Exploring the Molecular Mechanism underlying the Stable Purple-Red Leaf Phenotype in Lagerstroemia indica cv. Ebony Embers


Abstract
:1. Introduction
2. Results
2.1. Anthocyanin Analysis in the Leaves of the Two Lagerstroemia indica Cutlivars
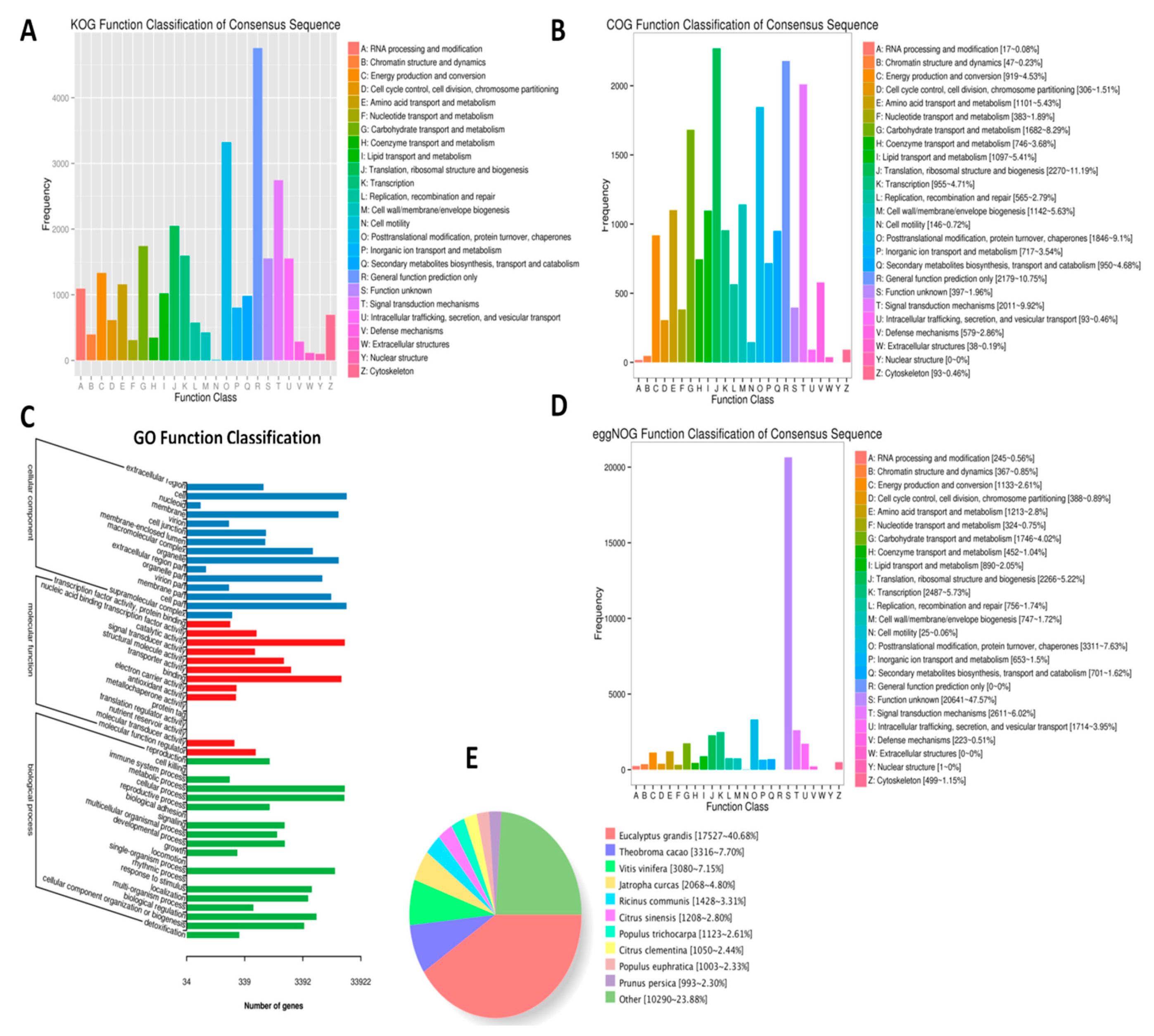
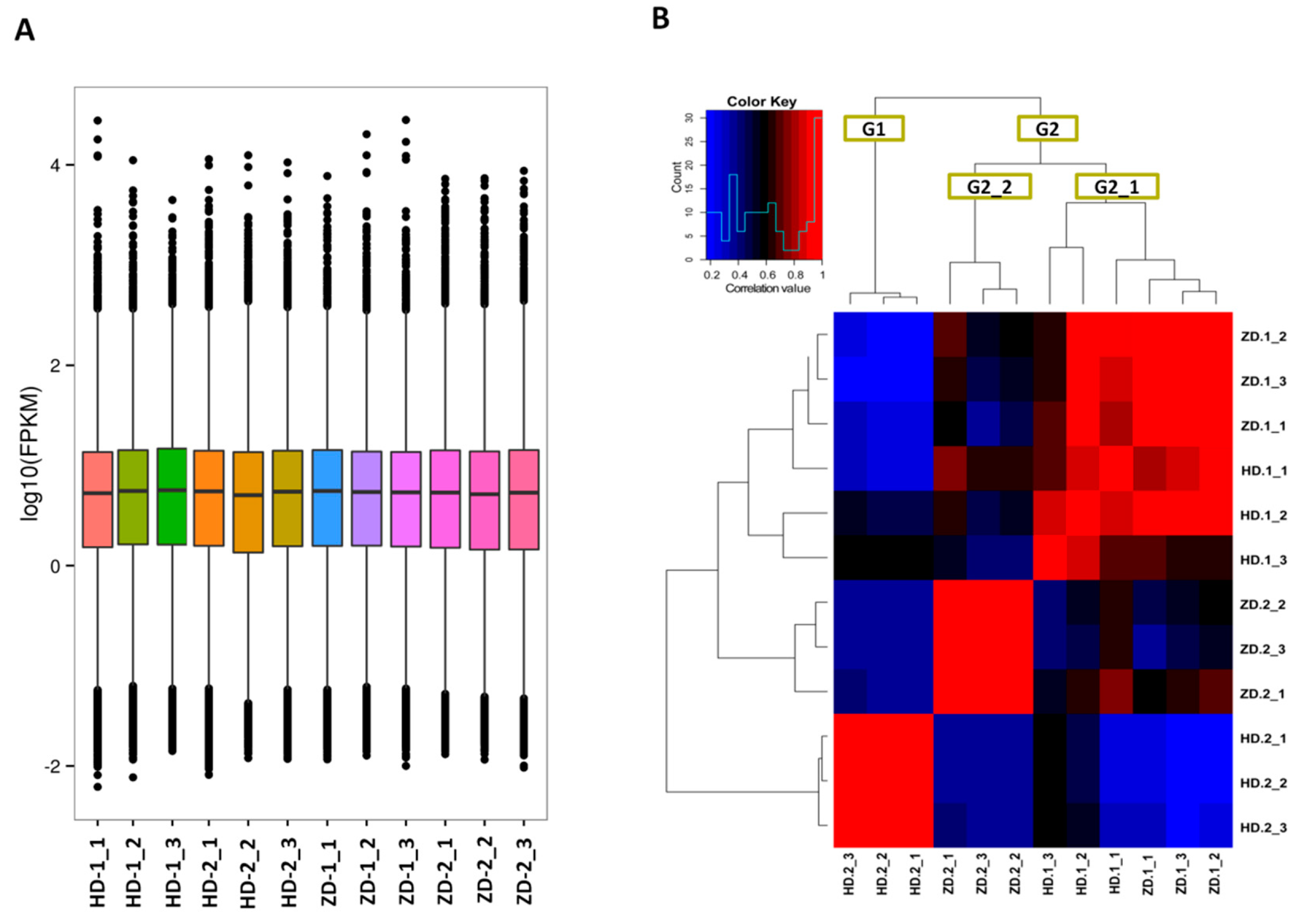
2.2. De Novo Transcriptome Assembly and Gene Expression Profiles in the Two L. indica Cutlivars at Different Leaf Developmental Stages
2.3. Differentially Expressed Genes between the Two L. indica Cutlivars
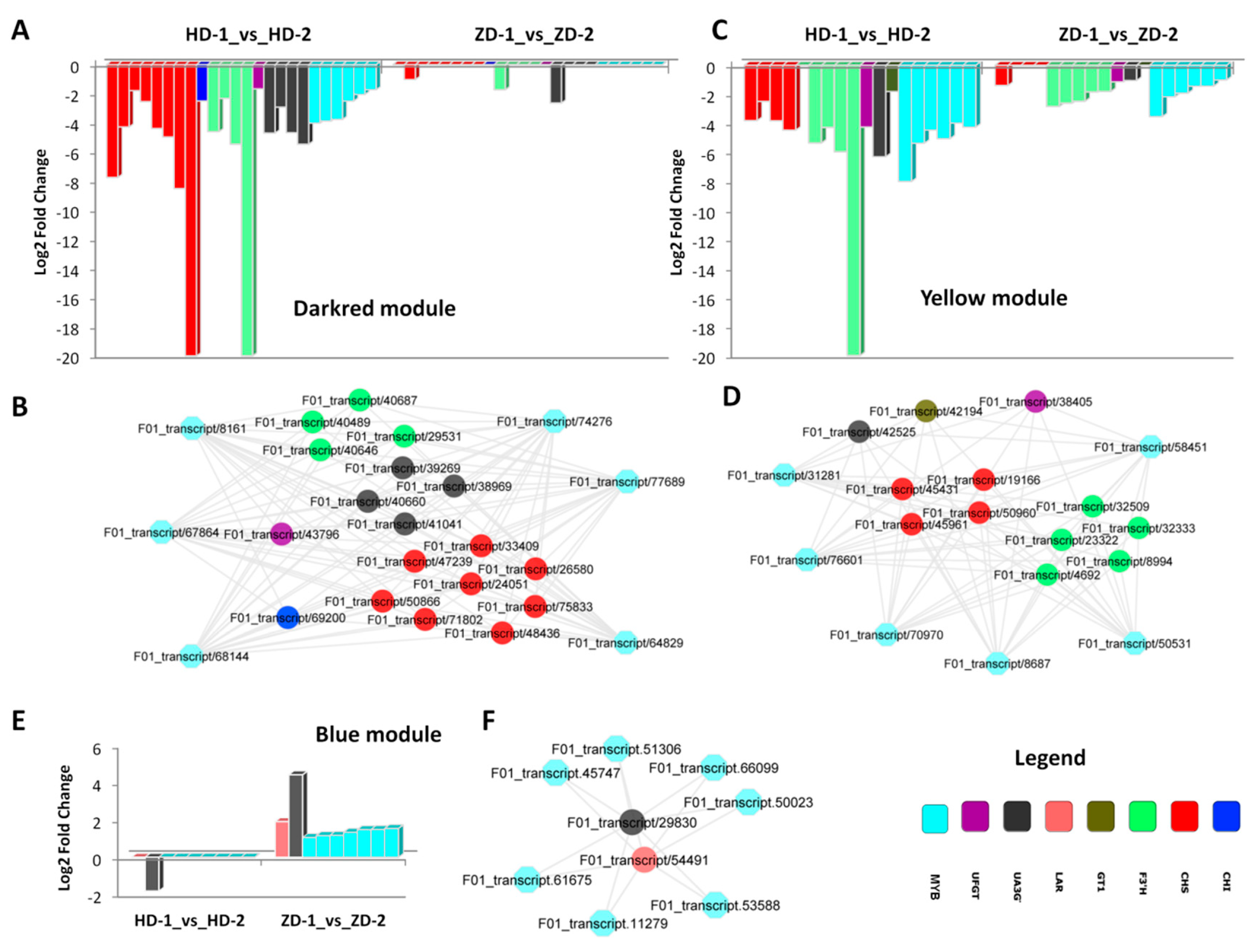
2.4. DEGs Related to the Flavonoid–Anthocyanin Biosynthesis and Mechanisms Underlying the Differential Leaf Color Phenotypes
2.5. Active MYB Transcripion Factors Regulating Gene Expression for the Differential Leaf Color Phenotypes
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Anthocyanin Analysis
4.3. Transcriptome Sequencing and Data Analysis
4.4. Gene Co-Expression Analysis
4.5. Quantitative RT-PCR Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pounders, C.; Rinehart, T.; Edwards, N.; Knight, P. An analysis of combining ability for height, leaf out, bloom date, and flower color for crapemyrtle. HortScience 2007, 42, 1496–1499. [Google Scholar] [CrossRef]
- Liu, Y.S.; Zetter, R.; Ferguson, D.K.; Zou, C. Lagerstroemia (Lythraceae) pollen from the Miocene of eastern China. Grana 2008, 47, 262–271. [Google Scholar] [CrossRef]
- Cabrera, R.I. Evaluating and promoting the cosmopolitan and multipurpose Lagerstroemia. In Proceedings of the XXVI International Horticultural Congress, Toronto, ON, Canada, 11−17 August 2002; pp. 177–184. [Google Scholar]
- Pounders, C.; Scheffler, B.E.; Rinehart, T.A. ‘Ebony Embers’, ‘Ebony Fire’, ‘Ebony Flame’, ‘EbonyGlow’, and ‘Ebony and Ivory’ Dark-leaf Crapemyrtles. HortScience 2013, 48, 1568–1570. [Google Scholar] [CrossRef]
- Zhengkang, P. Cultivation Managements of Lagerstroemia indica and its Application in the Landscape. J. Anhui Agric. Sci. 2006, 34, 5832–5833. [Google Scholar]
- Knox, G. New Crapemyrtles with Burgundy Leaves from Spring through Fall. UF/IFAS Extension. 21 July 2014. Available online: http://nwdistrict.ifas.ufl.edu/hort/2014/07/21/new-crapemyrtles-with-burgundy-leaves-from-spring-through-fall/ (accessed on 15 February 2019).
