Understanding the Role of the Transcription Factor Sp1 in Ovarian Cancer: from Theory to Practice


 and
and 
Abstract
:1. Introduction
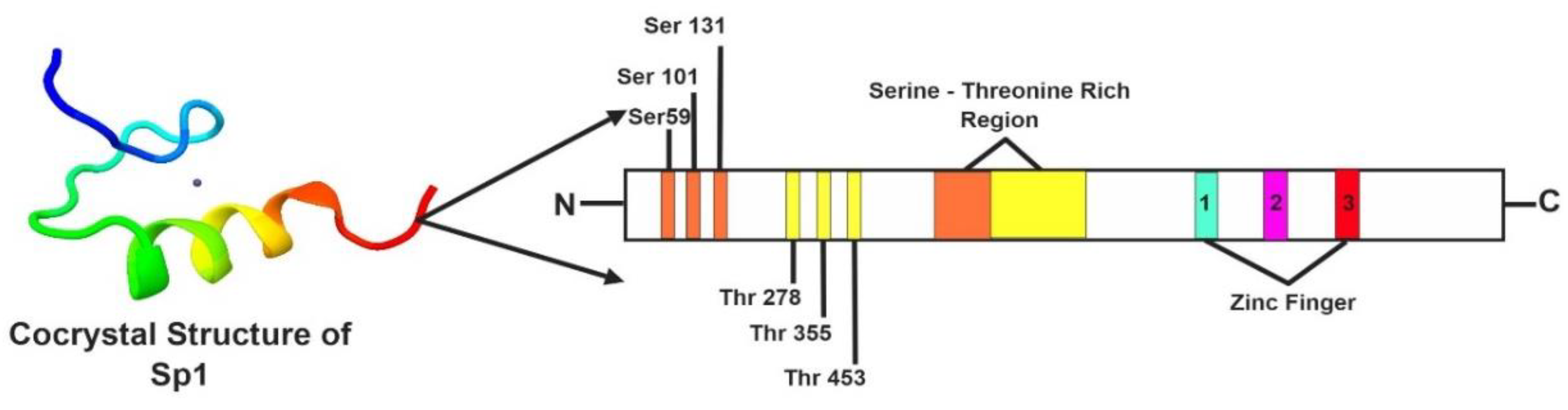
2. Genetic Makeup and Structure of Sp1
3. Regulation of Sp1
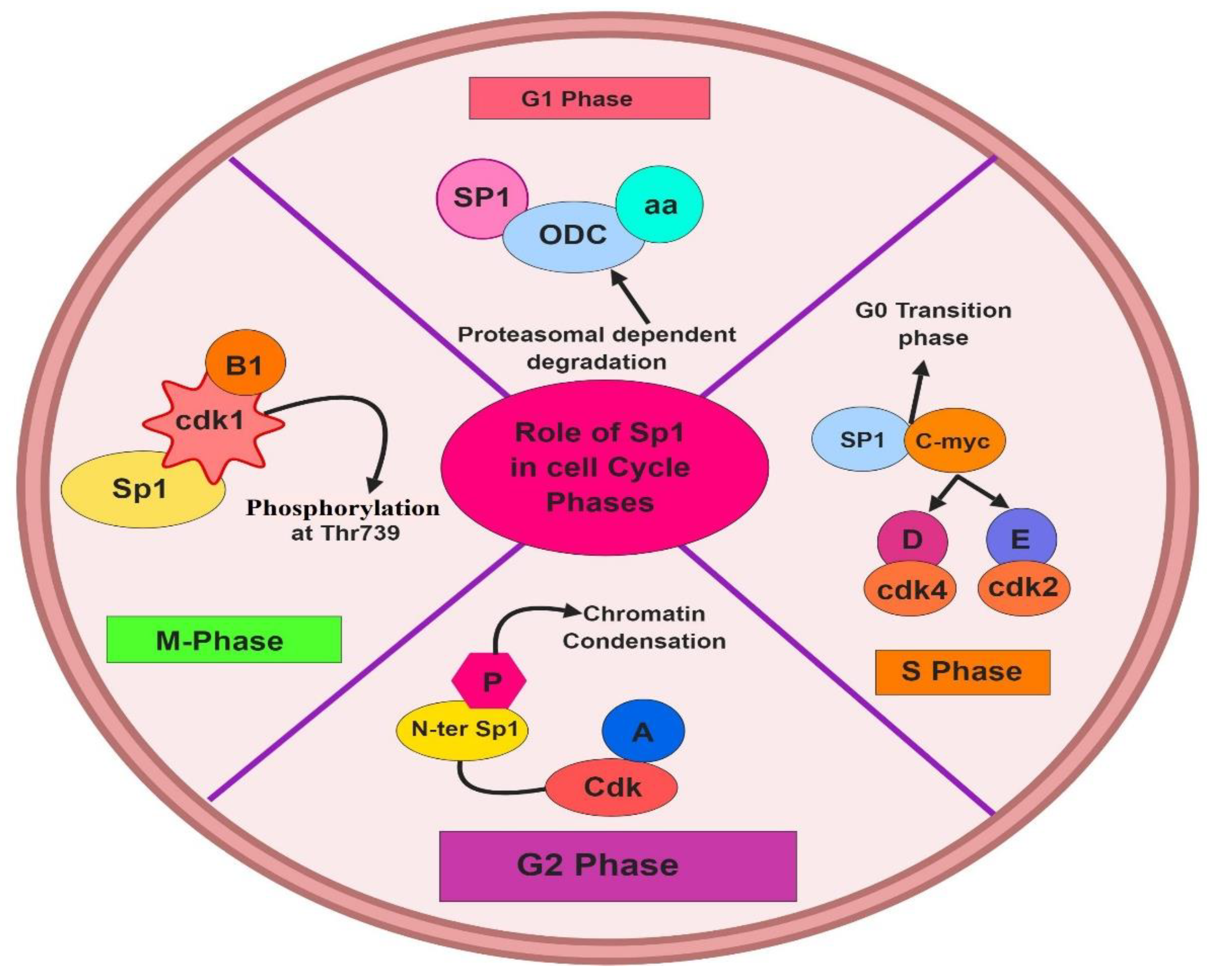
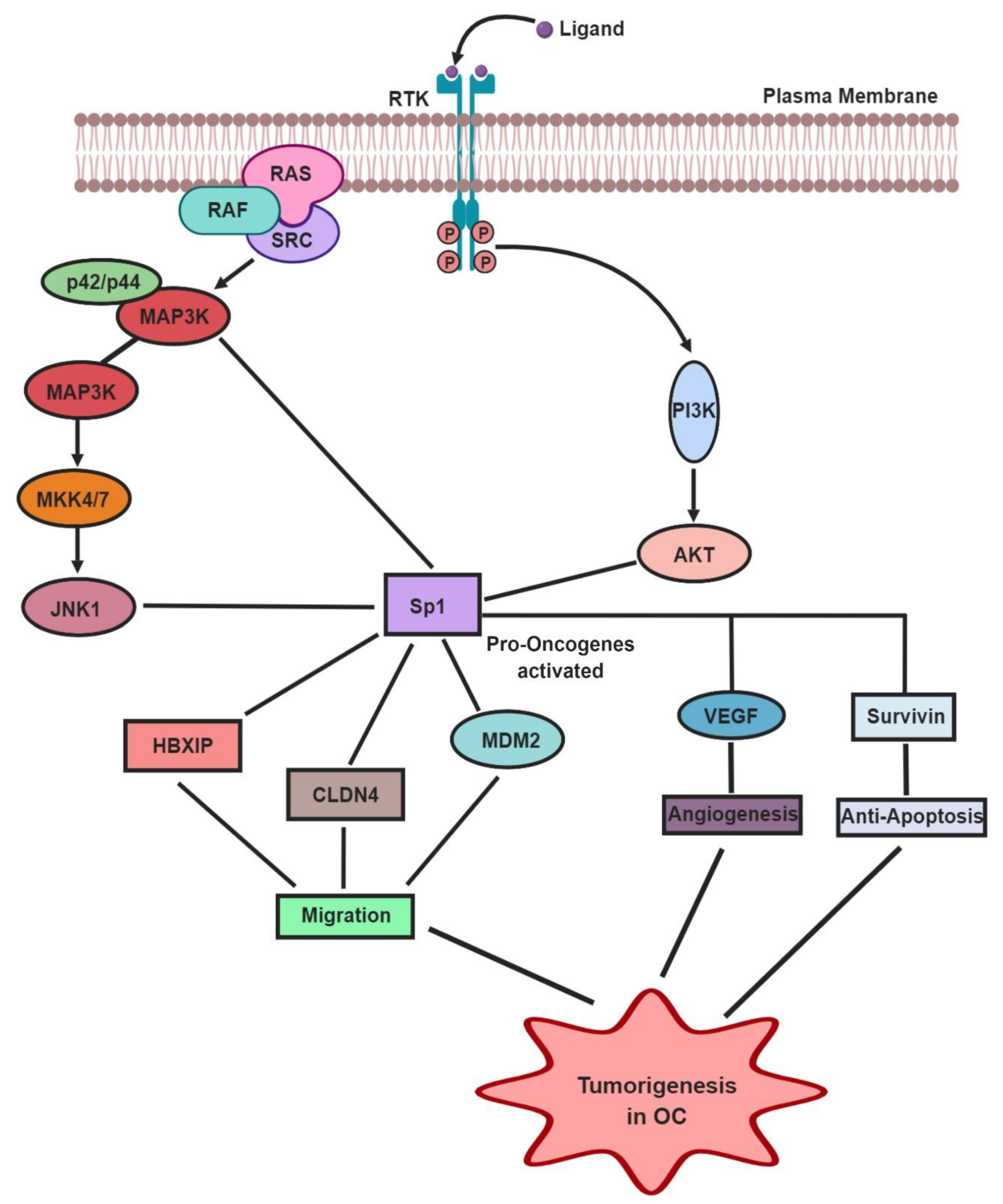
4. Role of Sp1 in the Normal Cell Cycle and OC Tumorigenesis
5. Effect of Sp1 TF in Angiogenesis and Anti-Apoptosis in OC
6. Sp1 as a Therapeutic Target in OC
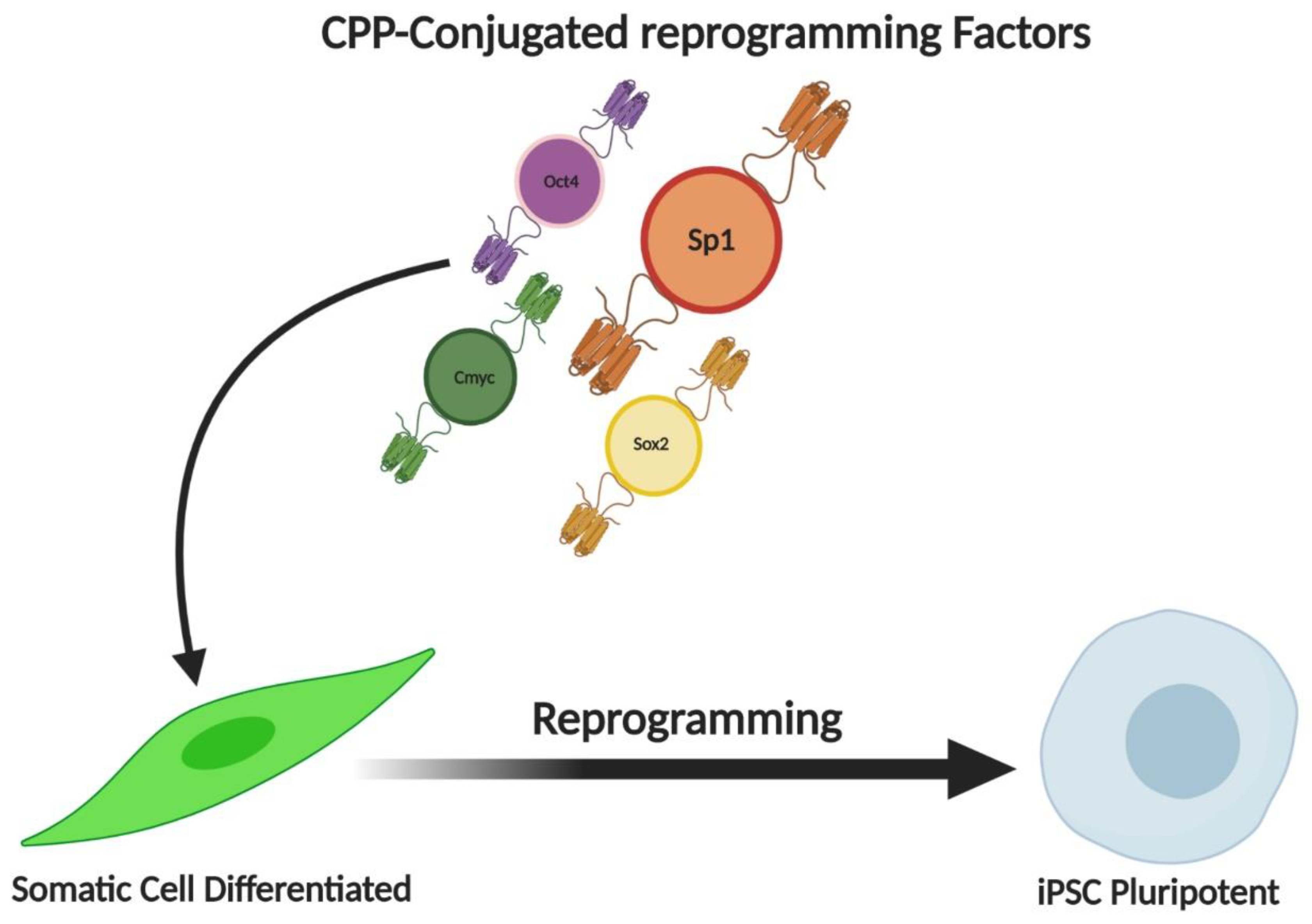
7. Influence of Sp1 on Cellular Reprogramming
8. Future Perspective
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| OC | Ovarian cancer |
| Sp1 | Specificity protein 1 |
| KLF | Kruppel-like family |
| TF(s) | Transcription factors |
| VEGF | Vascular endothelial growth factor |
| CCC | Ovarian clear cell carcinoma |
| JNK1 | Jun N—terminal kinase 1 |
| MAPK/ERK | mitogen-activated protein kinase//extracellular signal-regulated kinase |
| MDR | Multidrug resistance |
| IAP | Inhibitor of apoptosis protein |
| MTA | Mithramycin A |
| BCL2L1 | Bcl-2-like 1 |
| hTERT | Human telomerase Reverse Transcriptase |
| CLDN4 | Claudin 4 |
| HIFs | Hypoxia—Inducible Factors |
| LCFA | Long Chain Fatty Acid |
| HBXIP | Hepatitis B X—interacting protein |
| Skp2 | S—phase kinase—associated protein 2 |
| ROS | Reactive Oxygen Species |
| NO | Nitric Oxide |
| Raf | Rapidly Accelerated Fibrosarcoma |
| Sox2 | SRY-Box Transcription Factor 2 |
| Oct4 | Octamer-binding transcription factor 4 |
| Klf4 | Kruppel-like factor 4 |
| JNK1 | c-Jun N-terminal kinases |
| HBXIP | Hepatitis B X-interacting protein |
| CLDN | Claudin |
| VEGF | Vascular endothelial growth factor |
| ODC | Ornithine decarboxylase |
| CDK | Cyclin-dependent protein kinases |
References
