The Transcriptome of Pig Spermatozoa, and Its Role in Fertility

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Experiment 1: Screening of Sperm miRNAs in the Sperm-rich Fraction (SRF) of the Boar Ejaculate
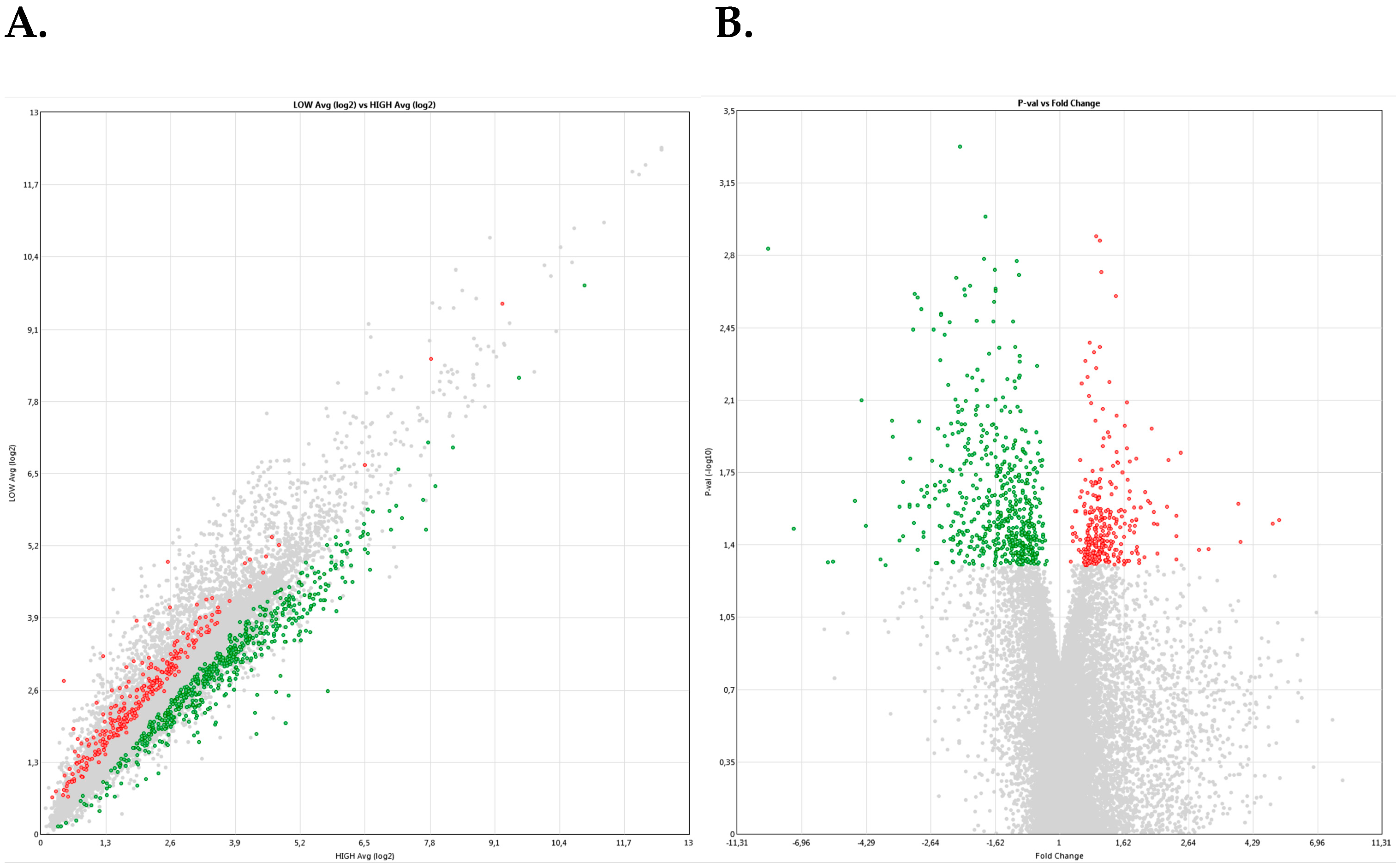
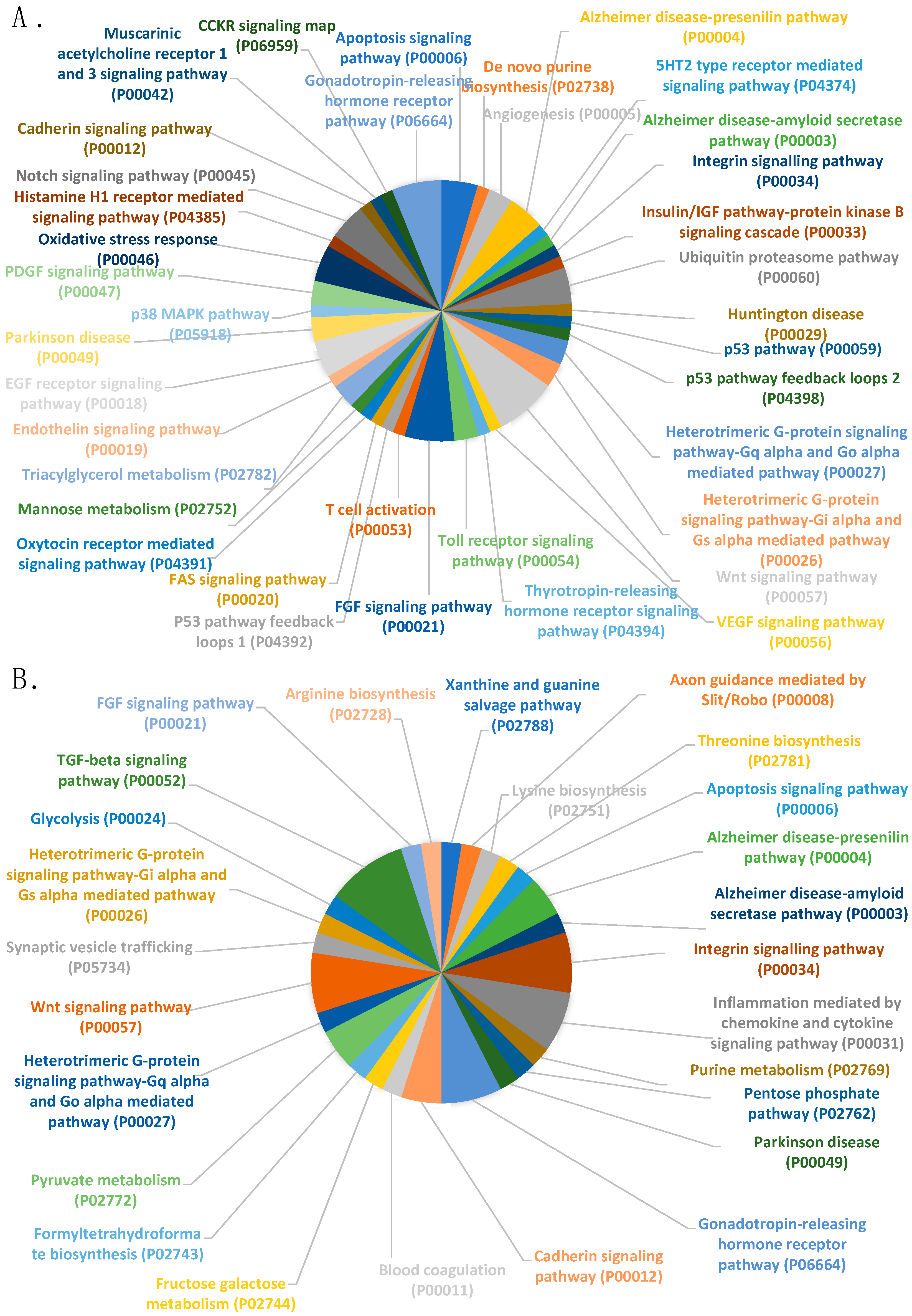
2.2. Experiment 2. Differential Cargo of RNAs in Ejaculated Spermatozoa from Breeding Boars with High- or Low-fertility (as Farrowing Rate and Litter Size) after Artificial Insemination (AI)
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Ejaculated Spermatozoa
4.3. Experimental Design
4.3.1. Experiment 1. Screening of Sperm miRNAs in the Sperm-rich Fraction (SRF) of the Boar Ejaculate
4.3.2. Experiment 2: Assessment of the Cargo of RNAs in Ejaculated Spermatozoa from Breeding Boars with High- or Low-fertility (as Farrowing Rate and Litter Size) after AI.
4.4. RNA Isolation
4.5. MiRNA Microarray Protocol (Experiment 1)
4.6. Microarray Protocol (Experiment 2)
4.7. miRNA Arrays and Microarrays Hybridization and Scanning
4.8. Bioinformatics and Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baker, R.D.; Degen, A.A. Transport of live and dead boar spermatozoa within the reproductive tract of gilts. J. Reprod. Fertil. 1972, 28, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Langendijk, P.; Soede, N.M.; Kemp, B. Uterine activity, sperm transport, and the role of boar stimuli around insemination in sows. Theriogenology 2005, 63, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Alminana, C.; Caballero, I.; Heath, P.R.; Maleki-Dizaji, S.; Parrilla, I.; Cuello, C.; Gil, M.A.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; et al. The battle of the sexes starts in the oviduct: Modulation of oviductal transcriptome by X and Y-bearing spermatozoa. BMC Genom. 2014, 15, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Rodriguez, M.; Atikuzzaman, M.; Venhoranta, H.; Wright, D.; Rodriguez-Martinez, H. Expression of Immune Regulatory Genes in the Porcine Internal Genital Tract Is Differentially Triggered by Spermatozoa and Seminal Plasma. Int. J. Mol. Sci. 2019, 20, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Martinez, H.; Kvist, U.; Saravia, F.; Wallgren, M.; Johannisson, A.; Sanz, L.; Pena, F.J.; Martinez, E.A.; Roca, J.; Vazquez, J.M.; et al. The physiological roles of the boar ejaculate. Soc. Reprod. Fertil. Suppl. 2009, 66, 1–21. [Google Scholar] [PubMed]
- Robertson, S.A.; Sharkey, D.J. The role of semen in induction of maternal immune tolerance to pregnancy. Semin. Immunol. 2001, 13, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martinez, H.; Courtens, J.L.; Kvist, U.; Ploen, L. Immunocytochemical localization of nuclear protamine in boar spermatozoa during epididymal transit. J. Reprod. Fertil. 1990, 89, 591–595. [Google Scholar] [CrossRef] [Green Version]
- Grunewald, S.; Paasch, U.; Glander, H.J.; Anderegg, U. Mature human spermatozoa do not transcribe novel RNA. Andrologia 2005, 37, 69–71. [Google Scholar] [CrossRef]
- Yang, C.C.; Lin, Y.S.; Hsu, C.C.; Wu, S.C.; Lin, E.C.; Cheng, W.T.K. Identification and sequencing of remnant messenger RNAs found in domestic swine (Sus scrofa) fresh ejaculated spermatozoa. Anim. Reprod. Sci. 2009, 113, 143–155. [Google Scholar] [CrossRef]
- Dai, D.-H.; Qazi, I.H.; Ran, M.-X.; Liang, K.; Zhang, Y.; Zhang, M.; Zhou, G.-B.; Angel, C.; Zeng, C.-J. Exploration of miRNA and mRNA Profiles in Fresh and Frozen-Thawed Boar Sperm by Transcriptome and Small RNA Sequencing. Int. J. Mol. Sci. 2019, 20, 802. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C. Novel functions for small RNA molecules. Curr. Opin. Mol. Ther. 2009, 11, 641–651. [Google Scholar] [PubMed]
- Miller, D. Analysis and Significance of Messenger RNA in Human Ejaculated Spermatozoa. Mol. Reprod. Dev. 2000, 56, 259–264. [Google Scholar] [CrossRef]
