Potential Use of Biotherapeutic Bacteria to Target Colorectal Cancer-Associated Taxa

Abstract
:1. Introduction
2. The Gut Microbiota of CRC Patients
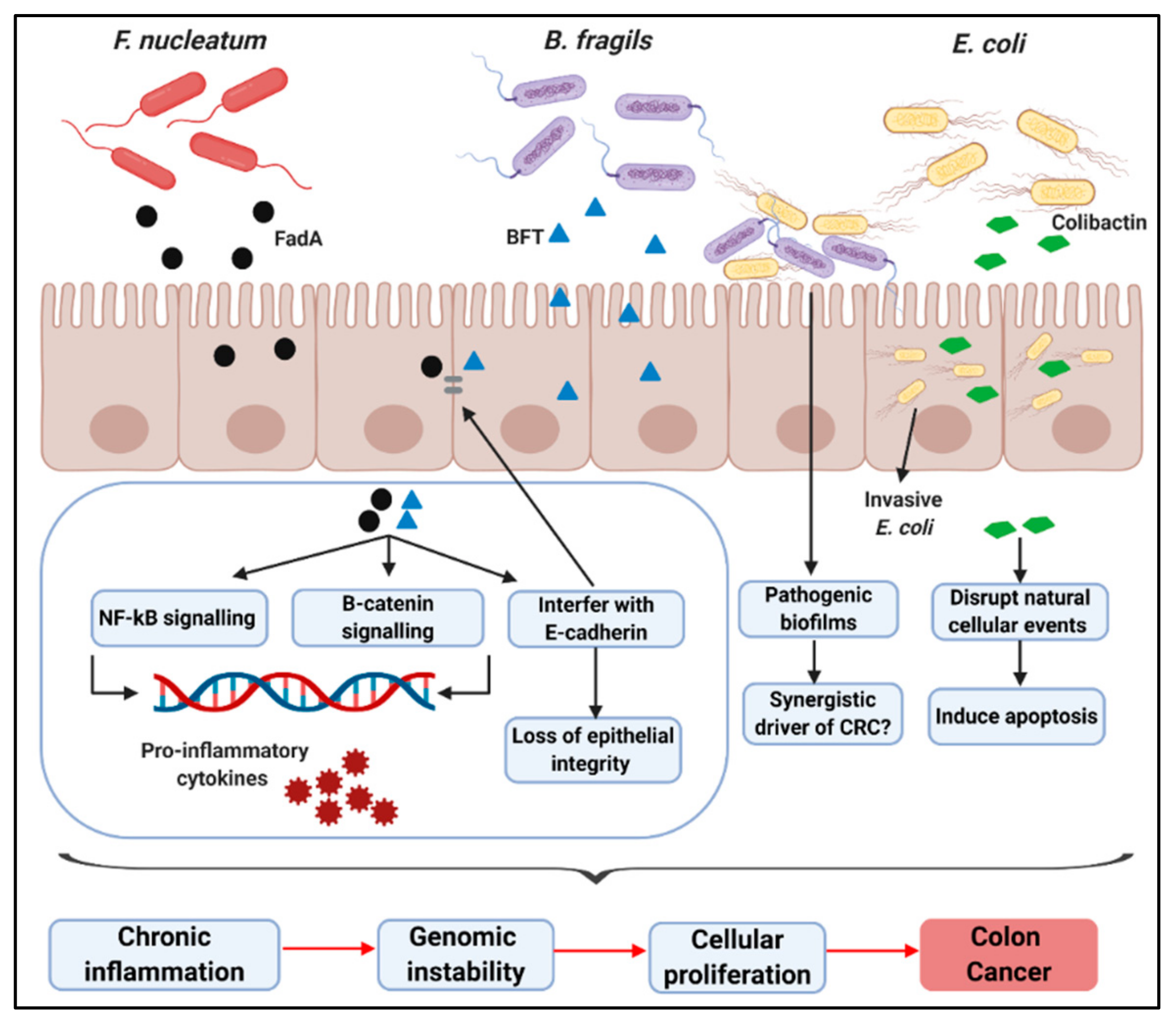
3. Specific Taxa Associated with CRC Development
3.1. F. nucleatum
3.2. B. fragilis
3.3. E. coli
3.4. Other Bacteria
4. Potential Use of Biotherapeutic Bacteria to Target Colorectal Cancer-Associated Taxa