- Mahalaxmi, I.; Devi, S.M.; Kaavya, J.; Arul, N.; Balachandar, V.; Santhy, K.S. New insight into NANOG: A novel therapeutic target for ovarian cancer (OC). Eur. J. Pharmacol. 2019, 852, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Mahalaxmi, I.; Santhy, K.S. An overview about mitochondrial DNA mutations in ovarian cancer. Alexandria J. Med. 2017, 53, 307–310. [Google Scholar] [CrossRef]
- Meinhold-Heerlein, I.; Hauptmann, S. The heterogeneity of ovarian cancer. Arch. Gynecol. Obstet. 2014, 289, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Mahalaxmi, I.; Santhy, K.S. Role and hallmarks of Sp1 in promoting ovarian cancer. J. Oncol. Sci. 2018, 4, 102–105. [Google Scholar] [CrossRef]
- Venugopal, A.; Chandran, M.; Eruppakotte, N.; Kizhakkillach, S.; Breezevilla, S.C.; Vellingiri, B. Monogenic diseases in India. Mutat. Res. 2018, 776, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Goto, S.; Takahashi, M.; Yasutsune, N.; Inayama, S.; Kato, D.; Fukuoka, M.; Kashiwaba, S.; Murakami, Y. Identification of GA-Binding Protein Trancription Factor Alpha Subunit (GABPA) as a Novel Bookmarking Factor. Int. J. Mol. Sci. 2019, 20, 1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabrowska, K.; Zielinska, M. Silencing of Transcription Factor SP1 Promotes SN1 Transporter Regulation by Ammonia in Mouse Cortical Astrocytes. Int. J. Mol. Sci. 2019, 20, 234. [Google Scholar] [CrossRef] [Green Version]
- Szpirer, J.; Szpirer, C.; Riviere, M.; Levan, G.; Marynen, P.; Cassiman, J.J.; Wiese, R.; DeLuca, H.F. The Sp1 transcription factor gene (SP1) and the 1,25-dihydroxyvitamin D3 receptor gene (VDR) are colocalized on human chromosome arm 12q and rat chromosome 7. Genomics 1991, 11, 168–173. [Google Scholar] [CrossRef]
- Safe, S.; Kim, K. Nuclear receptor-mediated transactivation through interaction with Sp proteins. Prog Nucleic Acid Res. Mol. Biol. 2004, 77, 1–36. [Google Scholar]
