Cbx2, a PcG Family Gene, Plays a Regulatory Role in Medaka Gonadal Development

1
Key Laboratory of Freshwater Aquatic Genetic Resources, Ministry of Agriculture, Shanghai Collaborative Innovation for Aquatic Animal Genetics and Breeding, National Demonstration Center for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306, China
2
Laboratory for Marine Fisheries Science and Food Production Processes, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China
3
Laboratory of Quality and Safety Risk Assessment for Aquatic Product on Storage and Preservation, Ministry of Agriculture, Shanghai Ocean University, Shanghai 201306, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(4), 1288; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041288
Submission received: 3 January 2020
/
Revised: 5 February 2020
/
Accepted: 7 February 2020
/
Published: 14 February 2020
(This article belongs to the Special Issue Novel Physiology and Molecular Pathology of Reproduction, Novel Treatments of Infertility)
Abstract
:Chromobox homolog 2 (CBX2), a key member of the polycomb group (PcG) family, is essential for gonadal development in mammals. A functional deficiency or genetic mutation in cbx2 can lead to sex reversal in mice and humans. However, little is known about the function of cbx2 in gonadal development in fish. In this study, the cbx2 gene was identified in medaka, which is a model species for the study of gonadal development in fish. Transcription of cbx2 was abundant in the gonads, with testicular levels relatively higher than ovarian levels. In situ hybridization (ISH) revealed that cbx2 mRNA was predominately localized in spermatogonia and spermatocytes, and was also observed in oocytes at stages I, II, and III. Furthermore, cbx2 and vasa (a marker gene) were co-localized in germ cells by fluorescent in situ hybridization (FISH). After cbx2 knockdown in the gonads by RNA interference (RNAi), the sex-related genes, including sox9 and foxl2, were influenced. These results suggest that cbx2 not only plays a positive role in spermatogenesis and oogenesis but is also involved in gonadal differentiation through regulating the expression levels of sex-related genes in fish.
1. Introduction
The polycomb group (PcG) protein family was initially identified in Drosophila [1]. In higher eukaryotes, it is mainly involved in maintaining a series of cellular physiological activities, such as cell differentiation, cell cycle regulation, cell senescence, and X chromosome inactivation [2]. PcG proteins are assembled into two Polycomb Repressive Complexes, PRC1 and PRC2 [3]. In embryonic stem cells, whole-genome target analysis revealed that the two complexes were localized in the promoter region of developmental regulators [4]. Functionally, PRC1 and PRC2 exhibit enzymatic activities to modify histones and are crucial for epigenetic regulation via histone methylation [5,6,7].