- Brunet, A.; Sweeney, L.B.; Sturgill, J.F.; Chua, K.F.; Greer, P.L.; Lin, Y.; Tran, H.; Ross, S.E.; Mostoslavsky, R.; Cohen, H.Y.; et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science 2004, 303, 2011–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Liu, Y.; Chen, L.; Ellis, M.; Li, M.; Wang, J.; Zhang, Y.; Fu, P.; Wang, K.; Li, X.; et al. microRNA profiling in three main stages during porcine spermatogenesis. J. Assist. Reprod. Genet. 2015, 32, 451–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, L.; Ye, L.; Liu, G.; Shao, G.; Zheng, R.; Ren, Z.; Zuo, B.; Xu, D.; Lei, M.; Jiang, S.; et al. Microarray-Based approach identifies differentially expressed MicroRNAs in porcine sexually immature and mature testes. PLoS ONE 2010, 5, e11744. [Google Scholar] [CrossRef] [PubMed]
- Curry, E.; Safranski, T.J.; Pratt, S.L. Differential expression of porcine sperm microRNAs and their association with sperm morphology and motility. Theriogenology 2011, 76, 1532–1539. [Google Scholar] [CrossRef]
- Tian, H.; Li, Z.; Peng, D.; Bai, X.; Liang, W. Expression difference of miR-10b and miR-135b between the fertile and infertile semen samples (p). Forensic Sci. Int. Genet. Suppl. Ser. 2017, 6, e257–e259. [Google Scholar] [CrossRef] [Green Version]
- Salas-Huetos, A.; Blanco, J.; Vidal, F.; Godo, A.; Grossmann, M.; Pons, M.C.; F-Fernández, S.; Garrido, N.; Anton, E. Spermatozoa from patients with seminal alterations exhibit a differential micro-ribonucleic acid profile. Fertil. Steril. 2015, 104, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J.; Frazier, K.S. Swine as Models in Biomedical Research and Toxicology Testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef]
- Roca, J.; Parrilla, I.; Bolarin, A.; Martinez, E.A.; Rodriguez-Martinez, H. Will AI in pigs become more efficient? Theriogenology 2016, 86, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Alcivar, A.A.; Hake, L.E.; Millette, C.F.; Trasler, J.M.; Hecht, N.B. Mitochondrial gene expression in male germ cells of the mouse. Dev. Biol. 1989, 135, 263–271. [Google Scholar] [CrossRef]
- Gur, Y.; Breitbart, H. Mammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomes. Genes Dev. 2006, 20, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gòdia, M.; Estill, M.; Castelló, A.; Balasch, S.; Rodríguez-Gil, J.E.; Krawetz, S.A.; Sánchez, A.; Clop, A. A RNA-seq analysis to describe the boar sperm transcriptome and its seasonal changes. Front. Genet. 2019, 10, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Beaulieu, J.F.; Huggett, J.; Jaggi, R.; Kibenge, F.S.B.; Olsvik, P.A.; Penning, L.C.; Toegel, S. MIQE précis: Practical implementation of minimum standard guidelines for fluorescence-based quantitative real-time PCR experiments. BMC Mol. Biol. 2010, 11, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Z.J.; Yi, W.; Rong, Y.W.; Kee, J.D.; Zhong, W.X. MicroRNA-1285 regulates 17β-estradiol-inhibited immature boar sertoli cell proliferation via adenosine monophosphate-Activated protein kinase activation. Endocrinology 2015, 156, 4059–4070. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, L.; Dinç, E. Evaluation of microRNA responses in ARPE-19 cells against the oxidative stress. Cutan. Ocul. Toxicol. 2018, 37, 121–126. [Google Scholar] [CrossRef]
- Jin, W.; Chen, F.; Wang, K.; Song, Y.; Fei, X.; Wu, B. miR-15a/miR-16 cluster inhibits invasion of prostate cancer cells by suppressing TGF-β signaling pathway. Biomed. Pharmacother. 2018, 104, 637–644. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Q.; Chamba, Y.; Zhang, B.; Shang, P.; Zhang, H.; Wu, C. Identification of Genes Related to Growth and Lipid Deposition from Transcriptome Profiles of Pig Muscle Tissue. PLoS ONE 2015, 10, e0141138. [Google Scholar] [CrossRef] [Green Version]
- He, K.; Niu, G.; Gao, J.; Liu, J.X.; Qu, H. MicroRNA-92 expression may be associated with reduced estrogen receptor β1 mRNA levels in cervical portion of uterosacral ligaments in women with pelvic organ prolapse. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 198, 94–99. [Google Scholar] [CrossRef]
- Shin, V.Y.; Siu, M.-T.; Liu, X.; Ng, E.K.O.; Kwong, A.; Chu, K.-M. MiR-92 suppresses proliferation and induces apoptosis by targeting EP4/Notch1 axis in gastric cancer. Oncotarget 2018, 9, 24209. [Google Scholar] [CrossRef] [Green Version]
- Qiu, T.; Wang, K.; Li, X.; Jin, J. MiR-671-5p inhibits gastric cancer cell proliferatioand promotes cell apoptosis by targeting URGCP. Exp. Ther. Med. 2018, 16, 4753–4758. [Google Scholar] [PubMed]
