A2AR Transmembrane 2 Peptide Administration Disrupts the A2AR-A2AR Homoreceptor but Not the A2AR-D2R Heteroreceptor Complex: Lack of Actions on Rodent Cocaine Self-Administration

 , ,
, ,
Abstract
:1. Introduction
2. Results
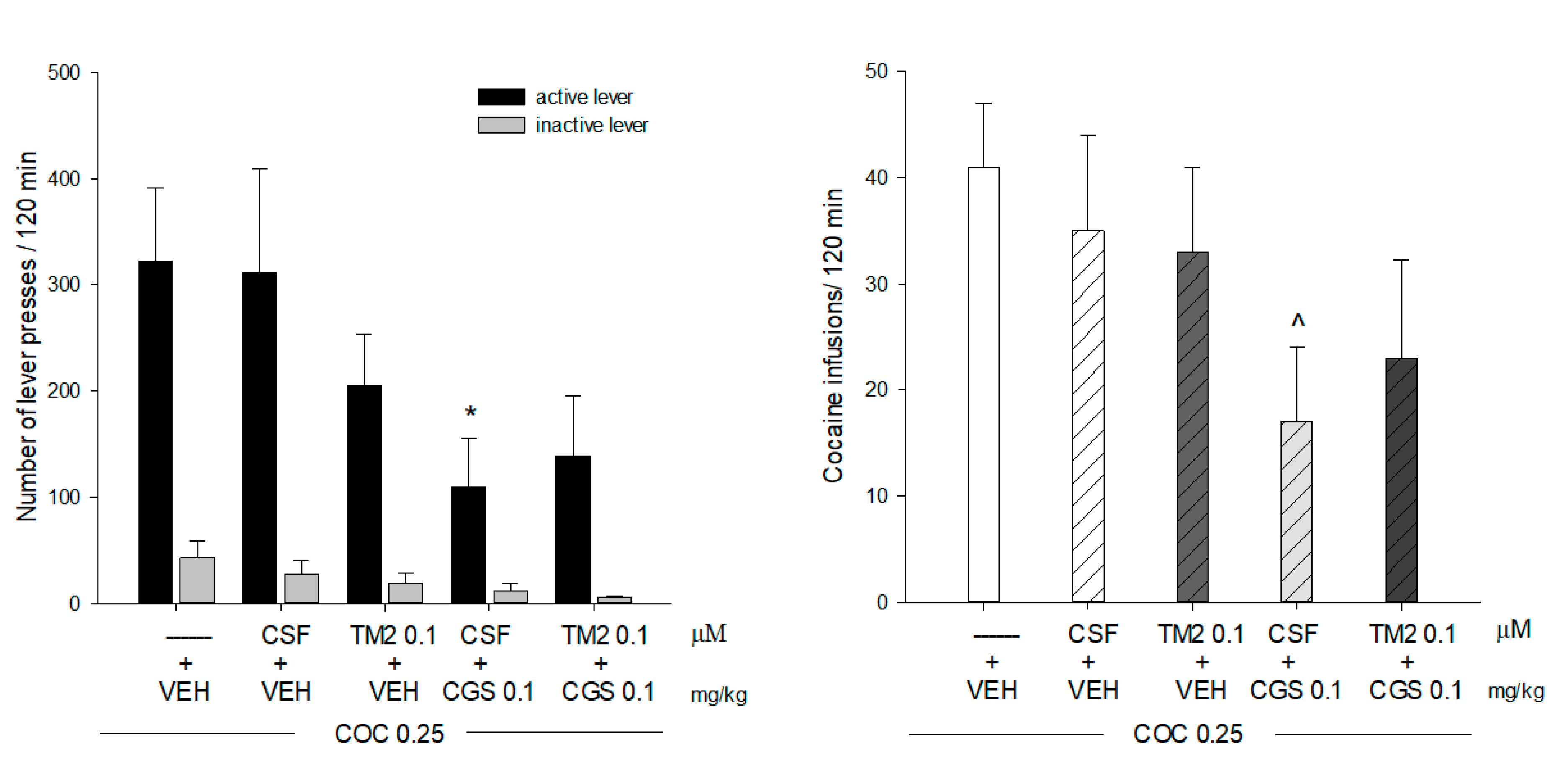
2.1. Effects of Intra-Accumbal Microinjections of Rat A2AR TM2 Peptide on Cocaine Self-Administration
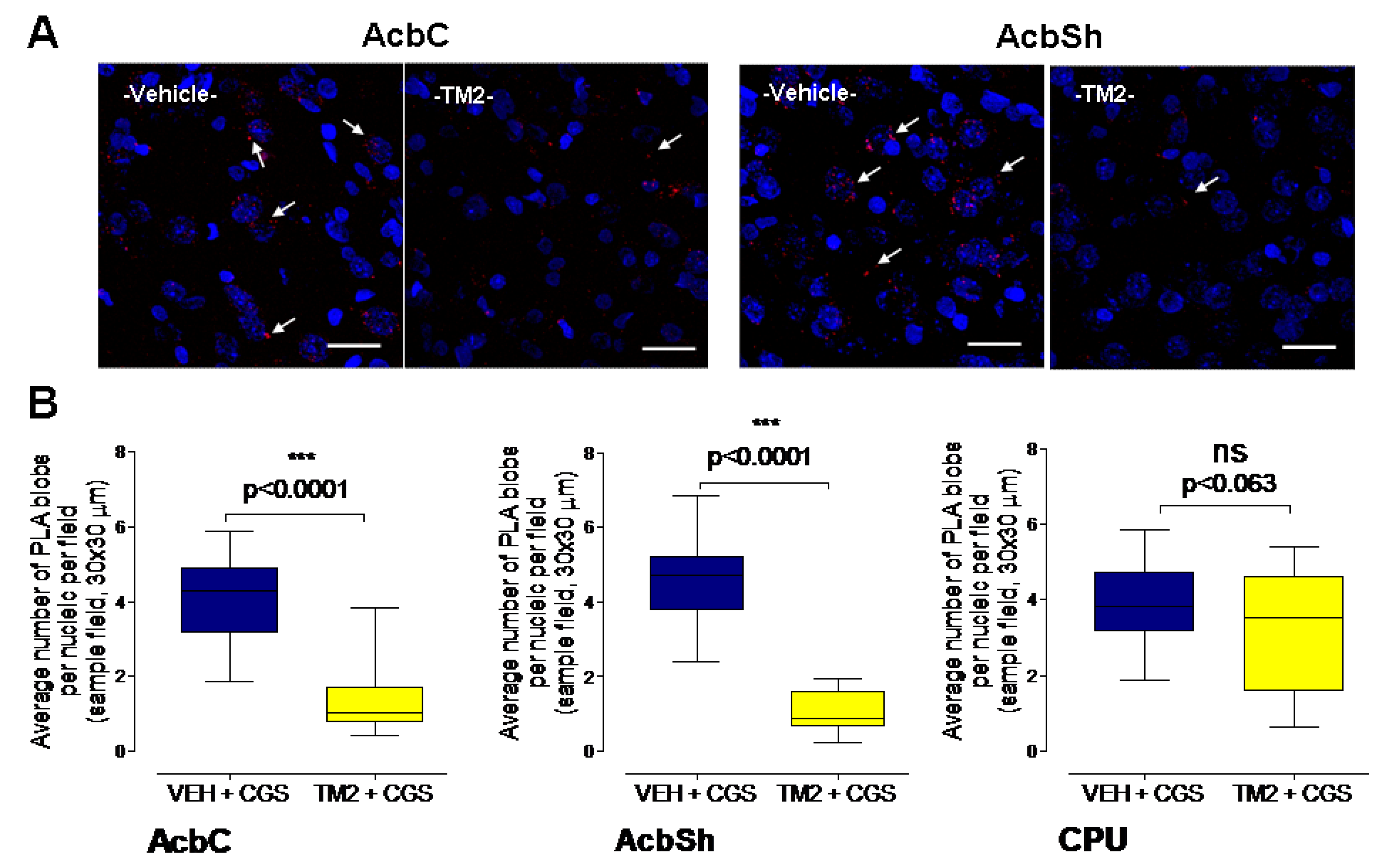
2.2. Effects of Intra Accumbal Microinjections of Rat A2AR TM2 Peptide on A2AR-D2R Heteroreceptor Complexes in the Nucleus Accumbens Using In Situ PLA
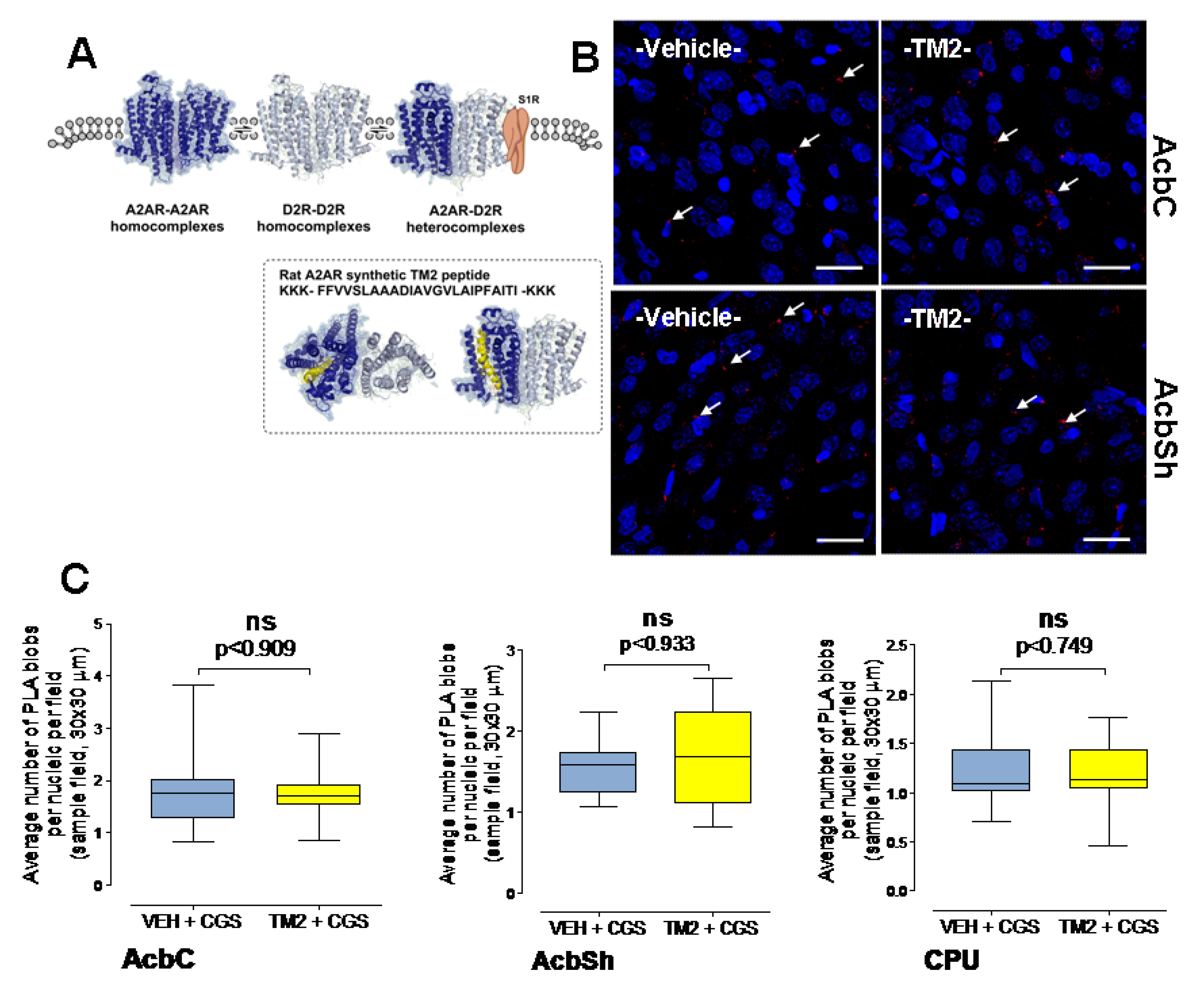
2.3. Effects of Intra Accumbal Microinjections of Rat A2AR TM2 Peptide on A2AR-A2AR Homoreceptor Complexes in the Nucleus Accumbens Using In Situ PLA
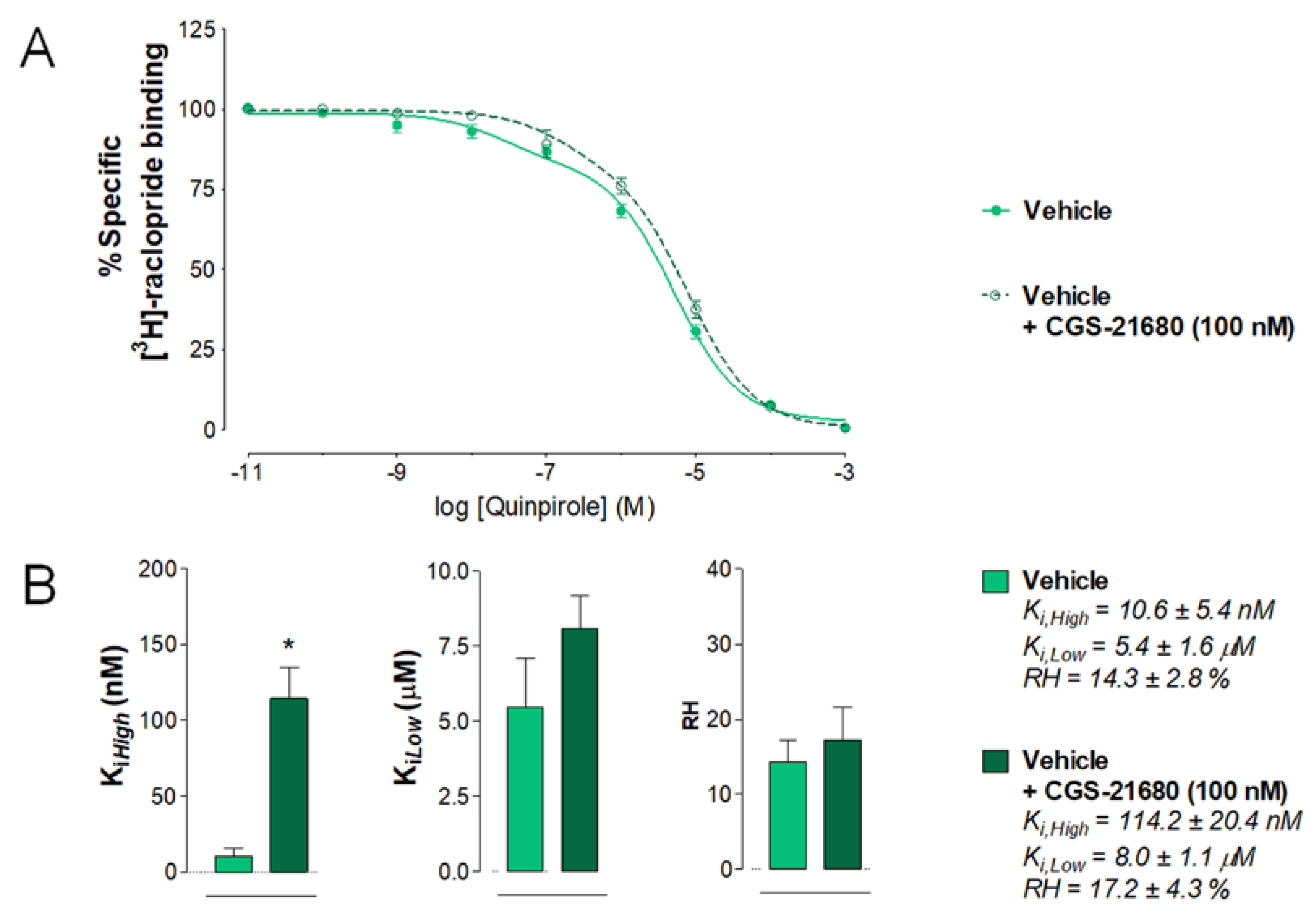
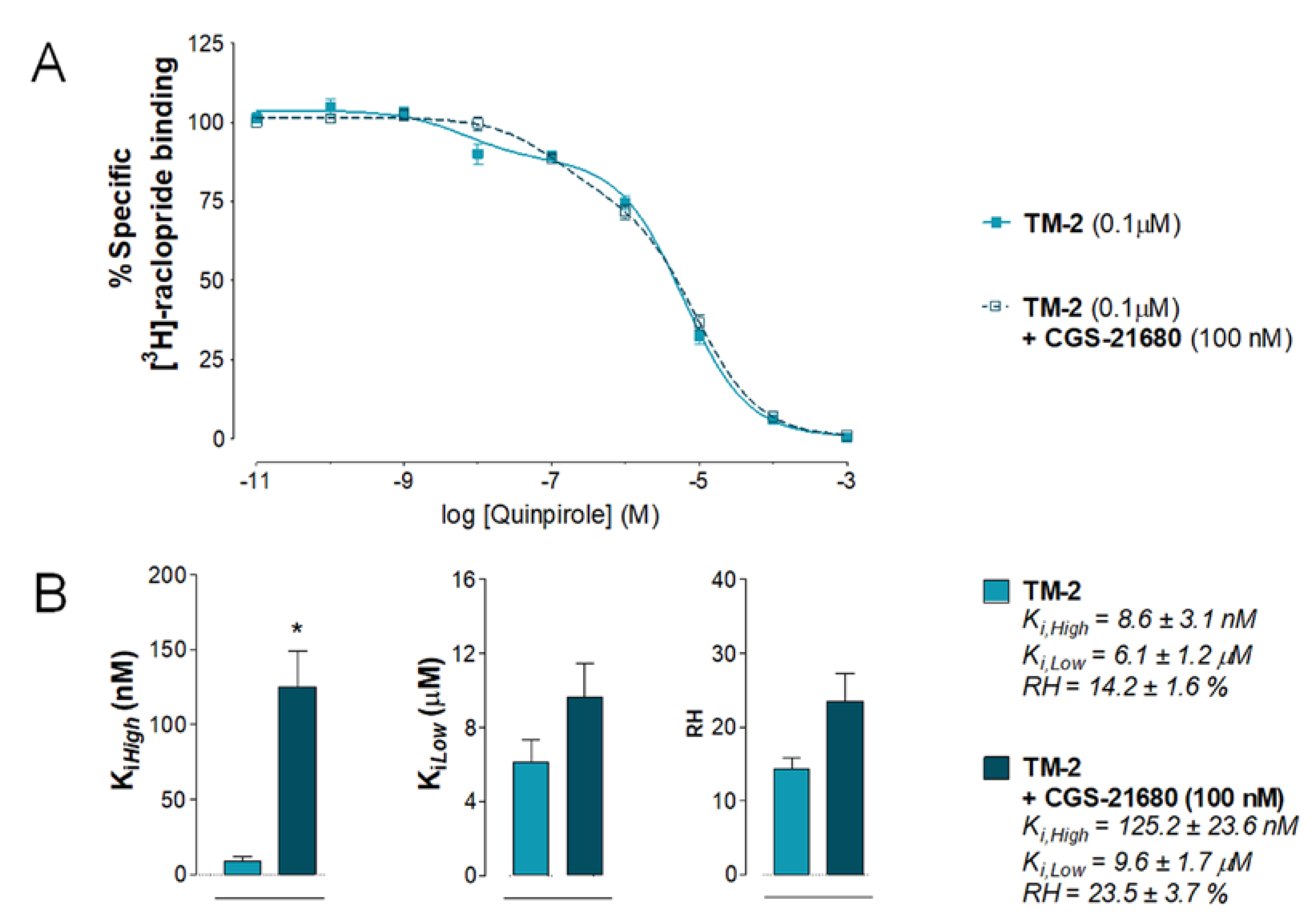
2.4. Effects of Intra Accumbal Microinjections of Rat A2AR TM2 Peptide on the Antagonistic A2AR-D2R Interactions in the Ventral Striatum Using [3H]-Raclopride/Quinpirole Competition Binding Assay
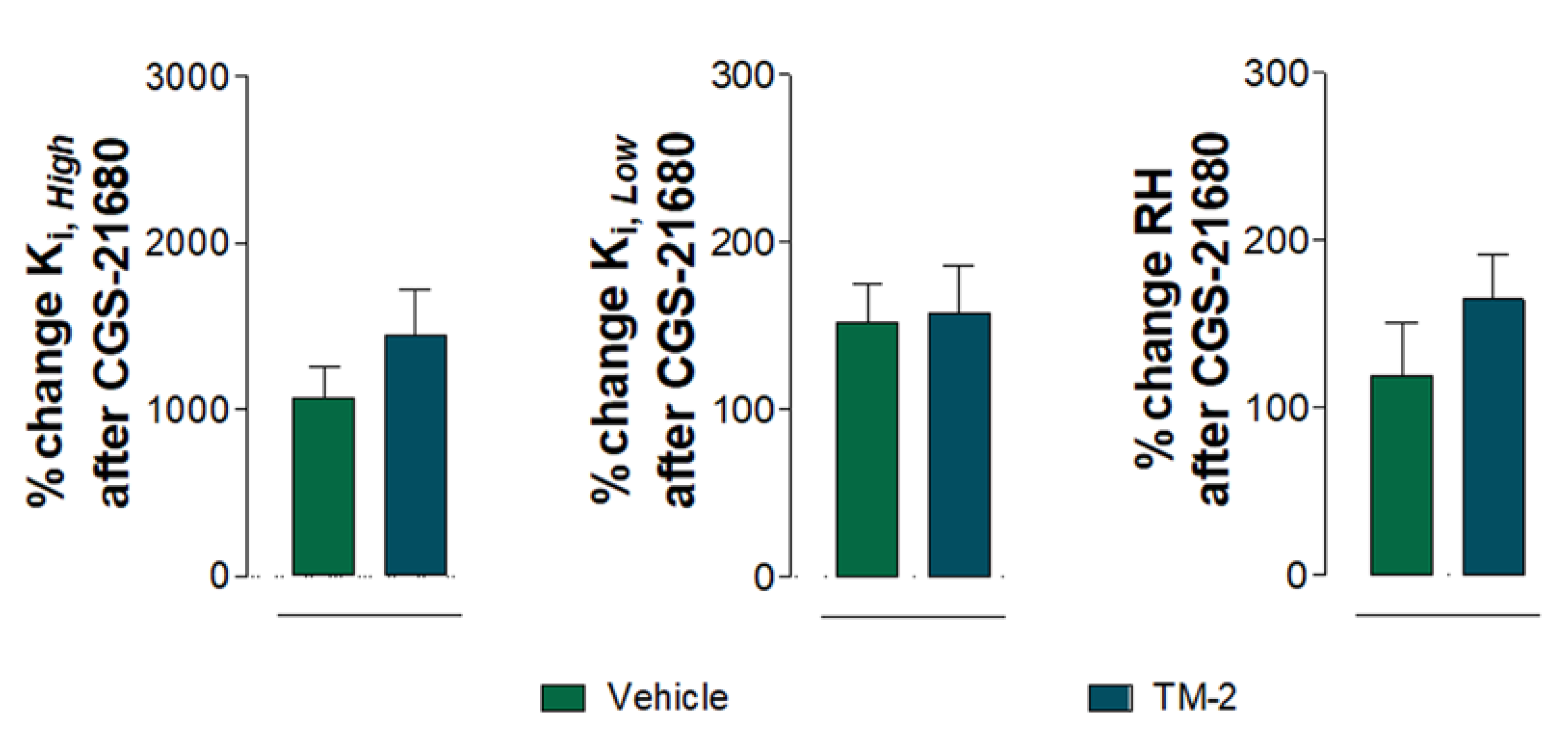
2.4.1. Vehicle Group Treated with CGS 21680
2.4.2. A2AR TM2 Groups Treated with CGS 21680
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Surgery
4.4. Cocaine Self-Administration
4.5. In Situ Proximity Ligation Assay (In Situ PLA)
4.6. Biochemical Binding Experiments
4.6.1. Membrane Preparation
4.6.2. [3H]-Raclopride Competition Binding Experiments
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| A2AR | Adenosine A2A receptor subtype |
| D2R | Dopamine D2 receptor subtype |
| GPCR | G protein-coupled receptor |
| PLA | Proximity ligation assay |
| TM | Transmembrane |
References
