Docking and Molecular Dynamics Predictions of Pesticide Binding to the Calyx of Bovine β-Lactoglobulin


Abstract
:
1. Introduction
2. Results
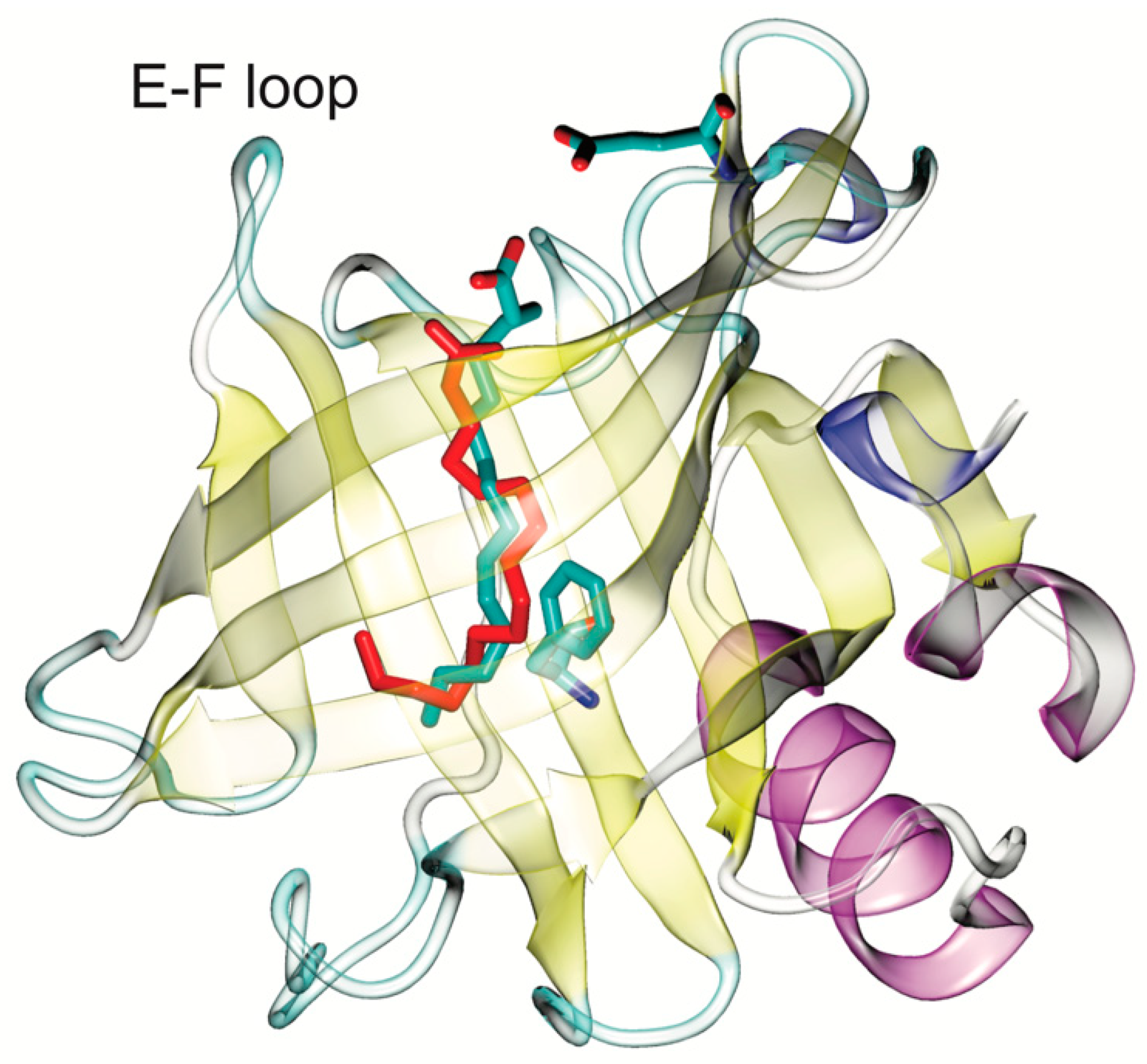
2.1. Pesticide Docking to β-Lactoglobulin
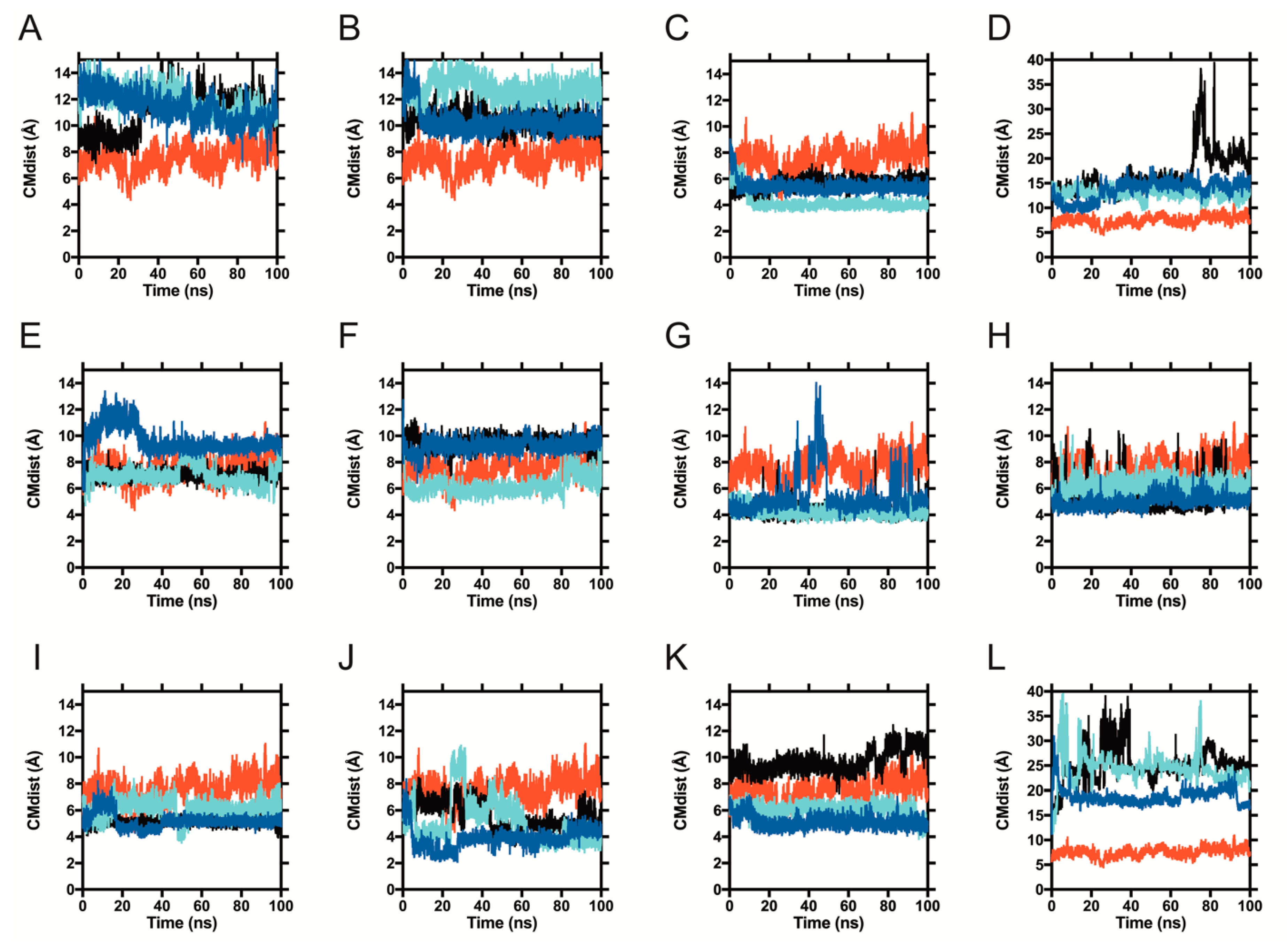
2.2. Molecular Dynamics of Pesticide-BLG Complexes
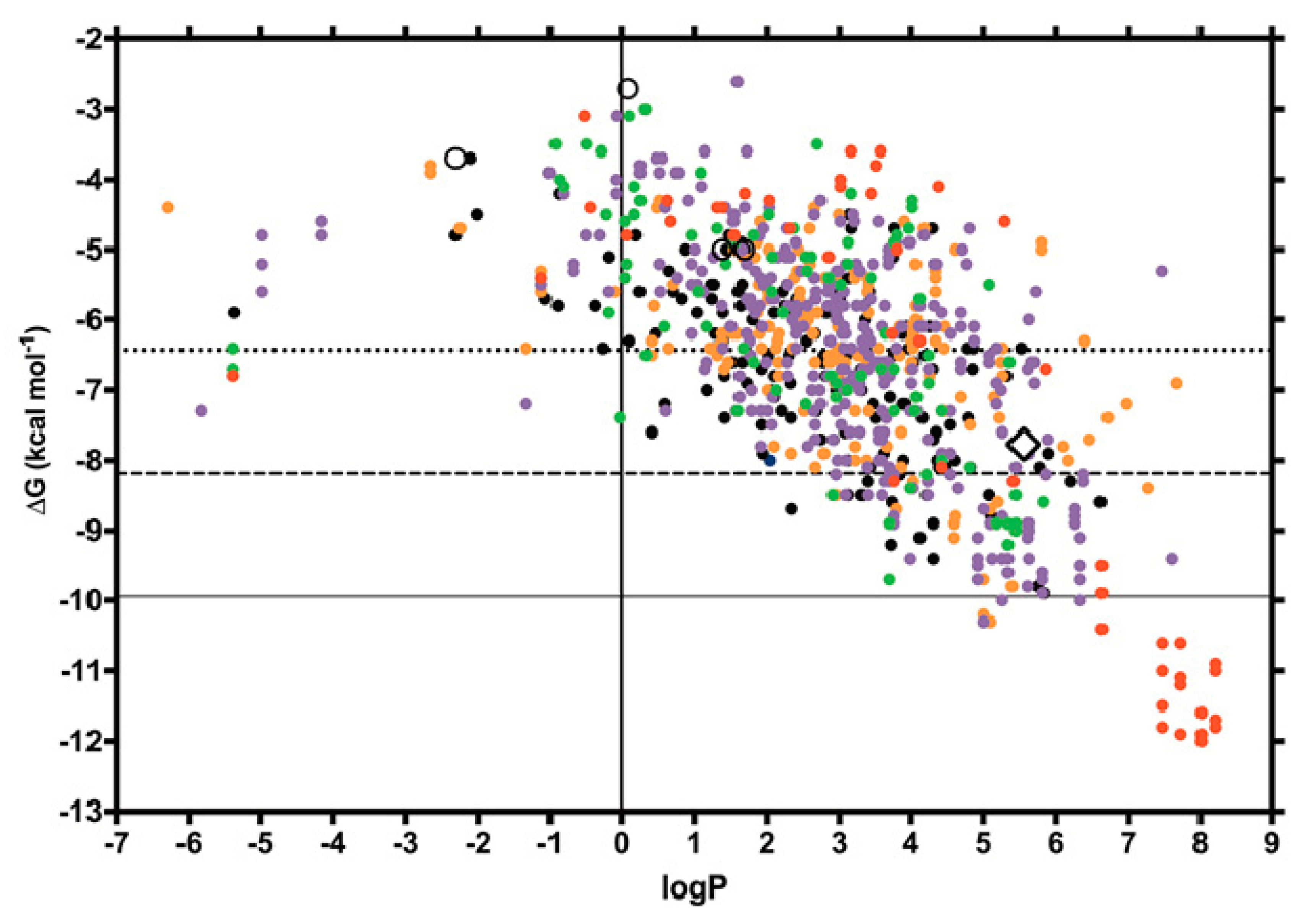
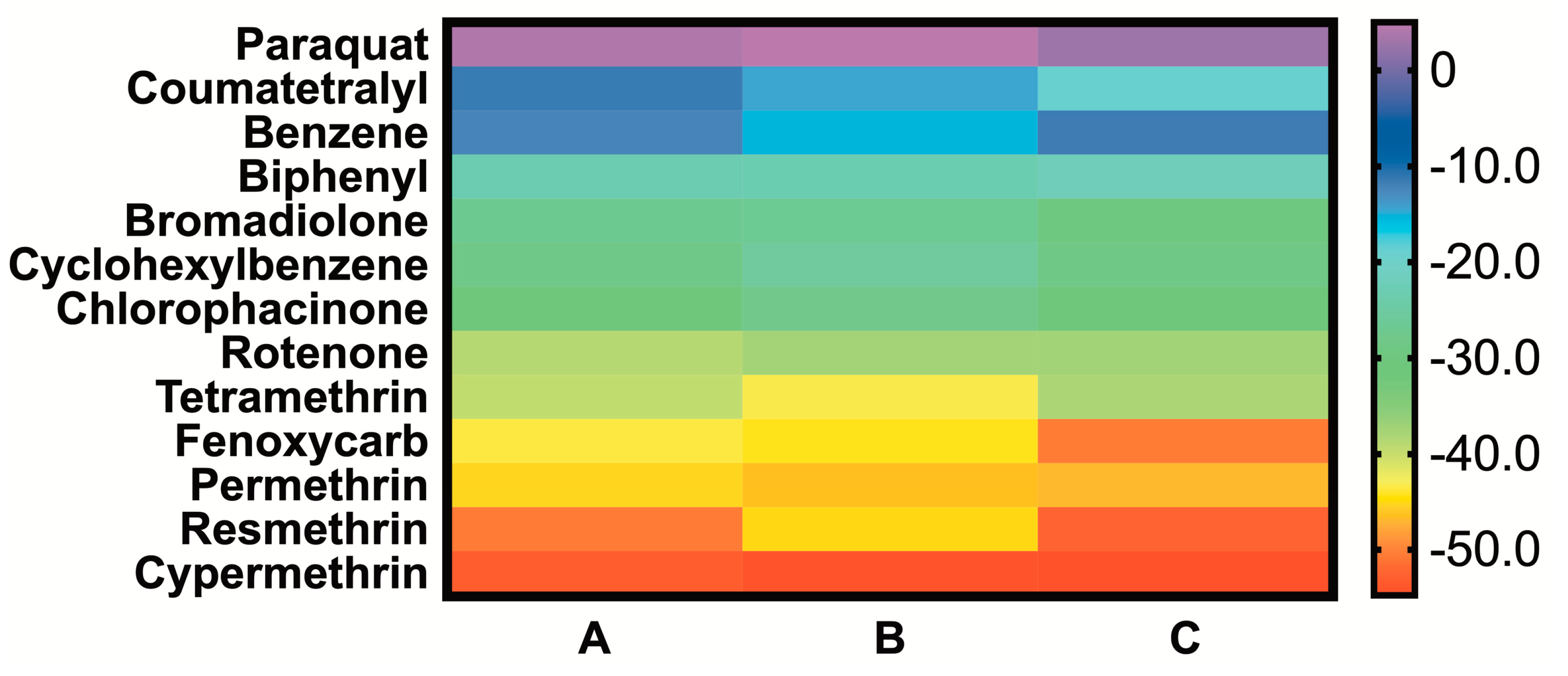
2.3. MMGBSA of the Simulated BG-Pesticide Complexes
3. Discussion
4. Conclusions
5. Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. International Code of Conduct on the Distribution and Use of Pesticides (Revised Version); Food and Agriculture Organization of the United Nations: Rome, Italy, 2003. [Google Scholar]
- FAO. FAOSTAT Pesticides Indicators; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- American Association for the Advancement of Science. Infographic: Pesticide Planet. Science 2013, 341, 730–731. [Google Scholar]
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical Pesticides and Human Health: The Urgent Need for a New Concept in Agriculture. Front. Public Health 2016, 4, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verger, P.J.P.; Boobis, A.R. Global Food Supply. Reevaluate Pesticides for Food Security and Safety. Science 2013, 341, 717–718. [Google Scholar] [CrossRef] [PubMed]
- WHO. The WHO Recommended Classification of Pesticides by Hazard and Guidelines to Classification 2009; WHO: Geneva, Switzerland, 2016; pp. 1–81. [Google Scholar]
