The Anti-Inflammatory and Antioxidant Effects of Sodium Propionate

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. In Vitro Studies
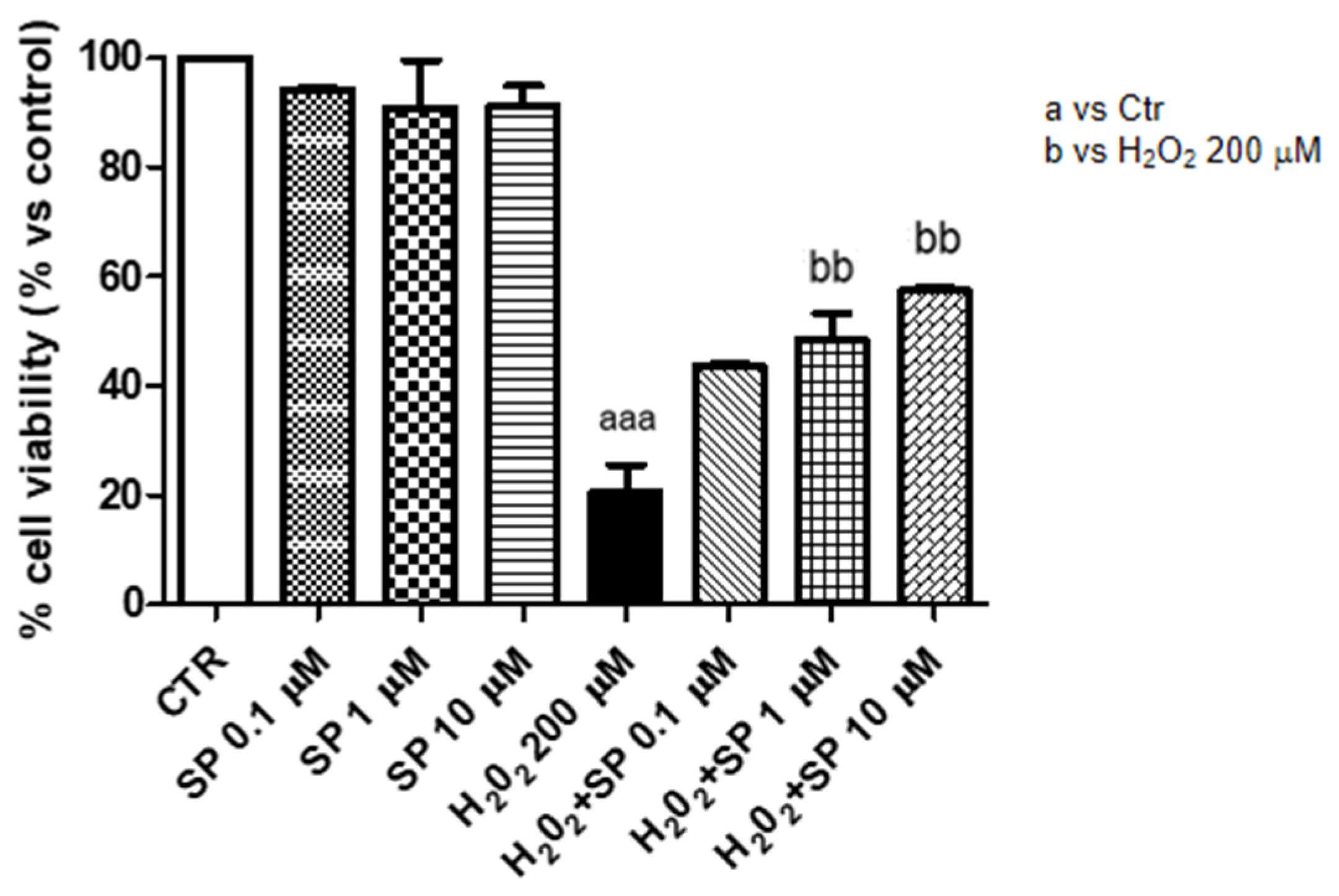
2.1.1. Effect of SP on Vitality Following LPS Stimulation
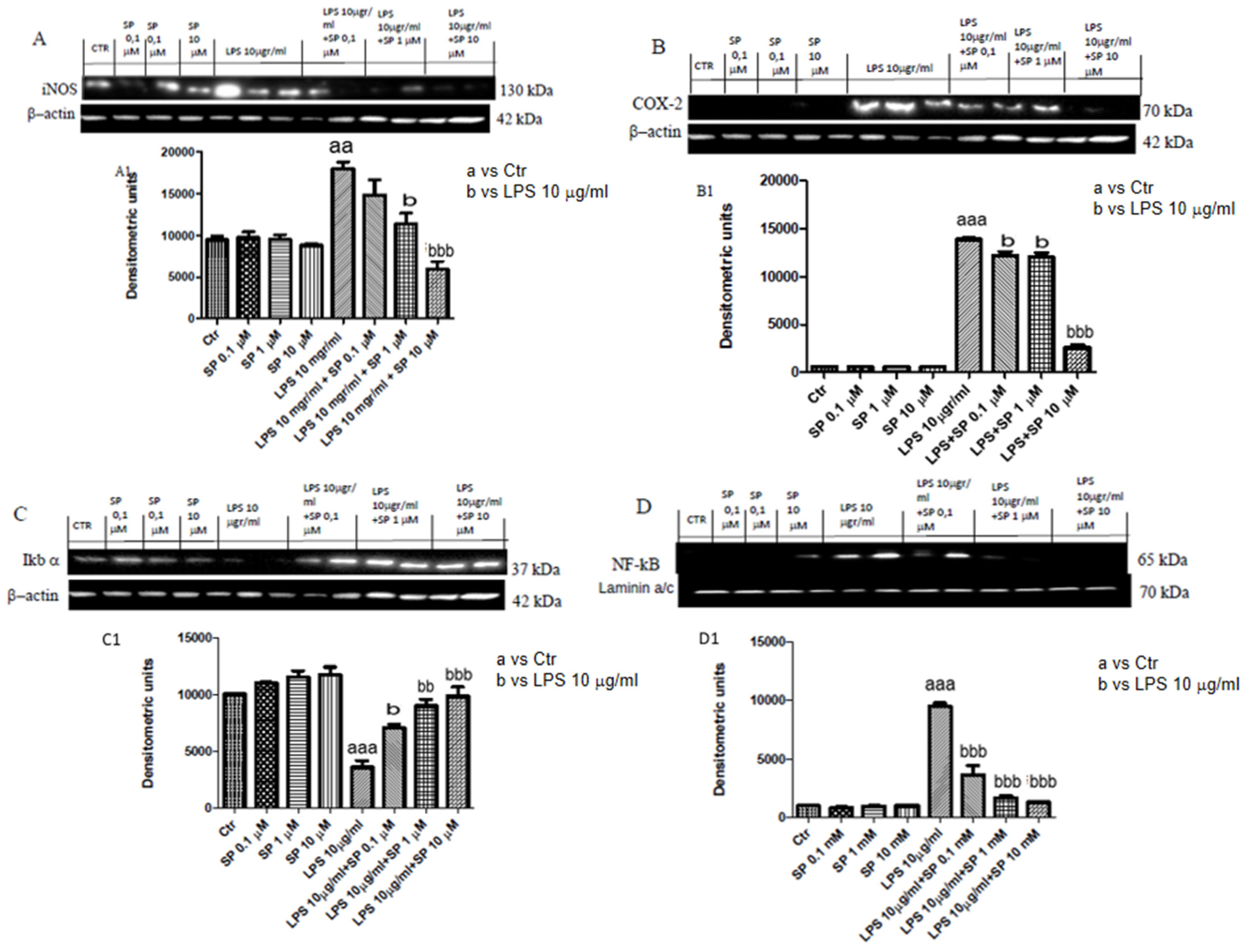
2.1.2. Effect of SP on the Expression of iNOS and COX-2 Following LPS Stimulation
2.1.3. Effect of SP on the Expression of IκBα and NF-κB following LPS Stimulation
2.1.4. Anti-Oxidant Effect of SP in J774-A1 Cell Cultures Stimulated with H2O2
2.1.5. SP Reduces the Nitrite Production and MDA Level in J774-A1
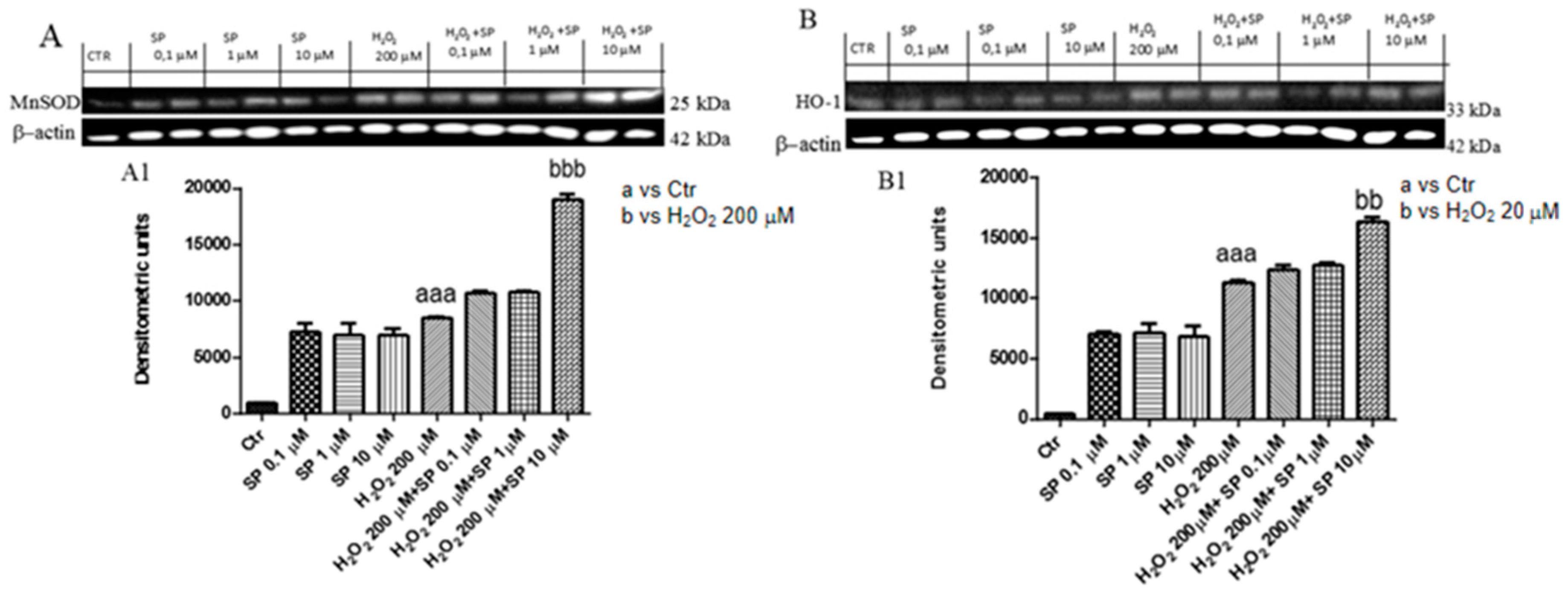
2.1.6. Effect of SP on Anti-Oxidant Enzymes In Vitro
2.2. In Vivo Studies
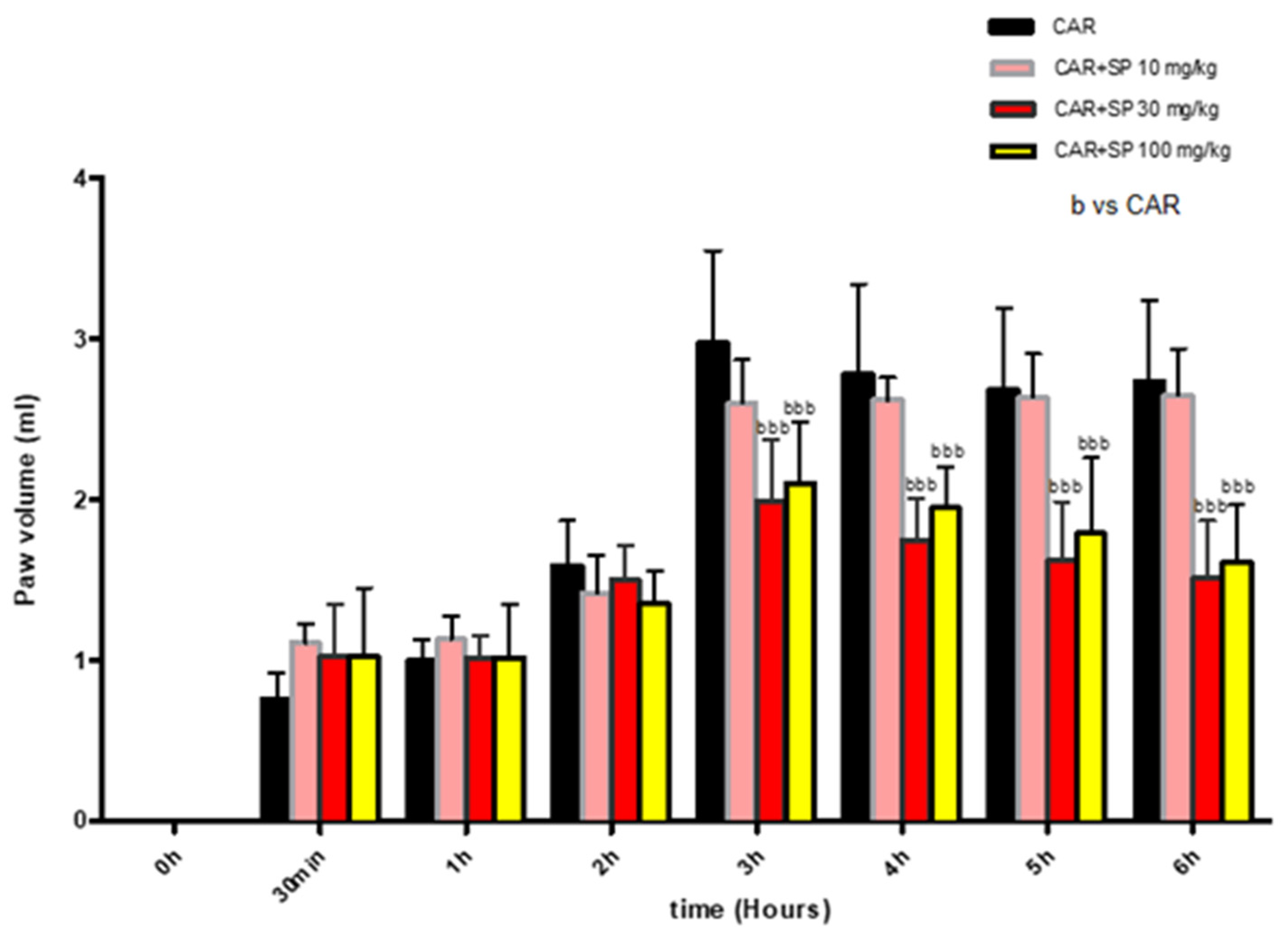
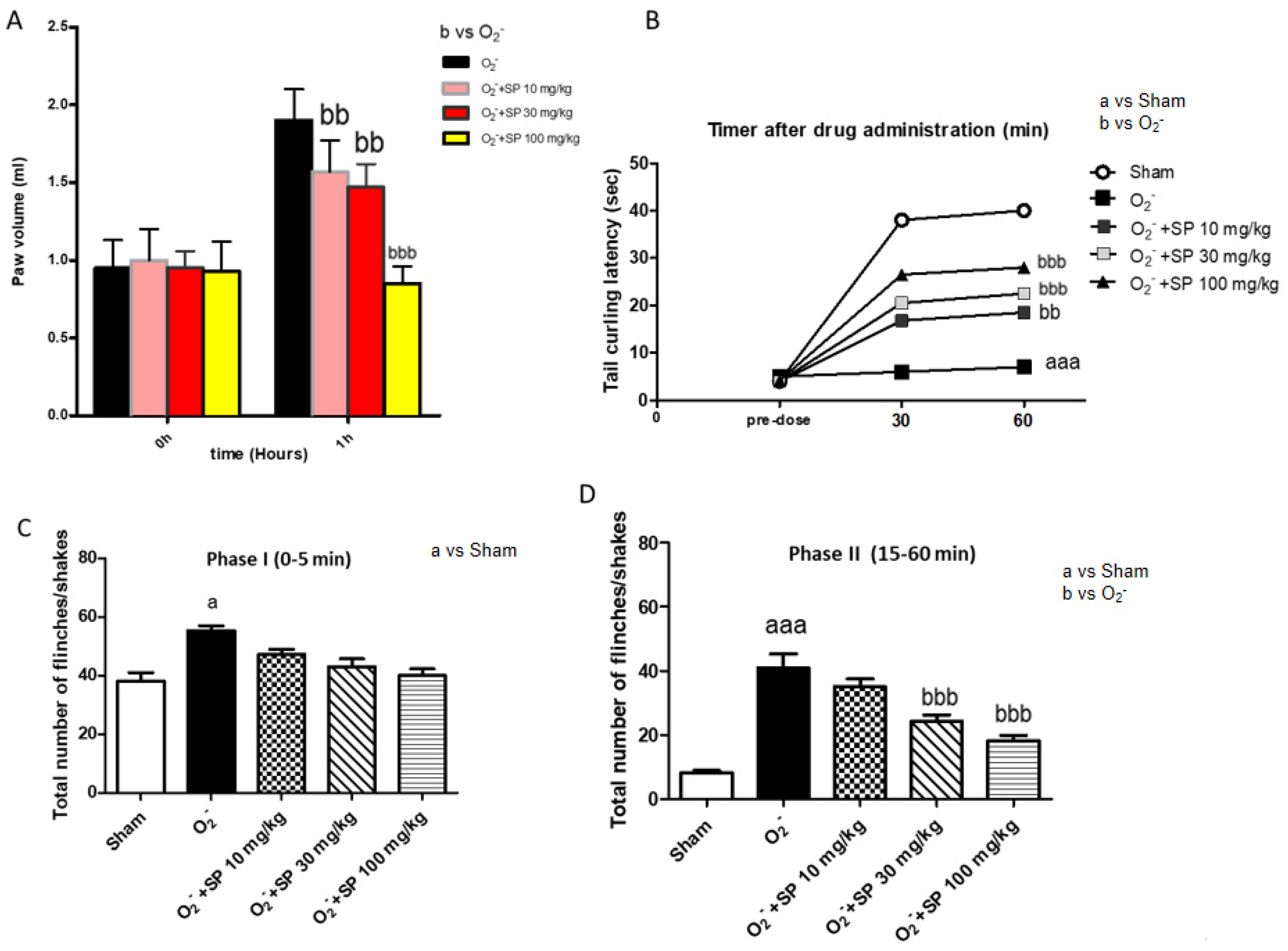
2.2.1. Effect of SP on Time-Course of CAR-Induced Paw Edema in Rats
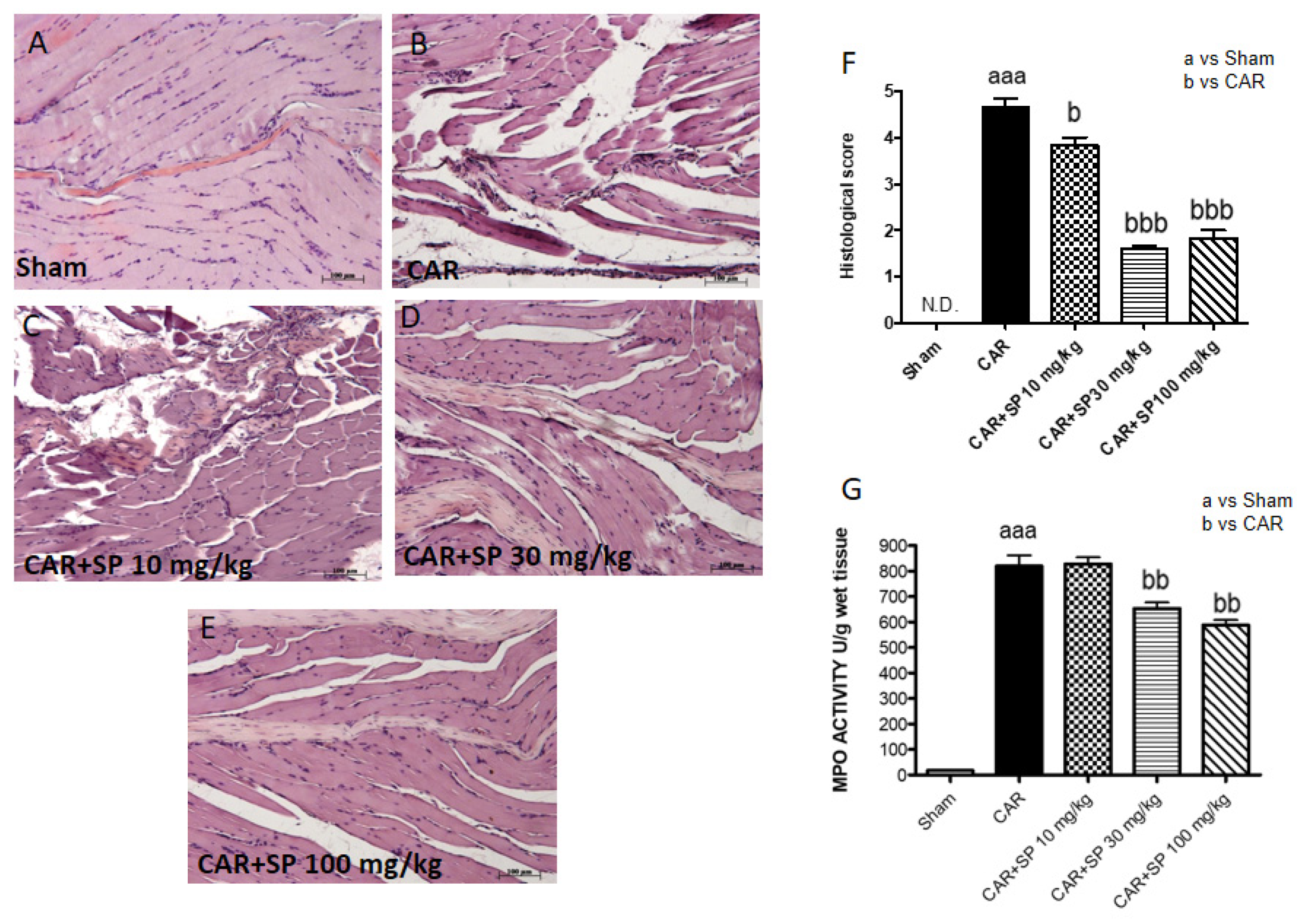
2.2.2. Histological Analyses of Paw Tissues and MPO Activity in CAR-Treated Rats
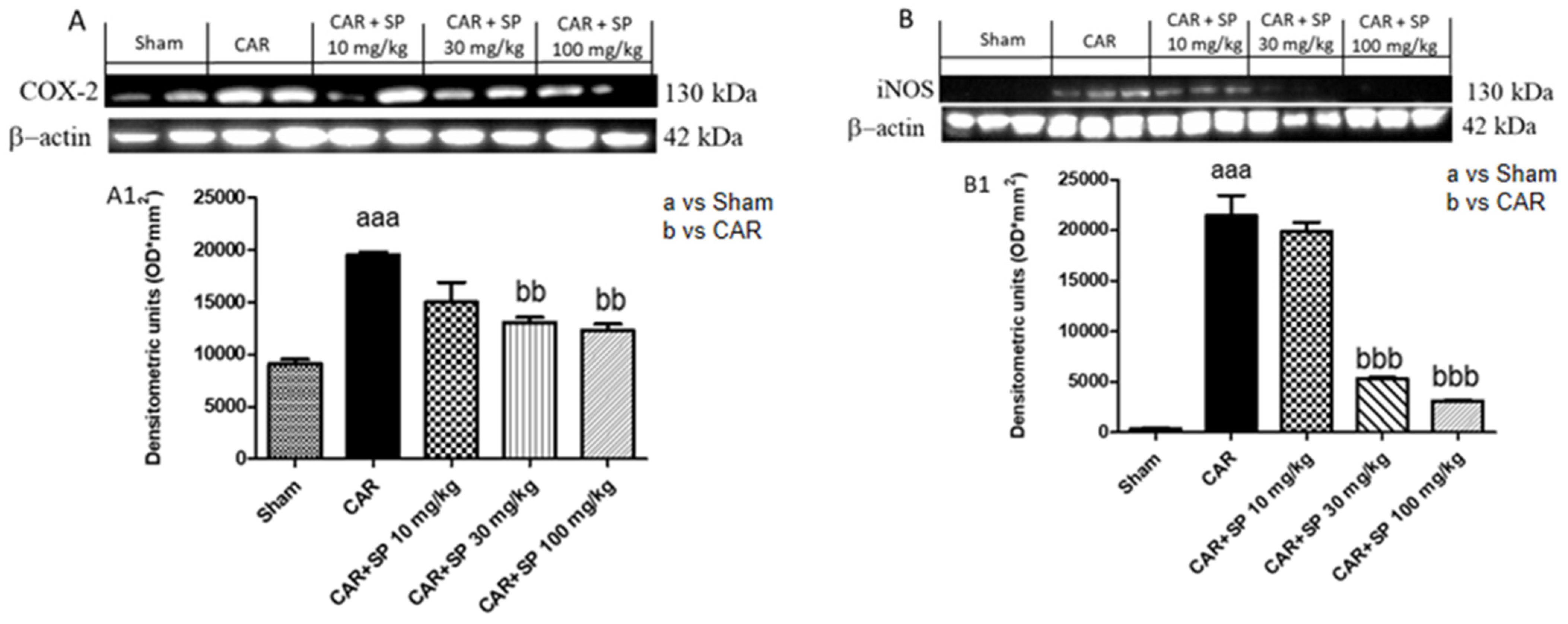
2.2.3. Effect of SP on the Expression of iNOS and COX-2 in Hind Paw Tissue from CAR-Treated Rats
2.2.4. Effect of SP on Time-Course of O2− Anion-Induced Inflammatory Pain
2.2.5. The Nociceptive Response Following O2− Intraplantar Injection
2.2.6. Analgesic Effect of SP Evaluated by Formalin Test
2.2.7. Histological Evaluation of Paw Tissues and MPO Activity following O2− Intraplantar Injection
