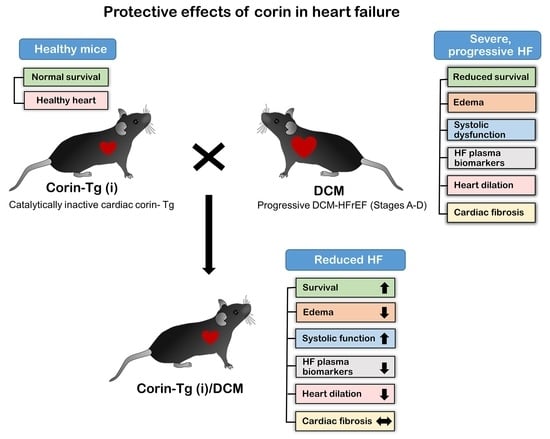
Cardiac-Specific Overexpression of Catalytically Inactive Corin Reduces Edema, Contractile Dysfunction, and Death in Mice with Dilated Cardiomyopathy

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
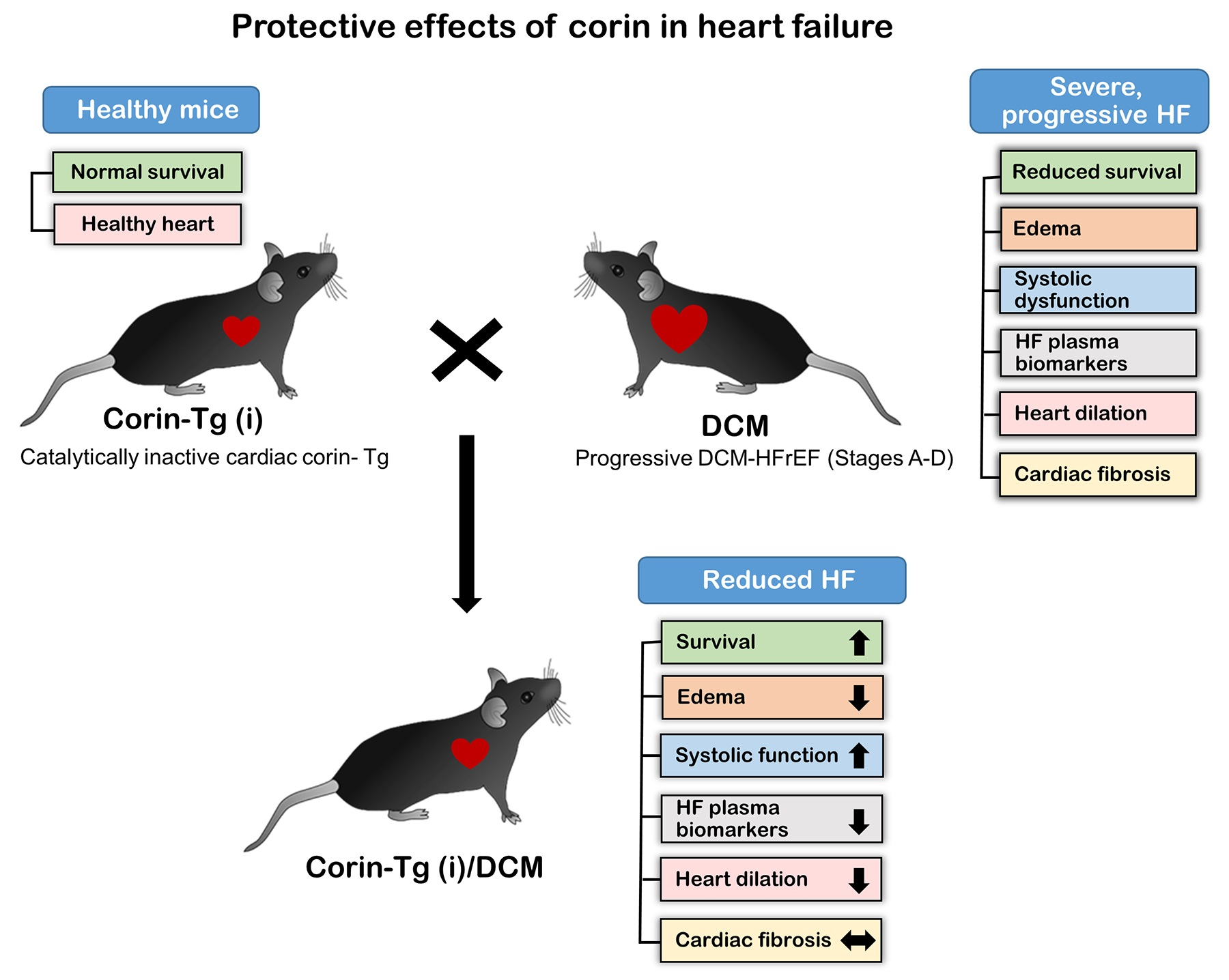
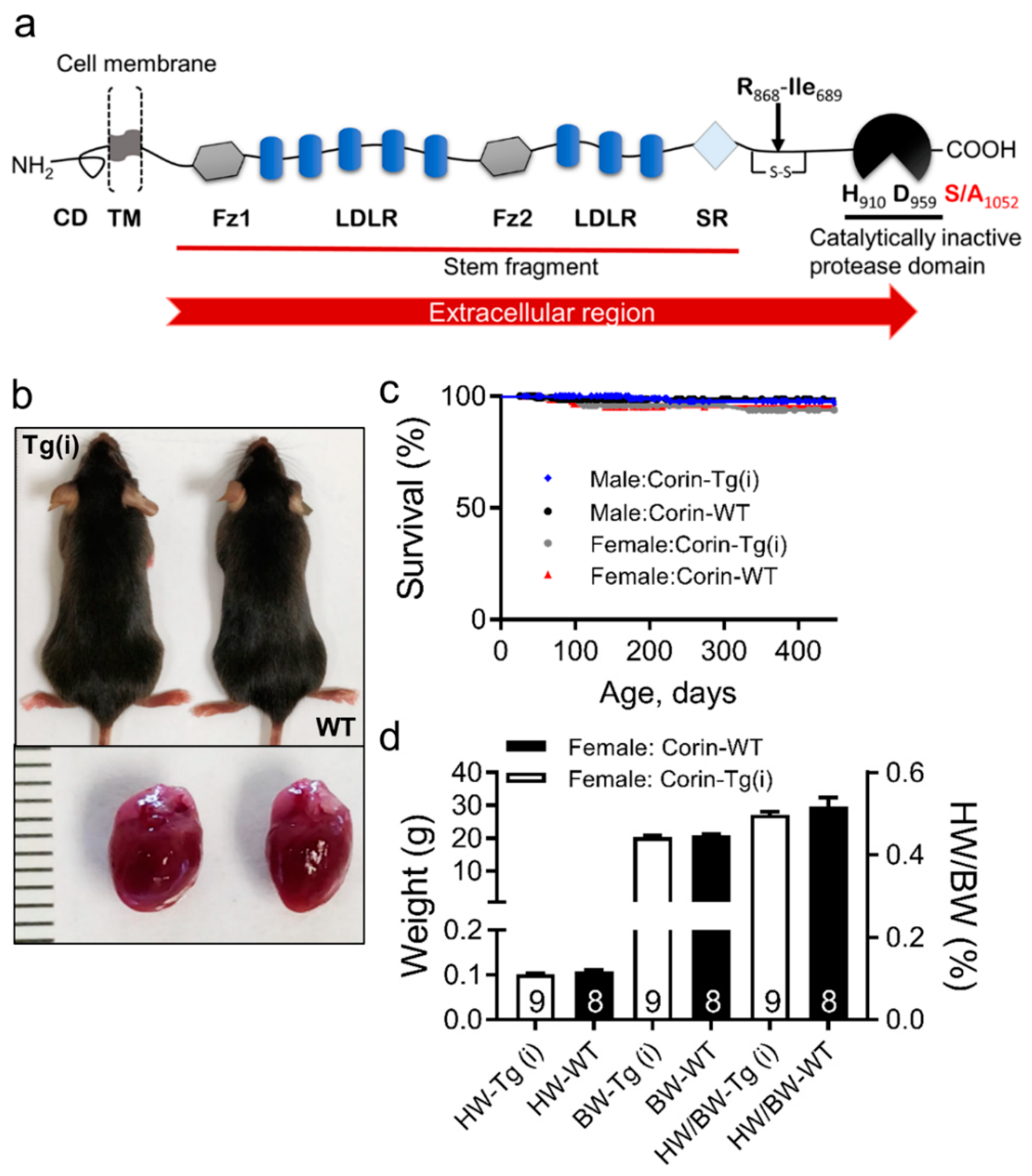
2.1. Effects of Overexpression of Catalytically Inactive Cardiac Corin-Tg(i) onWild-Type Mice
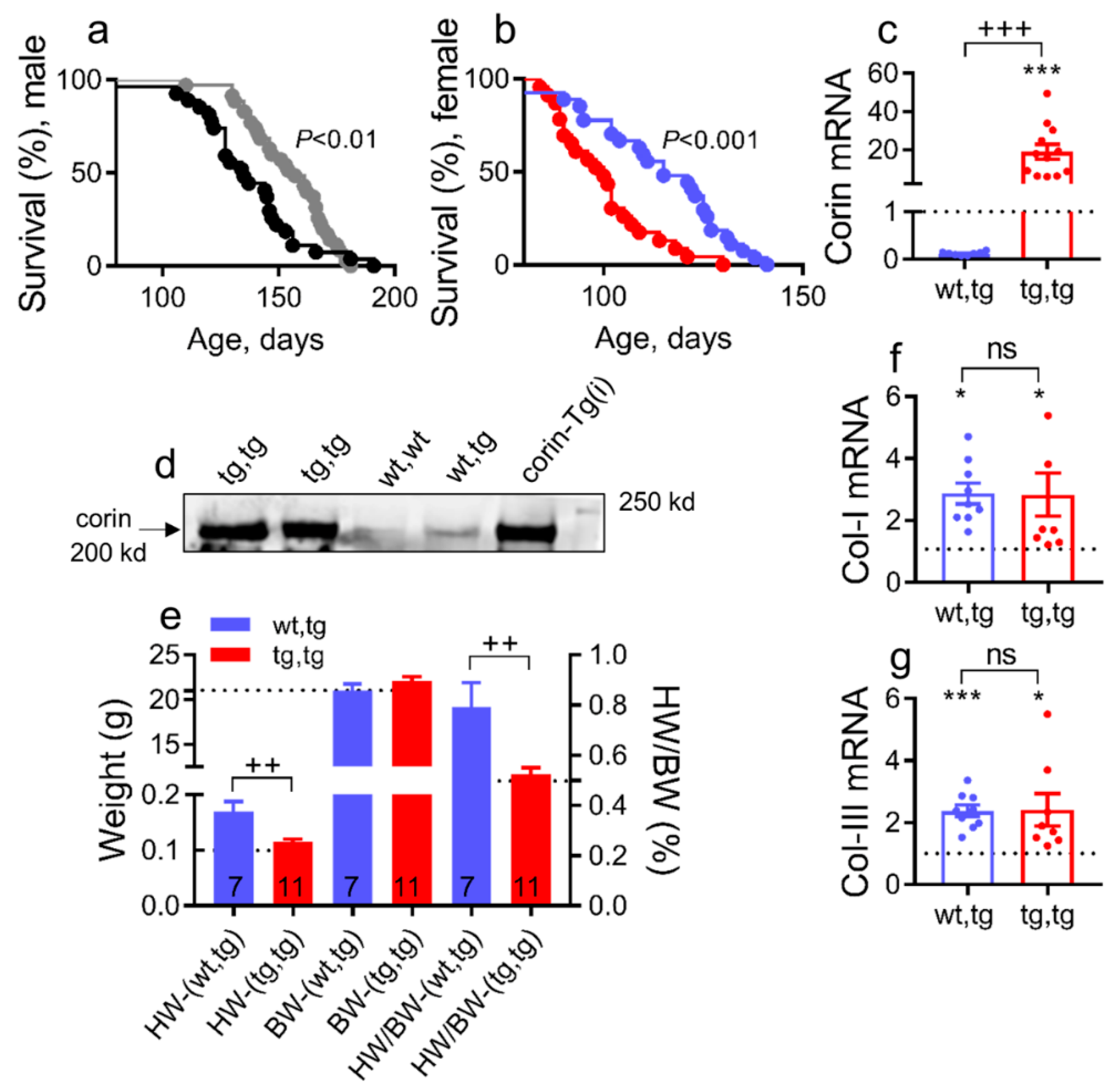
2.2. Catalytically Inactive Cardiac Corin-Tg(i) Improves Survival in Mice with DCM
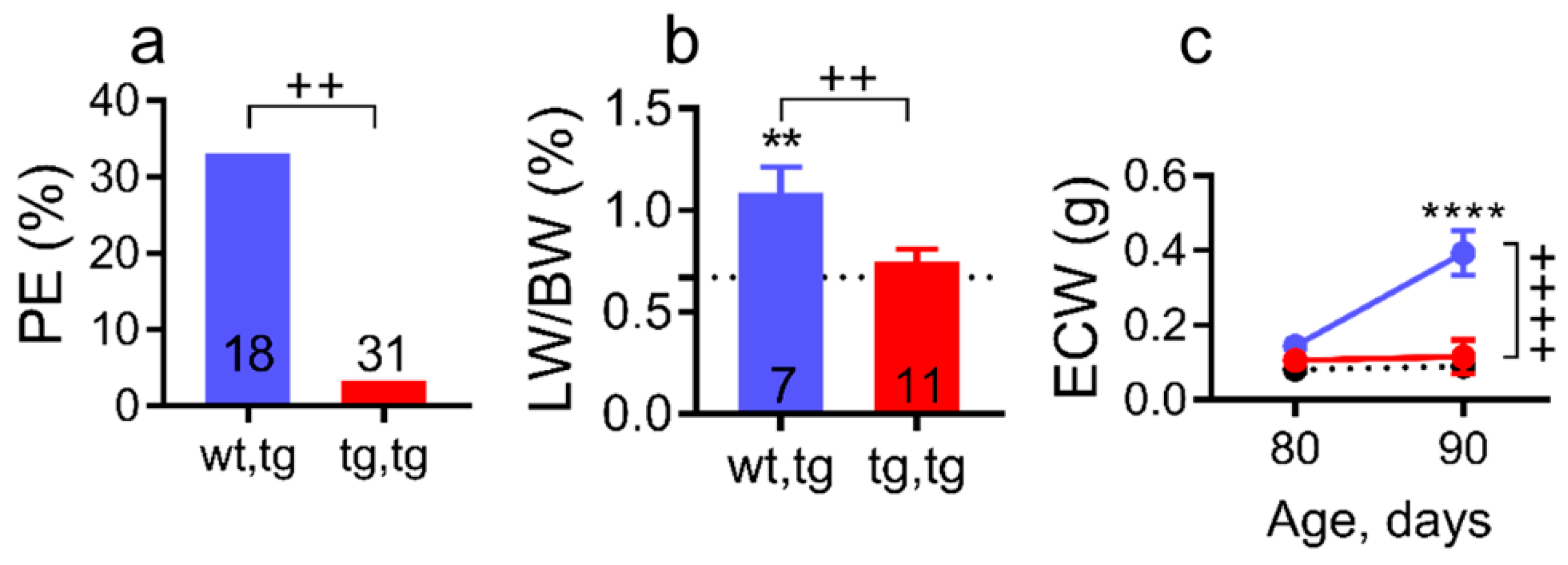
2.3. Corin-Tg(i) Overexpression Reduces Pleural Effusion, Lung Edema, and Systemic Water Retention in Mice with DCM
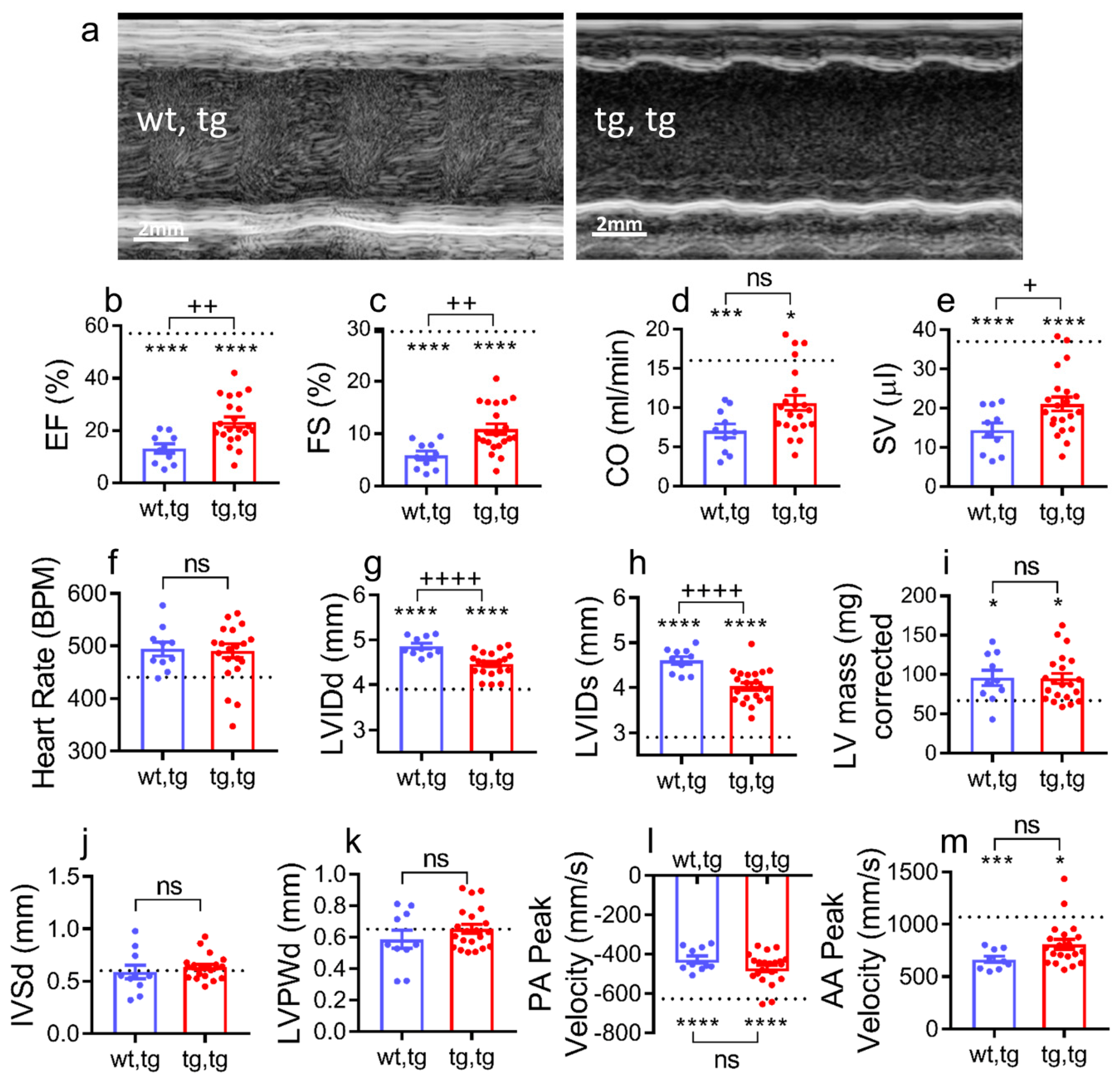
2.4. Corin-Tg(i) Overexpression Reduces Heart Systolic Dysfunction and Ventricular Dilation in DCM
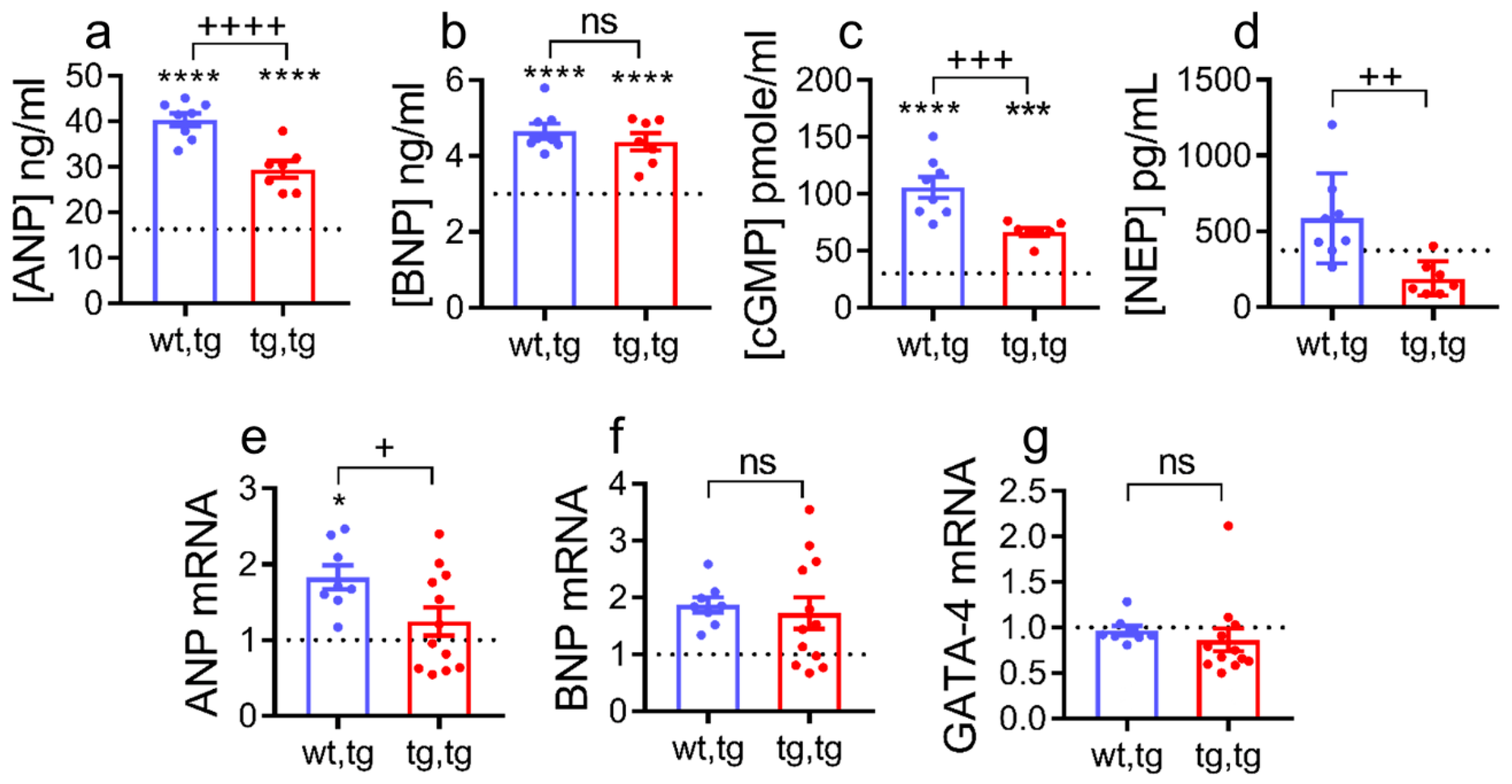
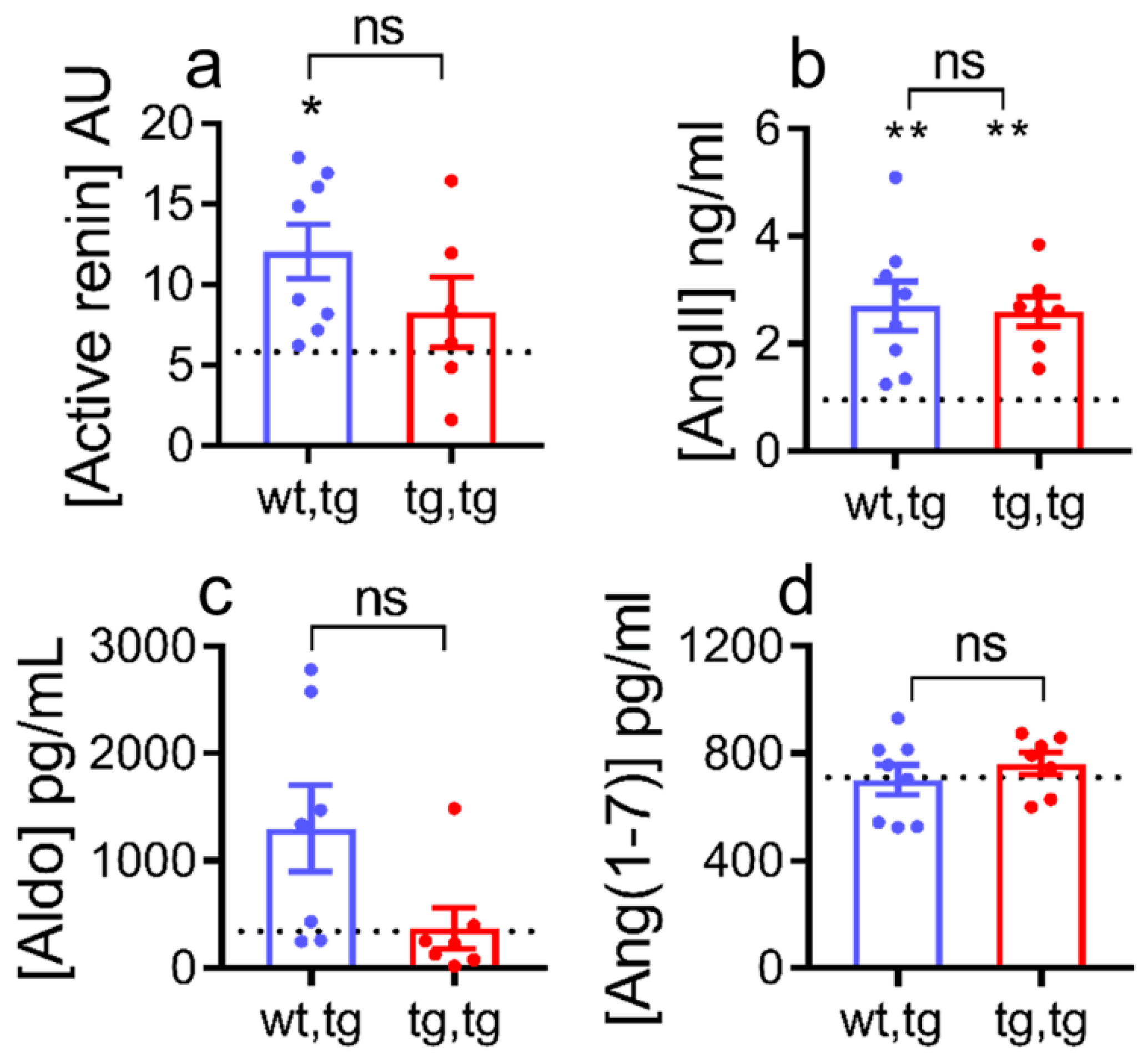
2.5. Effect of Cardiac Corin-Tg(i) Overexpression on HF Plasma Biomarkers
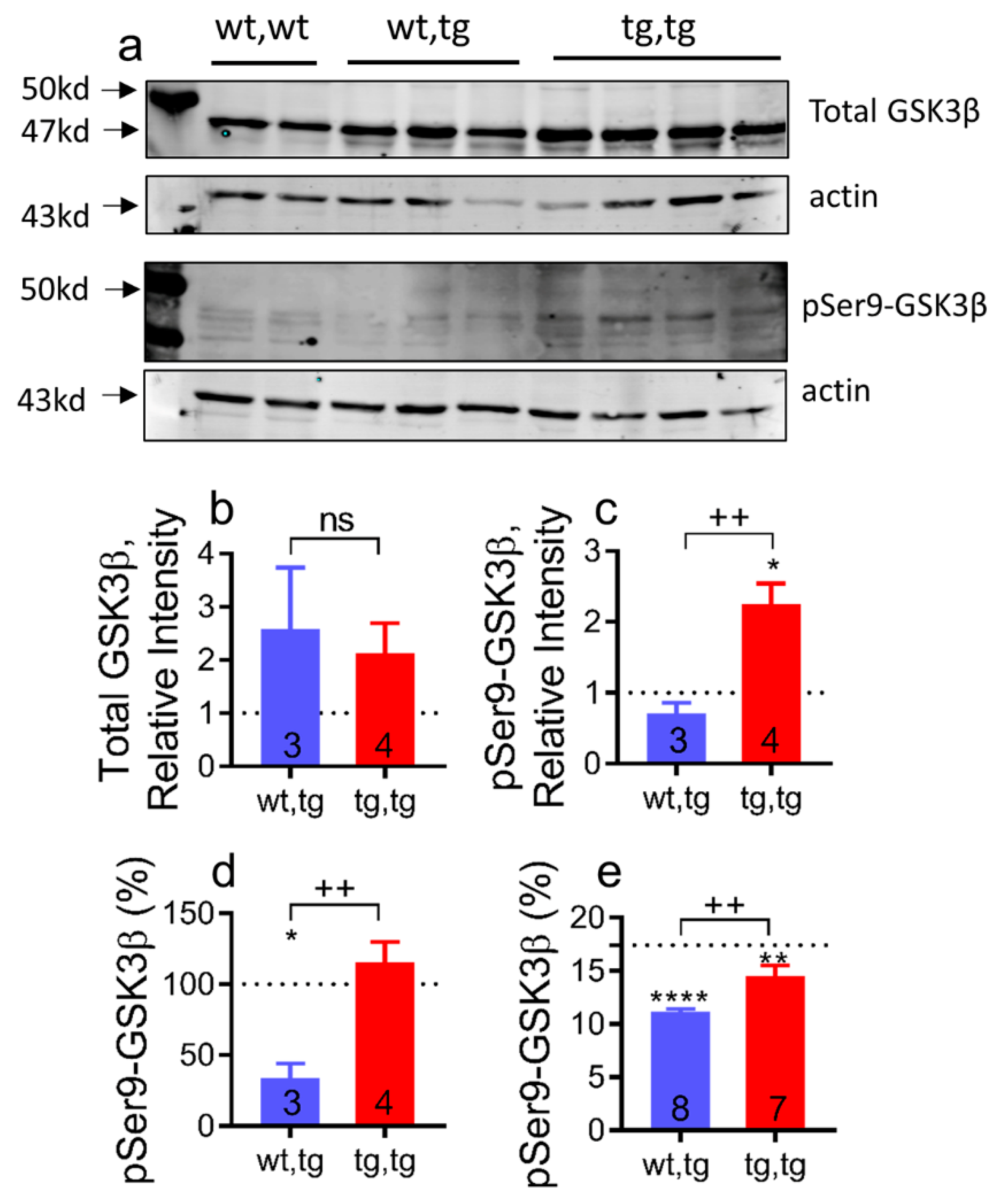
2.6. Cardiac Corin-Tg(i) Overexpression Increases pSer9-GSK3β Levels in DCM
3. Discussion
4. Materials and Methods
4.1. Institution and Environment
4.2. Mice
4.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.4. Echocardiography
4.5. Lung Edema and Pleural Effusion Analysis
4.6. Body Composition
4.7. Mouse Heart Tissue Extraction
4.8. Western Blot Analysis
4.9. Enzyme Immunoassay
4.10. Plasma Renin Activity Assay
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| wt,tg | corin-WT/DCM, |
| tg,tg | corin-Tg(i)/DCM |
| wt,wt | WT/WT |
| Corin-Tg (i) | corinS1052A-Tg |
| Tg | transgenic |
| WT | wild-type |
| HF | heart failure |
| rEF | reduced ejection fraction |
| HFrEF | heart failure with reduced ejection fraction |
| DCM | dilated cardiomyopathy |
| RAAS | renin-angiotensin-aldosterone-system |
| NP | natriuretic peptide |
| ANP | atrial natriuretic peptide |
| BNP | b-type natriuretic peptide |
| Ang II | angiotensin II |
| Aldo | aldosterone |
| Ang (1–7) | angiotensin (1–7) |
| NEP | neprilysin |
| cGMP | cyclic guanosine monophosphate |
| GSK3β | glycogen synthase kinase 3β |
| pSer9-GSK3β | glycogen synthase kinase 3β phosphorylated at Ser9 |
| ELISA | enzyme-linked immunosorbent assay |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| AU | arbitrary units |
| SEM | standard error of the mean |
| CD | cytoplasmic domain |
| TM | transmembrane |
| LDLR | low density lipoprotein receptor |
| Fz | frizzled |
| SR | macrophage scavenger receptor |
| H | histidine |