As a core component of PRC1, chromobox (CBX) homolog protein performs an indispensable function regarding stem cell maintenance and embryo development [8]. In mammals, the CBX family contains five proteins, CBX2, CBX4, CBX6, CBX7, and CBX8 [9]. Although they have a common conserved domain, their molecular size, tissue distribution, and biological functions are quite different. In recent years, cbx2 has become increasingly important due to the discovery of the role of CBX2 in human and mouse gonadal development. In mice, a lack of cbx2 can lead to gonadal hypoplasia and sex reversal (testis–ovary) [7]. Sex reversal was also observed in humans due to cbx2 gene mutation [10]. The CBX2 protein mainly participates in the recruitment and stabilization of PRC1 in mitotic chromosomes [11]. Previous reports suggested that CBX2, a member of PcG, was an epigenetically modified transcriptional repressor. Recently, it was also found to serve as a transcriptional activator to regulate the expression of downstream genes [12,13]. However, the functional role of cbx2 in gonadal development is still unclear in vertebrates other than mammals.
Medaka (Oryzias latipes) is regarded as a model animal to investigate the gonadal development of vertebrates [14]. Like mammals, medaka has XY sex chromosomes and was the first species to define the sex-determining gene dmy in fish [15,16]. Due to its small size (2-3 cm), short sexual maturity cycle (3 months), and gender plasticity, medaka is usually used to explore gonadal development in vertebrates. In this work, we identified medaka cbx2 via PCR cloning and sequencing, detected the relative expression of cbx2 mRNA in different tissues using real-time PCR, and established an expression profile of cbx2 mRNA in medaka gonads via digoxigenin (DIG)-labeled in situ hybridization (ISH) and fluorescent in situ hybridization (FISH). Furthermore, the expression of cbx2 mRNA was knocked down using RNA interference (RNAi), then the expression levels of cbx2 and sex-related genes, including foxl2 and sox9, were analyzed to clarify the function of cbx2 in the gonadal development of fish.
2. Results
2.1. Molecular Characterization and Phylogenetic Analyses of cbx2
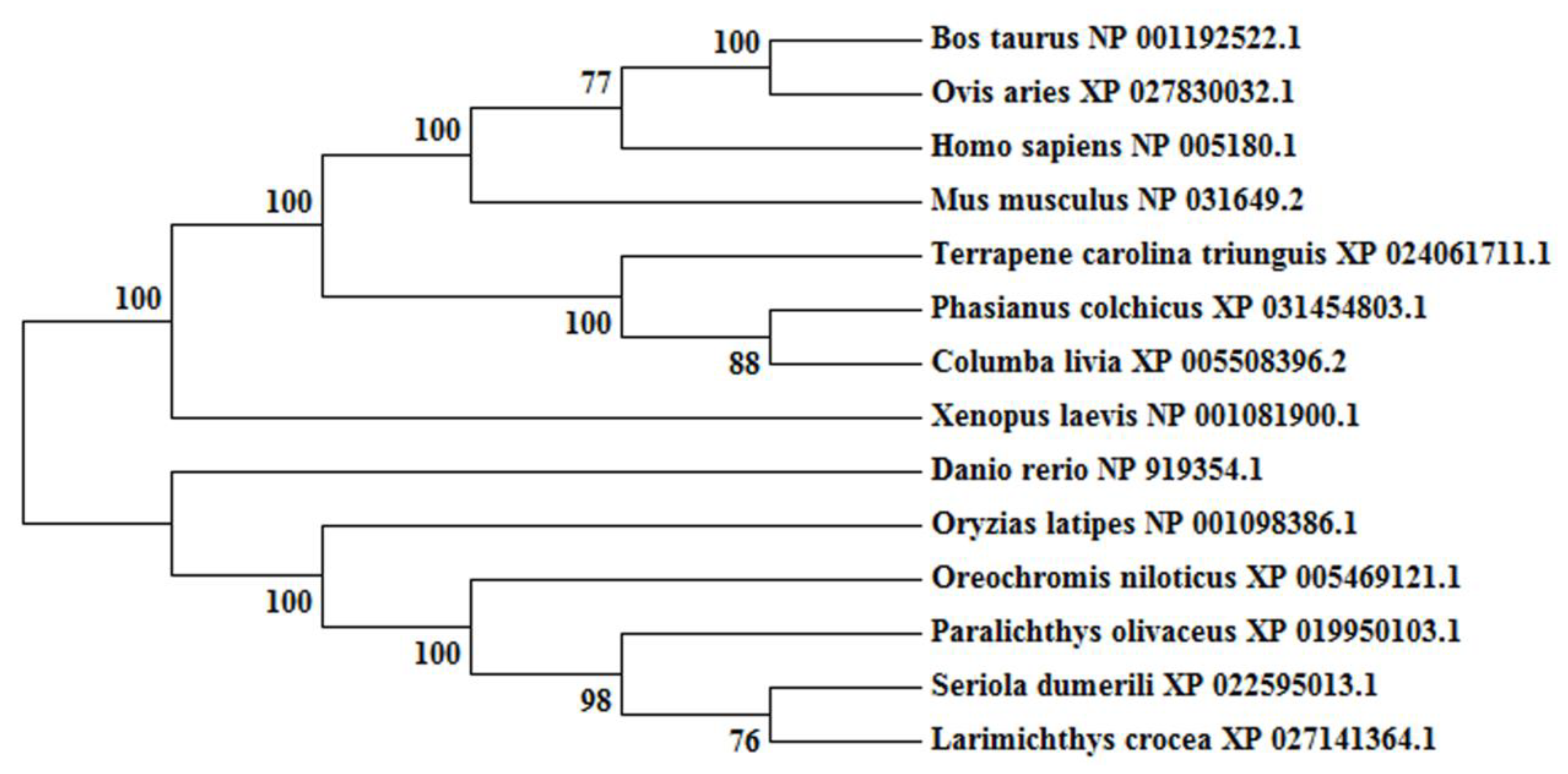
The obtained cDNA of cbx2 was 1443 bp in length and encoded 480 amino acid residues (Figure 1A). The molecular mass of the deuced CBX2 protein was 51.5 kDa and its theoretical isoelectric point was 10.12. Among these amino acids, the ratios of serine were the highest (14.0%). The CBX2 protein had no signal peptide or transmembrane region, indicating that it was neither secreted nor transmembrane-located. It contained one Chromo domain (11–63 bp), four low complex domains, and a Pfam domain (439–471 bp) (Figure 1B). Comparing the amino acid sequences of CBX2 in different species, we found that the anterior and posterior amino acid sequences of the CBX2 protein were more conservative and the middle parts were less conservative (Figure 2). A phylogenetic tree showed that CBX2 was evolutionarily conserved from fish to humans. The CBX2 proteins of medaka and all other fish were grouped together, whereas those of amphibians, reptiles, birds, and mammals were grouped together (Figure 3).
2.2. Tissue Expression of cbx2 mRNA
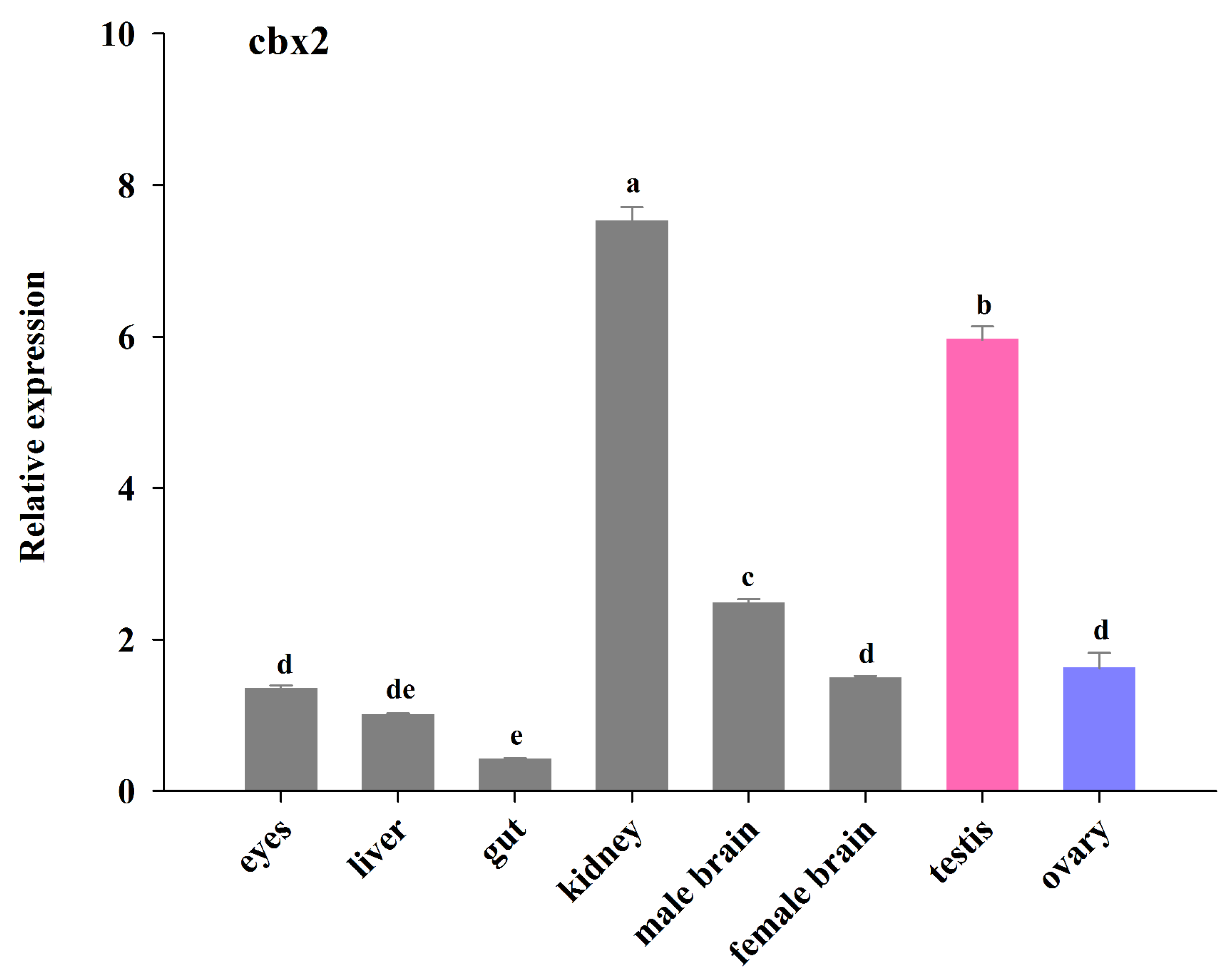
The relative expression of cbx2 mRNA in different tissues was evaluated using real-time PCR (Figure 4). The cbx2 mRNA was detected in different tissues of the adult medaka. The expression of cbx2 mRNA was higher in the kidneys and testes than in other tissues. Furthermore, the expression of cbx2 mRNA in the testes was about three times that in the ovaries, and its level in male brains was relatively higher than in female brains (P < 0.05).