- Zoli, M.; Agnati, L.F.; Hedlund, P.B.; Li, X.M.; Ferre, S.; Fuxe, K. Receptor-receptor interactions as an integrative mechanism in nerve cells. Mol. Neurobiol. 1993, 7, 293–334. [Google Scholar] [CrossRef] [PubMed]
- Ferre, S.; von Euler, G.; Johansson, B.; Fredholm, B.B.; Fuxe, K. Stimulation of high-affinity adenosine A2 receptors decreases the affinity of dopamine D2 receptors in rat striatal membranes. Proc. Natl. Acad. Sci. USA 1991, 88, 7238–7241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuxe, K.; Ungerstedt, U. Action of caffeine and theophyllamine on supersensitive dopamine receptors: Considerable enhancement of receptor response to treatment with dopa and dopamine receptor agonists. Med. Biol. 1974, 52, 48–54. [Google Scholar] [PubMed]
- Ciruela, F.; Burgueno, J.; Casado, V.; Canals, M.; Marcellino, D.; Goldberg, S.R.; Bader, M.; Fuxe, K.; Agnati, L.F.; Lluis, C.; et al. Combining mass spectrometry and pull-down techniques for the study of receptor heteromerization. Direct epitope-epitope electrostatic interactions between adenosine A2A and dopamine D2 receptors. Anal. Chem. 2004, 76, 5354–5363. [Google Scholar] [CrossRef]
- Ciruela, F.; Ferre, S.; Casado, V.; Cortes, A.; Cunha, R.A.; Lluis, C.; Franco, R. Heterodimeric adenosine receptors: A device to regulate neurotransmitter release. Cell. Mol. Life Sci. 2006, 63, 2427–2431. [Google Scholar] [CrossRef] [Green Version]
- Azdad, K.; Gall, D.; Woods, A.S.; Ledent, C.; Ferre, S.; Schiffmann, S.N. Dopamine D2 and adenosine A2A receptors regulate nmda-mediated excitation in accumbens neurons through A2A-D2 receptor heteromerization. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2009, 34, 972–986. [Google Scholar] [CrossRef] [Green Version]