| D | aspartate |
| S | serine |
| A | alanine |
| BW | body weight |
| HW/BW | heart weight to bodyweight ratio |
| LW/BW | lung weight to bodyweight ratio |
| QMR | quantitative magnetic resonance |
| ECW | extracellular water |
| LV | left ventricle |
| EF | ejection fraction |
| FS | fraction shortening |
| CO | cardiac output |
| SV | stroke volume |
| BPM | beats per minute |
| LVIDd | left ventricular internal diameter, diastole |
| LVIDs | left ventricular internal diameter, systole |
| LVMc | left ventricular mass corrected |
| IVSd | interventricular septum, diastole |
| LVPWd | left ventricular posterior wall, diastole |
| PA | pulmonary artery |
| AA | ascending aorta |
| PE | pleural effusion |
References
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. Heart Disease and Stroke Statistics-2019 Update: A Report from the American Heart Association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef] [PubMed]
- Reed, B.N.; Sueta, C.A. Stage B: What is the evidence for treatment of asymptomatic left ventricular dysfunction? Curr. Cardiol. Rev. 2015, 11, 18–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Japp, A.G.; Gulati, A.; Cook, S.A.; Cowie, M.R.; Prasad, S.K. The Diagnosis and Evaluation of Dilated Cardiomyopathy. J. Am. Coll. Cardiol. 2016, 67, 2996–3010. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, R.G.; Semsarian, C.; Macdonald, P. Dilated cardiomyopathy. Lancet 2017, 390, 400–414. [Google Scholar] [CrossRef]
- Hershberger, R.E.; Hedges, D.J.; Morales, A. Dilated cardiomyopathy: The complexity of a diverse genetic architecture. Nat. Rev. Cardiol. 2013, 10, 531–547. [Google Scholar] [CrossRef] [PubMed]
- Yancy, C.W.; Jessup, M.; Bozkurt, B.; Butler, J.; Casey, D.E., Jr.; Drazner, M.H.; Fonarow, G.C.; Geraci, S.A.; Horwich, T.; Januzzi, J.L.; et al. 2013 ACCF/AHA guideline for the management of heart failure: Executive summary: A report of the American College of Cardiology Foundation/American Heart Association Task Force on practice guidelines. Circulation 2013, 128, 1810–1852. [Google Scholar] [CrossRef]
- Bozkurt, B.; Colvin, M.; Cook, J.; Cooper, L.T.; Deswal, A.; Fonarow, G.C.; Francis, G.S.; Lenihan, D.; Lewis, E.F.; McNamara, D.M.; et al. Current Diagnostic and Treatment Strategies for Specific Dilated Cardiomyopathies: A Scientific Statement from the American Heart Association. Circulation 2016, 134, e579–e646. [Google Scholar] [CrossRef]