2.3. Gonadal Localization of cbx2 mRNA by ISH and FISH
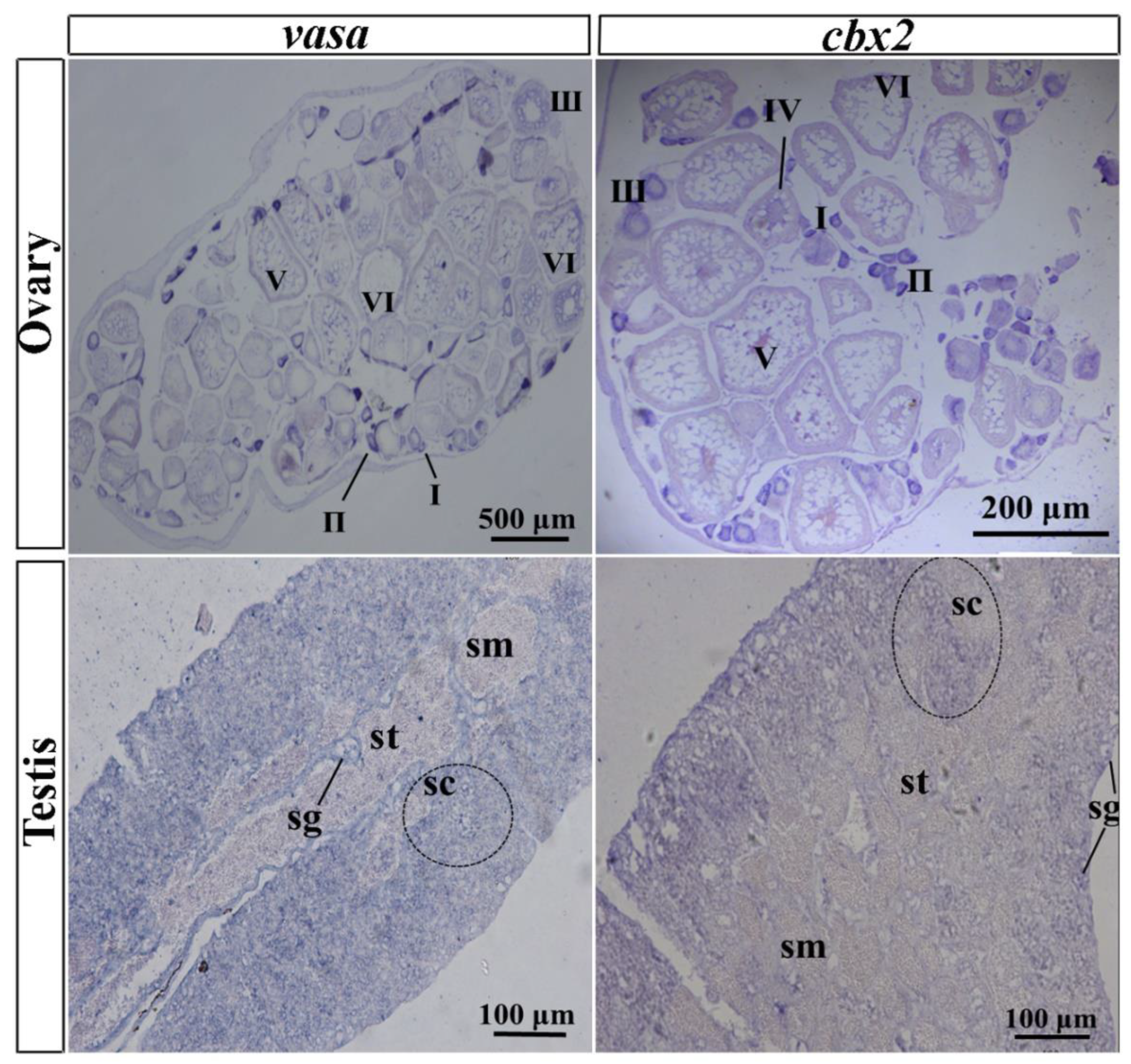
In the ovary, oocyte development is not synchronized because adult female medaka produce eggs daily. A small number of oocytes exist at stage I and a large number of oocytes exist in the ovary from stage II to stage V. In this study, vasa was employed as a marker gene, with its expression pattern first being verified by DIG-labeled ISH. The results showed that vasa mRNA was expressed throughout the whole oogenesis process, with higher expression in stages I, II, and III, whereas it was expressed at a lesser extent in stages IV and V. Similarly, cbx2 mRNA was highly expressed in oocytes at stages I, II, and III, and expressed less in stage IV and V oocytes (Figure 5). No positive signals were sensed by ISH in the probe.
Medaka testes are small in size, and the reproductive epithelium divides them into many irregular efferent cysts containing a variety of germ cells during gametogenesis (Figure 5). From the outside in, a testis is divided into spermatogonia, spermatocytes, spermatids, and sperm. The ISH results showed that cbx2 mRNA was chiefly located in spermatocytes and spermatogonia, and it was hardly distributed at all in mature sperms and spermatids (Figure 5). Similarly, vasa mRNA was localized predominately in the spermatogonia and spermatocytes, with no obvious signal in sperm.
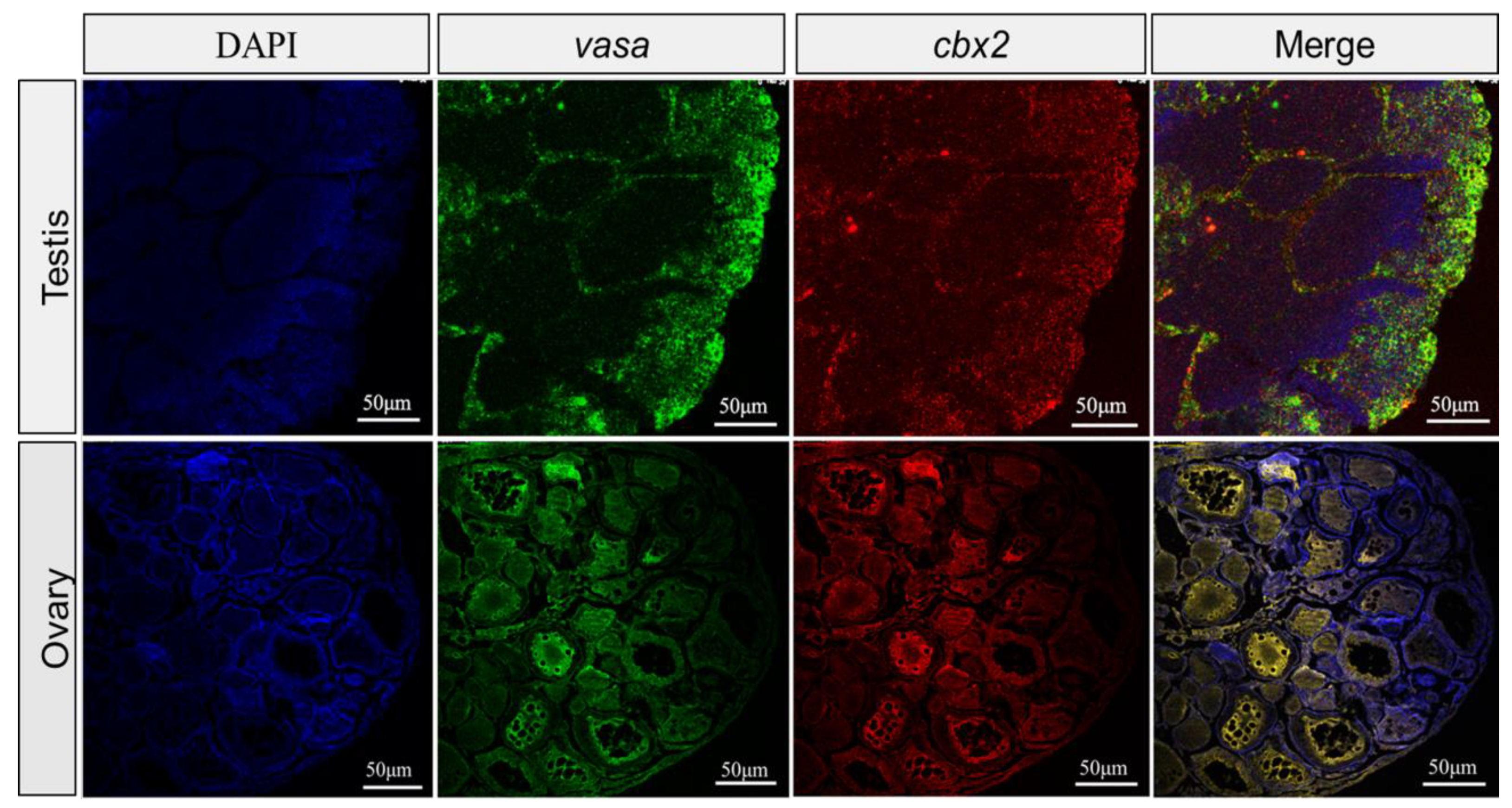
To further localize the cbx2 mRNA, we utilized vasa as a reference to perform FISH. The results showed that the cellular localization of cbx2 and vasa mRNA almost completely overlapped in the ovary and the testis (Figure 6), which was consistent with the ISH results.