- Tang, W.; Wang, D.; Wang, J.; Wu, Z.; Li, L.; Huang, M.; Xu, S.; Yan, D. Pyrethroid Pesticide Residues in the Global Environment: An Overview. Chemosphere 2018, 191, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Sassine, A.; Moura, S.; Léo, V.M.; Bustillos, O.V. Cypermethrin Residues Determination in the Milk of a Lactating Dairy Cow by Gas Chromatography-Ion Trap Mass Spectrometry. J. Anal. Toxicol. 2004, 28, 238–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, J.H.; González, M.M.; Noa, M.; Pérez, N.A.; Díaz, G.; Gutiérrez, R.; Zazueta, H.; Osuna, I. Organophosphorus Pesticide Residues in Mexican Commercial Pasteurized Milk. J. Agric. Food Chem. 2003, 51, 4468–4471. [Google Scholar] [CrossRef]
- Ng, P.K.; Snyder, M.A. Purification of Β-Lactoglobulin with a High-Capacity Anion Exchanger: High-Throughput Process Development and Scale-Up Considerations. J. Sci. Food Agric. 2013, 93, 3231–3236. [Google Scholar] [CrossRef]
- Ragona, L.; Fogolari, F.; Zetta, L.; Pérez, D.M.; Puyol, P.; De Kruif, K.; Löhr, F.; Rüterjans, H.; Molinari, H. Bovine Β-Lactoglobulin: Interaction Studies with Palmitic Acid. Protein Sci. 2000, 9, 1347–1356. [Google Scholar] [CrossRef]
- Sahihi, M.; Heidari-Koholi, Z.; Bordbar, A.-K. The Interaction of Polyphenol Flavonoids with Β-Lactoglobulin: Molecular Docking and Molecular Dynamics Simulation Studies. J. Macromol. Sci. Part B 2012, 51, 2311–2323. [Google Scholar] [CrossRef]
- Sawyer, L.; Kontopidis, G. The Core Lipocalin, Bovine Β-Lactoglobulin. Biochim. Biophys. Acta 2000, 1482, 136–148. [Google Scholar] [CrossRef]
