carlson, J.W.; Lundqvist, A.; Kiessling, R.; Seliger, B.; Masucci, G. V Non-classical HLA-class I expression in serous ovarian carcinoma: Correlation with the HLA-genotype, tumor infiltrating immune cells and prognosis. Oncoimmunology 2016, 5, e1052213. [Google Scholar] [CrossRef]
- Gooden, M.; Lampen, M.; Jordanova, E.S.; Leffers, N.; Trimbos, J.B.; van der Burg, S.H.; Nijman, H.; van Hall, T. HLA-E expression by gynecological cancers restrains tumor-infiltrating CD8(+) T lymphocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 10656–10661. [Google Scholar] [CrossRef]
- Hodgkinson, A.D.; Millward, B.A.; Demaine, A.G. The HLA-E locus is associated with age at onset and susceptibility to type 1 diabetes mellitus. Hum. Immunol. 2000, 61, 290–295. [Google Scholar] [CrossRef]
- Paladini, F.; Belfiore, F.; Cocco, E.; Carcassi, C.; Cauli, A.; Vacca, A.; Fiorillo, M.T.; Mathieu, A.; Cascino, I.; Sorrentino, R. HLA-E gene polymorphism associates with ankylosing spondylitis in Sardinia. Arthritis Res. Ther. 2009, 11, R171. [Google Scholar] [CrossRef]
- Santos, M.R.; Couto, A.R.; Foroni, I.; Bettencourt, B.F.; Li, Z.; Meneses, R.; Wheeler, L.; Pereira, J.; Pimentel-Santos, F.; Fonseca, J.E.; et al. Non-classical human leucocyte antigens in ankylosing spondylitis: Possible association with HLA-E and HLA-F. RMD Open 2018, 4, e000677. [Google Scholar] [CrossRef] [PubMed]
- Bhanusali, D.G.; Sachdev, A.; Rahmanian, A.; Gerlach, J.A.; Tong, J.C.; Seiffert-Sinha, K.; Sinha, A.A. HLA-E*0103X is associated with susceptibility to Pemphigus vulgaris. Exp. Dermatol. 2013, 22, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Altun, E.; Yayli, S.; Toraman, B.; Arica, D.A.; Kalay, E.; Selcuk, L.B.; Bahadir, S. HLA-E * 0101 / 0103X is Associated with Susceptibility to Pemphigus Vulgaris: A Case-control Study. Acta Dermatovenerol. Croat. 2017, 25, 189–194. [Google Scholar] [PubMed]
- Iwaszko, M.; Swierkot, J.; Kolossa, K.; Jeka, S.; Wiland, P.; Bogunia-Kubik, K. Polymorphisms within the human leucocyte antigen-E gene and their associations with susceptibility to rheumatoid arthritis as well as clinical outcome of anti-tumour necrosis factor therapy. Clin. Exp. Immunol. 2015, 182, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Sokolik, R.; Gebura, K.; Iwaszko, M.; Swierkot, J.; Korman, L.; Wiland, P.; Bogunia-Kubik, K. Significance of association of HLA-C and HLA-E with psoriatic arthritis. Hum. Immunol. 2014, 75, 1188–1191. [Google Scholar] [CrossRef]
- Zeng, X.; Chen, H.; Gupta, R.; Paz-Altschul, O.; Bowcock, A.M.; Liao, W. Deletion of the activating NKG2C receptor and a functional polymorphism in its ligand HLA-E in psoriasis susceptibility. Exp. Dermatol. 2013, 22, 679–681. [Google Scholar] [CrossRef] [Green Version]
- van Bijnen, S.T.A.; Cossu, M.; Roeven, M.W.H.; Jansen, T.L.; Preijers, F.; Spanholtz, J.; Dolstra, H.; Radstake, T.R.D.J. Functionally active NKG2A-expressing natural killer cells are elevated in rheumatoid arthritis patients compared to psoriatic arthritis patients and healthy donors. Clin. Exp. Rheumatol. 2015, 33, 795–804. [Google Scholar]
- Walsh, C.E.; Ryan, E.J.; O’Farrelly, C.; Golden-Mason, L.; FitzGerald, O.; Veale, D.J.; Bresnihan, B.; Fearon, U. Differential expression of NK receptors CD94 and NKG2A by T cells in rheumatoid arthritis patients in remission compared to active disease. PLoS ONE 2011, 6, e27182. [Google Scholar] [CrossRef]