2.4. Expression Changes of cbx2, sox9, and foxl2 mRNA after RNA Interference
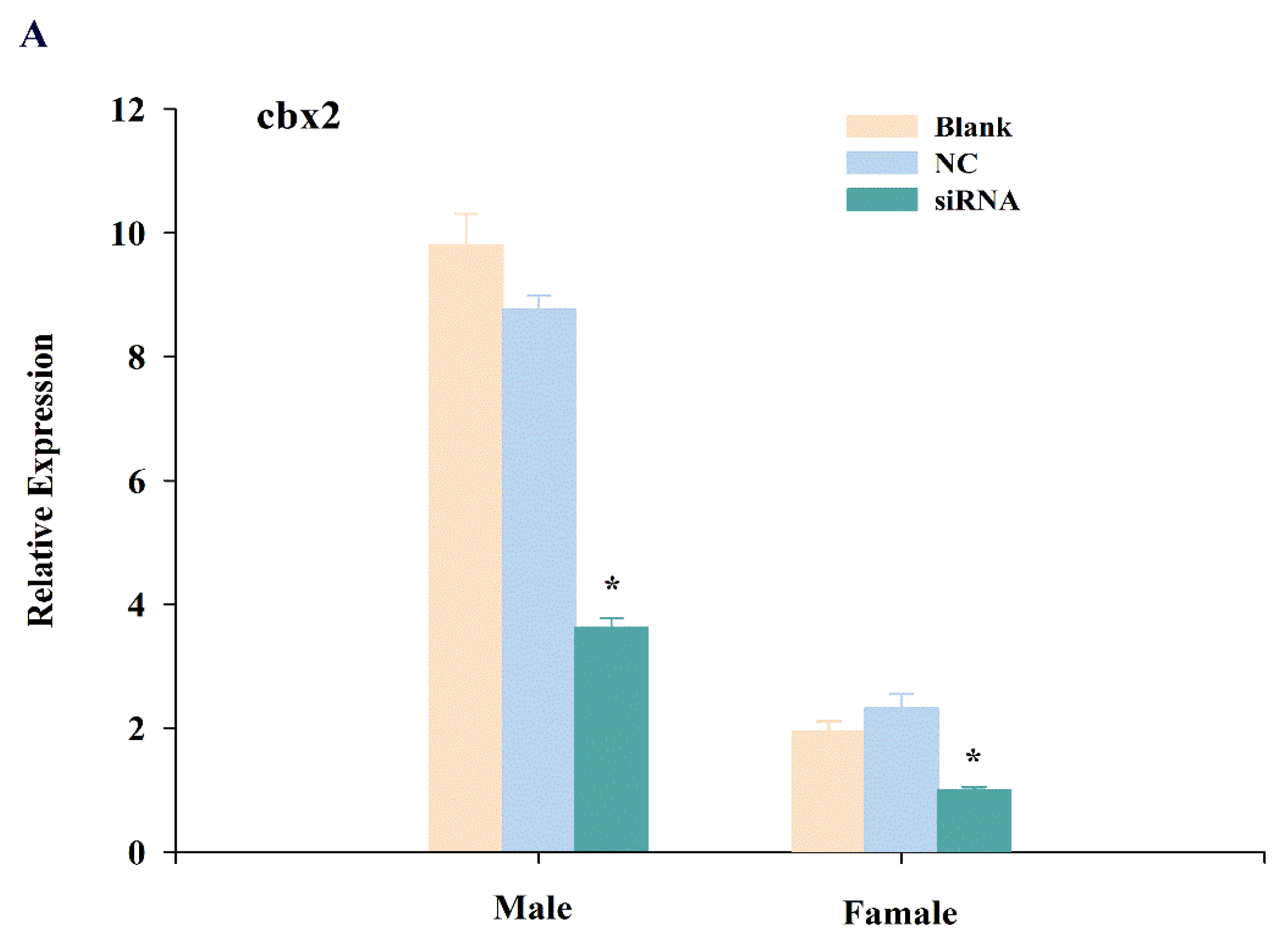
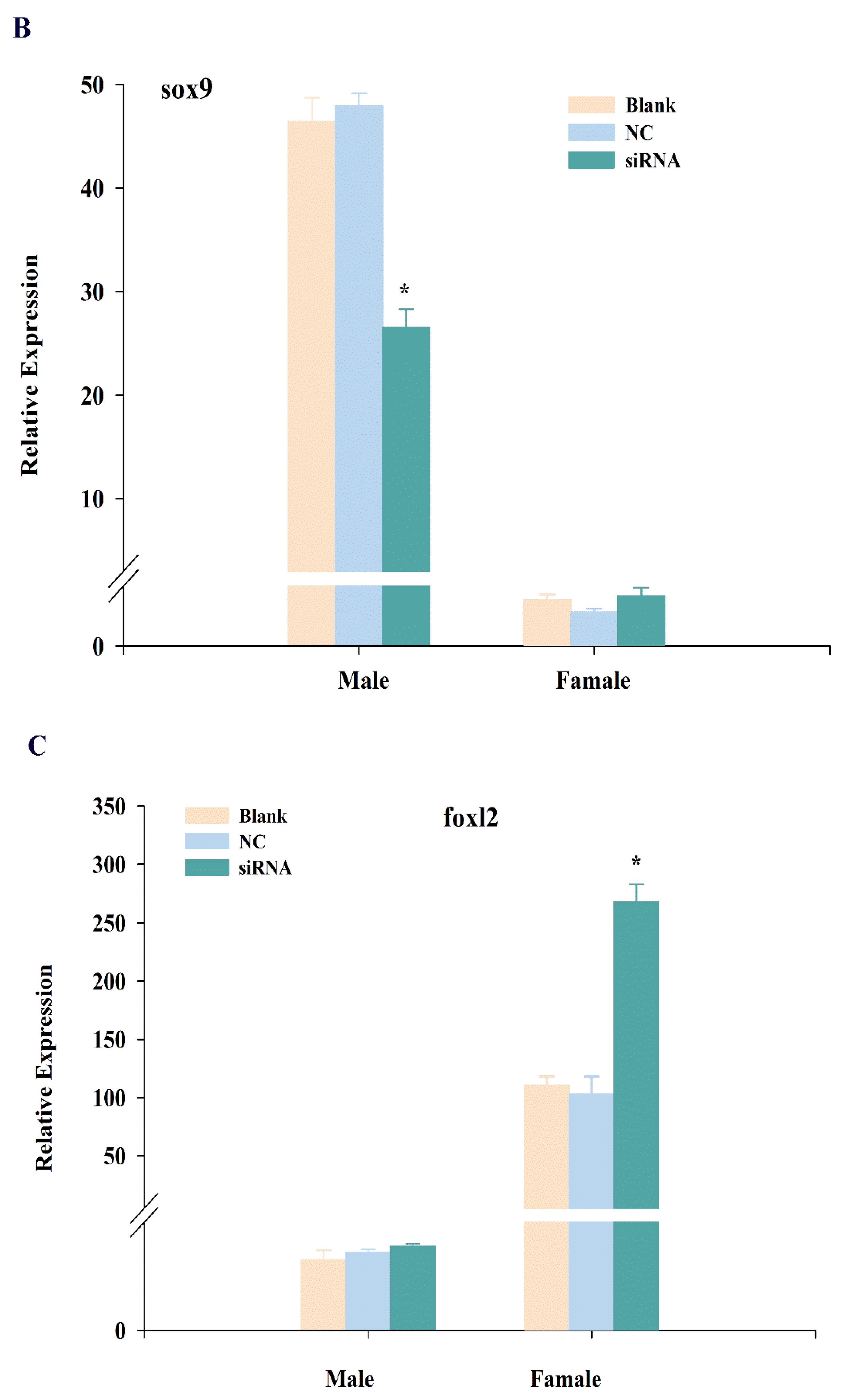
To further explore the function of cbx2, we knocked down cbx2 and investigated the expression changes of sex-related genes (i.e., sox9 and foxl2). The results showed that the expression of cbx2 in the testis was distinctly inhibited after injecting siRNA (P < 0.05), and its expression was also significantly decreased in the ovary (Figure 7A). Furthermore, the level of sox9, a male sex-related gene, was decreased in the testis when cbx2 expression was inhibited (Figure 7B). However, the level of foxl2, a female sex-related gene, was increased in the ovary (Figure 7C).
3. Discussion
In this study, cbx2 was identified in medaka and shown to be conserved in vertebrate evolution. The CBX2 protein had the highest ratio of serine. Kawaguchi et al. found that the serine-rich region is the phosphorylation site of CBX2 and is stably phosphorylated in cells [17]. In fish-to-human evolution, the length of the serine-rich region decreased from 24 serine residues to 16 serine residues [18]. Sequence alignment suggested that the anterior-Chromo domain and the posterior-Pfam domain possessed high sequence similarity. H3K27me3 is a typical epigenetic gene silencing marker, often located in the promoter region of developmental genes. The Chromo domain recognizes H3K27me3 and recruits PRC1 to the target gene to regulate gene expression. The Pfam domain is a conserved region at the C-terminus of the CBX protein family and is involved not only in transcriptional silence but also often binds the RING1B domain, causing the protein complexes to participate in the alternative recruitment pathway of chromatin targeting [19]. These results suggest that the CBX2 protein plays an important role in epigenetic regulation.
This work showed that cbx2 was expressed relatively abundantly in medaka gonads and kidneys. In humans, RNA sequencing data analysis revealed that cbx2 was more abundant in testes, followed by ovaries, placenta, and then lungs, but was expresses to a lesser extent in other tissues [20]. In mice, sequencing results showed that cbx2 was expressed mainly in the brain and limbs, with greater expression also gonads but less in other tissues. Moreover, our real-time PCR results indicated that the expression of cbx2 in testes was higher than in ovaries, suggesting that cbx2 may perform a vital function regarding gonadal development. In addition, cbx2 is also involved in balancing the self-renewal and differentiation of haematopoietic stem cells [21], lymphopoiesis [22], and the antiviral immunity response [23]. The high expression of cbx2 in medaka kidneys indicates that cbx2 is associated with the immune system.