- Pérez, M.D.; de Villegas, C.D.; Sánchez, L.; Aranda, P.; Ena, J.M.; Calvo, M. Interaction of Fatty Acids with Beta-Lactoglobulin and Albumin from Ruminant Milk. J. Biochem. 1989, 106, 1094–1097. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Ramírez, L.; del Moral-Ramírez, E.; Cortés-Hernández, P.; García-Garibay, M.; Jiménez-Guzmán, J. Β-Lactoglobulin’s Conformational Requirements for Ligand Binding at the Calyx and the Dimer Interphase: A Flexible Docking Study. PLoS ONE 2013, 8, e79530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayalakshmi, L.; Krishna, R.; Sankaranarayanan, R.; Vijayan, M. An Asymmetric Dimer of Β-Lactoglobulin in a Low Humidity Crystal Form—Structural Changes That Accompany Partial Dehydration and Protein Action. Proteins 2008, 71, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Evoli, S.; Guzzi, R.; Rizzuti, B. Molecular Simulations of Β-Lactoglobulin Complexed with Fatty Acids Reveal the Structural Basis of Ligand Affinity to Internal and Possible External Binding Sites. Proteins 2014, 82, 2609–2619. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-C.; Chen, N.C.; Chen, C.-J.; Wu, C.Y.; Mao, S.J.T. Evidence for Beta-Lactoglobulin Involvement in Vitamin D Transport in Vivo--Role of the Gamma-Turn (Leu-Pro-Met) of Beta-Lactoglobulin in Vitamin D Binding. FEBS J. 2009, 276, 2251–2265. [Google Scholar] [CrossRef] [PubMed]
- Loch, J.I.; Bonarek, P.; Polit, A.; Riès, D.; Dziedzicka-Wasylewska, M.; Lewiński, K. Binding of 18-Carbon Unsaturated Fatty Acids to Bovine Β-Lactoglobulin-Structural and Thermodynamic Studies. Int. J. Biol. Macromol. 2013, 57, 226–231. [Google Scholar] [CrossRef]
- Qvist, J.; Davidovic, M.; Hamelberg, D.; Halle, B. A Dry Ligand-Binding Cavity in a Solvated Protein. Proc. Natl. Acad. Sci. USA 2008, 105, 6296–6301. [Google Scholar] [CrossRef] [Green Version]