- Azizkhan, J.C.; Jensen, D.E.; Pierce, A.J.; Wade, M. Transcription from TATA-less promoters: Didhydrofolatereductase as a model. Crit. Rev. Eukaryotic Gene Exp. 1993, 3, 229–254. [Google Scholar]
- Dynan, W.S.; Tjian, R. The promoter-specific transcription factor Sp1 binds to upstream sequences in the SV40 early promoter. Cell 1983, 35, 79–87. [Google Scholar] [CrossRef]
- Dynan, W.S.; Sazer, S.; Tjian, R.; Schimke, R.T. Transcription factor Sp1 recognizes a DNA sequence in the mouse dihydrofolate reductase promoter. Nature 1986, 319, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wu, W.; Han, Q.; Wang, Y.; Li, C.; Zhang, P.; Xu, H. Post-translational modification control of RNA-binding protein hnRNPK function. Open Biol. 2019, 9, 180239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupuis-Maurin, V.; Brinza, L.; Baguet, J.; Plantamura, E.; Schicklin, S.; Chambion, S.; Macari, C.; Tomkowiak, M.; Deniaud, E.; Leverrier, Y.; et al. Overexpression of the transcription factor Sp1 activates the OAS-RNAse L-RIG-I pathway. PLoS ONE 2015, 10, e0118551. [Google Scholar] [CrossRef]
- Skubitz, A.P.; Pambuccian, S.E.; Argenta, P.A.; Skubitz, K.M. Differential gene expression identifies subgroups of ovarian carcinoma. Transl. Res. 2006, 148, 223–248. [Google Scholar] [CrossRef]
- Kelly, Z.L.; Michael, A.; Butler-Manuel, S.; Pandha, H.S.; Morgan, R.G.L. HOX genes in ovarian cancer. J. Ovarian Res. 2011, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.W.; Pan, C.W.; Yang, X.M.; Zhou, L.; Zheng, Z.Q.; Li, D.C. SP1 reduces autophagic flux through activating p62 in gastric cancer cells. Mol. Med. Rep. 2018, 17, 4633–4638. [Google Scholar] [CrossRef] [Green Version]
- ACE View, NCBI. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/IEB/Research/Acembly/av.cgi?db=human&c=Gene&l=SP1 (accessed on 12 August 2018).