- Schiffmann, S.N.; Fisone, G.; Moresco, R.; Cunha, R.A.; Ferre, S. Adenosine A2A receptors and basal ganglia physiology. Prog. Neurobiol. 2007, 83, 277–292. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Carlsson, J.; Ambrogini, P.; Narvaez, M.; Wydra, K.; Tarakanov, A.O.; Li, X.; Millon, C.; Ferraro, L.; Cuppini, R.; et al. Understanding the role of gpcr heteroreceptor complexes in modulating the brain networks in health and disease. Front. Cell. Neurosci. 2017, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Fuxe, K.; Borroto-Escuela, D.O.; Romero-Fernandez, W.; Palkovits, M.; Tarakanov, A.O.; Ciruela, F.; Agnati, L.F. Moonlighting proteins and protein-protein interactions as neurotherapeutic targets in the g protein-coupled receptor field. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2014, 39, 131–155. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Tarakanov, A.O.; Gomez-Soler, M.; Corrales, F.; Marcellino, D.; Narvaez, M.; Frankowska, M.; Flajolet, M.; Heintz, N.; et al. Characterization of the A2AR-D2R interface: Focus on the role of the c-terminal tail and the transmembrane helices. Biochem. Biophys. Res. Commun. 2010, 402, 801–807. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Rodriguez, D.; Romero-Fernandez, W.; Kapla, J.; Jaiteh, M.; Ranganathan, A.; Lazarova, T.; Fuxe, K.; Carlsson, J. Mapping the interface of a gpcr dimer: A structural model of the A2A adenosine and D2 dopamine receptor heteromer. Front. Pharmacol. 2018, 9, 829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borroto-Escuela, D.O.; Perez De La Mora, M.; Manger, P.; Narvaez, M.; Beggiato, S.; Crespo-Ramirez, M.; Navarro, G.; Wydra, K.; Diaz-Cabiale, Z.; Rivera, A.; et al. Brain dopamine transmission in health and parkinson’s disease: Modulation of synaptic transmission and plasticity through volume transmission and dopamine heteroreceptors. Front. Synaptic Neurosci. 