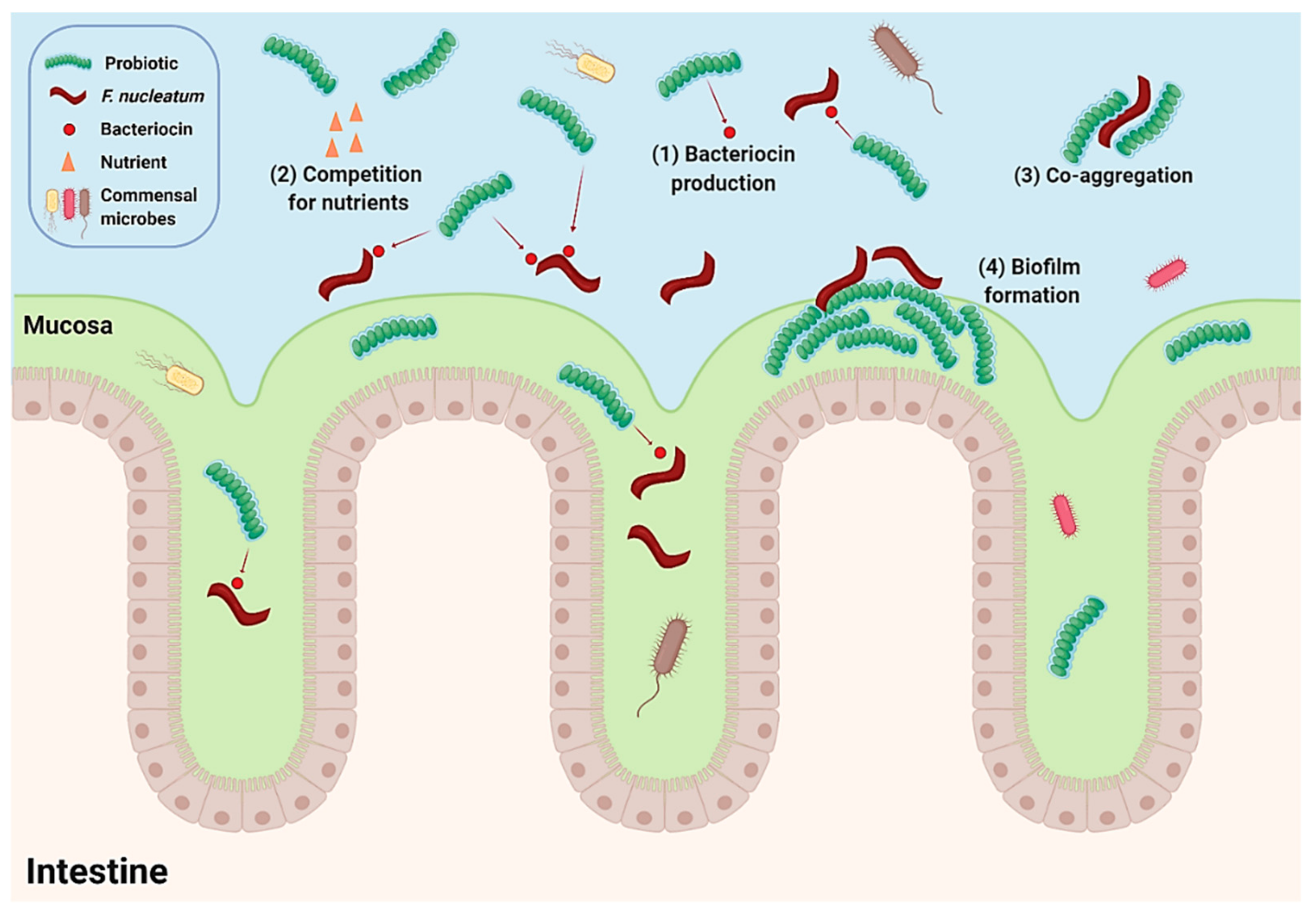
5. Antimicrobial Activity of Biotherapeutic Strains against Fusobacterium nucleatum
6. Probiotic Intervention Modulates the CRC-Associated Microbiome
7. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- What Is Colorectal Cancer? Available online: http://www.cancer.org/cancer/colon-rectal-cancer/about/what-is-colorectal-cancer.html (accessed on 20 January 2020).