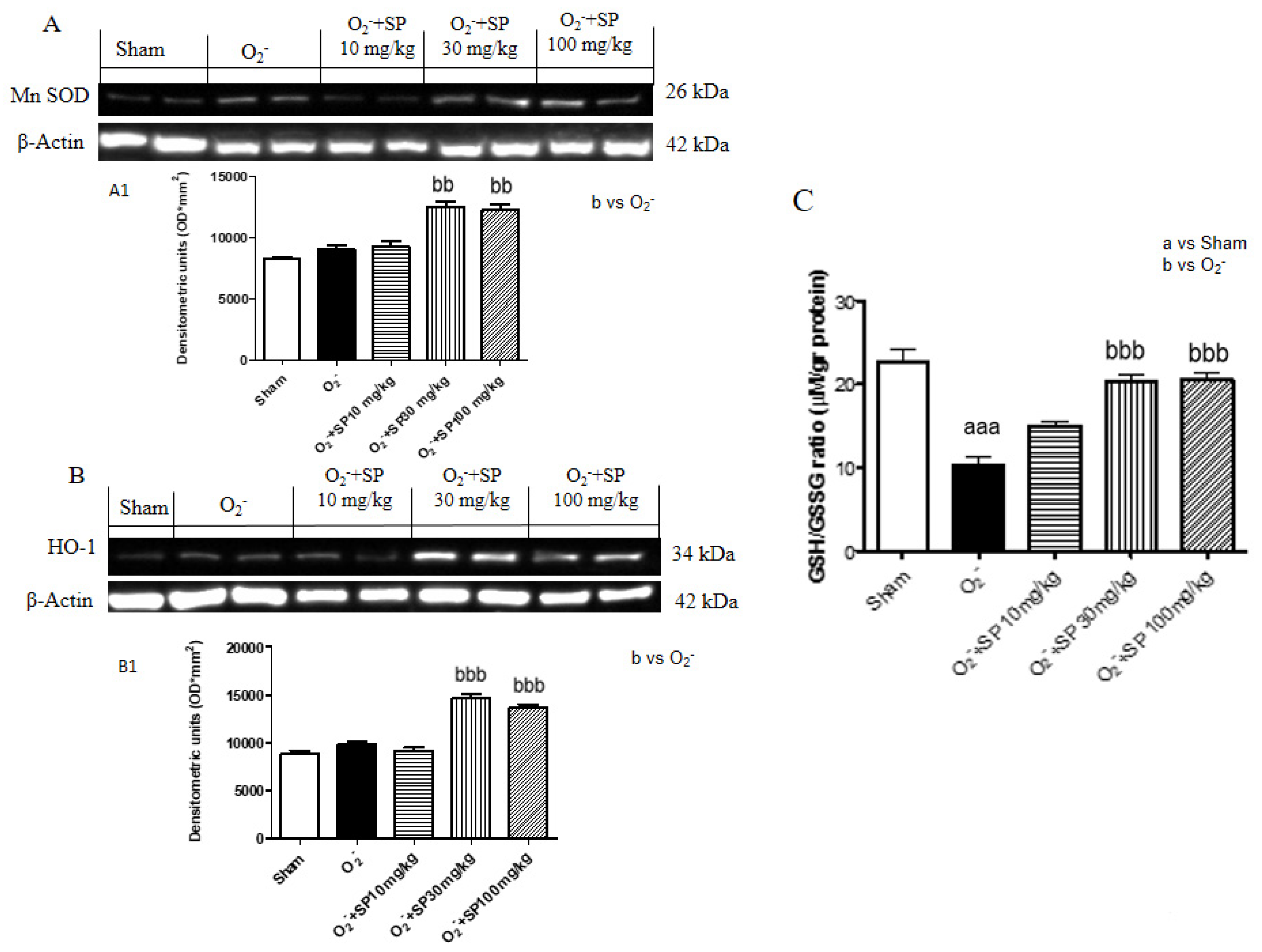
2.2.8. SP Reduce O2− Induced Oxidative Stress
2.2.9. SP Reduce GSH Levels in Rat Paws Following O2− Intraplantar Injection
3. Discussion
4. Materials and Methods
4.1. In Vitro Studies
4.1.1. Murine Macrophage Cell Cultures and Treatments
4.1.2. Western Blot Analysis
4.1.3. NOX Assay
4.1.4. Malondialdehyde (MDA) Assay
4.1.5. Data Analysis
4.2. In Vivo Studies
4.2.1. Animals
4.2.2. Carrageenan-Induced Paw Edema
4.2.3. Induction of edema by O2−
4.2.4. Experimental Groups
4.2.5. Paw Edema Measurement
4.2.6. Behavioral Tests
Tail-Flick Test
Formalin Test
4.2.7. Myeloperoxidase Activity (MPO Activity)
4.2.8. Histological Examination of the CAR-Inflamed Hind Paw
4.2.9. Histological Examination of the O2−-Inflamed Hind Paw
4.2.10. Western Blot Analysis for COX-2, iNOS, MnSOD and HO-1
4.2.11. Glutathione Assay (GSH Assay)
4.2.12. Statistical Evaluation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SP | sodium propionate |
| SCFAs | Short Chain Fatty Acids |
| LPS | lipopolisaccharide from E. coli |
| H2O2 | hydrogen peroxide |
| COX-2 | cicloxigenase-2 |
| iNOS | inducible nitric oxide synthase |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NO | nitric oxide |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DMSO | dimethyl sulfoxide |
| IκBα | nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor |
| MPO | myeloperoxidase |
| KO2 | potassium peroxide |
| GSH | Glutathione |
| MnSOD | manganese superoxide dismutase |
| CAR | carrageenan |
| MDA | Malondialdehyde |
References
- Venegas, D.P.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera4 RDijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, L.-C.; Wang, Y.; Wang, Z.-B.; Liu, W.-Y.; Sun, S.; Li, L.; Su, D.-F.; Zhang, L.-C. Propionate Ameliorates Dextran Sodium Sulfate-Induced Colitis by Improving Intestinal Barrier Function and Reducing Inflammation and Oxidative Stress. Front. Pharmacol. 2016, 7, 253. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.R.; Rodrigues, H.G.; Hatanaka, E.; Sato, F.T.; Sampaio, S.C.; Curi, R. Suppressive effect of short-chain fatty acids on production of proinflammatory mediators by neutrophils. J. Nutr. Biochem. 2011, 22, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2017, 17, 1519–1528. [Google Scholar] [CrossRef]
- Inan, M.S.; Rasoulpour, R.J.; Yin, L.; Hubbard, A.K.; Rosenberg, D.W.; Giardina, C. The luminal short-chain fatty acid butyrate modulates NF-kappaB activity in a human colonic epithelial cell line. Gastroenterology 2000, 118, 724–734. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.M.; Bast, A.; Vanhoutvin, S.A.; Fischer, M.A.; Kodde, A.; Troost, F.J.; Venema, K.; Brummer, R.J. Butyrate modulates oxidative stress in the colonic mucosa of healthy humans. Clin. Nutr. 2009, 28, 88–93. [Google Scholar] [CrossRef]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [Green Version]