- Li, X.; Nie, C.; Tian, B.; Tan, X.; Han, W.; Wang, J.; Jin, Y.; Li, Y.; Guan, X.; Hong, A.; et al. MiR-671-5p blocks the progression of human esophageal squamous cell carcinoma by suppressing FGFR2. Int. J. Biol. Sci. 2019, 15, 1892–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Lan, Z.-J.; Li, X.; Lin, J.; Lei, Z. Role of postnatal expression of fgfr1 and fgfr2 in testicular germ cells on spermatogenesis and fertility in mice. J. Reprod. Infertil. 2014, 15, 122–133. [Google Scholar] [PubMed]
- Xie, M.Y.; Hou, L.J.; Sun, J.J.; Zeng, B.; Xi, Q.Y.; Luo, J.Y.; Chen, T.; Zhang, Y.L. Porcine Milk Exosome MiRNAs Attenuate LPS-Induced Apoptosis through Inhibiting TLR4/NF-κB and p53 Pathways in Intestinal Epithelial Cells. J. Agric. Food Chem. 2019, 67, 9477–9491. [Google Scholar] [CrossRef]
- Cai, Q.; Zhao, A.; Ren, L.G.; Chen, J.; Liao, K.S.; Wang, Z.S.; Zhang, W. MiR-425 Involves in the Development and Progression of Renal Cell Carcinoma by Inhibiting E2F6. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6300–6307. [Google Scholar]
- Chen, F.F.; Xiong, Y.; Peng, Y.; Gao, Y.; Qin, J.; Chu, G.Y.; Pang, W.J.; Yang, G.S. MiR-425-5p inhibits differentiation and proliferation in porcine intramuscular preadipocytes. Int. J. Mol. Sci. 2017, 18, 2101. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Wu, X.; Wang, J. Correlation of miR-425-5p and IL-23 with pancreatic cancer. Oncol. Lett. 2019, 17, 4595–4599. [Google Scholar] [CrossRef]
- Kastelein, R.A.; Hunter, C.A.; Cua, D.J. Discovery and Biology of IL-23 and IL-27: Related but Functionally Distinct Regulators of Inflammation. Annu. Rev. Immunol. 2007, 25, 221–242. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wu, M.; Chong, Q.Y.; Zhang, W.; Qian, P.; Yan, H.; Qian, W.; Zhang, M.; Lobie, P.E.; Zhu, T. Amplification of hsa-miR-191/425 locus promotes breast cancer proliferation and metastasis by targeting DICER1. Carcinogenesis 2018, 39, 1506–1516. [Google Scholar] [CrossRef]
- Gu, Y.; Ampofo, E.; Menger, M.D.; Laschke, M.W. MIR-191 suppresses angiogenesis by activation of NF-kB signaling. FASEB J. 2017, 31, 3321–3333. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Zhang, Z. miR-191/DAB2 axis regulates the tumorigenicity of estrogen receptor-positive breast cancer. IUBMB Life 2018, 70, 71–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Zhang, F.; Tong, J.; Liu, F. MiR-191-5p inhibits lung adenocarcinoma by repressing SATB1 to inhibit Wnt pathway. Mol. Genet. Genomic Med. 2020, 8, e1043. [Google Scholar] [CrossRef] [PubMed]
- Broekhuijse, M.; Sostaric, E.; Feitsma, H.; Gadella, B. The value of microscopic semen motility assessment at collection for a commercial artificial insemination center, a retrospective study on factors explaining variation in pig fertility. Theriogenology 2012, 77, 1466–1479. [Google Scholar] [CrossRef] [PubMed]
- Riesco, M.F.; Valcarce, D.G.; Martínez-Vázquez, J.M.; Robles, V. Effect of low sperm quality on progeny: A study on zebrafish as model species. Sci. Rep. 2019, 9, 11192. [Google Scholar] [CrossRef] [Green Version]
- Abu-Halima, M.; Hammadeh, M.; Schmitt, J.; Leidinger, P.; Keller, A.; Meese, E.; Backes, C. Altered microRNA expression profiles of human spermatozoa in patients with different spermatogenic impairments. Fertil. Steril. 2013, 99, 1249–1255. [Google Scholar] [CrossRef]
- Michailov, Y.; Ickowicz, D.; Breitbart, H. Zn2+-stimulation of sperm capacitation and of the acrosome reaction is mediated by EGFR activation. Dev. Biol. 2014, 396, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Lian, N.; Niu, Q.; Lei, Y.; Li, X.; Li, Y.; Song, X. MiR-221 is involved in depression by regulating Wnt2/CREB/BDNF axis in hippocampal neurons. Cell Cycle 2018, 17, 2745–2755. [Google Scholar] [CrossRef] [Green Version]
- Jacob, A. Molecular characterization of a voltage-gated potassium channel expressed in rat testis. Mol. Hum. Reprod. 2000, 6, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Carrillo, A.; Álvarez-Rodríguez, M.; Rodríguez-Martínez, H. The CatSper channel modulates boar sperm motility during capacitation. Reprod. Biol. 2017, 17, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, J.; Cho, K.-H.; Ren, D. A Novel, Single, Transmembrane Protein CATSPERG Is Associated with CATSPER1 Channel Protein1. Biol. Reprod. 2009, 81, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Ghanbari, H.; Keshtgar, S.; Zare, H.R.; Gharesi-Fard, B. Inhibition of CatSper and Hv1 Channels and NOX5 Enzyme Affect Progesterone-Induced Increase of Intracellular Calcium Concentration and ROS Generation in Human Sperm. Iran. J. Med. Sci. 2019, 44, 127–134. [Google Scholar] [PubMed]