- Qin, B.Y.; Bewley, M.C.; Creamer, L.K.; Baker, H.M.; Baker, E.N.; Jameson, G.B. Structural Basis of the Tanford Transition of Bovine Beta-Lactoglobulin. Biochemistry 1998, 37, 14014–14023. [Google Scholar] [CrossRef]
- Bello, M.; García-Hernández, E. Ligand Entry Into the Calyx of Β-Lactoglobulin. Biopolymers 2014, 101, 744–757. [Google Scholar] [CrossRef]
- Sakurai, K.; Konuma, T.; Yagi, M.; Goto, Y. Structural Dynamics and Folding of Beta-Lactoglobulin Probed by Heteronuclear NMR. Biochim. Biophys. Acta 2009, 1790, 527–537. [Google Scholar] [CrossRef]
- Eberini, I.; Rocco, A.G.; Mantegazza, M.; Gianazza, E.; Baroni, A.; Vilardo, M.C.; Donghi, D.; Galliano, M.; Beringhelli, T. Computational and Experimental Approaches Assess the Interactions Between Bovine Beta-Lactoglobulin and Synthetic Compounds of Pharmacological Interest. J. Mol. Graph. Model. 2008, 26, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Loch, J.I.; Polit, A.; Bonarek, P.; Olszewska, D.; Kurpiewska, K.; Dziedzicka-Wasylewska, M.; Lewiński, K. Structural and Thermodynamic Studies of Binding Saturated Fatty Acids to Bovine Β-Lactoglobulin. Int. J. Biol. Macromol. 2012, 50, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Gangemi, S.; Miozzi, E.; Teodoro, M.; Briguglio, G.; De Luca, A.; Alibrando, C.; Polito, I.; Libra, M. Occupational Exposure to Pesticides as a Possible Risk Factor for the Development of Chronic Diseases in Humans (Review). Mol. Med. Rep. 2016, 14, 