2018, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Ferre, S.; Zoli, M.; Agnati, L.F. Integrated events in central dopamine transmission as analyzed at multiple levels. Evidence for intramembrane adenosine A2A/dopamine D2 and adenosine A1/dopamine D1 receptor interactions in the basal ganglia. Brain Res. Rev. 1998, 26, 258–273. [Google Scholar] [CrossRef]
- Navarro, G.; Moreno, E.; Aymerich, M.; Marcellino, D.; McCormick, P.J.; Mallol, J.; Cortes, A.; Casado, V.; Canela, E.I.; Ortiz, J.; et al. Direct involvement of sigma-1 receptors in the dopamine D1 receptor-mediated effects of cocaine. Proc. Natl. Acad. Sci. USA 2010, 107, 18676–18681. [Google Scholar] [CrossRef] [Green Version]
- Navarro, G.; Moreno, E.; Bonaventura, J.; Brugarolas, M.; Farre, D.; Aguinaga, D.; Mallol, J.; Cortes, A.; Casado, V.; Lluis, C.; et al. Cocaine inhibits dopamine D2 receptor signaling via sigma-1-D2 receptor heteromers. PLoS ONE 2013, 8, e61245. [Google Scholar] [CrossRef] [Green Version]
- Pinton, L.; Borroto-Escuela, D.O.; Narváez, M.; Jiménez-Beristain, A.; Oflijan, J.; Ferraro, L.; Agnati, L.F.; Fuxe, K. Dopamine D2 receptor dynamic and modulation in the D2R-Sigma 1 heteroreceptor complexes: Role in cocaine actions. In European Neuropsychopharmacology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 25, pp. S609–S610. [Google Scholar]
- Pintsuk, J.; Borroto-Escuela, D.O.; Pomierny, B.; Wydra, K.; Zaniewska, M.; Filip, M.; Fuxe, K. Cocaine self-administration differentially affects allosteric A2A-D2 receptor-receptor interactions in the striatum. Relevance for cocaine use disorder. Pharmacol. Biochem. Behav. 2016, 144, 85–91. [Google Scholar] [CrossRef]