- Grace, P.M.; Gaudet, A.; Staikopoulos, V.; Maier, S.F.; Hutchinson, M.R.; Salvemini, D.; Watkins, L.R. Nitroxidative Signaling Mechanisms in Pathological Pain. Trends Neurosci. 2016, 39, 862–879. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid. Med. Cell. Longev. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef] [Green Version]
- Besten, G.D.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippone, A.; Lanza, M.; Campolo, M.; Casili, G.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Protective effect of sodium propionate in Aβ1-42 -induced neurotoxicity and spinal cord trauma. Neuropharmacology 2020, 166, 107977. [Google Scholar] [CrossRef] [PubMed]
- Segain, J.P.; de la Bletiere, D.R.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottiere, H.M.; Galmiche, J.P. Butyrate inhibits inflammatory responses through NF kappa B inhibition: Implications for Crohn’s disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheeba, M.; Asha, V. Cardiospermum halicacabum ethanol extract inhibits LPS induced COX-2, TNF-α and iNOS expression, which is mediated by NF-κB regulation, in RAW264.7 cells. J. Ethnopharmacol. 2009, 124, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.-P.-P.; Rahman, H.S. Antioxidant and Oxidative Stress: A Mutual Interplay in Age-Related Diseases. Front. Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhou, T.; Ziegler, A.C.; Dimitrion, P.; Zuo, L. Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications. Oxidative Med. Cell. Longev. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lugrin, J.; Rosenblatt-Velin, N.; Parapanov, R.; Liaudet, L. The role of oxidative stress during inflammatory processes. Boil. Chem. 2014, 395, 203–230. [Google Scholar] [CrossRef] [Green Version]
- Fehrenbacher, J.C.; Vasko, M.R.; Duarte, D.B. Models of inflammation: Carrageenan- or complete Freund’s Adjuvant (CFA)-induced edema and hypersensitivity in the rat. Curr. Protoc. Pharmacol. 2012, 56. [Google Scholar] [CrossRef] [Green Version]
- Ndengele, M.M.; Cuzzocrea, S.; Esposito, E.; Mazzon, E.; Di Paola, R.; Matuschak, G.M.; Salvemini, D. Cyclooxygenases 1 and 2 contribute to peroxynitrite-mediated inflammatory pain hypersensitivity. FASEB J. 2008, 22, 3154–3164. [Google Scholar] [CrossRef] [Green Version]
- Taubert, D. Reaction rate constants of superoxide scavenging by plant antioxidants. Free. Radic. Boil. Med. 2003, 35, 1599–1607. [Google Scholar] [CrossRef]
- Choi, D.C.; Lee, J.Y.; Lim, E.J.; Baik, H.H.; Oh, T.H.; Yune, T. Inhibition of ROS-induced p38MAPK and ERK activation in microglia by acupuncture relieves neuropathic pain after spinal cord injury in rats. Exp. Neurol. 