4475–4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-H.; Steffes, M.W.; Sjödin, A.; Jones, R.S.; Needham, L.L.; Jacobs, D.R. Low Dose Organochlorine Pesticides and Polychlorinated Biphenyls Predict Obesity, Dyslipidemia, and Insulin Resistance Among People Free of Diabetes. PLoS ONE 2011, 6, e15977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corcellas, C.; Feo, M.L.; Torres, J.P.; Malm, O.; Ocampo-Duque, W.; Eljarrat, E.; Barceló, D. Pyrethroids in Human Breast Milk: Occurrence and Nursing Daily Intake Estimation. Environ. Int. 2012, 47, 17–22. [Google Scholar] [CrossRef]
- Collotta, M.; Bertazzi, P.A.; Bollati, V. Epigenetics and Pesticides. Toxicology 2013, 307, 35–41. [Google Scholar] [CrossRef]
- Wardhaugh, K.G. Insecticidal Activity of Synthetic Pyrethroids, Organophosphates, Insect Growth Regulators, and Other Livestock Parasiticides: An Australian Perspective. Environ. Toxicol. Chem. 2005, 24, 789–796. [Google Scholar] [CrossRef]
- Singh, A.K.; Tiwari, M.N.; Prakash, O.; Singh, M.P. A Current Review of Cypermethrin-Induced Neurotoxicity and Nigrostriatal Dopaminergic Neurodegeneration. Curr. Neuropharmacol. 2012, 10, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Bissacot, D.Z.; Vassilieff, I. HPLC Determination of Flumethrin, Deltamethrin, Cypermethrin, and Cyhalothrin Residues in the Milk and Blood of Lactating Dairy Cows. J. Anal. Toxicol. 1997, 21, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Månsson, H.L. Fatty Acids in Bovine Milk Fat. Food Nutr. Res. 2008, 52, 1–3. [Google Scholar]