- Writing Group, M.; Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Das, S.R.; de Ferranti, S.; Despres, J.P.; et al. Heart Disease and Stroke Statistics-2016 Update: A Report from the American Heart Association. Circulation 2016, 133, e38–e360. [Google Scholar]
- Langenickel, T.H.; Pagel, I.; Buttgereit, J.; Tenner, K.; Lindner, M.; Dietz, R.; Willenbrock, R.; Bader, M. Rat corin gene: Molecular cloning and reduced expression in experimental heart failure. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1516–H1521. [Google Scholar] [CrossRef] [Green Version]
- Barth, A.S.; Kuner, R.; Buness, A.; Ruschhaupt, M.; Merk, S.; Zwermann, L.; Kaab, S.; Kreuzer, E.; Steinbeck, G.; Mansmann, U.; et al. Identification of a common gene expression signature in dilated cardiomyopathy across independent microarray studies. J. Am. Coll. Cardiol. 2006, 48, 1610–1617. [Google Scholar] [CrossRef] [Green Version]
- Ibebuogu, U.N.; Gladysheva, I.P.; Reed, G.L. Is heart failure due to impaired cleavage and activation of atrial natriuretic peptide? J. Am. Coll. Cardiol. 2009, 53, A468–A469. [Google Scholar]
- Rame, J.E.; Tam, S.W.; McNamara, D.; Worcel, M.; Sabolinski, M.L.; Wu, A.H.; Dries, D.L. Dysfunctional corin i555(p568) allele is associated with impaired brain natriuretic peptide processing and adverse outcomes in blacks with systolic heart failure: Results from the Genetic Risk Assessment in Heart Failure substudy. Circ. Heart Fail. 2009, 2, 541–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, N.; Chen, S.; Yang, J.; He, L.; Liu, P.; Zheng, D.; Li, L.; Zhou, Y.; Ruan, C.; Plow, E.; et al. Plasma soluble corin in patients with heart failure. Circ. Heart Fail. 2010, 3, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibebuogu, U.N.; Gladysheva, I.P.; Houng, A.K.; Reed, G.L. Decompensated heart failure is associated with reduced corin levels and decreased cleavage of pro-atrial natriuretic peptide. Circ. Heart Fail. 2011, 4, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Dries, D.L. Process matters: Emerging concepts underlying impaired natriuretic peptide system function in heart failure. Circ. Heart Fail. 2011, 4, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Gladysheva, I.P.; Wang, D.; McNamee, R.A.; Houng, A.K.; Mohamad, A.A.; Fan, T.M.; Reed, G.L. Corin overexpression improves cardiac function, heart failure, and survival in mice with dilated cardiomyopathy. Hypertension 2013, 61, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Ngo, D.T.; Horowitz, J.D.; Sverdlov, A.L. Heart failure: A corin-deficient state? Hypertension 2013, 61, 284–285. [Google Scholar] [CrossRef] [Green Version]