2012, 236, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Maioli, N.; Zarpelon, A.; Mizokami, S.; Calixto-Campos, C.; Guazelli, C.; Hohmann, M.; Ribeiro, F.P.; Carvalho, T.T.; Manchope, M.F.; Ferraz, C.R.; et al. The superoxide anion donor, potassium superoxide, induces pain and inflammation in mice through production of reactive oxygen species and cyclooxygenase-2. Braz. J. Med. Boil. Res. 2015, 48, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.-Q.; Porreca, F.; Cuzzocrea, S.; Galen, K.; Lightfoot, R.; Masini, E.; Muscoli, C.; Mollace, V.; Ndengele, M.; Ischiropoulos, H.; et al. A Newly Identified Role for Superoxide in Inflammatory Pain. J. Pharmacol. Exp. Ther. 2004, 309, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Toso, R.D.; Pressi, G.; Bramanti, P.; Meli, R.; Cuzzocrea, S. Protective effect of verbascoside in activated C6 glioma cells: Possible molecular mechanisms. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2009, 381, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Paterniti, I.; Cordaro, M.; Campolo, M.; Siracusa, R.; Cornelius, C.; Navarra, M.; Cuzzocrea, S.; Esposito, E. Neuroprotection by association of palmitoylethanolamide with luteolin in experimental Alzheimer’s disease models: The control of neuroinflammation. CNS Neurol. Disord. Drug Targets 2014, 13, 1530–1541. [Google Scholar] [CrossRef]