- Beggiato, S.; Borelli, A.C.; Borroto-Escuela, D.; Corbucci, I.; Tomasini, M.C.; Marti, M.; Antonelli, T.; Tanganelli, S.; Fuxe, K.; Ferraro, L. Cocaine modulates allosteric D2-sigma1 receptor-receptor interactions on dopamine and glutamate nerve terminals from rat striatum. Cell. Signal. 2017, 40, 116–124. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Narvaez, M.; Wydra, K.; Pintsuk, J.; Pinton, L.; Jimenez-Beristain, A.; Di Palma, M.; Jastrzebska, J.; Filip, M.; Fuxe, K. Cocaine self-administration specifically increases A2AR-D2R and D2R-sigma1R heteroreceptor complexes in the rat nucleus accumbens shell. Relevance for cocaine use disorder. Pharmacol. Biochem. Behav. 2017, 155, 24–31. [Google Scholar] [CrossRef]
- Hasbi, A.; Perreault, M.L.; Shen, M.Y.F.; Fan, T.; Nguyen, T.; Alijaniaram, M.; Banasikowski, T.J.; Grace, A.A.; O’Dowd, B.F.; Fletcher, P.J.; et al. Activation of dopamine D1-D2 receptor complex attenuates cocaine reward and reinstatement of cocaine-seeking through inhibition of darpp-32, erk, and deltafosb. Front. Pharmacol. 2017, 8, 924. [Google Scholar] [CrossRef]
- Borroto-Escuela, D. Acute cocaine enhances dopamine D2R recognition and signalling and counteracts D2R internalization in sigma1R-D2R heteroreceptor complexes. Mol. Neurobiol. (Accept. Revis.) 2019, 56, 7045–7055. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Wydra, K.; Filip, M.; Fuxe, K. A2AR-D2R heteroreceptor complexes in cocaine reward and addiction. Trends Pharmacol. Sci. 2018, 39, 1008–1020. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Wydra, K.; Li, X.; Rodriguez, D.; Carlsson, J.; Jastrzebska, J.; Filip, M.; Fuxe, K. Disruption of A2AR-D2R heteroreceptor complexes after A2AR transmembrane 5 peptide administration enhances cocaine self-administration in rats. Mol. Neurobiol. 2018, 55, 7038–7048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borroto-Escuela, D.O.; Hagman, B.; Woolfenden, M.; Pinton, L.; Jiménez-Beristain, A.; Oflijan, J.; Narvaez, M.; Di Palma, M.; Feltmann, K.; Sartini, S.; et al. In situ proximity ligation assay to study and understand the distribution and balance of gpcr homo- and heteroreceptor complexes in the brain. In Receptor and Ion Channel Detection in the Brain; Lujan, R., Ciruela, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 110, pp. 109–126. [Google Scholar]
- Fuxe, K.; Borroto-Escuela, D.O. Heteroreceptor complexes and their allosteric receptor-receptor interactions as a novel biological principle for integration of communication in the cns: Targets for drug development. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2016, 41, 380–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canals, M.; Burgueno, J.; Marcellino, D.; Cabello, N.; Canela, E.I.; Mallol, J.; Agnati, L.; Ferre, S.; Bouvier, M.; Fuxe, K.; et al. Homodimerization of adenosine A2A receptors: Qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J. Neurochem. 2004, 88, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Brito, I.; Romero-Fernandez, W.; Di Palma, M.; Oflijan, J.; Skieterska, K.; Duchou, J.; Van Craenenbroeck, K.; Suarez-Boomgaard, D.; Rivera, A.; et al. The g protein-coupled receptor heterodimer network (gpcr-hetnet) and its hub components. Int. J. Mol. Sci. 2014, 15, 8570–8590. [Google Scholar] [CrossRef] [PubMed]