- Ren, D.; Xia, J. Calcium signaling through CatSper channels in mammalian fertilization. Physiology 2010, 25, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassandri, M.; Smirnov, A.; Novelli, F.; Pitolli, C.; Agostini, M.; Malewicz, M.; Melino, G.; Raschellà, G. Zinc-finger proteins in health and disease. Cell Death Discov. 2017, 3, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonn Eisinger, K.R.; Woolfrey, K.M.; Swanson, S.P.; Schnell, S.A.; Meitzen, J.; Dell’Acqua, M.; Mermelstein, P.G. Palmitoylation of caveolin-1 is regulated by the same DHHC acyltransferases that modify steroid hormone receptors. J. Biol. Chem. 2018, 293, 15901–15911. [Google Scholar] [CrossRef] [Green Version]
- Aquila, S.; Sisci, D.; Gentile, M.; Middea, E.; Catalano, S.; Carpino, A.; Rago, V.; Andò, S. Estrogen Receptor (ER)α and ERβ Are Both Expressed in Human Ejaculated Spermatozoa: Evidence of Their Direct Interaction with Phosphatidylinositol-3-OH Kinase/Akt Pathway. J. Clin. Endocrinol. Metab. 2004, 89, 1443–1451. [Google Scholar] [CrossRef] [Green Version]
- Gou, D.M.; Sun, Y.; Gao, L.; Chow, L.M.C.; Huang, J.; Feng, Y.D.; Jiang, D.H.; Li, W.X. Cloning and characterization of a novel Krüppel-like zinc finger gene, ZNF268, expressed in early human embryo. Biochim. Biophys. Acta-Gene Struct. Expr. 2001, 1518, 306–310. [Google Scholar] [CrossRef]
- Kroft, T.L.; Patterson, J.; Won Yoon, J.; Doglio, L.; Walterhouse, D.O.; Iannaccone, P.M.; Goldberg, E. GLI1 Localization in the Germinal Epithelial Cells Alternates Between Cytoplasm and Nucleus: Upregulation in Transgenic Mice Blocks Spermatogenesis in Pachytene1. Biol. Reprod. 2001, 65, 1663–1671. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.T.; Bang, H.J.; Attipoe, S.A.; Johnston, D.S.; Tomsig, J.L. Sonic hedgehog pathway inhibition alters epididymal function as assessed by the development of sperm motility. J. Androl. 2006, 27, 225–232. [Google Scholar] [CrossRef]
- Edwards, D.R.; Handsley, M.M.; Pennington, C.J. The ADAM metalloproteinases. Mol. Asp. Med. 2009, 29, 258–289. [Google Scholar] [CrossRef]
- Atikuzzaman, M.; Alvarez-Rodriguez, M.; Vicente-Carrillo, A.; Johnsson, M.; Wright, D.; Rodriguez-Martinez, H. Conserved gene expression in sperm reservoirs between birds and mammals in response to mating. BMC Genom. 2017, 18, 98. [Google Scholar] [CrossRef] [Green Version]
- Atikuzzaman, M.; Hoglund, A.; Johnsson, M.; Alvarez-Rodriguez, M.; Mehta, R.; Wright, D.; Rodriguez-Martinez, H. Spermatozoa rather than seminal fluid drives immune-modulation in the sperm storage tubuli of Red Jungle Fowl hens. Reprod. Domest. Anim. 2015, 50, 44. [Google Scholar]
- Choi, H.; Han, C.; Jin, S.; Kwon, J.T.; Kim, J.; Jeong, J.; Kim, J.; Ham, S.; Jeon, S.; Yoo, Y.J.; et al. Reduced Fertility and Altered Epididymal and Sperm Integrity in Mice Lacking ADAM71. Biol. Reprod. 2015, 93, 1–11. [Google Scholar] [CrossRef]
- Wei, X.; Moncada-Pazos, A.; Cal, S.; Soria-Valles, C.; Gartner, J.; Rudloff, U.; Lin, J.C.; Rosenberg, S.A.; López-Otín, C.; Samuels, Y. Analysis of the disintegrin-metalloproteinases family reveals ADAM29 and ADAM7 are often mutated in melanoma. Hum. Mutat. 2011, 32, E2148–E2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinawath, N.; Shiao, M.S.; Norris, A.; Murphy, K.; Klein, A.P.; Yonescu, R.; Iacobuzio-Donahue, C.; Meeker, A.; Jinawath, A.; Yeo, C.J.; et al. Alterations of type II classical cadherin, cadherin-10 (CDH10), is associated with pancreatic ductal adenocarcinomas. Genes Chromosom. Cancer 2017, 56, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, A.; Matusiak, Ł.; Szczerkowska-Dobosz, A.; Nowak, I.; Kuśnierczyk, P. HLA-C*06:02-independent, gender-related association of PSORS1C3 and PSORS1C1/CDSN single-nucleotide polymorphisms with risk and severity of psoriasis. Mol. Genet. Genom. 