Furthermore, our results from ISH and FISH showed that the expression profile of cbx2 was similar to that of vasa in medaka gonads. Vasa is an evolutionarily conserved translation factor and is required for the polarization of the anterior–posterior axis and the formation of germ cells. So far, the expression of vasa mRNA has only been found in medaka germ cells [24]. Similar to vasa, cbx2 signals were mostly localized in spermatocytes and spermatogonia and not so much in mature sperms and spermatids in medaka testes. In medaka ovaries, cbx2 was also found in primary oocytes at stages I, II, and III. Previous studies in mammals showed that cbx2 plays a key role in cell cycle changes [25,26], meiosis, homologous chromosome synapsis, and germ cell differentiation [27]. These results suggest that cbx2 might be involved in spermatogenesis and oogenesis in medaka.
After cbx2 knockdown in medaka, sox9 expression was downregulated in testes. SRY-related HMG box 9 (sox9) is related to spermatogenesis and testis maintenance and development in mammals. Johnsen et al. found that sox9 negatively regulated the expression of aromatase to inhibit estrogen synthesis [28], with aromatase being the rate-limiting enzyme in estrogen synthesis [29]. Therefore, the high expression of sox9 may be one of the key factors in gonad masculinization. In teleost, sox9 is also significantly expressed during gonadal development and affects the differentiation process of both germ cells and supporting cells [30]. As a result, we speculated that cbx2 was possibly involved in male gonadal development by regulating sox9 expression in medaka.
On the other hand, the expression of sex-related foxl2 was upregulated in ovaries when cbx2 expression was inhibited in medaka. Forkhead transcriptional factor 2 (foxl2) is important in ovarian development and a key factor in female sex determination, and was also found to be abundantly expressed in Gobiocypris rarus [31] and Clarias gariepinus [32] ovaries. Foxl2 was shown to affect gonadal differentiation and development by regulating cyp19a1, a key gene in the female signaling pathway in Oreochromis niloticus [33]. These results indicate that cbx2 is also related to female gonadal development by controlling the gene expression of foxl2 in medaka.