- Cabello, N.; Gandia, J.; Bertarelli, D.C.; Watanabe, M.; Lluis, C.; Franco, R.; Ferre, S.; Lujan, R.; Ciruela, F. Metabotropic glutamate type 5, dopamine D2 and adenosine A2A receptors form higher-order oligomers in living cells. J. Neurochem. 2009, 109, 1497–1507. [Google Scholar] [CrossRef]
- Marcellino, D.; Carriba, P.; Filip, M.; Borgkvist, A.; Frankowska, M.; Bellido, I.; Tanganelli, S.; Muller, C.E.; Fisone, G.; Lluis, C.; et al. Antagonistic cannabinoid CB1/dopamine D2 receptor interactions in striatal CB1/D2 heteromers. A combined neurochemical and behavioral analysis. Neuropharmacology 2008, 54, 815–823. [Google Scholar] [CrossRef]
- Wydra, K.; Suder, A.; Borroto-Escuela, D.O.; Filip, M.; Fuxe, K. On the role of A(2)A and D(2) receptors in control of cocaine and food-seeking behaviors in rats. Psychopharmacology 2015, 232, 1767–1778. [Google Scholar] [CrossRef] [Green Version]
- Wydra, K.; Golembiowska, K.; Zaniewska, M.; Kaminska, K.; Ferraro, L.; Fuxe, K.; Filip, M. Accumbal and pallidal dopamine, glutamate and gaba overflow during cocaine self-administration and its extinction in rats. Addict. Biol. 2013, 18, 307–324. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Flajolet, M.; Agnati, L.F.; Greengard, P.; Fuxe, K. Bioluminescence resonance energy transfer methods to study g protein-coupled receptor-receptor tyrosine kinase heteroreceptor complexes. Methods Cell Biol. 2013, 117, 141–164. [Google Scholar]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Mudo, G.; Perez-Alea, M.; Ciruela, F.; Tarakanov, A.O.; Narvaez, M.; Di Liberto, V.; Agnati, L.F.; Belluardo, N.; et al. Fibroblast growth factor receptor 1-5-hydroxytryptamine 1a heteroreceptor complexes and their enhancement of hippocampal plasticity. Biol. Psychiatry 2012, 71, 84–91. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | No. Rats | Total Cocaine Intake (mg/rat) [Mean ± S.E.M] |