- Pounders, C.T.; Blythe, E.K.; Fare, D.C.; Knox, G.W.; Sibley, J.L. Crapemyrtle genotype × environment interactions, and trait stability for plant height, leaf-out, and flowering. HortScience 2010, 45, 198–207. [Google Scholar] [CrossRef]
- Gilman, E.F.; Watson, D.G.; Klein, R.W.; Koeser, A.K.; Hilbert, D.R.; McLean, D.C. Lagerstroemia indica: Crapemyrtle. UF/IFAS Extension. 2018. ENH-501. Available online: https://edis.ifas.ufl.edu (accessed on 15 February 2019).
- Mohan Jain, S.; Brar, D.S. Molecular Techniques in Crop Improvement, 2nd ed.; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar] [CrossRef]
- Chen, C. Overview of plant pigments. In Pigments in Fruits and Vegetables; Springer: New York, NY, USA, 2015; pp. 1–7. [Google Scholar]
- Tanaka, Y.; Ohmiya, A. Seeing is believing: Engineering anthocyanin and carotenoid biosynthetic pathways. Curr. Opin. Biotechnol. 2008, 19, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Steele, C.L. Flavonoids and isoflavonoids—A gold mine for metabolic engineering. Trends Plant Sci. 1999, 4, 394–400. [Google Scholar] [CrossRef]
- Moyano, E.; Martinez-Garcia, J.F.; Martin, C. Apparent redundancy in myb gene function provides gearing for the control of flavonoid biosynthesis in antirrhinum flowers. Plant Cell 1996, 8, 1519–1532. [Google Scholar] [PubMed]
- To, K.Y.; Wang, C.K. Molecular breeding of flower color. In Floriculture Ornamental and Plant Biotechnology: Advances and Topical Issues Volume I; Silva, T.D., Ed.; Global Science Books: London, UK, 2006; pp. 300–310. [Google Scholar]
- Li, Z.; Zhao, M.; Jin, J.; Zhao, L.; Xu, Z. Anthocyanins and their biosynthetic genes in three novel-colored Rosa rugosa cultivars and their parents. Plant Physiol. Biochem. 2018, 129, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Li, Z.; Song, Y.; Zhu, H.; Lin, S.; Huang, R.; Jiang, Y.; Duan, X. LcNAC13 Physically Interacts with LcR1MYB1 to Coregulate Anthocyanin Biosynthesis-Related Genes during Litchi Fruit Ripening. Biomolecules 2019, 9, 135. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, A.; Brockman, A.; Aguirre, L.; Campbell, A.; Bean, A.; Cantero, A.; Gonzalez, A. Advances in the MYB–bHLH–WD repeat (MBW) pigment regulatory model: Addition of a WRKY factor and co-option of an anthocyanin MYB for betalain regulation. Plant Cell Physiol. 2017, 58, 1431–1441. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Lin-Wang, K.; Wang, H.; Gu, C.; Dare, A.P.; Espley, R.V.; He, H.; Allan, A.C.; Han, Y. Molecular genetics of blood-fleshed peach reveals activation of anthocyanin biosynthesis by NAC transcription factors. Plant J. 2015, 82, 105–121. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Tang, R.; Shi, X.; Wang, W.; Cao, Q.; Liu, X.; Wang, T.; Sun, Y.; Zhang, H.; Li, R.; et al. Uncovering anthocyanin biosynthesis related microRNAs and their target genes by small RNA and degradome sequencing in tuberous roots of sweet potato. BMC Plant Biol. 2019, 19, 232. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, H.; Lou, Q.; Liu, H.; Han, H.; Wang, Q.; Tang, Z.; Ma, Y.; Wang, H. Differential Regulation of Anthocyanins in Green and Purple Turnips Revealed by Combined De Novo Transcriptome and Metabolome Analysis. Int. J. Mol. Sci. 2019, 20, 4387. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Ji, Y.; Li, S.; Lu, L.; Tian, M.; Yang, W.; Li, H. Extensive Metabolic Profiles of Leaves and Stems from the Medicinal Plant Dendrobium officinale Kimura et Migo. Metabolites 2019, 9, 215. [Google Scholar] [CrossRef] [PubMed]
- Fosket, D.E. The Genetic Basis of Plant Development. In Fosket, Plant Growth and Development; Donald, E., Ed.; Academic Press: Cambridge, MA, USA, 1994; pp. 41–78. [Google Scholar] [CrossRef]
- Allan, A.C.; Hellens, R.P.; Laing, W.A. MYB transcription factors that colour our fruit. Trends Plant Sci. 2008, 13, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Xu, L.; Dossa, K.; Zhou, K.; Zhu, M.; Xie, H.; Tang, S.; Yu, Y.; Guo, X.; Zhou, B. Identification of putative drought-responsive genes in rice using gene co-expression analysis. Bioinformation 2019, 15, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Hsu, A.K.; Sajdak, J.; Qin, J.; Pavlidis, P. Overexpression analysis of human genes across many microarray data sets. Genome Res. 2004, 14, 105–1094. [Google Scholar] [CrossRef] [PubMed]