- Liang, L.; Zhang, J.; Zhou, P.; Subirade, M. Protective Effect of Ligand-Binding Proteins Against Folic Acid Loss Due to Photodecomposition. Food Chem. 2013, 141, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Shimoyamada, M.; Yoshimura, H.; Tomida, K.; Watanabe, K. Stabilities of Bovine Β-Lactoglobulin/Retinol or Retinoic Acid Complexes Against Tryptic Hydrolysis, Heating and Light-Induced Oxidation. Lebenson Wiss Technol. 1996, 29, 763–766. [Google Scholar] [CrossRef]
- Ali, J.; Najeeb, J.; Ali, M.A.; Aslam, M.F.; Raza, A. Biosensors: Their Fundamentals, Designs, Types and Most Recent Impactful Applications: A Review. J. Biosens. Bioelectron. 2017, 8, 9. [Google Scholar] [CrossRef]
- Zhang, W.; Asiri, A.M.; Liu, D.; Du, D.; Lin, Y. Nanomaterial-Based Biosensors for Environmental and Biological Monitoring of Organophosphorus Pesticides and Nerve Agents. Trends Anal. Chem. 2014, 54, 1–10. [Google Scholar] [CrossRef]
- Mirpoor, S.F.; Hosseini, S.M.H.; Nekoei, A.-R. Efficient Delivery of Quercetin After Binding to Beta-Lactoglobulin Followed by Formation Soft-Condensed Core-Shell Nanostructures. Food Chem. 2017, 233, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Loch, J.I.; Bonarek, P.; Tworzydło, M.; Łazińska, I.; Szydłowska, J.; Lipowska, J.; Rzęsikowska, K.; Lewiński, K. The Engineered Β-Lactoglobulin with Complementarity to the Chlorpromazine Chiral Conformers. Int. J. Biol. Macromol. 2018, 114, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-Atom Structure Validation for Macromolecular Crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Tan, C.; Mongan, J.; Hornak, V. Amber 9; University of California: San Francisco, CA, USA, 2006. [Google Scholar]