- Ichiki, T.; Boerrigter, G.; Huntley, B.K.; Sangaralingham, S.J.; McKie, P.M.; Harty, G.J.; Harders, G.E.; Burnett, J.C., Jr. Differential expression of the pro-natriuretic peptide convertases corin and furin in experimental heart failure and atrial fibrosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R102–R109. [Google Scholar] [CrossRef] [Green Version]
- Ward, R.D.; Zaidi, S.S.; Ramanathan, K.; Yu, X.; Gladysheva, I.P.; Reed, G.L. Corin Levels Are Linked to Systolic Function and Serum Sodium. J. Card. Fail. 2013, 19, S12. [Google Scholar] [CrossRef]
- Barnet, C.S.; Liu, X.; Body, S.C.; Collard, C.D.; Shernan, S.K.; Muehlschlegel, J.D.; Jarolim, P.; Fox, A.A. Plasma corin decreases after coronary artery bypass graft surgery and is associated with postoperative heart failure: A pilot study. J. Cardiothorac. Vasc. Anesth. 2015, 29, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Chen, J.C.; Liu, Y.; Yang, H.; Du, K.; Kong, Y.; Xu, X.H. Plasma Corin as a Predictor of Cardiovascular Events in Patients with Chronic Heart Failure. JACC Heart Fail. 2016, 4, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.; Wang, D.; Sullivan, R.; Fan, T.H.; Gladysheva, I.P.; Reed, G.L. Depressed Corin Levels Indicate Early Systolic Dysfunction Before Increases of Atrial Natriuretic Peptide/B-Type Natriuretic Peptide and Heart Failure Development. Hypertension 2016, 67, 362–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.S.; Ward, R.D.; Ramanathan, K.; Yu, X.; Gladysheva, I.P.; Reed, G.L. Possible Enzymatic Downregulation of the Natriuretic Peptide System in Patients with Reduced Systolic Function and Heart Failure: A Pilot Study. Biomed Res. Int. 2018, 2018, 7279036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstreken, S.; Delrue, L.; Goethals, M.; Bartunek, J.; Vanderheyden, M. Natriuretic Peptide Processing in Patients with and Without Left Ventricular Dysfunction. Int. Heart J. 2019, 60, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, J.D.; Scarman, A.L.; Clarke, B.E.; Normyle, J.F.; Antalis, T.M. Localization of the mosaic transmembrane serine protease corin to heart myocytes. Eur. J. Biochem. 2000, 267, 6931–6937. [Google Scholar] [CrossRef]
- Gladysheva, I.P.; Robinson, B.R.; Houng, A.K.; Kovats, T.; King, S.M. Corin is co-expressed with pro-ANP and localized on the cardiomyocyte surface in both zymogen and catalytically active forms. J. Mol. Cell. Cardiol. 2008, 44, 131–142. [Google Scholar] [CrossRef]
- Armaly, Z.; Assady, S.; Abassi, Z. Corin: A new player in the regulation of salt-water balance and blood pressure. Curr. Opin. Nephrol. Hypertens. 2013, 22, 713–722. [Google Scholar] [CrossRef]