- Narayan, V.A.; Kriwacki, R.W.; Caradonna, J.P. Structures of zinc finger domains from transcription factor Sp1. Insights into sequence–specific protein–DNA recognition. J. Biol. Chem. 1997, 272, 7801–7809. [Google Scholar] [CrossRef] [Green Version]
- Oka, S.; Shiraishi, Y.; Yoshida, T.; Ohkubo, T.; Sugiura, Y.; Kobayashi, Y. NMR structure of transcription factor Sp1 DNA binding domain. Biochemistry 2004, 43, 16027–16035. [Google Scholar] [CrossRef]
- Nagaoka, M.; Shiraishi, Y.; Sugiura, Y. Selected base sequence outside the target binding site of zinc finger protein Sp1. Nucleic Acids Res. 2001, 29, 4920–4929. [Google Scholar] [CrossRef] [Green Version]
- Kaczynski, J.; Cook, T.; Urrutia, R. Sp1- and Krüppel-like transcription factors. Genome Biol. 2003, 4, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, T.; Gebelein, B.; Belal, M.; Mesa, K.; Urrutia, R. Three conserved transcriptional repressor domains are a defining feature of the TIEG subfamily of Sp1-like zinc finger proteins. J. Biol. Chem. 1999, 274, 29500–29504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadonag, J.T.; Courey, A.J.; Ladika, J.; Tjian, R. Distinct regions of Sp1 modulate DNA binding and transcriptional activation. Science 1988, 242, 1566–1570. [Google Scholar] [CrossRef] [PubMed]
- Kadonaga, J.T.; Jones, K.A.; Tjian, R. Promoter specific activation of RNA polymerase II transcription by Sp1. Trends Biochem. Sci. 1986, 11, 20–23. [Google Scholar] [CrossRef]
- Suske, G. The Sp-family of transcription factors. Gene 1999, 238, 291–300. [Google Scholar] [CrossRef]
- Tan, N.; Khachigian, L. Sp1 phosphorylation and its regulation of gene transcription. Mol. Cell Biol. 2009, 29, 2483–2488. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Kimura, A.; Nagai, R.; Horikoshi, M. Regulation of interaction of the acetyltransferase region of p300 and the DNA-binding domain of Sp1 on and through DNA binding. Genes Cells 2000, 5, 29–41. [Google Scholar] [CrossRef]
- Jackson, S.P.; Tjian, R. O-glycosylation of eukaryotic transcription factors: Implications for mechanisms of transcriptional regulation. Cell 1988, 55, 125–133. [Google Scholar] [CrossRef]
- Stielow, B.; Sapetschnig, A.; Wink, C.; Kruger, I.; Suske, G. SUMO-modified Sp3 represses transcription by provoking local heterochromatic gene silencing. EMBO Rep. 2008, 9, 899–906. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Chuang, H.C.; Tsai, W.C.; Yang, H.C.; Ho, S.R.; Paterson, A.J.; Kulp, S.K.; Chen, C.S. Thiazolidinediones mimic glucose starvation in facilitating Sp1 degradation through the up-regulation of beta-transducin repeat-containing protein. Mol. Pharmacol. 2009, 76, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Kaldis, P. Cdks, cyclins and CKIs: Roles beyond cell cycle regulation. Development 2013, 140, 3079–3093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinstein, E.; Jundt, F.; Weinert, I.; Wernet, P.; Royer, H.D. Sp1 as G1 cell cycle phase specific transcription factor in epithelial cells. Oncogene 2002, 21, 1485–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, J.Y.; Wang, S.A.; Yang, W.B.; Yang, H.C.; Hung, C.Y.; Su, T.P.; Chang, W.C.; Hung, J.J. Sp1 phosphorylation by cyclin-dependent kinase 1/cyclin B1 represses its DNA-binding activity during mitosis in cancer cells. Oncogene 2012, 31, 4946–4959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fojas De Borja, P.; Collins, N.K.; Du, P.; Azizkhan-Clifford, J.; Mudryj, M. Cyclin A-CDK phosphorylates Sp1 and enhances Sp1-mediated transcription. EMBO J. 2001, 20, 5737–5747. [Google Scholar] [CrossRef] [Green Version]
- Sherr, C.J.; Roberts, J.M. CDK inhibitors: Positive and negative regulators of G1- phase progression. Genes Dev. 1999, 13, 1501–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Zhu, X.; Han, T.; You, X.; Liu, F.; Ye, L.; Zhang, X.; Wang, X.; Yao, Y. The oncoprotein hepatitis B X-interacting protein promotes the migration of ovarian cancer cells through the upregulation of S-phase kinase-associated protein 2 by Sp. Int. J. Oncol. 2014, 45, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Beishline, K.; Azizkhan-Clifford, J. Sp1 and the Hallmarks of cancer. FEBS J. 2014, 282, 224–258. [Google Scholar] [CrossRef] [PubMed]
- Farley, J.; Smith, L.M.; Darcy, K.M.; Sobel, E.; O’Connor, D.; Henderson, B.; Larry, E.M.; Birrer, M.J. Cyclin E expression is a significant predictor of survival in advanced, suboptimally debulked ovarian epithelial cancers: A Gynecologic Oncology Group Study. Cancer Res. 2003, 63, 1235–1241. [Google Scholar]
- Honda, H.; Pazin, M.J.; Ji, H.; Wernyj, R.P.; Morin, P.J. Crucial roles of Sp1 and epigenetic modifications in the regulation of the CLDN4 promoter in ovarian cancer cells. J. Biol. Chem. 2006, 281, 21433–21444. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.J.; Jung, E.M.; Lee, J.T.; Kim, S.; Park, J.W.; Choi, K.S.; Kwon, T.K. Mithramycin A sensitizes cancer cells to TRAIL-mediated apoptosis by down-regulation of XIAP gene promoter through Sp1 sites. Mol. Cancer Ther. 2006, 5, 2737–2746. [Google Scholar] [CrossRef] [Green Version]
- Liguang, Z.; Peishu, L.; Hongluan, M.; Hong, J.; Rong, W.; Wachtel, M.S.; Frezza, E.E. Survivin expression in ovarian cancer. Exp. Oncol. 2007, 29, 121–125. [Google Scholar] [PubMed]
- Wang, X.; Urvalek, A.M.; Liu, J.; Zhao, J. Activation of KLF8 transcription by focal adhesion kinase in human ovarian epithelial and cancer cells. J. Biol. Chem. 2008, 282, 13934–13942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knappskog, S.; Bjørnslett, M.; Myklebust, M.; Huijts, P.E.; Vreeswijk, M.P.; Edvardsen, H.; Guo, Y.; Zhang, X.; Yang, M.; Ylisaukko-Oja, S.K.; et al. The MDM2 promoter SNP285C/309G haplotype diminishes Sp1 transcription factor binding and reduces risk for breast and ovarian cancer in Cauca-sians. Cancer Cell 2011, 9, 273–282. [Google Scholar] [CrossRef]