- Park, K.S.; Park, J.H.; Song, Y.W. Inhibitory NKG2A and activating NKG2D and NKG2C natural killer cell receptor genes: Susceptibility for rheumatoid arthritis. Tissue Antigens 2008, 72, 342–346. [Google Scholar] [CrossRef]
- Cauli, A.; Dessole, G.; Piga, M.; Angioni, M.M.; Pinna, S.; Floris, A.; Congia, M.; Mascia, E.; Paladini, F.; Tedeschi, V.; et al. Expression analysis of HLA-E and NKG2A and NKG2C receptors points at a role for natural killer function in ankylosing spondylitis. RMD Open 2018, 4, e000597. [Google Scholar] [CrossRef] [Green Version]
- Patel, F.; Marusina, A.I.; Duong, C.; Adamopoulos, I.E.; Maverakis, E. NKG2C, HLA-E and their association with psoriasis. Exp. Dermatol. 2013, 22, 797–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, S.W.; Kim, E.-O.; Ryu, E.S.; Kim, T.-J.; Kim, J.N.; Choi, J.E.; Kye, Y.C.; Lee, K.-M. Upregulation of Fas and downregulation of CD94/NKG2A inhibitory receptors on circulating natural killer cells in patients with new-onset psoriasis. Br. J. Dermatol. 2009, 161, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.H.; Jee, S.H.; Sheu, B.C.; Huang, Y.L.; Tseng, M.P.; Hsu, S.M.; Tsai, T.-F. Increased expression of the natural killer cell inhibitory receptor CD94/NKG2A and CD158b on circulating and lesional T cells in patients with chronic plaque psoriasis. Br. J. Dermatol. 2006, 155, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Di Cristofaro, J.; Pelardy, M.; Loundou, A.; Basire, A.; Gomez, C.; Chiaroni, J.; Thomas, P.; Reynaud-Gaubert, M.; Picard, C. HLA-01:03 Allele in Lung Transplant Recipients Correlates with Higher Chronic Lung Allograft Dysfunction Occurrence. J. Immunol. Res. 2016, 2016, 1910852. [Google Scholar] [CrossRef] [PubMed]
- Tamouza, R.; Busson, M.; Rocha, V.; Fortier, C.; Haddad, Y.; Brun, M.; Boukouaci, W.; Bleux, H.; Socie, G.; Krishnamoorthy, R.; et al. Homozygous status for HLA-E*0103 confers protection from acute graft-versus-host disease and transplant-related mortality in HLA-matched sibling hematopoietic stem cell transplantation. Transplantation 2006, 82, 1436–1440. [Google Scholar] [CrossRef]
- Danzer, M.; Polin, H.; Proll, J.; Haunschmid, R.; Hofer, K.; Stabentheiner, S.; Hackl, C.; Kasparu, H.; Konig, J.; Hauser, H.; et al. Clinical significance of HLA-E*0103 homozygosity on survival after allogeneic hematopoietic stem-cell transplantation. Transplantation 2009, 88, 528–532. [Google Scholar] [CrossRef]
- Mossallam, G.I.; Fattah, R.A.; El-Haddad, A.; Mahmoud, H.K. HLA-E polymorphism and clinical outcome after allogeneic hematopoietic stem cell transplantation in Egyptian patients. Hum. Immunol. 2015, 76, 161–165. [Google Scholar] [CrossRef]
- Furst, D.; Bindja, J.; Arnold, R.; Herr, W.; Schwerdtfeger, R.; Muller, C.H.; Recker, K.; Schrezenmeier, H.; Mytilineos, J. HLA-E polymorphisms in hematopoietic stem cell transplantation. Tissue Antigens 2012, 79, 287–290. [Google Scholar] [CrossRef]
- Mosaad, Y.M.; Abdel-Dayem, Y.; El-Deek, B.S.; El-Sherbini, S.M. Association between HLA-E *0101 homozygosity and recurrent miscarriage in Egyptian women. Scand. J. Immunol. 2011, 74, 205–209. [Google Scholar] [CrossRef]
- Tripathi, P.; Naik, S.; Agrawal, S. HLA-E and immunobiology of pregnancy. Tissue Antigens 2006, 67, 207–213. [Google Scholar] [CrossRef]