- Burnett, J.C., Jr.; Granger, J.P.; Opgenorth, T.J. Effects of synthetic atrial natriuretic factor on renal function and renin release. Am. J. Physiol. 1984, 247, F863–F866. [Google Scholar] [CrossRef] [Green Version]
- Johnston, C.I.; Hodsman, P.G.; Kohzuki, M.; Casley, D.J.; Fabris, B.; Phillips, P.A. Interaction between atrial natriuretic peptide and the renin angiotensin aldosterone system. Endogenous antagonists. Am. J. Med. 1989, 87, 24S–28S. [Google Scholar]
- Yamamoto, K.; Burnett, J.C., Jr.; Redfield, M.M. Effect of endogenous natriuretic peptide system on ventricular and coronary function in failing heart. Am. J. Physiol. 1997, 273, H2406–H2414. [Google Scholar] [CrossRef]
- Tomita, Y.; Kim, D.H.; Magoori, K.; Fujino, T.; Yamamoto, T.T. A novel low-density lipoprotein receptor-related protein with type II membrane protein-like structure is abundant in heart. J. Biochem. 1998, 124, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Hinzmann, B.; Yan, W.; Wu, F.; Morser, J.; Wu, Q. Genomic structures of the human and murine corin genes and functional GATA elements in their promoters. J. Biol. Chem. 2002, 277, 38390–38398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Wu, S.; Wang, W.; Chen, S.; Peng, J.; Zhang, X.; Wu, Q. Ectodomain shedding and autocleavage of the cardiac membrane protease corin. J. Biol. Chem. 2011, 286, 10066–10072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fentzke, R.C.; Korcarz, C.E.; Lang, R.M.; Lin, H.; Leiden, J.M. Dilated cardiomyopathy in transgenic mice expressing a dominant-negative CREB transcription factor in the heart. J. Clin. Investig. 1998, 101, 2415–2426. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.D.; Mehta, R.M.; Tripathi, R.; Gladysheva, I.P.; Reed, G.L. Normalizing Plasma Renin Activity in Experimental Dilated Cardiomyopathy: Effects on Edema, Cachexia, and Survival. Int. J. Mol. Sci. 2019, 20, 3886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, R.; Sullivan, R.; Fan, T.M.; Wang, D.; Sun, Y.; Reed, G.L.; Gladysheva, I.P. Enhanced heart failure, mortality and renin activation in female mice with experimental dilated cardiomyopathy. PLoS ONE 2017, 12, e0189315. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Gladysheva, I.P.; Fan, T.H.; Sullivan, R.; Houng, A.K.; Reed, G.L. Atrial natriuretic peptide affects cardiac remodeling, function, heart failure, and survival in a mouse model of dilated cardiomyopathy. Hypertension 2014, 63, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Watson, P.A.; Birdsey, N.; Huggins, G.S.; Svensson, E.; Heppe, D.; Knaub, L. Cardiac-specific overexpression of dominant-negative CREB leads to increased mortality and mitochondrial dysfunction in female mice. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H2056–H2068. [Google Scholar] [CrossRef] [Green Version]