- Koizume, S.; Ito, S.; Nakamura, Y.; Yoshihara, M.; Furuya, M.; Yamada, R.; Miyagi, E.; Hirahara, F.; Takano, Y.; Miyagi, Y. Lipid starvation and hypoxia synergistically activate ICAM1 and multiple genes in an Sp1-dependent manner to promote the growth of ovarian cancer. Mol. Cancer 2015, 14, 77. [Google Scholar] [CrossRef] [Green Version]
- MacLean, J.A.; Rao, M.K.; Doyle, K.M.; Richards, J.S.; Wilkinson, M.F. Regulation of the Rhox5 Homeobox Gene in Primary Granulosa Cells: Preovulatory Expression and Dependence on SP1/SP3 and GABP. Biol. Reprod. 2005, 73, 1126–1134. [Google Scholar] [CrossRef] [Green Version]
- Michael, C.A. Role of Sp Transcription Factors in the Regulation of Cancer Cell Metabolism. Genes Cancer 2011, 2, 712–719. [Google Scholar]
- Milanini-Mongiat, J.; Pouyssegur, J.; Pages, G. Identification of two Sp1 phosphorylation sites for p42/p44 mitogen-activated protein kinases. Their implication in vascular endothelial growth factor gene transcription. J. Biol. Chem. 2002, 277, 20631–20639. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Ye, W.; Wu, J.; Liu, L.; Yang, L.; Gao, L.; Cheng, B.; Zhang, F.L.; Yang, H.; Li, Y. Sp1-CD147 positive feedback loop promotes the invasion ability of ovarian cancer. Oncol. Rep. 2015, 34, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Sellak, H.; Yang, X.; Cao, X.; Cornwell, T.; Soff, G.A.; Lincoln, T. Sp1 transcription factor as a molecular target for nitric oxide– and cyclic nucleotide–mediated suppression of cGMP-dependent protein kinase-I expression in vascular smooth muscle cells. Circ. Res. 2002, 90, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Ji, Y.S.; Schmedtje, J.F. Sp1 increases expression of cyclooxygenase-2 in hypoxic vascular endothelium. Implications for the mechanisms of aortic aneurysm and heart failure. J. Biol. Chem. 2000, 275, 24583–24589. [Google Scholar] [CrossRef] [Green Version]
- Kieran, M.W.; Kalluri, R.; Cho, Y.J. The VEGF pathway in cancer and disease: Responses, resistance, and the path forward. Cold Spring Harb. Perspect. Med. 2012, 2, a006593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibuya, M. Vascular endothelial growth factor and its receptor system: Physiological functions in angiogenesis and pathological roles in various diseases. J. Biol. Chem. 2013, 153, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gille, H.; Kowalski, J.; Li, B.; LeCouter, J.; Moffat, B.; Zioncheck, T.F.; Pelletier, N.; Ferrara, N. Analysis of biological effects and signaling properties of Flt-1 (VEGFR-1) and KDR (VEGFR-2). A reassessment using novel receptor-specific vascular endothelial growth factor mutants. J. Biol. Chem. 2001, 276, 3222–3230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Li, Z.L.; Jiang, L.L.; Guo, Q.Q.; Liu, M.J.; Nan, K.J. Suppression of lung cancer cell invasion by LKB1 is due to the downregulation of tissue factor and vascular endothelial growth factor, partly dependent on SP1. Int. J. Oncol. 2014, 44, 1989–1997. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, W.; Guo, Z.; Ma, F.; Wu, Y.; Bai, Y.; Gong, W.; Chen, Y.; Cheng, T.; Zhi, Z.; et al. Inhibition of the transcription factor Sp1 suppresses colon cancer stem cell growth and induces apoptosis in vitro and in nude mouse xenografts. Oncol. Rep. 2013, 30, 1782–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, F.; Geng, J.; Li, X.; Qiao, C.; Luo, L.; Feng, J.; Dong, X.; Lv, M. SP1 promotes tumor angiogenesis and invasion by activating VEGF expression in an acquired trastuzumab‑resistant ovarian cancer model. Oncol. Rep. 2017, 38, 2677–2684. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Zhang, P.; Huang, J.; Ge, S.F. Sp1 and Sp3 regulate basal transcription of the survivin gene. Biochem. Biophys. Res. Commun. 2007, 356, 286–292. [Google Scholar] [CrossRef]
- Kayurma, M.M.; Khachigian, L.M. Sp1 Inhibits Proliferation and Induces Apoptosis in Vascular Smooth Muscle Cells by Repressing p21WAF1/Cip1 Transcription and Cyclin D1-Cdk4-p21WAF1/Cip1 Complex Formation. J. Biol. Chem. 2003, 278, 32537–32543. [Google Scholar] [CrossRef] [Green Version]
- Basha, R.; Ingersol, S.B.; Sankpal, U.T.; Ahmad, S.; Baker, C.H.; Edwards, J.R.; Holloway, R.W.; Kaja, S.; Abdelrahim, M. Tolfenamic acid inhibits ovarian cancer cell growth and decreases the expression of c-Met and survivin through suppressing specificity protein transcription factors. Gynecol. Oncol. 2011, 122, 163–170. [Google Scholar] [CrossRef]
- Miyata, K.; Yotsumoto, F.; Nam, S.O.; Odawara, T.; Manabe, S.; Ishikawa, T.; Itamochi, H.; Kigawa, J.; Takada, S.; Asahara, H.; et al. Contribution of transcription factor, SP1, to the promotion of HB-EGF expression in defense mechanism against the treatment of irinotecan in ovarian clear cell carcinoma. Cancer Med. 2014, 3, 1159–1169. [Google Scholar] [CrossRef]
- Chen, Y.T.; Tsai, H.P.; Wu, C.C.; Chen, C.Y.; Chai, C.Y.; Kwan, A.L. High-level Sp1 is Associated with Proliferation, Invasion, and Poor Prognosis in Astrocytoma. Pathol. Oncol. Res. 2019, 25, 1003–1013. [Google Scholar] [CrossRef]
- Gniazdowski, M.; Denny, W.A.; Nelson, S.M.; Czyz, M. Effects of anticancer drugs on transcription factor–DNA interactions. Expert Opin. Ther. Targets 2005, 9, 471–489. [Google Scholar] [CrossRef] [PubMed]