- Campolo, M.; Casili, G.; Biundo, F.; Crupi, R.; Cordaro, M.; Cuzzocrea, S.; Esposito, E. The Neuroprotective Effect of Dimethyl Fumarate in an MPTP-Mouse Model of Parkinson’s Disease:Involvement of Reactive Oxygen Species/Nuclear Factor-κB/Nuclear Transcription Factor Related to NF-E2. Antioxid. Redox. Signal. 2017, 27, 453–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elena, T.; Rosanna, D.P.; Emanuela, M.; Esposito, E.; Virginia, M.; Salvatore, C. Anti-Inflammatory Effects of Adrenomedullin on Acute Lung Injury Induced by Carrageenan in Mice. Mediat. Inflamm. 2012, 2012, 717851. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Di Paola, R.; Cordaro, M.; Gugliandolo, E.; Casili, G.; Morittu, V.M.; Britti, D.; Esposito, E.; Cuzzocrea, S. Adelmidrol, a palmitoylethanolamide analogue, as a new pharmacological treatment for the management of acute and chronic inflammation. Biochem. Pharmacol. 2016, 119, 27–41. [Google Scholar] [CrossRef]
- Salvemini, D.; Wang, Z.-Q.; Wyatt, P.S.; Bourdon, D.M.; Marino, M.H.; Manning, P.T.; Currie, M.G. Nitric oxide: A key mediator in the early and late phase of carrageenan-induced rat paw inflammation. Br. J. Pharmacol. 1996, 118, 829–838. [Google Scholar] [CrossRef] [Green Version]
- Petrosino, S.; Campolo, M.; Impellizzeri, D.; Paterniti, I.; Allarà, M.; Gugliandolo, E.; D’Amico, R.; Siracusa, R.; Cordaro, M.; Esposito, E.; et al. 2-Pentadecyl-2-Oxazoline, the Oxazoline of Pea, Modulates Carrageenan-Induced Acute Inflammation. Front. Pharmacol. 2017, 8, 308. [Google Scholar] [CrossRef] [Green Version]