- Roe, D.R.; Cheatham, T.E., III. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.R., III; McGee, T.D., Jr.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA.Py: An Efficient Program for End-State Free Energy Calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–8–27–8. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Representatives, Including Isomers | Unique Representatives, Excluding Isomers | Chemical Types of Unique Representatives, n (% of Class) | Simulated Ligands * (Chemical Type) | Chemical Structures of Simulated Ligands |
|---|---|---|---|---|---|
| Ia | 22 | 6 | CO, 3 (50.0) No Type, 3 (50.0) | Bromadiolone (CO) Chlorophacinone |  |
| Ib | 12 | 5 | PY, 4 (80.0) CO, 1 (20.0) | Cypermethrin (PY) Coumatetralyl (CO) |  |
| II | 56 | 24 | No type, 12 (50.0) PY, 11 (45.8) C, 1 (4.2) | Permethrin (PY) Rotenone |  |
| III | 11 | 8 | No type, 7 (87.5) PY, 1 (12.5) | Biphenyl Cyclohexylbenzene Resmethrin (PY) |  |
| U | 17 | 17 | No type, 13 (76.5) PY, 3 (17.6) C, 1 (5.9) | Tetramethrin (PY) Fenoxycarb (C) |  |
| Total | 118 | 60 | No type, 35 (58.3) PY, 19 (31.7) CO, 4 (6.7) C, 2 (3.3) | 11 |
| Simulated Ligands | Class | Number of Histogram Peaks | Peak Maxima (Å) |
|---|---|---|---|
| Bromadiolone | Ia | 2 | 9.4 and 12.2 |
| Chlorophacinone | Ia | 2 | 10.1 and 12.6 |
| Cypermethrin | Ib | 3 | 3.9, 5.3 and 7.0 |
| Coumatetralyl | Ib | 3 | 12.9, 14.5 and 19.6 |
| Permethrin | II | 3 | 6.9, 9.2 and 11.3 |
| Rotenone | II | 2 | 6.1 and 9.3 |
| Biphenyl | III | 2 | 4.2 and 8.1 |
| Cyclohexylbenzene | III | 2 | 4.8 and 5.4 |
| Resmethrin | III | 2 | 5.1 and 6.0 |
| Tetramethrin | U | 4 | 5.0, 6.2, 9.1 and 10.9 |