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grady, W.M.; Carethers, J.M. Genomic and Epigenetic Instability in Colorectal Cancer Pathogenesis. Gastroenterology 2008, 135, 1079–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colussi, D.; Brandi, G.; Bazzoli, F.; Ricciardiello, L. Molecular Pathways Involved in Colorectal Cancer: Implications for Disease Behavior and Prevention. Int. J. Mol. Sci. 2013, 14, 16365–16385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.N. Genetic and Epigenetic Alterations of Colorectal Cancer. Intest. Res. 2018, 16, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Larsson, S.C.; Wolk, A. Meat Consumption and Risk of Colorectal Cancer: A Meta-Analysis of Prospective Studies. Int. J. Cancer 2006, 119, 2657–2664. [Google Scholar] [CrossRef] [Green Version]
- Riboli, E.; Norat, T. Epidemiologic Evidence of the Protective Effect of Fruit and Vegetables on Cancer Risk. Am. J. Clin. Nutr. 2003, 78, 559–569. [Google Scholar] [CrossRef] [Green Version]
- Burnett-Hartman, A.N.; Newcomb, P.A.; Potter, J.D. Infectious Agents and Colorectal Cancer: A Review of Helicobacter Pylori, Streptococcus Bovis, JC Virus, and Human Papillomavirus. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 2970–2979. [Google Scholar] [CrossRef] [Green Version]
- Antonic, V.; Stojadinovic, A.; Kester, K.E.; Weina, P.J.; Brücher, B.L.; Protic, M.; Avital, I.; Izadjoo, M. Significance of Infectious Agents in Colorectal Cancer Development. J. Cancer 2013, 4, 227–240. [Google Scholar] [CrossRef]
- Zou, S.; Fang, L.; Lee, M.-H. Dysbiosis of Gut Microbiota in Promoting the Development of Colorectal Cancer. Gastroenterol. Rep. 2018, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Garrett, W.S. Cancer and the Microbiota. Science 2015, 348, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H. Chemotherapy for Colorectal Cancer in the Elderly. World J. Gastroenterol. 2015, 21, 5158–5166. [Google Scholar] [CrossRef]
- Assed Bastos, D.; Freitas, D.; Coelho Ribeiro, S.; Hoff, P.M. Review: Combination Therapy in High-Risk Stage II or Stage III Colon Cancer: Current Practice and Future Prospects. Ther. Adv. Med. Oncol. 2010, 2, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocarnik, J.M.; Shiovitz, S.; Phipps, A.I. Molecular Phenotypes of Colorectal Cancer and Potential Clinical Applications. Gastroenterol. Rep. 2015, 3, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, D.E. Targeted Therapies: A New Generation of Cancer Treatments. Am. Fam. Physician 2008, 77, 311–319. [Google Scholar] [PubMed]
- Stavrou, G.; Kotzampassi, K. Gut Microbiome, Surgical Complications and Probiotics. Ann. Gastroenterol. 2017, 30, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.-A.; Pezet, D.; Bonnet, M. Gut Microbiota Imbalance and Colorectal Cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Kato, I. Gut Microbiota, Inflammation and Colorectal Cancer. Genes Dis. 2016, 3, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Jahani-Sherafat, S.; Alebouyeh, M.; Moghim, S.; Ahmadi Amoli, H.; Ghasemian-Safaei, H. Role of Gut Microbiota in the Pathogenesis of Colorectal Cancer; A Review Article. Gastroenterol. Hepatol. Bed Bench 2018, 11, 101–109. [Google Scholar]
- Wang, T.; Cai, G.; Qiu, Y.; Fei, N.; Zhang, M.; Pang, X.; Jia, W.; Cai, S.; Zhao, L. Structural Segregation of Gut Microbiota between Colorectal Cancer Patients and Healthy Volunteers. ISME J. 2012, 6, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Kong, C.; Huang, L.; Li, H.; Qu, X.; Liu, Z.; Lan, P.; Wang, J.; Qin, H. Mucosa-Associated Microbiota Signature in Colorectal Cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2073–2083. [Google Scholar] [CrossRef]