- Veberic, R.; Slatnar, A.; Bizjak, J.; Stampar, F.; Mikulic-Petkovsek, M. Anthocyanin composition of different wild and cultivated berry species. LWT Food Sci. Technol. 2015, 60, 509–517. [Google Scholar] [CrossRef]
- Kong, J.M.; Chia, L.S.; Goh, N.K.; Chia, T.F.; Brouillard, R. Analysis and biological activities of anthocyanins. Phytochemistry 2003, 64, 923–933. [Google Scholar] [CrossRef]
- Egolf, D.R.; Santamour, F.S. Anthocyanin pigments and breeding potential in crape myrtle (Lagerstreomia indica L.) and rose of Sharon (Hibiscus syriacus L.). HortScience 1975, 10, 223–224. [Google Scholar]
- Toki, K. Anthocyanin pigments and breeding potential of blue flowers in Lagerstreomia indica. BioHort 1989, 73–77. [Google Scholar]
- Toki, K.; Katsuyama, N. Pigments and color variation in flowers of Lagerstroemia indica. J. Jpn. Soc. Hortic. Sci. 1995, 63, 853–861. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.-S.; Gao, J.-M.; Shu, Q.-Y.; Li, C.-H.; Yao, J.; Hao, Q.; Zhang, J.-J. Determination of Anthocyanins and Exploration of Relationship between Their Composition and Petal Coloration in Crape Myrtle (Lagerstroemia hybrid). J. Integr. Plant Biol. 2008, 50, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, S.; Craig, T.; de Magalhães, J.P. GeneFriends: A human RNA-seq-based gene and transcript co-expression database. Nucleic Acids Res. 2015, 43, D1124–D1132. [Google Scholar]
- Zhang, Z.Y.; Wang, P.; Li, Y.; Ma, L.L.; Li, L.F.; Yang, R.T.; Ma, Y.Z.; Wang, S.; Wang, Q. Global transcriptome analysis and identification of the flowering regulatory genes expressed in leaves of Lagerstroemia indica. DNA Cell Biol. 2014, 33, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shi, W.; Rinehart, T. Transcriptomes That Confer to Plant Defense against Powdery Mildew Disease in Lagerstroemia indica. Int. J. Genom. 2015. [Google Scholar] [CrossRef] [PubMed]
- Naing, A.H.; Kim, C.K. Roles of R2R3-MYB transcription factors in transcriptional regulation of anthocyanin biosynthesis in horticultural plants. Plant Mol. Biol. 2018, 98, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mao, Y.; Liu, H.; Yu, F.; Li, S.; Yin, T. Transcriptome analysis of differentially expressed genes relevant to variegation in peach flowers. PLoS ONE 2014, 9, e90842. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Ma, R.J.; Shen, Z.J.; Yan, J.; Yu, M.L. Gene regulation of anthocyanin biosynthesis in two blood-flesh peach (Prunus persica (L) Batsch) cultivars during fruit development. J. Zhejiang Univ. Sci. B 2014, 15, 809–819. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Pan, Q.H.; She, Y.; Duan, C.Q. Biosynthesis and genetic regulation of proanthocyanidins in plants. Molecules 2008, 13, 2674–2703. [Google Scholar] [CrossRef] [PubMed]
- Hellström, J.; Mattila, P.; Karjalainen, R.O. Stability of anthocyanins in berry juices stored at different temperatures. J. Food Compos. Anal. 2013, 31, 12–19. [Google Scholar] [CrossRef]
- Ono, E.; Homma, Y.; Horikawa, M.; Kunikane-Doi, S.; Imai, H.; Takahashi, S.; Kawai, Y.; Ishiguro, M.; Fukui, Y.; Nakayama, T. Functional differentiation of the glycosyltransferases that contribute to the chemical diversity of bioactive flavonol glycosides in grapevines (Vitis vinifera). Plant Cell 2010, 22, 2856–2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Wei, G.; Zhou, H.; Gu, C.; Vimolmangkang, S.; Liao, L.; Han, Y.P. Unraveling the mechanism underlying the glycosylation and methylation of anthocyanins in peach. Plant Physiol. 2014, 166, 1044–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangan, S.; Alon, U. Structure and function of the feed-forward loop network motif. Proc. Natl. Acad. Sci. USA 2003, 100, 11980–11985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Li, X.; Zheng, J. Transcriptome profiling using Illumina- and SMRT-based RNAseq of hot pepper for in-depth understanding of genes involved in CMV infection. Gene 2018, 666, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full length transcriptome assembly from RNA Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A. The COG database: A tool for genome scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2013, 42, 222–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The Universal Protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Cepas, J.; Szklarczyk, D.; Forslund, K.; Cook, H.; Heller, D.; Walter, M.C.; Rattei, T.; Mende, D.R.; Sunagawa, S.; Kuhn, M.; et