- Gladysheva, I.P.; King, S.M.; Houng, A.K. N-glycosylation modulates the cell-surface expression and catalytic activity of corin. Biochem. Biophys. Res. Commun. 2008, 373, 130–135. [Google Scholar] [CrossRef]
- Ware, L.B.; Matthay, M.A. Clinical practice. Acute pulmonary edema. N. Engl. J. Med. 2005, 353, 2788–2796. [Google Scholar] [CrossRef] [Green Version]
- Chow, S.L.; Maisel, A.S.; Anand, I.; Bozkurt, B.; de Boer, R.A.; Felker, G.M.; Fonarow, G.C.; Greenberg, B.; Januzzi, J.L., Jr.; Kiernan, M.S.; et al. Role of Biomarkers for the Prevention, Assessment, and Management of Heart Failure: A Scientific Statement from the American Heart Association. Circulation 2017, 135, e1054–e1091. [Google Scholar] [CrossRef] [PubMed]
- Dzau, V.J.; Colucci, W.S.; Hollenberg, N.K.; Williams, G.H. Relation of the renin-angiotensin-aldosterone system to clinical state in congestive heart failure. Circulation 1981, 63, 645–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrier, R.W.; Abraham, W.T. Hormones and hemodynamics in heart failure. N. Engl. J. Med. 1999, 341, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.T. Aldosterone in congestive heart failure. N. Engl. J. Med. 2001, 345, 1689–1697. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, Y.; Yoshimura, M.; Yasue, H.; Sakamoto, T.; Ogawa, H.; Kugiyama, K.; Harada, E.; Nakayama, M.; Nakamura, S.; Ito, T.; et al. Aldosterone production is activated in failing ventricle in humans. Circulation 2001, 103, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Vasan, R.S.; Evans, J.C.; Benjamin, E.J.; Levy, D.; Larson, M.G.; Sundstrom, J.; Murabito, J.M.; Sam, F.; Colucci, W.S.; Wilson, P.W. Relations of serum aldosterone to cardiac structure: Gender-related differences in the Framingham Heart Study. Hypertension 2004, 43, 957–962. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, R.D.; Mehta, R.M.; Tripathi, R.; Reed, G.L.; Gladysheva, I.P. Renin Activity in Heart Failure with Reduced Systolic Function-New Insights. Int. J. Mol. Sci. 2019, 20, 3182. [Google Scholar] [CrossRef] [Green Version]
- Miura, T.; Miki, T. GSK-3beta, a therapeutic target for cardiomyocyte protection. Circ. J. 2009, 73, 1184–1192. [Google Scholar] [CrossRef] [Green Version]