2018, 293, 957–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Soni, U.K.; Maurya, V.K.; Singh, K.; Jha, R.K. Integrin beta8 (ITGB8) activates VAV-RAC1 signaling via FAK in the acquisition of endometrial epithelial cell receptivity for blastocyst implantation. Sci. Rep. 2017, 7, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santulli, G. Angiopoietin-like proteins: A comprehensive look. Front. Endocrinol. 2014, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, M.; Mulders, J.; Könst, A.; Janssens, B.; van Roy, F.; Blankenstein, M.; Oudejans, C. Differential downregulation of αT-catenin expression in placenta: Trophoblast cell type-dependent imprinting of the CTNNA3 gene. Gene Expr. Patterns 2004, 5, 61–65. [Google Scholar] [CrossRef]
- Chapman, E.M.; Lant, B.; Ohashi, Y.; Yu, B.; Schertzberg, M.; Go, C.; Dogra, D.; Koskimäki, J.; Girard, R.; Li, Y.; et al. A conserved CCM complex promotes apoptosis non-autonomously by regulating zinc homeostasis. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Bragantini, B.; Tiotiu, D.; Rothé, B.; Saliou, J.M.; Marty, H.; Cianférani, S.; Charpentier, B.; Quinternet, M.; Manival, X. Functional and Structural Insights of the Zinc-Finger HIT protein family members Involved in Box C/D snoRNP Biogenesis. J. Mol. Biol. 2016, 428, 2488–2506. [Google Scholar] [CrossRef]
- Stephan, J.S.; Fioriti, L.; Lamba, N.; Colnaghi, L.; Karl, K.; Derkatch, I.L.; Kandel, E.R. The CPEB3 Protein Is a Functional Prion that Interacts with the Actin Cytoskeleton. Cell Rep. 2015, 11, 1772–1785. [Google Scholar] [CrossRef] [PubMed]
- Drisaldi, B.; Colnaghi, L.; Fioriti, L.; Rao, N.; Myers, C.; Snyder, A.M.; Metzger, D.J.; Tarasoff, J.; Konstantinov, E.; Fraser, P.E.; et al. SUMOylation Is an Inhibitory Constraint that Regulates the Prion-like Aggregation and Activity of CPEB3. Cell Rep. 2015, 11, 1694–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Zhang, G.; Lv, S.; Wen, X.; Liu, P. miRNA-301b-3p accelerates migration and invasion of high-grade ovarian serous tumor via targeting CPEB3/EGFR axis. J. Cell. Biochem. 2019, 120, 12618–12627. [Google Scholar] [CrossRef]
- Wilson, V.G. Introduction to sumoylation. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2017; Volume 963, pp. 1–12. [Google Scholar]
- Mei, J.; Yan, Y.; Li, S.Y.; Zhou, W.J.; Zhang, Q.; Li, M.Q.; Sun, H.X. CXCL16/CXCR6 interaction promotes endometrial decidualization via the PI3K/AKT pathway. Reproduction 2019, 157, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Guerin, L.R.; Moldenhauer, L.M.; Prins, J.R.; Bromfield, J.J.; Hayball, J.D.; Robertson, S.A. Seminal Fluid Regulates Accumulation of FOXP3+ Regulatory T Cells in the Preimplantation Mouse Uterus Through Expanding the FOXP3+ Cell Pool and CCL19-Mediated Recruitment1. Biol. Reprod. 2011, 85, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Krikun, G. The CXL12/CXCR4/CXCR7 axis in female reproductive tract disease: Review. Am. J. Reprod. Immunol. 2018, 80, e13028. [Google Scholar] [CrossRef]
- Mendis, S.H.S.; Meachem, S.J.; Sarraj, M.A.; Loveland, K.L. Activin A Balances Sertoli and Germ Cell Proliferation in the Fetal Mouse Testis. Biol. Reprod. 2011, 84, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Yuan, S.; Swiggin, H.M.; Zheng, H.; Yan, W. A testis-specific gene, Ubqlnl, is dispensable for mouse embryonic development and spermatogenesis. Mol. Reprod. Dev. 2015, 82, 408–409. [Google Scholar] [CrossRef] [Green Version]
- Marín, I. The ubiquilin gene family: Evolutionary patterns and functional insights. BMC Evol. Biol. 2014, 14, 63. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Lan, Y.; Xu, J.; Zhang, W.; Wen, Z. SUMO1-activating enzyme subunit 1 is essential for the survival of hematopoietic stem/progenitor cells in zebrafish. Development 2012, 139, 4321–4329. [Google Scholar] [CrossRef] [Green Version]