- Hibberd, A.A.; Lyra, A.; Ouwehand, A.C.; Rolny, P.; Lindegren, H.; Cedgård, L.; Wettergren, Y. Intestinal Microbiota Is Altered in Patients with Colon Cancer and Modified by Probiotic Intervention. BMJ Open Gastroenterol. 2017, 4, e000145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, R.; Ahn, J.; Sampson, J.N.; Shi, J.; Yu, G.; Xiong, X.; Hayes, R.B.; Goedert, J.J. Fecal Microbiota, Fecal Metabolome, and Colorectal Cancer Interrelations. PLoS ONE 2016, 11, e0152126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of Human Gut Microbiota in Control Subjects and Patients with Colorectal Carcinoma in Adenoma: Terminal Restriction Fragment Length Polymorphism and next-Generation Sequencing Analyses. Oncol. Rep. 2016, 35, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, R.L.; Freeman, D.J.; Pleasance, S.; Watson, P.; Moore, R.A.; Cochrane, K.; Allen-Vercoe, E.; Holt, R.A. Co-Occurrence of Anaerobic Bacteria in Colorectal Carcinomas. Microbiome 2013, 1, 16. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Qin, H. Microbiota Disbiosis Is Associated with Colorectal Cancer. Front. Microbiol. 2015, 6, 20. [Google Scholar] [CrossRef]
- Flemer, B.; Warren, R.D.; Barrett, M.P.; Cisek, K.; Das, A.; Jeffery, I.B.; Hurley, E.; O’Riordain, M.; Shanahan, F.; O’Toole, P.W. The Oral Microbiota in Colorectal Cancer Is Distinctive and Predictive. Gut 2018, 67, 1454–1463. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wu, Y.; He, L.; Wu, L.; Wang, X.; Liu, Z. Effects of the Intestinal Microbial Metabolite Butyrate on the Development of Colorectal Cancer. J. Cancer 2018, 9, 2510–2517. [Google Scholar] [CrossRef]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Pedeta, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef]
- Rezasoltani, S.; Aghdaei, H.A.; Dabiri, H.; Sepahi, A.A.; Modarressi, M.H.; Mojarad, N. The Association between Fecal Microbiota and Different Types of Colorectal Polyp as Precursors of Colorectal Cancer. Microb. Pathog. 2018, 124, 244–249. [Google Scholar] [CrossRef]
- Purcell, R.V.; Pearson, J.; Aitchison, A.; Dixon, L.; Frizelle, F.A.; Keenan, J.I. Colonization with Enterotoxigenic Bacteroides Fragilis Is Associated with Early-Stage Colorectal Neoplasia. PLoS ONE 2017, 12, e0171602. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, M.; Buc, E.; Sauvanet, P.; Darcha, C.; Dubois, D.; Pereira, B.; Dechelotte, P.; Bonnet, R.; Pezet, D.; Darfeuille-Michaud, A. Colonization of the Human Gut by E. Coli and Colorectal Cancer Risk. Clin. Cancer Res. 2014, 20, 859–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, T.N.Y.; Wang, X.; Nakatsu, G.; Chow, T.C.; Tipoe, T.; Dai, R.Z.W.; Tsoi, K.K.K.; Wong, M.C.S.; Tse, G.; Chan, M.T.V.; et al. Association Between Bacteremia From Specific Microbes and Subsequent Diagnosis of Colorectal Cancer. Gastroenterology 2018, 155, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Liu, Y.; Wang, J.; Guo, Y.; Zhang, Y.; Xiao, S. Detection of Fusobacterium Nucleatum and FadA Adhesin Gene in Patients with Orthodontic Gingivitis and Non-Orthodontic Periodontal Inflammation. PLoS ONE 2014, 9, e85280. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Eun, C.S.; Lee, A.R.; Park, C.H.; Han, D.S. Fusobacterium Isolates Recovered From Colonic Biopsies of Inflammatory Bowel Disease Patients in Korea. Ann. Lab. Med. 2016, 36, 387–389. [Google Scholar] [CrossRef] [Green Version]
- Swidsinski, A.; Dorffel, Y.; Loening-Baucke, V.; Theissig, F.; Ruckert, J.C.; Ismail, M.; Rau, W.A.; Gaschler, D.; Weizenegger, M.; Kuhn, S.; et al. Acute Appendicitis Is Characterised by Local Invasion with Fusobacterium Nucleatum/Necrophorum. Gut 2011, 60, 34–40. [Google Scholar] [CrossRef]
- Vander Haar, E.L.; So, J.; Gyamfi-Bannerman, C.; Han, Y.W. Fusobacterium Nucleatum and Adverse Pregnancy Outcomes: Epidemiological and Mechanistic Evidence. Anaerobe 2018, 50, 55–59. [Google Scholar] [CrossRef]
- Chhibber-Goel, J.; Singhal, V.; Bhowmik, D.; Vivek, R.; Parakh, N.; Bhargava, B.; Sharma, A. Linkages between Oral Commensal Bacteria and Atherosclerotic Plaques in Coronary Artery Disease Patients. NPJ Biofilms Microbiomes 2016, 2, 7. [Google Scholar] [CrossRef]
- Koziel, J.; Mydel, P.; Potempa, J. The Link Between Periodontal Disease and Rheumatoid Arthritis: An Updated Review. Curr. Rheumatol. Rep. 2014, 16, 408. [Google Scholar] [CrossRef] [Green Version]