- Albertini, V.; Jain, A.; Vignati, S.; Napoli, S.; Rinaldi, A.; Kwee, I.; Nur-e-Alam, M.; Bergant, J.; Bertoni, F.; Carbone, G.M.; et al. Novel GC-rich DNA-binding compound produced by a genetically engineered mutant of the mithramycin producer Streptomyces argillaceus exhibits improved transcriptional repressor activity: Implications for cancer therapy. Nucleic Acids Res. 2006, 34, 1721–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcelo, F.; Scotta, C.; Ortiz-Lombardía, M.; Méndez, C.; Salas, J.A.; Portugal, J. Entropically-driven binding of mithramycin in the minor groove of C/G-rich DNA sequences. Nucleic Acids Res. 2007, 35, 2215–2226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcelo, F.; Ortiz-Lombardıa, M.; Martorell, M.; Oliver, M.; Méndez, C.; Salas, J.A.; Portugal, J. DNA binding characteristics of mithramycin and chromomycin analogues obtained by combinatorial biosynthesis. Biochemistry 2010, 49, 10543–10552. [Google Scholar] [CrossRef]
- Fernández-Guizán, A.; Mansilla, S.; Barcelo, F.; Vizcaíno, C.; Núñezd, L.N.; Morís, F.; Gonzalez, S.; Portugal, J. The activity of a novel mythramycin analogue is related to its binding to DNA, cellular uptake, and inhibition of Sp1- driven gene transcription. Chem. Biol. Interact. 2014, 219, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Martín, B.; Vaquero, A.; Priebe, W.; Portugal, J. BisanthracyclineWP631 inhibits basal and Sp1-activated transcription initiation in vitro. Nucleic Acids Res. 1999, 27, 3402–3409. [Google Scholar] [CrossRef] [Green Version]
- Mansilla, S.; Priebe, W.; Portugal, J. Sp1-targeted inhibition of gene transcription by WP631 in transfected lymphocytes. Biochemistry 2004, 43, 7584–7592. [Google Scholar] [CrossRef]
- Mansilla, S.; Priebe, W.; Portugal, J. Transcriptional changes facilitate mitotic catastrophe in tumour cells that contain functional p53. Eur. J. Pharmacol. 2006, 540, 34–45. [Google Scholar] [CrossRef]
- Mansilla, S.; Portugal, J. Sp1 transcription factor as a target for anthracyclines: Effects on gene transcription. Biochimie 2008, 90, 976–987. [Google Scholar] [CrossRef]
- Dalvai, M.; Mondesert, O.; Bugler, B.; Manenti, S.; Ducommun, B.; Dozier, C. Doxorubicin promotes transcriptional upregulation of Cdc25B in cancer cells by releasing Sp1 from the promoter. Oncogene 2013, 32, 5123–5128. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ling, X.; Pan, D.; Apontes, P.; Song, L.; Liang, P.; Altieri, D.C.; Beerman, T.; Li, F. Molecular mechanism of inhibition of survivin transcription by the GC-rich sequence-selective DNA binding antitumor agent, hedamycin: Evidence of survivin down-regulation associated with drug sensitivity. J. Biol. Chem. 2005, 280, 9745–9751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaquero, A.; Portugal, J. Modulation of DNA–protein interactions in the P1 and P2 c-myc promoters by two intercalating drugs. Eur. J. Biochem. 1998, 251, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.H.; Lenzi, M.; Schwartz, E.L. The Sp1 transcription factor contributes to the tumor necrosis factor-induced expression of the angiogenic factor thymidine phosphorylase in human colon carcinoma cells. Oncogene 2002, 21, 8477–8485. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahim, M.; Baker, C.H.; Abbruzzese, J.L.; Safe, S. Tolfenamic acid and pancreatic cancer growth, angiogenesis, and Sp protein degradation. J. Natl. Cancer Inst. 2006, 98, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahim, M.; Safe, S. Cyclooxygenase-2 inhibitors decrease vascular endothelial growth factor expression in colon cancer cells by enhanced degradation of Sp1 and Sp4 proteins. Mol. Pharmacol. 2005, 68, 317–329. [Google Scholar] [CrossRef]
- Pathi, S.; Jutooru, I.; Chadalapaka, G.; Nair, V.; Lee, S.O.; Safe, S. Aspirin inhibits colon cancer cell and tumor growth and downregulates specificity protein (Sp) transcription factors. PLoS ONE 2012, 7, e48208. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Sun, M.; Shi, W.; Yang, Q.; Chen, C.; Wang, Z.; Zhou, X. Arsenic trioxide suppresses transcription of hTERT through down-regulation of multiple transcription factors in HL-60 leukemia cells. Toxicol. Lett. 2015, 232, 481–489. [Google Scholar] [CrossRef]
- Chadalapaka, G.; Jutooru, I.; Chintharlapalli, S.; Papineni, S.; Smith, R.; Li, X.; Safe, S. Curcumin decreases specificity protein expression in bladder cancer cells. Cancer Res. 2008, 68, 5345–5354. [Google Scholar] [CrossRef] [Green Version]
- Jutooru, I.; Chadalapaka, G.; Lei, P.; Safe, S. Inhibition of NFκB and pancreatic cancer cell and tumor growth by curcumin is dependent on specificity protein downregulation. J. Biol Chem 2010, 285, 25332–25344. [Google Scholar] [CrossRef] [Green Version]
- Sreevalsan, S.; Safe, S. The cannabinoid WIN 55,212-2 decreases specificity protein transcription factors and the oncogenic cap protein eIF4E in colon cancer cells. Mol. Cancer Ther. 2013, 12, 2483–2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.A.; Lee, Y.J.; Ban, J.O.; Lee, Y.J.; Lee, S.H.; Cho, M.K.; Nam, H.S.; Hong, J.T.; Shim, J.H. The flavonoid resveratrol suppresses growth of human malignant pleural mesothelioma cells through direct inhibition of specificity protein 1. Int. J. Mol. Med. 2012, 30, 21–27. [Google Scholar] [PubMed] [Green Version]
- Lombo, F.; Menendez, N.; Salas, J.A.; Mendez, C. The aureolic acid family of antitumor compounds: Structure, mode of action, biosynthesis, and novel derivatives. Appl. Microbiol. Biotechnol. 2006, 73, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vizcaıno, C.; Mansilla, S.; Nu´n˜ez, L.E.; Méndez, C.; Salas, J.A.; Morís, F.; Portugal, J. Novel mithramycins abrogate the involvement of protein factors in the transcription of cell cycle control genes. Biochem. Pharmacol. 2012, 84, 1133–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, T.J.; Lacasse, E.C.; Durkin, J.P.; Vanderhyden, B.C. Downregulation of XIAP expression in ovarian cancer cells induces cell death in vitro and in vivo. Int. J. Cancer 2008, 122, 1430–1434. [Google Scholar] [CrossRef]