al. eggNOG 4.5: A hierarchical orthology framework with improved functional annotations for eukaryotic, prokaryotic and viral sequences. Nucleic Acids Res. 2015, 44, 286–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.Y.; Li, J.Q.; Wu, S.F.; Zhu, Y.P.; Chen, Y.W.; He, F.C. Integrated nr Database in Protein Annotation System and Its Localization. Comput. Eng. 2006, 32, 71–74. [Google Scholar]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Krylov, D.M.; Makarova, K.S.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; Rao, B.S.; et al. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome Biol. 2004, 5, R7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, S.R. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Colin, N.D. RSEM: Accurate transcript quantification from RNA Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, G.; Morris, J.H.; Demchak, B.; Bader, G.D. Biological network exploration with Cytoscape 3. Curr. Protoc. Bioinform. 2014, 47, 1–24. [Google Scholar]
- Dossa, K.; Mmadi, M.A.; Zhou, R.; Zhang, T.; Su, R.; Zhang, Y.; Wang, L.; You, J.; Zhang, X. Depicting the Core Transcriptome Modulating Multiple Abiotic Stresses Responses in Sesame (Sesamum indicum L.). Int. J. Mol. Sci. 2019, 20, 3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dossa, K.; Mmadi, M.A.; Zhou, R.; Zhou, Q.; Yang, M.; Cisse, N.; Diouf, D.; Wang, L.; Zhang, X. The contrasting response to drought and waterlogging is underpinned by divergent DNA methylation programs associated with transcript accumulation in sesame. Plant Sci. 2018, 277, 207–217. [Google Scholar]
- Lalitha, S. Primer premier 5. Biotechnol. Softw. Internet Rep. 2000, 1, 270–272. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | HD-1 | ZD-1 | HD-2 | ZD-2 | Log2 Fold Change | |||
|---|---|---|---|---|---|---|---|---|
| HD-1_vs_HD-2 | ZD-1_vs_ZD-2 | ZD-1_vs_HD-1 | ZD-2_vs_HD-2 | |||||
| Peonidin O-hexoside | 24257333.3 | 13935333.3 | 2037100 | 12121666.7 | −3.57 | −0.20 | 0.80 | −2.57 |
| Rosinidin O-hexoside | 89211.3333 | 144180 | 0 | 43131 | −13.28 | −1.74 | −0.69 | −12.23 |
| Cyanidin O-syringic acid | 2205133.33 | 4702233.33 | 144803.333 | 2828266.67 | −3.93 | −0.73 | −1.09 | −4.29 |