|---|---|---|
| Vehicle (intra accumbal) + Vehicle (i.p.) + cocaine (i.v.) | 10 | 173 ± 26 |
| TM2 (intra accumbal) + Vehicle (i.p.) + cocaine (i.v.) | 9 | 224 ± 49 |
| Vehicle (intra accumbal) + CGS 21680 (i.p.) + cocaine (i.v.) | 7 | 213 ± 24 |
| TM2 (intra accumbal) + CGS 21680 (i.p.) + cocaine (i.v.) | 7 | 174 ± 16 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borroto-Escuela, D.O.; Wydra, K.; Romero-Fernandez, W.; Zhou, Z.; Frankowska, M.; Filip, M.; Fuxe, K. A2AR Transmembrane 2 Peptide Administration Disrupts the A2AR-A2AR Homoreceptor but Not the A2AR-D2R Heteroreceptor Complex: Lack of Actions on Rodent Cocaine Self-Administration. Int. J. Mol. Sci. 2019, 20, 6100. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20236100
Borroto-Escuela DO, Wydra K, Romero-Fernandez W, Zhou Z, Frankowska M, Filip M, Fuxe K. A2AR Transmembrane 2 Peptide Administration Disrupts the A2AR-A2AR Homoreceptor but Not the A2AR-D2R Heteroreceptor Complex: Lack of Actions on Rodent Cocaine Self-Administration. International Journal of Molecular Sciences. 2019; 20(23):6100. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20236100
Chicago/Turabian StyleBorroto-Escuela, Dasiel O., Karolina Wydra, Wilber Romero-Fernandez, Zilong Zhou, Malgorzata Frankowska, Malgorzata Filip, and Kjell Fuxe. 2019. "A2AR Transmembrane 2 Peptide Administration Disrupts the A2AR-A2AR Homoreceptor but Not the A2AR-D2R Heteroreceptor Complex: Lack of Actions on Rodent Cocaine Self-Administration" International Journal of Molecular Sciences 20, no. 23: 6100. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20236100