- Previdi, S.; Malek, A.; Albertini, V.; Riva, C.; Capella, C.; Broggini, M.; Carbone, G.M.; Rohr, J.; Catapano, C.V. Inhibition of Sp1-dependent transcription and antitumor activity of the new aureolic acid analogue mithramycin SDK and SK in human ovarian cancer xenografts. Gynecol. Oncol. 2010, 118, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Guizan, A.; Lopez-Soto, A.; Acebes-Huerta, A.; Huergo-Zapico, L.; Villa-Álvarez, M.; Núñez, L.E.; Moris, F.; Gonzalez, S. Pleiotropic Anti-Angiogenic and Anti-Oncogenic Activities of the Novel Mithralog Demycarosyl-3D-ß-D-Digitoxosyl-Mithramycin SK (EC-8042). PLoS ONE 2015, 10, e0140786. [Google Scholar] [CrossRef] [Green Version]
- Chintharlapalli, S.; Papineni, S.; Lei, P.; Pathi, S.; Safe, S. Betulinic acid inhibits colon cancer cell and tumor growth and induces proteasome-dependent and -independent downregulation of specificity proteins (Sp) transcription factors. BMC Cancer 2011, 11, 371. [Google Scholar] [CrossRef] [Green Version]
- Chintharlapalli, S.; Papineni, S.; Ramaiah, S.K.; Safe, S. Betulinic acid inhibits prostate cancer growth through inhibition of specificity protein transcription factors. Cancer Res. 2007, 67, 2816–2823. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Li, Y.; Xie, C.; Yin, X.; Liu, Y.; Cao, Y.; Fang, Y.; Lin, X.; Xu, Y.; Xu, W.; et al. miR-145 sensitizes ovarian cancer cells to paclitaxel by targeting Sp1 and Cdk6. Int. J. Cancer 2014, 135, 1286–1296. [Google Scholar] [CrossRef]
- Weidle, U.H.; Birzele, F.; Kollmorgen, G.; Nopora, A. Potential microRNA-related Targets for Therapeutic Intervention with Ovarian Cancer Metastasis. Cancer Genomics Proteomics 2018, 15, 1–15. [Google Scholar] [PubMed] [Green Version]
- Mak, C.S.; Yung, M.M.; Hui, L.M.; Leung, L.L.; Liang, R.; Chen, K.; Liu, S.S.; Qin, Y.; Leung, T.H.; Lee, K.F.; et al. MicroRNA-141 enhances anoikis resistance in metastatic progression of ovarian cancer through targeting KLF12/Sp1/survivin axis. Mol. Cancer 2017, 16, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.R.; Chen, X.J.; Chen, W.; Wu, L.F.; Jiang, H.P.; Lin, D.; Wang, L.J.; Wang, W.; Guo, S.Q. Sp1 contributes to radioresistance of cervical cancer through targeting G2/M cell cycle checkpoint CDK1. Cancer Manag. Res. 2019, 11, 5835–5844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bairong, X.; Yan, H.; Hong, C.; Shanshan, Y.; Tianbo, L.; Mei, L.; Ge, L. Long non-coding RNA ZFAS1 interacts with miR-150-5p to regulate Sp1 expression and ovarian cancer cell malignancy. Oncotarget 2017, 8, 19534–19546. [Google Scholar]
- Hedrick, E.; Li, X.; Safe, S. Penfluridol Represses Integrin Expression in Breast Cancer through Induction of Reactive Oxygen Species and Downregulation of Sp Transcription Factors. Mol. Cancer Ther. 2017, 16, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, E.; Lee, S.O.; Doddapaneni, R.; Singh, M.; Safe, S. NR4A1 antagonists inhibit β1-integrin-dependent breast cancer cell migration. Mol. Cell Biol. 2016, 36, 1383–1394. [Google Scholar] [CrossRef] [Green Version]
- Lacey, A.; Hedrick, E.; Li, X.; Patel, K.; Doddapaneni, R.; Singh, M.; Safe, S. Nuclear receptor 4A1 (NR4A1) as a drug target for treating rhabdomyosarcoma (RMS). Oncotarget 2016, 7, 31257–31269. [Google Scholar] [CrossRef]
- Lee, S.O.; Li, X.; Hedrick, E.; Jin, U.H.; Tjalkens, R.B.; Backos, D.S.; Li, L.; Zhang, Y.; Wu, Q.; Safe, S. Diindolylmethane analogs bind NR4A1 and are NR4A1 antagonists in colon cancer cells. Mol. Endocrinol. 2014, 28, 1729–1739. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.O.; Jin, U.H.; Kang, J.H.; Kim, S.B.; Guthrie, A.S.; Sreevalsan, S.; Lee, J.S.; Safe, S. The orphan nuclear receptor NR4A1 (Nur77) regulates oxidative and endoplasmic reticulum stress in pancreatic cancer cells. Mol. Cancer Res. 2014, 12, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Jutooru, I.; Lei, P.; Kim, K.; Lee, S.O.; Brents, L.K.; Prather, P.L.; Safe, S. Betulinic acid targets YY1 and ErbB2 through cannabinoid receptor-dependent disruption of microRNA-27a: ZBTB10 in breast cancer. Mol. Cancer Ther. 2012, 11, 1421–1431. [Google Scholar] [CrossRef] [Green Version]
- Potashkin, J.A.; Bottero, V.; Santiago, J.A.; Quinn, J.P. Computational identification of key genes that may regulate gene expression reprogramming in Alzheimer’s patients. PLoS ONE. 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jin, K.; Song, J.; Zuo, Q.; Yang, H.; Zhang, Y.; Li, B. Functional characterization of the Sox2, c-Myc, and Oct4 promoters. J. Cell. Biochem. 2019, 120, 332–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zuo, Q.; Wang, Y.; Song, J.; Yang, H.; Zhang, Y.; Li, B. Inducing goat pluripotent stem cells with four transcription factor mRNAs that activate endogenous promoters. BMC Biotech. 2017, 17, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soleimani, T.; Falsafi, N.; Fallahi, H. Dissection of regulatory elements during direct conversion of somatic cells into neurons. J. Cell Biochem. 2017, 118, 3158–3170. [Google Scholar] [CrossRef]
- Singh, V.P.; Mathison, M.; Patel, V.; Sanagasetti, D.; Gibson, B.W.; Yang, J.; Rosengart, T.K. MiR-590 Promotes Transdifferentiation of Porcine and Human Fibroblasts Toward a Cardiomyocyte-Like Fate by Directly Repressing Specificity Protein 1. J. Am. Heart Ass. 2016, 5, e003922. [Google Scholar]