| Fenoxycarb | U | 5 | 2.7, 3.7, 4.9, 6.2 and 9.1 |
| Paraquat | II | 3 | 17.0, 24.3 and 32.0 |
| Ligand | ΔΔG 1 | ΔΔG 2 | ΔΔG 3 |
|---|---|---|---|
| Paraquat | 37.82 | 38.78 | 38.78 |
| Coumatetralyl | 22.22 | 19.36 | 19.36 |
| Benzene | 21.51 | 18.39 | 18.39 |
| Biphenyl | 10.76 | 10.52 | 10.52 |
| Bromadiolone | 6.81 | 7.18 | 7.18 |
| Cyclohexylbenzene | 5.25 | 8.09 | 8.09 |
| Chlorophacinone | 3.23 | 5.96 | 5.96 |
| Rotenone | −4.91 | −3.57 | −3.57 |
| Tetramethrin # | −5.84 | −9.55 | −9.55 |
| Fenoxycarb | −9.78 | −10.48 | −10.48 |
| Permethrin # | −11.38 | −12.65 | −12.65 |
| Resmethrin # | −16.87 | −11.24 | −11.24 |
| Cypermethrin # | −19.19 | −20.46 | −20.46 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortes-Hernandez, P.; Vázquez Nuñez, R.; Domínguez-Ramírez, L. Docking and Molecular Dynamics Predictions of Pesticide Binding to the Calyx of Bovine β-Lactoglobulin. Int. J. Mol. Sci. 2020, 21, 1988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21061988
Cortes-Hernandez P, Vázquez Nuñez R, Domínguez-Ramírez L. Docking and Molecular Dynamics Predictions of Pesticide Binding to the Calyx of Bovine β-Lactoglobulin. International Journal of Molecular Sciences. 2020; 21(6):1988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21061988
Chicago/Turabian StyleCortes-Hernandez, Paulina, Roberto Vázquez Nuñez, and Lenin Domínguez-Ramírez. 2020. "Docking and Molecular Dynamics Predictions of Pesticide Binding to the Calyx of Bovine β-Lactoglobulin" International Journal of Molecular Sciences 21, no. 6: 1988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21061988