- Shin, E.J.; Shin, H.M.; Nam, E.; Kim, W.S.; Kim, J.H.; Oh, B.H.; Yun, Y. DeSUMOylating isopeptidase: A second class of SUMO protease. EMBO Rep. 2012, 13, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Judith, D.; Jefferies, H.B.J.; Boeing, S.; Frith, D.; Snijders, A.P.; Tooze, S.A. ATG9A shapes the forming autophagosome through Arfaptin 2 and phosphatidylinositol 4-kinase IIIβ. J. Cell Biol. 2019, 218, 1634–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukamachi, K.; Matsuoka, Y.; Ohno, H.; Hamaguchi, T.; Tsuda, H. Neuronal leucine-rich repeat protein-3 amplifies MAPK activation by epidermal growth factor through a carboxyl-terminal region containing endocytosis motifs. J. Biol. Chem. 2002, 277, 43549–43552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Duan, H.; Duan, C.; Zhu, H.; Zhou, R.; Pei, H.; Shen, L. TCF21 functions as a tumor suppressor in colorectal cancer through inactivation of PI3K/AKT signaling. Onco Targets Ther. 2017, 10, 1603–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad Aslam, M.K.; Kumaresan, A.; Yadav, S.; Mohanty, T.K.; Datta, T.K. Comparative proteomic analysis of high- and low-fertile buffalo bull spermatozoa for identification of fertility-associated proteins. Reprod. Domest. Anim. 2019, 54, 786–794. [Google Scholar]
- Du, Y.; Huang, Y.; Gao, Y.; Song, B.; Mao, J.; Chen, L.; Bai, L.; Tang, J. Annexin A7 modulates BAG4 and BAG4-binding proteins in mitochondrial apoptosis. Biomed. Pharmacother. 2015, 74, 30–34. [Google Scholar] [CrossRef]
- Thomas, P.D.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. PANTHER: A library of protein families and subfamilies indexed by function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript ID | Expression Mean ± Standard Deviation (log2) | Accession | Sequence Length | Sequence |
|---|---|---|---|---|
| ssc-miR-1285 | 13.98 ± 0.13 | MIMAT0013954 | 24 | CUGGGCAACAUAGCGAGACCCCGU |
| ssc-miR-16 | 12.6 ± 0.74 | MIMAT0007754 | 22 | UAGCAGCACGUAAAUAUUGGCG |
| ssc-miR-4332 | 12.32 ± 0.29 | MIMAT0017962 | 20 | CACGGCCGCCGCCGGGCGCC |
| ssc-miR-92a | 12.06 ± 0.09 | MIMAT0013908 | 22 | UAUUGCACUUGUCCCGGCCUGU |
| ssc-miR-671-5p | 11.73 ± 0.54 | MIMAT0025381 | 24 | AGGAAGCCCUGGAGGGGCUGGAGG |
| ssc-miR-4334-5p | 11.31 ± 0.05 | MIMAT0017966 | 19 | CCCUGGAGUGACGGGGGUG |
| ssc-miR-425-5p | 10.99 ± 0.15 | MIMAT0013917 | 23 | AAUGACACGAUCACUCCCGUUGA |
| ssc-miR-191 | 10.57 ± 0.22 | MIMAT0013876 | 23 | CAACGGAAUCCCAAAAGCAGCUG |
| ssc-miR-92b-5p | 10.53 ± 0.18 | MIMAT0017377 | 24 | AGGGACGGGACGCGGUGCAGUGUU |
| ssc-miR-15b | 10.01 ± 0.9 | MIMAT0002125 | 22 | UAGCAGCACAUCAUGGUUUACA |
| UP-REGULATED GENES in High-Fertile Boars vs. Low-Fertile Boars | DESCRIPTION | Fold Change | DOWN-REGULATED GENES in High-Fertile Boars vs. Low-Fertile Boars | DESCRIPTION | Fold Change |
|---|---|---|---|---|---|
| LOC100739568 | replication factor C subunit 1-like | 5.72 | IFN-DELTA-4; LOC100736862 | interferon-delta-4; interferon tau-11-like | −2 |
| AGPAT2 | 1-acylglycerol-3-phosphate O-acyltransferase 2 | 3.53 | FOXP2; LOC100620234 | forkhead box P2; forkhead box protein P2-like | −2.03 |
| FBXW5 | F-box and WD repeat domain containing 5 | 3.52 | LOC100155405 | olfactory receptor 4K1 | −2.03 |
| PPP1R16A | protein phosphatase 1, regulatory subunit 16A | 3.34 | LOC100516093 | olfactory receptor 52N4 | −2.27 |
| PIM1 | Pim-1 proto-oncogene, serine/threonine kinase | 3.26 | LOC100517289 | E3 ubiquitin-protein ligase RNF220 | −2.41 |
| RAB7A | RAB7A, member RAS oncogene family | 3.26 | LOC100738965; LOC100739445 | olfactory receptor 1J4-like | −2.49 |
| C12H17orf97 | chromosome 12 open reading frame, human C17orf97 | 3.1 | LOC100525437 | ATP synthase subunit d, mitochondrial-like | −3.06 |
| TMEM239; C17H20orf141 | transmembrane protein 239; chromosome 17 open reading frame, human C20orf141 | 3.1 | LOC100157815 | olfactory receptor 4K15 | −3.9 |
| SAE1 | SUMO1 activating enzyme subunit 1 | 3.01 | LOC100522664; LOC100622735 | olfactory receptor 2AJ1-like | −4.97 |
| TSSK6 | testis-specific serine kinase 6 | 2.91 | LOC100157785 | olfactory receptor 8H1-like | −5.22 |
| WNT3 | wingless-type MMTV integration site family, member 3 | 2.91 | |||
| CARHSP1 | calcium regulated heat stable protein 1, 24kDa | 2.88 | |||
| UBQLNL | ubiquilin-like | 2.79 | |||
| TSSK2 | testis-specific serine kinase 2 | 2.77 | |||
| MAP1LC3A | microtubule-associated protein 1 light chain 3 alpha | 2.71 | |||
| RABAC1 | Rab acceptor 1 (prenylated) | 2.68 | |||
| PHKG2 | phosphorylase kinase, gamma 2 (testis) | 2.66 | |||
| MTHFSD | methenyltetrahydrofolate synthetase domain containing | 2.61 | |||
| CSNK1G2 | casein kinase 1, gamma 2 | 2.56 | |||
| C9H11orf71 | chromosome 9 open reading frame, human C11orf71 | 2.54 | |||