- Chia, J.S.M.; Izham, N.A.M.; Farouk, A.A.O.; Sulaiman, M.R.; Mustafa, S.; Hutchinson, M.R.; Perimal, E.K. Zerumbone Modulates α2A-Adrenergic, TRPV1, and NMDA NR2B Receptors Plasticity in CCI-Induced Neuropathic Pain In Vivo and LPS-Induced SH-SY5Y Neuroblastoma In Vitro Models. Front. Pharmacol. 2020, 11, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippone, A.; Lanza, M.; Campolo, M.; Casili, G.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. The Anti-Inflammatory and Antioxidant Effects of Sodium Propionate. Int. J. Mol. Sci. 2020, 21, 3026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083026
Filippone A, Lanza M, Campolo M, Casili G, Paterniti I, Cuzzocrea S, Esposito E. The Anti-Inflammatory and Antioxidant Effects of Sodium Propionate. International Journal of Molecular Sciences. 2020; 21(8):3026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083026
Chicago/Turabian StyleFilippone, Alessia, Marika Lanza, Michela Campolo, Giovanna Casili, Irene Paterniti, Salvatore Cuzzocrea, and Emanuela Esposito. 2020. "The Anti-Inflammatory and Antioxidant Effects of Sodium Propionate" International Journal of Molecular Sciences 21, no. 8: 3026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083026