- Kwon, Y.W.; Jang, S.; Paek, J.S.; Lee, J.W.; Cho, H.J.; Yang, H.M.; Kim, H.S. E-Ras improves the efficiency of reprogramming by facilitating cell cycle progression through JNK–Sp1 pathway. Stem Cell Res. 2015, 15, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Jhamb, D.; Rao, N.; Milner, D.J.; Song, F.; Cameron, J.A.; Stocum, D.L.; Palakal, M.J. Network based transcription factor analysis of regenerating axolotl limbs. BMC Bioinform. 2011, 12, 80. [Google Scholar] [CrossRef] [Green Version]
- Gurdon, J.; Elsdale, T.; Fischberg, M. Sexually Mature Individuals of Xenopus laevis from the Transplantation of Single Somatic Nuclei. Nature 1958, 182, 64–65. [Google Scholar] [CrossRef]
- Stadtfeld, M.; Hochedlinger, K. Induced pluripotency: History, mechanisms, and applications. Genes Dev. 2010, 24, 2239–2263. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, H.; Balasubramanian, V.; Iyer, M.; Venugopal, A.; Subramaniam, M.D.; Cho, S.G.; Vellingiri, B. mTOR signalling pathway - A root cause for idiopathic autism? BMB Rep. 2019, 52, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.K.; Jeong, Y.; Cho, S.; Cho, S.G. Systematic expression alteration analysis of master reprogramming factor OCT4 and its three pseudogenes in human cancer and their prognostic outcomes. Sci. Rep. 2018, 8, 14806. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Islam, S.M.R.; Kwak, K.; Rahman, M.S.; Cho, S.G. PROM1 and PROM2 expression differentially modulates clinical prognosis of cancer: A multiomics analysis. Cancer Gene Ther. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Adams, C.C.; Workman, J.L. Nucleosome binding by the constitutive transcription factor Sp1. J. Biol. Chem. 1994, 269, 7756–7763. [Google Scholar] [PubMed]
- Harrington, M.A.; Jones, P.A.; Imagawa, M.; Karin, M. Cytosine methylation does not affect binding of transcription factor Sp1. Proc. Natl. Acad. Sci. USA 1988, 85, 2066–2070. [Google Scholar] [CrossRef] [Green Version]
- Balachandar, V.; Kumar, R.K.; Varsha, P.; Mohana Devi, S.; Lakshman Kumar, B.; Manikantan, P.; Sasikala, K.; Malathi, J.; Brahmanandan, G.M.; Khanna, D.; et al. Evaluation of genetic alterations in inhabitants of a naturally high-level background radiation and Kudankulam nuclear power project site in India. Asian Pac. J. Cancer Prev. 2011, 12, 35–41. [Google Scholar]
- Balachandar, V.; Sureshkumar, S.; Mohana Devi, S.; Balamuralikrishnan, B.; Arun, M.; Karthickkumar, A.; Prakash, V.; Shafiahammedkhan, M.; Kathannan, S.; Pappuswamy, M.; et al. Cytogenetic endpoints and Xenobiotic gene polymorphism in lymphocytes of hospital workers chronically exposed to ionizing radiation in Cardiology, Radiology and Orthopedic Laboratories. Ecotox. Environ. Safe. 2014, 100, 266–274. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Compound Name | Binding Site | Reactions | References |
|---|---|---|---|---|
| 1 | Mithramycin A | Sp1 gene promoters | Alters Sp1 and DNA interactions | [64,65,66,67] |
| 2 | Daunorubicin | Binds to DNA with higher affinity | Inhibits Sp1-DNA interactions & gene transcription | [68,69,70] |
| 3 | WP631(bis-intercalating anthracycline) | Binds to DNA with higher affinity | Efficient inhibitor for transcription initiation of Sp1 containing binding sites & Sp1—activated transcription | [69,71] |
| 4 | Doxorubicin | Activates promoter of Cdc25B | Inhibits Sp1 binding & increases NF-Y binding to promoter and keeps P53 alive | [72] |
| 5 | Hedamycin | Down—regulates surviving expression | Abolishes Sp1 binding to putative binding elements & modulates viability of cancer cells | [73] |
| 6 | Elsamicin A | DNA—protein interactions in c–myc promoters | Affects Sp1 binding in a dose—dependent manner | [74] |
| 7 | Actinomycin D | DNA—protein interactions | Sp1 TFs induce TNF expressions on angiogenic factors in cancer cells | [75] |
| 8 | Tolfenamic acid | Drug—DNA interaction | Increases ubiquitination of Sp1 as well as the proteasome–dependent degradation (downregulates Sp1) | [76,77] |
| 9 | Aspirin | Cells that response to sequestration of zinc ions | It induces caspase–dependent cleavage of Sp1 protein factors | [78] |
| 10 | Arsenic trioxide | Human telomerase reverse transcriptase (hTERT) gene | Suppresses transcription of hTERT gene through regulation Sp1 TF | [79] |
| 11 | Curcumin (diferuloylmethane) | Not described | Induces proteasome–dependent down—regulation of Sp1 proteins | [80,81] |
| 12 | Betulinic acid | Not described | Decreases expression of Sp1 TF in cancer cells | [82] |
| 13 | Resveratrol (3,5,4′-trihydroxy-trans-stilbene) | Not described | Inhibits cell growth and Sp1 TF directly | [83] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vellingiri, B.; Iyer, M.; Devi Subramaniam, M.; Jayaramayya, K.; Siama, Z.; Giridharan, B.; Narayanasamy, A.; Abdal Dayem, A.; Cho, S.-G. Understanding the Role of the Transcription Factor Sp1 in Ovarian Cancer: from Theory to Practice. Int. J. Mol. Sci. 2020, 21, 1153. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031153
Vellingiri B, Iyer M, Devi Subramaniam M, Jayaramayya K, Siama Z, Giridharan B, Narayanasamy A, Abdal Dayem A, Cho S-G. Understanding the Role of the Transcription Factor Sp1 in Ovarian Cancer: from Theory to Practice. International Journal of Molecular Sciences. 2020; 21(3):1153. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031153
Chicago/Turabian StyleVellingiri, Balachandar, Mahalaxmi Iyer, Mohana Devi Subramaniam, Kaavya Jayaramayya, Zothan Siama, Bupesh Giridharan, Arul Narayanasamy, Ahmed Abdal Dayem, and Ssang-Goo Cho. 2020. "Understanding the Role of the Transcription Factor Sp1 in Ovarian Cancer: from Theory to Practice" International Journal of Molecular Sciences 21, no. 3: 1153. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031153