| AGBL5 | ATP/GTP binding protein-like 5 | 2.51 | |||
| SGSM2 | small G protein signaling modulator 2 | 2.47 | |||
| TIMP2 | TIMP metallopeptidase inhibitor 2 | 2.47 | |||
| DUSP18 | dual specificity phosphatase 18 | 2.45 | |||
| SARNP | SAP domain containing ribonucleoprotein | 2.45 | |||
| TNP1 | transition protein 1 (during histone to protamine replacement) | 2.45 | |||
| FAM57A | family with sequence similarity 57, member A | 2.44 | |||
| EHD1 | EH-domain containing 1 | 2.43 | |||
| NSUN4 | NOP2/Sun domain family, member 4 | 2.4 | |||
| C7H6orf125 | uncharacterized protein C6orf125 homolog | 2.39 | |||
| EIF4A3; LOC100739660 | eukaryotic translation initiation factor 4A3; eukaryotic initiation factor 4A-III-like | 2.38 | |||
| NDUFA10 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 10, 42kDa | 2.36 | |||
| KHDRBS3; LOC100625959 | KH domain containing, RNA binding, signal transduction associated 3; KH domain-containing, RNA-binding, signal transduction-associated protein 3-like | 2.33 | |||
| CHCHD5 | coiled-coil-helix-coiled-coil-helix domain containing 5 | 2.32 | |||
| OSBP2 | oxysterol binding protein 2 | 2.3 | |||
| C1H9orf16; LCN2 | chromosome 1 open reading frame, human C9orf16; lipocalin 2 | 2.29 | |||
| GRK4 | G protein-coupled receptor kinase 4 | 2.28 | |||
| PSMF1; LOC100736938 | proteasome inhibitor subunit 1; proteasome inhibitor PI31 subunit | 2.28 | |||
| CATSPERG | catsper channel auxiliary subunit gamma | 2.27 | |||
| MFAP3L | microfibrillar-associated protein 3-like | 2.26 | |||
| TSACC | TSSK6 activating co-chaperone | 2.25 | |||
| LOC100155534 | verprolin-like | 2.24 | |||
| LOC102166949 | E3 ubiquitin-protein ligase RNF169 | 2.24 | |||
| NEURL1 | neuralized E3 ubiquitin protein ligase 1 | 2.24 | |||
| FAM46C | family with sequence similarity 46, member C | 2.22 | |||
| ISG20L2 | interferon stimulated exonuclease gene 20kDa-like 2 | 2.19 | |||
| MIEF1 | mitochondrial elongation factor 1 | 2.19 | |||
| HTR4; LOC100737835 | 5-hydroxytryptamine (serotonin) receptor 4, G protein-coupled; 5-hydroxytryptamine receptor 4 | 2.18 | |||
| VRK3 | vaccinia related kinase 3 | 2.16 | |||
| CARM1 | coactivator-associated arginine methyltransferase 1 | 2.15 | |||
| LOC100152461; LOC100627430; NFYA | nuclear transcription factor Y subunit alpha; nuclear transcription factor Y, alpha | 2.15 | |||
| LOC100157936; LOC102162592 | E3 ubiquitin-protein ligase NRDP1; E3 ubiquitin-protein ligase NRDP1-like | 2.13 | |||
| LOC102162328 | uncharacterized LOC102162328 | 2.13 | |||
| SYCE2 | synaptonemal complex central element protein 2 | 2.12 | |||
| UBE2D2 | ubiquitin-conjugating enzyme E2D 2 | 2.12 | |||
| LOC100625908; LOC102162228 | uncharacterized LOC100625908; uncharacterized LOC102162228 | 2.1 | |||
| ACTR3B | ARP3 actin-related protein 3 homolog B (yeast) | 2.09 | |||
| CCNY | cyclin Y | 2.09 | |||
| CPEB3 | cytoplasmic polyadenylation element binding protein 3 | 2.09 | |||
| LOC100511504; LOC100515480 | maestro heat-like repeat-containing protein family member 1 | 2.09 | |||
| LOC100737397; C1H9orf9 | uncharacterized protein C9orf9 homolog; chromosome 1 open reading frame, human C9orf9 | 2.07 | |||
| LOC102158209 | uncharacterized LOC102158209 | 2.04 | |||
| REEP2 | receptor accessory protein 2 | 2.04 | |||
| TTC7A | tetratricopeptide repeat domain 7A | 2.04 | |||
| GNA12 | guanine nucleotide binding protein (G protein) alpha 12 | 2.02 | |||
| PIM3; BRD1 | Pim-3 proto-oncogene, serine/threonine kinase; bromodomain containing 1 | 2.02 | |||
| DESI1 | desumoylating isopeptidase 1 | 2 |
| A. PANTHER Molecular Function Class | Sus Scrofa Ref | Expected | Fold Enrichment | P Value | Gene IDs |
|---|---|---|---|---|---|
| C-C chemokine binding (GO:0019957) | 24 | 0.38 | 7.93 | 6.76E-03 | CCR7, XCR1, CXCR4 |
| chemokine binding (GO:0019956) | 24 | 0.38 | 7.93 | 6.76E-03 | CCR7, XCR1, CXCR4 |
| cytokine binding (GO:0019955) | 88 | 1.39 | 4.33 | 2.98E-03 | CCR7, XCR1, CXCR4, ACVR2B, IL23R |