In mice, M33 (cbx2) deficiency may cause sex reversal, i.e., male-to-female [34]. In humans, the expression levels of Nr5a1, sox9, and sry decrease significantly when cbx2 is knocked down using siRNA, and target identification by genome-wide sequencing suggested that cbx2 influences gonadal development by promoting the male signaling pathway while simultaneously inhibiting the female signaling pathway [35]. Therefore, the higher expression of cbx2 in gonads implied that this gene plays a role in spermatogenesis and oogenesis, and the gene knockdown results demonstrated its involvement in gonadal differentiation by controlling the sex-related genes, such as sox9 and foxl2, in medaka.
4. Materials and Methods
4.1. Experimental Materials
Medaka were raised at the Experimental Center of Shanghai Ocean University. Adult fishes at 3 months of age (n = 3 pools, 10 specimens/pool) were collected. The female brains, male brains, ovaries, testes, livers, kidneys, guts, and eyes were dissected, swiftly frozen, and stored at −80 °C in a refrigerator for RNA extraction. In addition, the ovaries and testes were cut and fixed in 4% paraformaldehyde overnight, then stored into 70% ethanol to make sections for ISH and FISH. Animal experiments were carried out in strict accordance with the guidance of the committee for the Laboratory Animal Research of Shanghai Ocean University.
4.2. Molecular Cloning and Sequence Analysis
Total RNA of different tissues was extracted using TRizol (Invitrogen, Carlsbad, CA, USA). To eliminate genome contamination, cDNA was reversed transcribed using the PrimeScript™ RT reagent Kit with the gDNA Eraser (Takara Biotechnology, Dalian, China). By searching the NCBI database, we obtained the predicted CDS sequences encoding the medaka cbx2 gene (Genebank accession No. EF537027.1). The CDS region of cbx2 was verified by PCR cloning and sequencing. Then, the physicochemical properties of the predicted proteins were analyzed online using the ExPASy-ProtParam tool, and the secondary and tertiary structures were predicted using SMART and Swiss-model. Protein sequence alignment was executed by BioEdit, and the neighbor-joining (NJ) tree was established by MEGA 7.0 based on CBX2 protein sequences from various species according to BLASTP.
4.3. Real-Time PCR of cbx2 in Different Tissues
Real-time PCR was performed with specific primers of cbx2 and 18S, as a reference gene, following the same set of cDNA amplification. The reaction underwent 39 cycles of 10 s at 95 °C and 30 s at 60 °C in a 20 μL system using CFX96 Touch real-time (Bio-Rad, Hercules, CA, USA). The PCR primers are described in Table 1.
The relative expression for cbx2 mRNA was evaluated using the 2-ΔΔCt method and the results were presented according to the mean ± standard deviation (SD). Statistical analysis was performed with one-way of variance (ANOVA) using SPSS 24 software. Differences were considered to be statistically significant at P < 0.05.
4.4. Gonadal Expression of cbx2 by ISH and FISH
In order to understand the subcellular distributions of cbx2, ISH and FISH were performed on gonadal sections and verified by hematoxylin and eosin staining. ISH was performed as previously described [24]. Briefly, the 552 nt of cbx2 cDNA and 794 nt of vasa cDNA were inserted into a pGEM-T vector, respectively. The recombinant plasmids were linearized with spel or sphl to transcribe DIG-labeled or FITC-labeled antisense or sense probes according to RNA Labeling kit instructions (Roche, Basel, Switzerland). The synthetic probe was incubated with DNaseI (Thermo Fisher, Carlsbad, CA, USA) and precipitated by LiCl at –80 °C. The ISH experiments of cbx2 and vasa as the marker gene were performed and stained with BCIP/NBT. FISH was carried out with a TSA™ Plus Fluorescence Systems kit according to the instructions (NEL756, PerkinElmer, Waltham, MA, USA). The nucleus was dyed with DAPI and the slide was fixed with Anti-Fade Mounting Medium (Invitrogen, Carlsbad, CA, USA).
Photography and observation of ISH were performed using an upright microscope (Leica DM500, Heerbrugg, Switzerland) and FISH images were observed and photographed using a Leica SP8 FALCON with the confocal image system (LAS X).
4.5. RNA Interference
siRNA sequences were designed and synthesized by Shanghai GeneBio Co. Ltd (Table 2). According to the instructions, cbx2-siRNA-1, cbx2-siRNA-2, and cbx2-siRNA-3 were mixed and diluted with Rnase-free water. For RNA interference, a pre-experiment was performed to optimize the injection concentration of the siRNA mixture (25 ng, 50 ng, 75 ng, or 100 ng). The optimal 50 ng siRNA mixture was then injected from the cloacal aperture of the medaka using a medical syringe with a diameter of 0.38 µm. A negative control of siRNA was synthesized, injected, and a blank group was set up synchronously. A total of 10 adult fishes at 3 months of age were injected in each assay. Each assay was repeated in triplicate.
After 48 h, the injected fishes were collected and their gonadal tissues were dissected for RNA extraction. Real-time PCR was performed to detect the expression changes of cbx2, sox9, and foxl2 mRNA after RNA interference. The experimental methods and procedures were consistent with those described above.