- Sparks Stein, P.; Steffen, M.J.; Smith, C.; Jicha, G.; Ebersole, J.L.; Abner, E.; Dawson, D. Serum Antibodies to Periodontal Pathogens Are a Risk Factor for Alzheimer’s Disease. Alzheimer’s Dement. 2012, 8, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Guinane, C.M.; Tadrous, A.; Fouhy, F.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.; Andrews, E.; Cotter, P.D.; Stanton, C.; Ross, P.R. Microbial composition of human appendices from patients following appendectomy. mBio 2013, 4, e00366-12. [Google Scholar] [CrossRef] [Green Version]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium Nucleatum Infection Is Prevalent in Human Colorectal Carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human Intestinal Lumen and Mucosa-Associated Microbiota in Patients with Colorectal Cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, L.; Schmid, J.; Ebert, M.; Soucek, P.; Kunicka, T.; Liska, V.; Bruha, J.; Neary, P.; Dezeeuw, N.; Tommasino, M.; et al. Fusobacterium Nucleatum Associates with Stages of Colorectal Neoplasia Development, Colorectal Cancer and Disease Outcome. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium Nucleatum Potentiates Intestinal Tumorigenesis and Modulates the Tumor-Immune Microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Mima, K.; Sukawa, Y.; Nishihara, R.; Qian, Z.R.; Yamauchi, M.; Inamura, K.; Kim, S.A.; Masuda, A.; Nowak, J.A.; Nosho, K.; et al. Fusobacterium Nucleatum and T Cells in Colorectal Carcinoma. JAMA Oncol. 2015, 1, 653. [Google Scholar] [CrossRef] [Green Version]
- Tahara, T.; Yamamoto, E.; Suzuki, H.; Maruyama, R.; Chung, W.; Garriga, J.; Jelinek, J.; Yamano, H.-O.; Sugai, T.; An, B.; et al. Fusobacterium in Colonic Flora and Molecular Features of Colorectal Carcinoma. Cancer Res. 2014, 74, 1311–1318. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Han, D.S.; Oh, Y.-H.; Lee, A.-R.; Lee, Y.-R.; Eun, C.S. Role of Fusobacteria in the Serrated Pathway of Colorectal Carcinogenesis. Sci. Rep. 2016, 6, 25271. [Google Scholar] [CrossRef]
- Advani, S.M.; Advani, P.; DeSantis, S.M.; Brown, D.; VonVille, H.M.; Lam, M.; Loree, J.M.; Mehrvarz Sarshekeh, A.; Bressler, J.; Lopez, D.S.; et al. Clinical, Pathological, and Molecular Characteristics of CpG Island Methylator Phenotype in Colorectal Cancer: A Systematic Review and Meta-Analysis. Transl. Oncol. 2018, 11, 1188–1201. [Google Scholar] [CrossRef]
- Xu, M.; Yamada, M.; Li, M.; Liu, H.; Chen, S.G.; Han, Y.W. FadA from Fusobacterium Nucleatum Utilizes Both Secreted and Nonsecreted Forms for Functional Oligomerization for Attachment and Invasion of Host Cells. J. Biol. Chem. 2007, 282, 25000–25009. [Google Scholar] [CrossRef] [Green Version]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium Nucleatum Promotes Colorectal Carcinogenesis by Modulating E-Cadherin/β-Catenin Signaling via Its FadA Adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.-T.; Luo, H.-S.; Gao, F.; Tang, Q.-C.; Chen, W. Fusobacterium Nucleatum Promotes the Progression of Colorectal Cancer by Interacting with E-Cadherin. Oncol. Lett. 2018, 16, 2606–2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quah, S.Y.; Bergenholtz, G.; Tan, K.S. Fusobacterium Nucleatum Induces Cytokine Production through Toll-like-Receptor-Independent Mechanism. Int. Endod. J. 2014, 47, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S.; et al. Binding of the Fap2 Protein of Fusobacterium Nucleatum to Human Inhibitory Receptor TIGIT Protects Tumors from Immune Cell Attack. Immunity 2015, 42, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Li, Q.; Wu, J.; Wu, Y.; Peng, W.; Li, H.; Wang, J.; Tang, X.; Peng, Y.; Fu, X. Fusobacterium Nucleatum Promotes M2 Polarization of Macrophages in the Microenvironment of Colorectal Tumours via a TLR4-Dependent Mechanism. Cancer Immunol. Immunother. 2018, 67, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.C.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium Nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 70, 548–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Li, Z. TOX Gene: A Novel Target for Human Cancer Gene Therapy. Am. J. Cancer Res. 2015, 5, 3516–3524. [Google Scholar]
- Chen, T.; Li, Q.; Zhang, X.; Wu, Y.; Wu, J. Original Contribution TOX Expression Decreases with Progression of Colorectal Cancers and Is Associated with CD4 T-Cell Density and Fusobacterium Nucleatum Infection. Hum. Pathol. 2018, 79, 93–101. [Google Scholar] [CrossRef]