| Cyanidin 3-O-glucoside | 7512833.33 | 17313666.7 | 443633.333 | 8068766.67 | −4.08 | −1.10 | −1.20 | −4.18 |
| Delphinidin 3-O-glucoside | 3606100 | 5734733.33 | 2009400 | 5926300 | –0.84 | 0.05 | −0.67 | −1.56 |
| Cyanidin 3,5-O-diglucoside | 4405800 | 13112000 | 394216.667 | 7217400 | −3.48 | −0.86 | −1.57 | −4.19 |
| Cyanidin | 801780 | 0 | 484516.667 | 0 | −0.73 | 0.00 | 16.44 | 15.72 |
| SampleID. | Raw Read Number | Base Number | GC (%) | Q20 (%) | Q30 (%) |
|---|---|---|---|---|---|
| HD-1_1 | 29963667 | 8930322034 | 51.21 | 97.91 | 94.18 |
| HD-2_1 | 29373060 | 8751104486 | 51.26 | 97.85 | 94.09 |
| HD-2_2 | 20168830 | 6023966824 | 50.7 | 97.82 | 93.98 |
| HD-2_3 | 23631048 | 7060390512 | 50.81 | 97.83 | 94 |
| HD-1_2 | 23919615 | 7129091888 | 51.23 | 98 | 94.41 |
| HD-1_2 | 23467265 | 7003557242 | 50.74 | 97.88 | 94.23 |
| ZD-1_1 | 24273602 | 7232878206 | 51.32 | 97.69 | 93.8 |
| ZD-1_2 | 21891679 | 6530271554 | 51.24 | 97.97 | 94.22 |
| ZD-1_3 | 22505602 | 6715319880 | 51.25 | 97.88 | 94.12 |
| ZD-2_1 | 21339681 | 6361383236 | 51.24 | 97.85 | 93.99 |
| ZD-2_2 | 21255491 | 6336179806 | 51.32 | 97.8 | 93.93 |
| ZD-2_3 | 22210402 | 6625694066 | 51.27 | 97.88 | 94.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, Z.; Liu, S.; Zeng, H.; Li, Y.; Wang, X.; Chen, Y.; Wang, X.; Cai, N. Exploring the Molecular Mechanism underlying the Stable Purple-Red Leaf Phenotype in Lagerstroemia indica cv. Ebony Embers. Int. J. Mol. Sci. 2019, 20, 5636. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225636
Qiao Z, Liu S, Zeng H, Li Y, Wang X, Chen Y, Wang X, Cai N. Exploring the Molecular Mechanism underlying the Stable Purple-Red Leaf Phenotype in Lagerstroemia indica cv. Ebony Embers. International Journal of Molecular Sciences. 2019; 20(22):5636. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225636
Chicago/Turabian StyleQiao, Zhongquan, Sisi Liu, Huijie Zeng, Yongxin Li, Xiangying Wang, Yi Chen, Xiaoming Wang, and Neng Cai. 2019. "Exploring the Molecular Mechanism underlying the Stable Purple-Red Leaf Phenotype in Lagerstroemia indica cv. Ebony Embers" International Journal of Molecular Sciences 20, no. 22: 5636. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225636