Author Contributions
Q.C. completed the experiments; F.S. and Y.X. helped with the experiment; J.W. was responsible for data analysis and writing of the manuscript; J.Z. was responsible for experimental design and writing of the manuscript. All authors read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (No. 31972772) and the Laboratory for Marine Fisheries Science and Food Production Processes, Qingdao National Laboratory for Marine Science and Technology, China (ZZ-A11).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lewis, E.B. A gene complex controlling segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Simon, J.A.; Kingston, R.E. Mechanisms of polycomb gene silencing: Knowns and unknowns. Nat. Rev. Mol. Cell Biol. 2009, 10, 697–708. [Google Scholar] [CrossRef]
- King, H.W.; Fursova, N.A.; Blackledge, N.P.; Klose, R.J. Polycomb repressive complex 1 shapes the nucleosome landscape but not accessibility at target genes. Genome Res. 2018, 28, 1494–1507. [Google Scholar] [CrossRef] [Green Version]
- Boyer, L.A.; Plath, K.; Zeitlinger, J.; Brambrink, T.; Medeiros, L.A.; Lee, T.I.; Levine, S.S.; Wernig, M.; Tajonar, A.; Ray, M.K.; et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature 2006, 441, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Kerppola, T.K. Polycomb group complexes-many combinations, many functions. Trends Cell Biol. 2009, 19, 692–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Croce, L.; Helin, K. Transcriptional regulation by Polycomb group proteins. Nat. Struct. Mol. Biol. 2013, 20, 1147–1155. [Google Scholar] [CrossRef]
- Katoh-Fukui, Y.; Miyabayashi, K.; Komatsu, T.; Owaki, A.; Baba, T.; Shima, Y.; Kidokoro, T.; Kanai, Y.; Schedl, A.; Wilhelm, D.; et al. Cbx2, a polycomb group gene, is required for Sry gene expression in mice. Endocrinology 2012, 153, 913–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plys, A.J.; Davis, C.P.; Kim, J.; Rizki, G.; Keenen, M.M.; Marr, S.K.; Kingston, R.E. Phase separation of polycomb-repressive complex 1 is governed by a charged disordered region of CBX2. Genes Dev. 2019, 33, 799–813. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.G.; Zhang, Y.; Sun, T.T.; Cheng, B. Epigenetic regulation by polycomb group complexes: Focus on roles of CBX proteins. J. Zhejiang Univ. Sci. B 2014, 15, 412–428. [Google Scholar] [CrossRef] [Green Version]
- Biason-Lauber, A.; Konrad, D.; Meyer, M.; DeBeaufort, C.; Schoenle, E.J. Ovaries and female phenotype in a girl with 46, XY karyotype and mutations in the CBX2 gene. Am. J. Hum. Genet. 2009, 84, 658–663. [Google Scholar] [CrossRef] [Green Version]
- Zhen, C.Y.; Duc, H.N.; Kokotovic, M.; Phiel, C.J.; Ren, X. Cbx2 stably associates with mitotic chromosomes via a PRC2-or PRC1-independent mechanism and is needed for recruiting PRC1 complex to mitotic chromosomes. Mol. Biol. Cell 2014, 25, 3726–3739. [Google Scholar] [CrossRef] [PubMed]
- Katoh-Fukui, Y.; Owaki, A.; Toyama, Y.; Kusaka, M.; Shinohara, Y.; Maekawa, M.; Toshimori, K.; Morohashi, K.I. Mouse polycomb M33 is required for splenic vascular and adrenal gland formation through regulating Ad4BP/SF1 expression. Blood 2005, 106, 1612–1620. [Google Scholar] [CrossRef] [PubMed]
- Sproll, P.; Eid, W.; Biason-Lauber, A. CBX2-dependent transcriptional landscape: Implications for human sex development and its defects. Sci. Rep. 2019, 9, 16552. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, H.; Saito, D.; Nakamura, S.; Katoh-Fukui, Y.; Ohta, K.; Baba, T.; Morohashi, K.; Tanaka, M. Germ cells are essential for sexual dimorphism in the medaka gonad. Proc. Natl. Acad. Sci. USA 2007, 104, 16958–16963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanda, I.; Kondo, M.; Hornung, U.; Asakawa, S.; Winkler, C.; Shimizu, A.; Shan, Z.; Haaf, T.; Shimizu, N.; Shima, A.; et al. A duplicated copy of Dmrt1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc. Natl. Acad. Sci. USA 2002, 99, 11778–11783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N.; et al. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 2002, 417, 559–563. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Machida, S.; Kurumizaka, H.; Tagami, H.; Nakayama, J.I. Phosphorylation of CBX2 controls its nucleosome-binding specificity. J. Biochem. 2017, 162, 343–355. [Google Scholar] [CrossRef]
- Senthilkumar, R.; Mishra, R.K. Novel motifs distinguish multiple homologues of polycombin vertebrates: Expansion and diversification of the epigenetic toolkit. BMC Genom. 2009, 10, 549–566. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Taylor, A.B.; Leal, B.Z.; Chadwell, L.V.; Ilangovan, U.; Robinson, A.K.; Schirf, V.; Hart, P.J.; Lafer, E.M.; Demeler, B.; et al. Polycomb group targeting through different binding partners of RING1B C-terminal domain. Structure 2010, 18, 966–975. [Google Scholar] [CrossRef] [Green Version]
- Fagerberg, L.; Hallström, B.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Klauke, K.; Radulović, V.; Broekhuis, M.; Weersing, E.; Zwart, E.; Olthof, S.; Bystrykh, L. Polycomb Cbx family members mediate the balance between haematopoietic stem cell self-renewal and differentiation. Nat. Cell Biol. 2013, 15, 353–374. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Koseki, H.; Vidal, M.; Nakauchi, H.; Iwama, A. Unique composition of polycomb repressive complex 1 in hematopoietic stem cells. Int. J. Hematol. 2007, 85, 179–181. [Google Scholar] [CrossRef]
- Sun, D.H.; Cao, X.T.; Wang, C.M. Polycomb chromobox Cbx2 enhances antiviral innate immunity by promoting Jmjd3-mediated demethylation of H3K27 at the Ifnb promoter. Protein Cell 2019, 10, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.Y.; Shen, Q.; Xu, H.Y.; Wong, F.M.; Cui, J.Z.; Li, Z.D.; Hong, N.; Wang, L.; Zhao, H.B.; Ma, B.; et al. Differential conservation and divergence of fertility genes Boule and Dazl in the rainbow trout. PLoS ONE 2011, 6, e15910. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, K.; Shiurba, R.; Higashinakagawa, T. Nuclear translocation of mouse polycomb M33 protein in regenerating liver. Biochem. Biophys. Res. Commun. 2002, 291, 508–515. [Google Scholar] [CrossRef]
- Hirose, S.; Komoike, Y.; Higashinakagawa, T. Identification of a nuclear localization signal in mouse polycomb protein, M33. Zool. Sci. 2006, 23, 785–791. [Google Scholar] [CrossRef]
- Baumann, C.; Rabindranath, D.L.F. Role of polycomb group protein Cbx2/M33 in meiosis onset and maintenance of chromosome stability in the mammalian germline. Genes 2011, 2, 59–80. [Google Scholar] [CrossRef] [Green Version]
- Johnsen, H.; Tveiten, H.; Torgersen, J.S.; Andersen, O. Divergent and sex-dimorphic expression of the paralogs of the Sox9-Amh-Cyp19a1 regulatory cascade in developing and adult-atlantic cod (Gadus morhua L.). Mol. Reprod. Dev. 2013, 80, 358–370. [Google Scholar] [CrossRef]
- Wang, X.Y.; Simpson, E.R.; Brown, K.A. Aromatase overexpression in dysfunctional adipose tissue links obesity to postmenopausal breast cancer. J. Steroid Biochem. Mol. Biol. 2015, 153, 35–44. [Google Scholar] [CrossRef]
- Chiang, E.F.; Pai, C.I.; Wyatt, M.; Yan, Y.L.; Postlethwait, J.; Chung, B. Two Sox9 genes on duplicated zebrafish chromosomes:expression of similar transcription activators in distinct sites. Dev. Biol. 2001, 231, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.P.; Wu, T.T.; Qin, F.; Wang, L.H.; Wang, Z.Z. Molecular cloning of Foxl2 gene and the effects of endocrine-disrupting chemicals on its mRNA level in rare minnow, Gobiocypris rarus. Fish Physiol. Biochem. 2012, 38, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Sridevi, P.; Senthilkumaran, B. Cloning and differential expression of Foxl2 during ovarian development and recrudescence of the catfish, Clarias gariepinus. Gen. Comp. Endocrinol. 2011, 174, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.S.; Kobayashi, T.; Zhou, L.Y.; Paul-Prasanth, B.; Ijiri, S.; Sakai, F.; Okubo, K.; Morohashi, K.; Nagahama, Y. Foxl2 up-regulates aromatase gene transcription in a female-specific manner by binding to the promoter as well as interacting with Ad4 binding protein/steroidogenic factor 1. Mol. Endocrinol. 2007, 21, 712–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh-Fukui, Y.; Tsuchiya, R.; Shiroishi, T.; Nakahara, Y.; Hashimoto, N.; Noguchi, K.; Higashinakagawa, T. Male-to-female sex reversal in M33 mutant mice. Nature 1998, 393, 688–692. [Google Scholar] [CrossRef]
- Eid, W.; Opitz, L.; Biason-Lauber, A. Genome-wide identification of CBX2 targets: Insights in the human sex development network. Mol. Endocrinol. 2015, 29, 247–257. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The cDNA sequences and the derived protein sequence of medaka cbx2 (A). The red font position represents the cloning primers, the Chromo domain is marked by a green box, the Pfam domain is marked by a blue box, and the underlined sequence indicates the serine-rich region. The secondary structure of CBX2 in medaka (B).