- Könönen, E.; Jousimies-Somer, H.; Asikainen, S. Relationship between Oral Gram-Negative Anaerobic Bacteria in Saliva of the Mother and the Colonization of Her Edentulous Infant. Oral Microbiol. Immunol. 1992, 7, 273–276. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, H.; Li, Z.; Tan, Y.; Han, Y.; Wang, X.; Du, Z.; Liu, Y.; Yang, R.; Bai, Y.; et al. Safety Evaluation of a Novel Strain of Bacteroides Fragilis. Front. Microbiol. 2017, 8, 435. [Google Scholar] [CrossRef] [Green Version]
- Culpepper, B.S.T.; Mai, V. Evidence for Contributions of Gut Microbiota to Colorectal Carcinogenesis. Curr. Nutr. Rep. 2013, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Kling, J.J.; Wright, R.L.; Moncrief, J.S.; Wilkins, T.D. Cloning and Characterization of the Gene for the Metalloprotease Enterotoxin of Bacteroides Fragilis. FEMS Microbiol. Lett. 1997, 146, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lim, K.C.; Huang, J.; Saidi, R.F.; Sears, C.L. Bacteroides Fragilis Enterotoxin Cleaves the Zonula Adherens Protein, E-Cadherin. Proc. Natl. Acad. Sci. USA 1998, 95, 14979–14984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.; Gwon, S.-Y.; Kim, M.S.; Lee, S.; Rhee, K.-J. Bacteroides Fragilis Toxin Induces IL-8 Secretion in HT29/C1 Cells through Disruption of E-Cadherin Junctions. Immune Netw. 2013, 13, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanfilippo, L.; Li, C.K.; Seth, R.; Balwin, T.J.; Menozzi, M.G.; Mahida, Y.R. Bacteroides Fragilis Enterotoxin Induces the Expression of IL-8 and Transforming Growth Factor-Beta (TGF-Beta) by Human Colonic Epithelial Cells. Clin. Exp. Immunol. 2000, 119, 456–463. [Google Scholar] [CrossRef]
- Sears, C.L.; Geis, A.L.; Housseau, F. Bacteroides Fragilis Subverts Mucosal Biology: From Symbiont to Colon Carcinogenesis. J. Clin. Invest. 2014, 124, 4166–4172. [Google Scholar] [CrossRef] [Green Version]
- Chung, L.; Thiele Orberg, E.; Geis, A.L.; Chan, J.L.; Fu, K.; DeStefano Shields, C.E.; Dejea, C.M.; Fathi, P.; Chen, J.; Finard, B.B.; et al. Bacteroides Fragilis Toxin Coordinates a Pro-Carcinogenic Inflammatory Cascade via Targeting of Colonic Epithelial Cells. Cell Host Microbe 2018, 23, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Boleij, A.; Hechenbleikner, E.M.; Goodwin, A.C.; Badani, R.; Stein, E.M.; Lazarev, M.G.; Ellis, B.; Carroll, K.C.; Albesiano, E.; Wick, E.C.; et al. The Bacteroides Fragilis Toxin Gene Is Prevalent in the Colon Mucosa of Colorectal Cancer Patients. Clin. Infect. Dis. 2015, 60, 208–215. [Google Scholar] [CrossRef]
- Wassenaar, T.M. Insights from 100 Years of Research with Probiotic E. Coli. Eur. J. Microbiol. Immunol. (Bp) 2016, 6, 147. [Google Scholar] [CrossRef] [Green Version]
- Falzano, L.; Filippini, P.; Travaglione, S.; Miraglia, A.G.; Fabbri, A.; Fiorentini, C. Escherichia Coli Cytotoxic Necrotizing Factor 1 Blocks Cell Cycle G2/M Transition in Uroepithelial Cells. Infect. Immun. 2006, 74, 3765–3772. [Google Scholar] [CrossRef] [Green Version]
- Malorni, W.; Fiorentini, C. Is the Rac GTPase-Activating Toxin CNF1 a Smart Hijacker of Host Cell Fate? FASEB J. 2006, 20, 606–609. [Google Scholar] [CrossRef]
- Jubelin, G.; Chavez, C.V.; Taieb, F.; Banfield, M.J.; Samba-Louaka, A.; Nobe, R.; Nougayrède, J.-P.; Zumbihl, R.; Givaudan, A.; Escoubas, J.-M.; et al. Cycle Inhibiting Factors (CIFs) Are a Growing Family of Functional Cyclomodulins Present in Invertebrate and Mammal Bacterial Pathogens. PLoS ONE 2009, 4, e4855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nougayrede, J.-P.; Homburg, S.; Taieb, F.; Boury, M.; Brzuszkiewicz, E.; Gottschalk, G.; Buchrieser, C.; Hacker, J.; Dobrindt, U.; Oswald, E. Escherichia Coli Induces DNA Double-Strand Breaks in Eukaryotic Cells. Science 2006, 313, 848–851. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Ramos, G.; Petit, C.R.; Marcq, I.; Boury, M.; Oswald, E.; Nougayrède, J.-P. Escherichia Coli Induces DNA Damage in Vivo and Triggers Genomic Instability in Mammalian Cells. Proc. Natl. Acad. Sci. USA 2010, 107, 11537–11542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faïs, T.; Delmas, J.; Barnich, N.; Bonnet, R.; Dalmasso, G. Colibactin: More Than a New Bacterial Toxin. Toxins 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut Microbiome Development along the Colorectal Adenoma–carcinoma Sequence. Nat. Commun. 2015, 6, 6528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, J.C.; Perez-Chanona, E.; Muhlbauer, M.; Tomkovich, S.; Uronis, J.M.; Fan, T.-J.; Campbell, B.J.; Abujamel, T.; Dogan, B.; Rogers, A.B.; et al. Intestinal Inflammation Targets Cancer-Inducing Activity of the Microbiota. Science 2012, 338, 120–123. [Google Scholar] [CrossRef] [Green Version]