| protein binding (GO:0005515) | 2824 | 44.49 | 1.51 | 4.32E-04 | WDR34, MTF2, ELOF1, KIF15, KAT7, FGFR2, GPAT3, ATP6V0A2, AGBL5, CCNY,BAG6, TRAF3, EHD1, WDTC1, PPP1R16A, CXCL16, MICU1, AKAP11, MAP1LC3A, SGSM2, DVL1, UBQLNL, ERC1, MOBKL3, SNX14, WNT3, BAG6, NRG4, SHC3, GNA12, UBE2D2, STYX, FIBCD1, TSACC, ATN1, CDH10, STXBP6, TMEFF2, CCR7, SERPINE2, FGD4, AKAP7, ACVR1C, CDSN, NUF2, ACVR2B, PPARGC1B, SYT1, GDF9, ANKRD55, GNAT3, ANGPTL1, WFDC9, SYT10, IL23R, SERPINI2, TFPI2, CTNNA3, XCR1, TEAD3, CXCR4, BRCC3, CDSN, CCNJ |
| cytokine receptor activity (GO:0005126) | 75 | 1.18 | 3.39 | 3.20E-02 | CCR7, XCR1, CXCR4, IL23R |
| B. PANTHER Protein Class | Sus Scrofa Ref | Expected | Fold Enrichment | P Value | Gene IDs |
| TGF-beta receptor (PC00035) | 15 | 0.24 | 12.69 | 1.83E-03 | ACVR1C, ACVR2B |
| serine/threonine protein kinase receptor (PC00205) | 20 | 0.32 | 9.52 | 4.10E-03 | ACVR1C, ACVR2B |
| protein kinase (PC00193) | 266 | 4.19 | 2.39 | 1.05E-02 | TSSK2, TSSK6, RPS6KC1, PRKCQ, VRK3, CSNK1G2, GRK4, ACVR1C, ACVR2B |
| kinase (PC00137) | 386 | 6.08 | 1.97 | 2.10E-02 | TSSK2, TSSK6, NDUFA10, RPS6KC1, PRKCQ, VRK3, CSNK1G2, PI4K2A, GRK4, ACVR1C, ACVR2B |
| transferase (PC00220) | 907 | 14.29 | 1.82 | 2.79E-03 | NTMT1, CHST13, NSUN4, TSSK2, TSSK6, KAT7, GPAT3, METTL2A, GALNT18, NDUFA10, RPS6KC1, PRKCQ, VRK3, FAM177A1, CSNK1G2, CHPT1, PI4K2A, RNGTT, GRK4, ACCSL, ME1, ACVR1C, ACVR2B, TGM6 |
| miRNA ID | Gene Target | GO – Molecular Function | GO – Biological Process |
|---|---|---|---|
| miR-16 | UBQLNL (Ubiquilin-like protein) | GO:0031593 polyubiquitin modification-dependent protein binding | GO:0006511 ubiquitin-dependent protein catabolic process |
| CPEB3 (Cytoplasmic polyadenylation element-binding protein 3) | GO:0000900 translation repressor activity, mRNA regulatory element binding | GO:0071230 cellular response to amino acid stimulus GO:0007616 long-term memory | |
| DESI1 (Desumoylating isopeptidase 1) | GO:0042802 identical protein binding | ||
| ATG9A (Autophagy-related protein 9A) | GO:0000045 autophagosome assembly | ||
| BAG4 (BAG family molecular chaperone regulator 4) | GO:0031625 ubiquitin protein ligase binding | GO:0071364 cellular response to epidermal growth factor stimulus GO:0045785 positive regulation of cell adhesion | |
| LRRN3 (Leucine-rich repeat neuronal protein 3) | GO:0051965 positive regulation of synapse assembly | ||
| miR-92a | CPEB3 (Cytoplasmic polyadenylation element-binding protein 3) | GO:0000900 translation repressor activity, mRNA regulatory element binding | GO:0071230 cellular response to amino acid stimulus GO:0007616 long-term memory |
| TCF21 (ranscription factor 21) | GO:0050681 androgen receptor binding | GO:0060766 negative regulation of androgen receptor signaling pathway | |
| PDZD8 (PDZ domain-containing protein 8) | GO:0008289 lipid binding | GO:0035556 intracellular signal transduction | |
| miR-15b | UBQLNL (Ubiquilin-like protein) | GO:0031593 polyubiquitin modification-dependent protein binding | GO:0006511 ubiquitin-dependent protein catabolic process |
| CPEB3 (Cytoplasmic polyadenylation element-binding protein 3) | GO:0000900 translation repressor activity, mRNA regulatory element binding | GO:0071230 cellular response to amino acid stimulus GO:0007616 long-term memory | |
| DESI1 (Desumoylating isopeptidase 1) | GO:0042802 identical protein binding | ||
| ATG9A (Autophagy-related protein 9A) | GO:0000045 autophagosome assembly | ||
| LRRN3 (Leucine-rich repeat neuronal protein 3) | GO:0051965 positive regulation of synapse assembly |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez-Rodriguez, M.; Martinez, C.; Wright, D.; Barranco, I.; Roca, J.; Rodriguez-Martinez, H. The Transcriptome of Pig Spermatozoa, and Its Role in Fertility. Int. J. Mol. Sci. 2020, 21, 1572. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051572
Alvarez-Rodriguez M, Martinez C, Wright D, Barranco I, Roca J, Rodriguez-Martinez H. The Transcriptome of Pig Spermatozoa, and Its Role in Fertility. International Journal of Molecular Sciences. 2020; 21(5):1572. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051572
Chicago/Turabian StyleAlvarez-Rodriguez, Manuel, Cristina Martinez, Dominic Wright, Isabel Barranco, Jordi Roca, and Heriberto Rodriguez-Martinez. 2020. "The Transcriptome of Pig Spermatozoa, and Its Role in Fertility" International Journal of Molecular Sciences 21, no. 5: 1572. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051572