Figure 1.
The cDNA sequences and the derived protein sequence of medaka cbx2 (A). The red font position represents the cloning primers, the Chromo domain is marked by a green box, the Pfam domain is marked by a blue box, and the underlined sequence indicates the serine-rich region. The secondary structure of CBX2 in medaka (B).

Figure 2.
Sequence alignment of the CBX2 protein between different species. Abbreviations: Ol, Oryzias latipes; On, Oreochromis niloticus; Po, Paralichthys olivaceus; Dr, Danio rerio; Xl, Xenopus laevis; Tct, Terrapene carolina triunguis; Pc, Phasianus colchicus; Mm, Mus musculus; Hs, Homo sapiens.
Figure 2.
Sequence alignment of the CBX2 protein between different species. Abbreviations: Ol, Oryzias latipes; On, Oreochromis niloticus; Po, Paralichthys olivaceus; Dr, Danio rerio; Xl, Xenopus laevis; Tct, Terrapene carolina triunguis; Pc, Phasianus colchicus; Mm, Mus musculus; Hs, Homo sapiens.

Figure 3.
Phylogenetic tree of CBX2 in vertebrates. Bootstrap values are marked at the branch points.
Figure 3.
Phylogenetic tree of CBX2 in vertebrates. Bootstrap values are marked at the branch points.

Figure 4.
The relative expression of cbx2 mRNA in different medaka tissues. The different alphabetic signs indicate statistically significant differences (P < 0.05).
Figure 4.
The relative expression of cbx2 mRNA in different medaka tissues. The different alphabetic signs indicate statistically significant differences (P < 0.05).

Figure 5.
Digoxigenin (DIG)-labeled in situ hybridization (ISH) in medaka ovaries and testes. Paraffin sections were hybridized with the antisense probe of cbx2 and vasa. Signals were visualized using chromogenic staining. I–VI, oocyte stage; sg, spermatogonia; sc, spermatocytes; st, spermatid; sm, sperm.
Figure 5.
Digoxigenin (DIG)-labeled in situ hybridization (ISH) in medaka ovaries and testes. Paraffin sections were hybridized with the antisense probe of cbx2 and vasa. Signals were visualized using chromogenic staining. I–VI, oocyte stage; sg, spermatogonia; sc, spermatocytes; st, spermatid; sm, sperm.

Figure 6.
Fluorescent in situ hybridization (FISH) in medaka ovaries and testes. The mRNA signals were stained with fluorescence, with green indicating vasa and red indicating cbx2, and the nuclei were dyed with DAPI (blue).
Figure 6.
Fluorescent in situ hybridization (FISH) in medaka ovaries and testes. The mRNA signals were stained with fluorescence, with green indicating vasa and red indicating cbx2, and the nuclei were dyed with DAPI (blue).

Figure 7.
Expression changes of cbx2 (A), sox9 (B), and foxl2 (C) mRNA in gonads after RNA interference. The asterisk represents a statistically significant difference (P < 0.05).
Figure 7.
Expression changes of cbx2 (A), sox9 (B), and foxl2 (C) mRNA in gonads after RNA interference. The asterisk represents a statistically significant difference (P < 0.05).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers for real-time PCR and probe synthesis.
| Primer | Primer Sequence (5′→3′) |
|---|---|
| cbx2-F | GCCCAAGTCCAGCACCTCA |
| cbx2-R | GCTCCTTCGCCTCACCTCT |
| cbx2-spel-F | GGACTAGTTGGTGGGCGACTCCTTGA |
| cbx2-sphl-R | CATGCATGCACCTGAACCCTTCCAAACAC |
| vasa-spel-F | GGACTAGTCCCGCTTGTTGAATTTGG |
| vasa-sphl-R | CATGCATGCTGTGCGAGTCGTTGGAGAA |
| sox9-F | AAACTGGCCGACCAATAC |
| sox9-R | CTCAGCCTCCTCCACAAA |
| foxl2-F | TCCTACACGTCCTGCCAGAT |
| foxl2-R | CCCATGCCGTTGTAAGAGTT |
| 18s-F | CTGAGAAACGGCTACCACAG |
| 18s-R | CAGCAACTTTAAGATACGC |
Codes were used to degenerate the primers. F, sense primer; R, antisense primer.
Table 2.
siRNA target site and sequence.
| Gene | Direction | Sequence |
|---|---|---|
| cbx2-siRNA-1(251) | Sense (5′-3′) | CCGACUCUGACCGCACUAATT |
| Anti-sense (3′-5′) | UUAGUGCGGUCAGAGUCGGTT | |
| cbx2-siRNA-2(853) | Sense (5′-3′) | GCGCUGCACCUGAACCCUUTT |
| Anti-sense (3′-5′) | AAGGGUUCAGGUGCAGCGCTT | |
| cbx2-siRNA-3(1337) | Sense (5′-3′) | GCCUCAUCGAGCACGUGUUTT |
| Anti-sense (3′-5′) | AACACGUGCUCGAUGAGGCTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chao, Q.; Shen, F.; Xue, Y.; Wu, J.; Zhang, J. Cbx2, a PcG Family Gene, Plays a Regulatory Role in Medaka Gonadal Development. Int. J. Mol. Sci. 2020, 21, 1288. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041288
AMA Style
Chao Q, Shen F, Xue Y, Wu J, Zhang J. Cbx2, a PcG Family Gene, Plays a Regulatory Role in Medaka Gonadal Development. International Journal of Molecular Sciences. 2020; 21(4):1288. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041288
Chicago/Turabian StyleChao, Qinghe, Fengfeng Shen, Yidong Xue, Jikui Wu, and Junling Zhang. 2020. "Cbx2, a PcG Family Gene, Plays a Regulatory Role in Medaka Gonadal Development" International Journal of Molecular Sciences 21, no. 4: 1288. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041288
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.