- Dejea, C.M.; Fathi, P.; Craig, J.M.; Boleij, A.; Taddese, R.; Geis, A.L.; Wu, X.; DeStefano Shields, C.E.; Hechenbleikner, E.M.; Huso, D.L.; et al. Patients with Familial Adenomatous Polyposis Harbor Colonic Biofilms Containing Tumorigenic Bacteria. Science 2018, 359, 592–597. [Google Scholar] [CrossRef] [Green Version]
- Garcia, E.F.; Luciano, W.A.; Xavier, D.E.; da Costa, W.C.A.; de Sousa Oliveira, K.; Franco, O.L.; de Morais Júnior, M.A.; Lucena, B.T.L.; Picão, R.C.; Magnani, M.; et al. Identification of Lactic Acid Bacteria in Fruit Pulp Processing Byproducts and Potential Probiotic Properties of Selected Lactobacillus Strains. Front. Microbiol. 2016, 7, 1371. [Google Scholar] [CrossRef] [Green Version]
- Mughini-Gras, L.; Schaapveld, M.; Kramers, J.; Mooij, S.; Neefjes-Borst, E.A.; Pelt, W.V.; Neefjes, J. Increased Colon Cancer Risk after Severe Salmonella Infection. PLoS ONE 2018, 13, e0189721. [Google Scholar] [CrossRef] [Green Version]
- Pillar, C.M.; Gilmore, M.S. Enterococcal Virulence--Pathogenicity Island of E. Faecalis. Front. Biosci. 2004, 9, 2335–2346. [Google Scholar] [CrossRef] [Green Version]
- Balamurugan, R.; Rajendiran, E.; George, S.; Samuel, G.V.; Ramakrishna, B.S. Real-Time Polymerase Chain Reaction Quantification of Specific Butyrate-Producing Bacteria, Desulfovibrio and Enterococcus Faecalis in the Feces of Patients with Colorectal Cancer. J. Gastroenterol. Hepatol. 2008, 23, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Komiya, M.; Fujii, G.; Hamoya, T.; Nakanishi, R.; Fujimoto, K.; Tamura, S.; Kurokawa, Y.; Takahashi, M.; Ijichi, T.; et al. Preventive Effects of Heat-Killed Enterococcus Faecalis Strain EC-12 on Mouse Intestinal Tumor Development. Int. J. Mol. Sci. 2017, 18, 826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aisu, N.; Tanimura, S.; Yamashita, Y.; Yamashita, K.; Maki, K.; Yoshida, Y.; Sasaki, T.; Takeno, S.; Hoshino, S. Impact of Perioperative Probiotic Treatment for Surgical Site Infections in Patients with Colorectal Cancer. Exp. Ther. Med. 2015, 10, 966–972. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Xia, Y.; Chen, H.; Hong, L.; Feng, J.; Yang, J.; Yang, Z.; Shi, C.; Wu, W.; Gao, R.; et al. The Effect of Perioperative Probiotics Treatment for Colorectal Cancer: Short-Term Outcomes of a Randomized Controlled Trial. Oncotarget 2016, 7, 8432–8440. [Google Scholar] [CrossRef] [Green Version]
- Prakash, S.; Urbanska, A.M. Colon-targeted delivery of live bacterial cell biotherapeutics including microencapsulated live bacterial cells. Biologics: Targets and Therapy 2008, 2, 355–378. [Google Scholar] [CrossRef] [Green Version]
- Samot, J.; Badet, C. Clinical Microbiology Antibacterial Activity of Probiotic Candidates for Oral Health. Anaerobe 2013, 19, 34–38. [Google Scholar] [CrossRef]
- Samot, J.; Belkhelfa, H.; Haddioui, L.; Badet, C. Probiotic Properties of Lactobacilli That Could Be Used Against Periodontitis. Probiotics Heal. 2017, 5, 3. [Google Scholar] [CrossRef]
- Zhu, Y.; Xiao, L.; Shen, D.; Hao, Y. Competition between Yogurt Probiotics and Periodontal Pathogens in Vitro. Acta Odontol. Scand. 2010, 68, 261–268. [Google Scholar] [CrossRef]
- Jang, H.J.; Kang, M.S.; Yi, S.H.; Hong, J.Y.; Hong, S.P. Comparative Study on the Characteristics of Weissella Cibaria CMU and Probiotic Strains for Oral Care. Molecules 2016, 21. [Google Scholar] [CrossRef]