- Takahashi-Yanaga, F. Activator or inhibitor? GSK-3 as a new drug target. Biochem. Pharmacol. 2013, 86, 191–199. [Google Scholar] [CrossRef]
- Dawson, K.; Aflaki, M.; Nattel, S. Role of the Wnt-Frizzled system in cardiac pathophysiology: A rapidly developing, poorly understood area with enormous potential. J. Physiol. 2013, 591, 1409–1432. [Google Scholar] [CrossRef]
- Buckley, C.L.; Stokes, A.J. Corin-deficient W-sh mice poorly tolerate increased cardiac afterload. Regul. Pept. 2011, 172, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaraj, S.; Claggett, B.; Pozzi, A.; McMurray, J.J.V.; Jhund, P.S.; Packer, M.; Desai, A.S.; Lewis, E.F.; Vaduganathan, M.; Lefkowitz, M.P.; et al. Prognostic Implications of Congestion on Physical Examination Among Contemporary Patients with Heart Failure and Reduced Ejection Fraction:PARADIGM-HF. Circulation 2019, 140, 1369–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, M.; Yasue, H.; Ogawa, H. Pathophysiological significance and clinical application of ANP and BNP in patients with heart failure. Can. J. Physiol. Pharmacol. 2001, 79, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Dewan, P.; Docherty, K.F.; McMurray, J.J.V. Sacubitril/Valsartan in Asian Patients with Heart Failure with Reduced Ejection Fraction. Korean Circ. J. 2019, 49, 469–484. [Google Scholar] [CrossRef] [Green Version]
- Mentz, R.J.; Bakris, G.L.; Waeber, B.; McMurray, J.J.; Gheorghiade, M.; Ruilope, L.M.; Maggioni, A.P.; Swedberg, K.; Pina, I.L.; Fiuzat, M.; et al. The past, present and future of renin-angiotensin aldosterone system inhibition. Int. J. Cardiol. 2013, 167, 1677–1687. [Google Scholar] [CrossRef] [Green Version]
- von Lueder, T.G.; Sangaralingham, S.J.; Wang, B.H.; Kompa, A.R.; Atar, D.; Burnett, J.C., Jr.; Krum, H. Renin-angiotensin blockade combined with natriuretic peptide system augmentation: Novel therapeutic concepts to combat heart failure. Circ. Heart Fail. 2013, 6, 594–605. [Google Scholar] [CrossRef] [Green Version]
- Houser, S.R.; Margulies, K.B.; Murphy, A.M.; Spinale, F.G.; Francis, G.S.; Prabhu, S.D.; Rockman, H.A.; Kass, D.A.; Molkentin, J.D.; Sussman, M.A.; et al. Animal models of heart failure: A scientific statement from the American Heart Association. Circ. Res. 2012, 111, 131–150. [Google Scholar] [CrossRef] [Green Version]
- von Haehling, S.; Ebner, N.; Dos Santos, M.R.; Springer, J.; Anker, S.D. Muscle wasting and cachexia in heart failure: Mechanisms and therapies. Nat. Rev. Cardiol. 2017, 14, 323–341. [Google Scholar] [CrossRef]