- Kanai, T.; Fujii, T.; Keicho, N.; Tokunaga, K.; Yamashita, T.; Hyodo, H.; Miki, A.; Unno, N.; Kozuma, S.; Taketani, Y. Polymorphism of human leukocyte antigen-E gene in the Japanese population with or without recurrent abortion. Am. J. Reprod. Immunol. 2001, 45, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, R.; Christiansen, O.B.; Bennett, E.P.; Jersild, C. HLA-E polymorphism in patients with recurrent spontaneous abortion. Tissue Antigens 1998, 52, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Meuleman, T.; Lashley, L.E.L.O.; Dekkers, O.M.; van Lith, J.M.M.; Claas, F.H.J.; Bloemenkamp, K.W.M. HLA associations and HLA sharing in recurrent miscarriage: A systematic review and meta-analysis. Hum. Immunol. 2015, 76, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Gelmini, G.F.; Costa, C.H.; da Silva Nardi, F.; Wowk, P.F.; Mattar, S.B.; Schuffner, A.; da Graça Bicalho, M.; Roxo, V.M.M.S. Is HLA-E a possible genetic marker relevant for natural conception? Am. J. Reprod. Immunol. 2016, 76, 439–442. [Google Scholar] [CrossRef]


{kind=link}
| VMAPRTLLL | HLA-A*01, -A*03, -A*11, -A*29, -A*30, -A*31, -A*32, -A*33, -A*36, and -A*74, HLA-C*02 and C*15 |
| VMAPRTLVL | HLA-A*02, -A*23, -A*24, -A*25, -A*26, -A*34:02, -A*34:06, -A*43, -A*66, and -A*69 |
| IMAPRTLVL | HLA-A*34:01 |
| VMPPRTLLL | HLA-A*80 |
| VMAPRTVLL | HLA-B*07, -B*08, -B*14, -B*38, -B*39, -B*42, -B48, -B*67, -B*73 and -B*81 |
| VTAPRTLLL | HLA-B*13, -B*18, -B*27, -B*37, -B*40, -B*44, -B*47, -B*54, -B*55 -B*56, -B*59, -B*82 and -B*83 |
| VTAPRTVLL | HLA-B*15, -B*35, -B*40, -B*41, -B*44:18, -B*45, -B*46, -B*49, -B*50, -B*51, -B*52, -B*53, -B*57, -B*58 and -B*78 |
| VMAPRTLIL | HLA-C*01, -C*03, -C*04, -C*05, -C*06, -C*08, -C*12, -C*14, -C*16 and -C*17:02 |
| VMAPRALLL | HLA-C*06:17, -C*07 and -C*18 |
| VMAPRTLTL | HLA-C*08:09 |
| VMAPQALLL | HLA-C*17:01, C*17:03 and -C*17:05 |
| VMAPRTLFL | HLA-G*01 |
| QMRPVSRVL | Hsp60 |
| ALALVRMLI | ATP-binding cassette protein MRP7 |
| VMAPRTL(I/V/L)L | HCMV UL40 protein |
| YLLPRRGPRL | Hepatitis C virus core protein |
| SQAPLPCVL | Epstein–Barr virus BZLF1 protein |
| AISPRTLNA | HIV, P24 |
| RMAATAQVL | Mycobacterium tuberculosis, Mtb14 |
| RMPPLGHEL | Mycobacterium tuberculosis, P49 |
| RLPAKAPLL | Mycobacterium tuberculosis, Mtb44 |
| GMQFDRGYL | Salmonella typhimurium serovar Typhi, GroEL |
| Pathologies | HLA-ER (*01:01) | HLA-EG (*01:03) |
|---|---|---|
| Viral infections | protection/risk | protection/risk |
| Cancer 1 | protection | risk |
| Bacterial infections | risk | protection |
| HSC and bone marrow transplantation 2 | risk | protection |
| Recurrent abortions | risk | protection |
| Autoimmune disorders | protection/risk | protection/risk |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanevskiy, L.; Erokhina, S.; Kobyzeva, P.; Streltsova, M.; Sapozhnikov, A.; Kovalenko, E. Dimorphism of HLA-E and Its Disease Association. Int. J. Mol. Sci. 2019, 20, 5496. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215496
Kanevskiy L, Erokhina S, Kobyzeva P, Streltsova M, Sapozhnikov A, Kovalenko E. Dimorphism of HLA-E and Its Disease Association. International Journal of Molecular Sciences. 2019; 20(21):5496. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215496
Chicago/Turabian StyleKanevskiy, Leonid, Sofya Erokhina, Polina Kobyzeva, Maria Streltsova, Alexander Sapozhnikov, and Elena Kovalenko. 2019. "Dimorphism of HLA-E and Its Disease Association" International Journal of Molecular Sciences 20, no. 21: 5496. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215496