- Schellenberg, J.; Smoragiewicz, W.; Karska-Wysocki, B. A Rapid Method Combining Immunofluorescence and Flow Cytometry for Improved Understanding of Competitive Interactions between Lactic Acid Bacteria (LAB) and Methicillin-Resistant S. Aureus (MRSA) in Mixed Culture. J. Microbiol. Methods 2005, 65, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Güzel-Seydim, Z.B.; Ece Cagdas, M.D.; Seydim, A.C. Effect of Kefir on Fusobacterium Nucleatum Potentially Causing Intestinal Cancer. Funct. Foods Heal. Dis. 2016, 6, 469. [Google Scholar] [CrossRef]
- Bourrie, B.C.T.; Willing, B.P.; Cotter, P.D. The Microbiota and Health Promoting Characteristics of the Fermented Beverage Kefir. Front. Microbiol. 2016, 7, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, A.M.; Crispie, F.; Kilcawley, K.; O’Sullivan, O.; O’Sullivan, M.G.; Claesson, M.J.; Cotter, P.D. Microbial Succession and Flavor Production in the Fermented Dairy Beverage Kefir. mSystems 2016, 1, e00052-16. [Google Scholar] [CrossRef] [Green Version]
- Romani Vestman, N.; Chen, T.; Lif Holgerson, P.; Öhman, C.; Johansson, I. Oral Microbiota Shift after 12-Week Supplementation with Lactobacillus Reuteri DSM 17938 and PTA 5289; A Randomized Control Trial. PLoS ONE 2015, 10, e0125812. [Google Scholar] [CrossRef] [PubMed]
- Olsen, I.; Yamazaki, K. Can Oral Bacteria Affect the Microbiome of the Gut? J. Oral Microbiol. 2019, 11, 1586422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Wu, W.; Qin, H. Probiotics Modify Human Intestinal Mucosa-Associated Microbiota in Patients with Colorectal Cancer. Mol. Med. Rep. 2015, 12, 6119–6127. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Sample Type | CRC Samples | Noncancer Controls | Detection Method | Probiotic/Antibiotic Exclusion Criteria | Overrepresented Taxa | Underrepresented Taxa | Reference | |
|---|---|---|---|---|---|---|---|---|
| Tissue | 65 | 65 | 16S rRNA gene (V4) pyrosequencing | No probiotic or antibiotic exposure within 4 weeks | Fusobacterium, Streptococcus, Peptostreptococcus, Porphyromonas, and Selenomonas | Roseburia | [21] | |
| Tissue | 31 b | 20 | 16S rRNA gene (V3) pyrosequencing | No antibiotic exposure within 2 months or probiotic exposure within 2 weeks | Firmicutes, Fusobacteria, Lactococcus, and Fusobacterium | Pseudomonas and Escherichia/Shigella | [26] | |
| Tissue | 65 | 65 | 16S rRNA gene (V1–V3) pyrosequencing | - | Fusobacterium, Leptotrichia, and Campylobacter | Ruminococcus, Parabacteroides, Pseudoflavonifractor, Ruminococcaceae, and Holdemania | [25] | |
| Tissue and Mucosa | 15 | 21 | 16S rRNA gene (V4) amplicon sequencing | Recent antibiotic exposure or regular use of probiotics | Fusobacterium, Peptostreptococcus, and Selenomonas | Roseburia and Streptococcus | [22] | |
| Faecal | 9 a | 49 | Next Generation Sequence (NGS) analysis & terminal restriction fragment length polymorphism (T-RFLP) | Current use of antibiotics or regular use of probiotics | Actinomyces, Atopobium, Fusobacterium, Haemophilus, Bacteroides fragilis, Clostridium nexile, Actinomyces odontolyticus Heamophilus parainfluenzae, Fusobacterium varium, Prevotella stercorea, Veillonella dispar, and Streptococcus gordonii | Slackia and Eubacterium coprostanoligens | [24] | |
| Faecal | 46 | 56 | 16S rRNA gene (V3) pyrosequencing | No antibiotic exposure within 3 months | Fusobacterium, Enterococcus, Escherichia/Shigella, Klebsiella, Streptococcus, and Peptostreptococcus | Roseburia | [20] | |
| Faecal | 42 | 89 | 16S rRNA gene (V3–V4) pyrosequencing | - | Fusobacterium and Porphyromonas | Clostridia and Lachnospiraceae | [23] | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawrence, G.W.; Begley, M.; Cotter, P.D.; Guinane, C.M. Potential Use of Biotherapeutic Bacteria to Target Colorectal Cancer-Associated Taxa. Int. J. Mol. Sci. 2020, 21, 924. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030924
Lawrence GW, Begley M, Cotter PD, Guinane CM. Potential Use of Biotherapeutic Bacteria to Target Colorectal Cancer-Associated Taxa. International Journal of Molecular Sciences. 2020; 21(3):924. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030924
Chicago/Turabian StyleLawrence, Garreth W., Máire Begley, Paul D. Cotter, and Caitriona M. Guinane. 2020. "Potential Use of Biotherapeutic Bacteria to Target Colorectal Cancer-Associated Taxa" International Journal of Molecular Sciences 21, no. 3: 924. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030924