- Shen, C.; Zhou, J.; Wang, X.; Yu, X.Y.; Liang, C.; Liu, B.; Pan, X.; Zhao, Q.; Song, J.L.; Wang, J.; et al. Angiotensin-II-induced Muscle Wasting is Mediated by 25-Hydroxycholesterol via GSK3beta Signaling Pathway. EBioMedicine 2017, 16, 238–250. [Google Scholar] [CrossRef]
- Foulquier, S.; Daskalopoulos, E.P.; Lluri, G.; Hermans, K.C.M.; Deb, A.; Blankesteijn, W.M. WNT Signaling in Cardiac and Vascular Disease. Pharmacol. Rev. 2018, 70, 68–141. [Google Scholar] [CrossRef]
- Iyer, L.M.; Nagarajan, S.; Woelfer, M.; Schoger, E.; Khadjeh, S.; Zafiriou, M.P.; Kari, V.; Herting, J.; Pang, S.T.; Weber, T.; et al. A context-specific cardiac beta-catenin and GATA4 interaction influences TCF7L2 occupancy and remodels chromatin driving disease progression in the adult heart. Nucleic Acids Res. 2018, 46, 2850–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janda, C.Y.; Waghray, D.; Levin, A.M.; Thomas, C.; Garcia, K.C. Structural basis of Wnt recognition by Frizzled. Science 2012, 337, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huggins, G.S.; Lepore, J.J.; Greytak, S.; Patten, R.; McNamee, R.; Aronovitz, M.; Wang, P.J.; Reed, G.L. The CREB leucine zipper regulates CREB phosphorylation, cardiomyopathy, and lethality in a transgenic model of heart failure. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1877–H1882. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripathi, R.; Sullivan, R.D.; Fan, T.-H.M.; Houng, A.K.; Mehta, R.M.; Reed, G.L.; Gladysheva, I.P. Cardiac-Specific Overexpression of Catalytically Inactive Corin Reduces Edema, Contractile Dysfunction, and Death in Mice with Dilated Cardiomyopathy. Int. J. Mol. Sci. 2020, 21, 203. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010203
Tripathi R, Sullivan RD, Fan T-HM, Houng AK, Mehta RM, Reed GL, Gladysheva IP. Cardiac-Specific Overexpression of Catalytically Inactive Corin Reduces Edema, Contractile Dysfunction, and Death in Mice with Dilated Cardiomyopathy. International Journal of Molecular Sciences. 2020; 21(1):203. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010203
Chicago/Turabian StyleTripathi, Ranjana, Ryan D. Sullivan, Tai-Hwang M. Fan, Aiilyan K. Houng, Radhika M. Mehta, Guy L. Reed, and Inna P. Gladysheva. 2020. "Cardiac-Specific Overexpression of Catalytically Inactive Corin Reduces Edema, Contractile Dysfunction, and Death in Mice with Dilated Cardiomyopathy" International Journal of Molecular Sciences 21, no. 1: 203. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010203