HIV-1-Infected Human Macrophages, by Secreting RANK-L, Contribute to Enhanced Osteoclast Recruitment

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
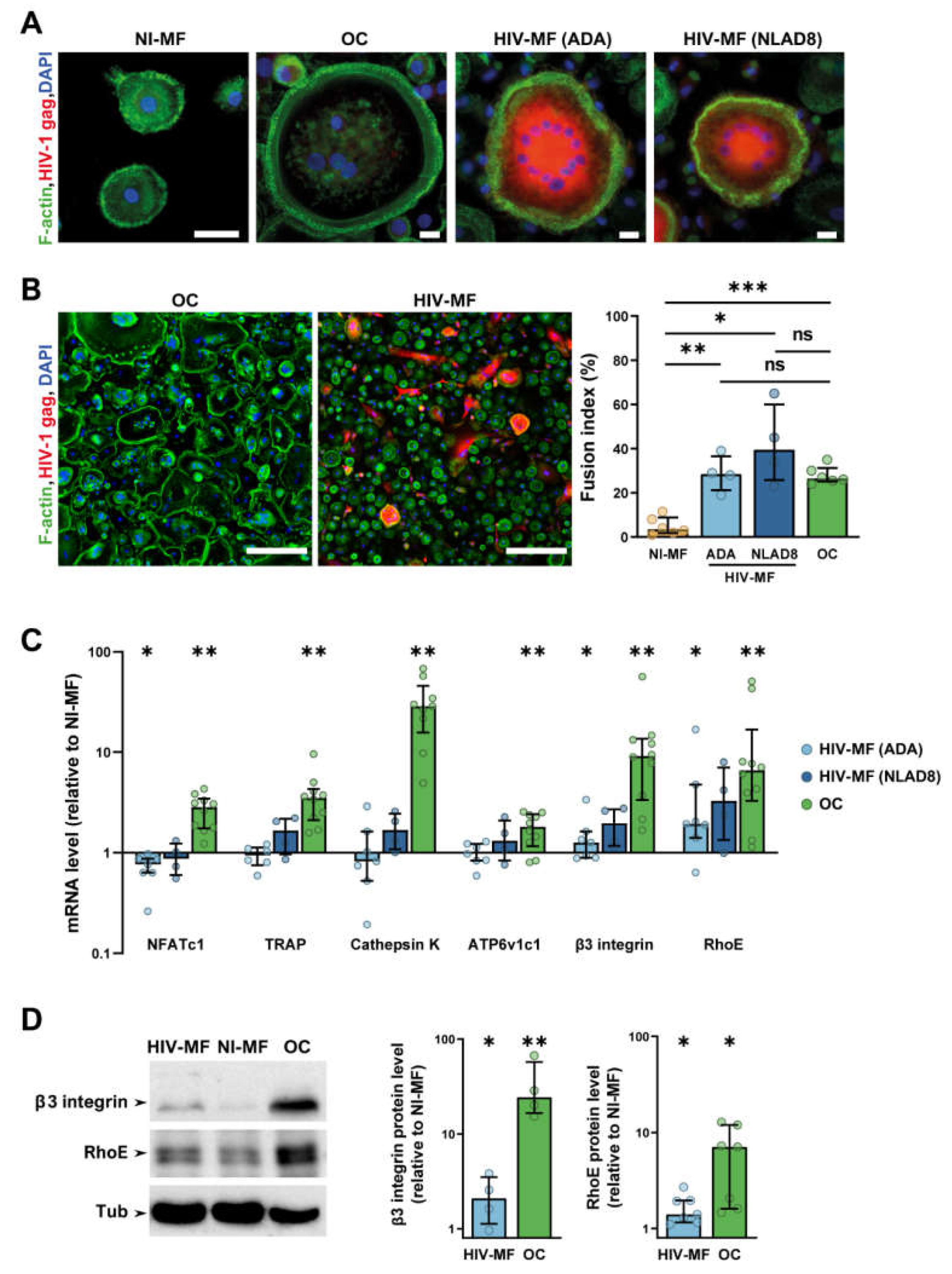
2.1. HIV-1 Infection of Macrophages Induces the Expression of a Subset of Osteoclast Markers
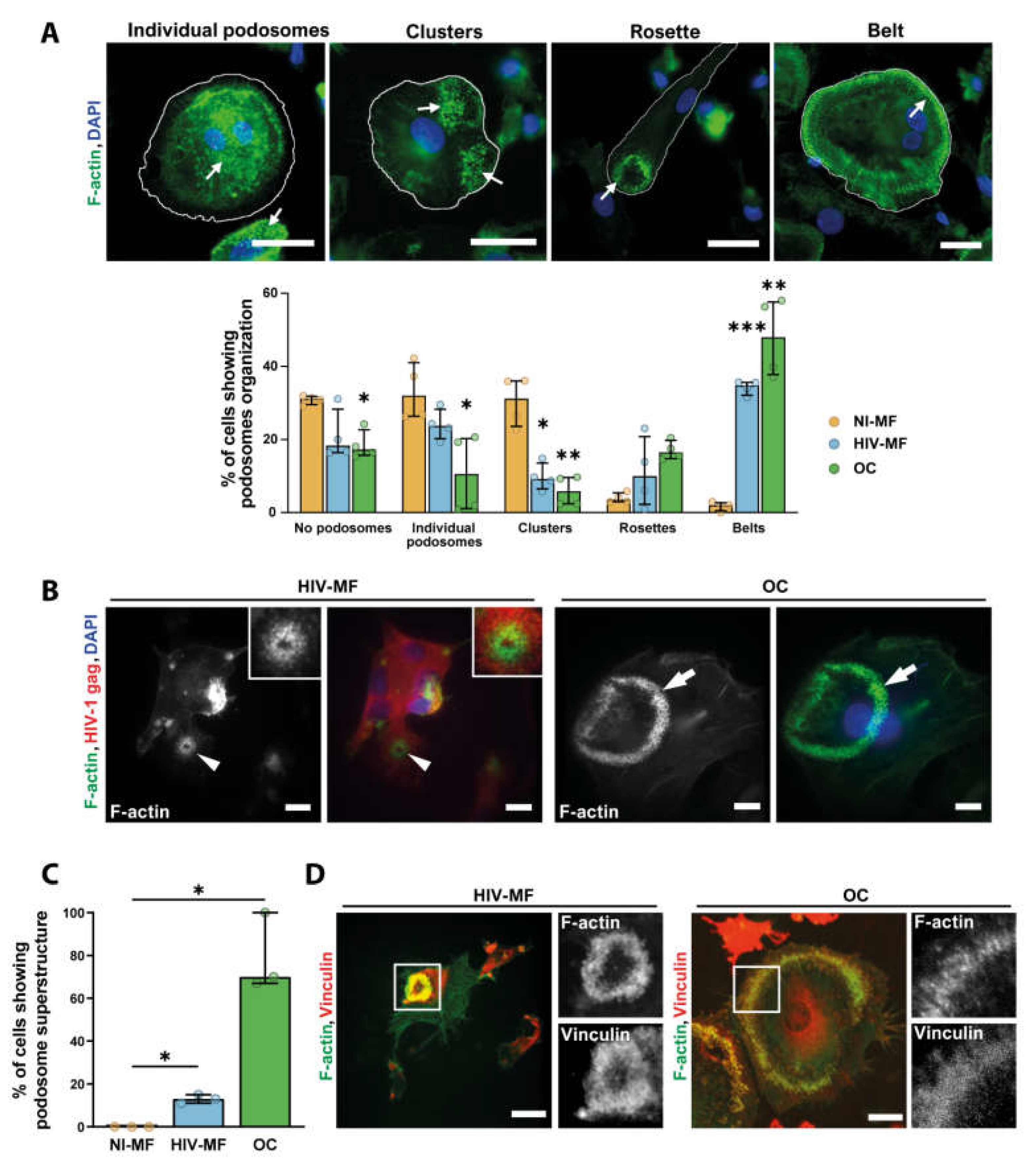
2.2. HIV-1 Infection of Macrophages Induces the Formation of Sealing Zone-Like Structures
2.3. HIV-1 Infection of Macrophages Does not Enhance Their Bone Degradation Activity
2.4. RANK-L Is Secreted by Infected Macrophages and Promotes 3D Migration of Osteoclast Precursors
3. Discussion
4. Material and Methods
4.1. Preparation of Monocyte-Derived Macrophages and Monocyte-Derived Osteoclasts
4.2. Viruses, HIV-1 Infection, and Preparation of Conditioned Media
4.3. RNA Extraction and qRT-PCR
4.4. ELISA
4.5. Immunofluorescence Microscopy
4.6. Scanning Electron Microscopy
4.7. Immunoblot Analyses
4.8. 3D Migration Assays
4.9. Bone Resorption Assays
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cotter, A.G.; Mallon, P.W. The effects of untreated and treated HIV infection on bone disease. Curr. Opin. HIV AIDS 2014, 9, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Moran, C.A.; Weitzmann, M.N.; Ofotokun, I. Bone Loss in HIV Infection. Curr. Treat. Options Infect. Dis. 2017, 9, 52–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruera, D.; Luna, N.; David, D.O.; Bergoglio, L.M.; Zamudio, J. Decreased bone mineral density in HIV-infected patients is independent of antiretroviral therapy. Aids 2003, 17, 1917–1923. [Google Scholar] [CrossRef] [PubMed]
- Gibellini, D.; Borderi, M.; De Crignis, E.; Cicola, R.; Vescini, F.; Caudarella, R.; Chiodo, F.; Re, M.C. RANK-L/OPG/TRAIL plasma levels and bone mass loss evaluation in antiretroviral naive HIV-1-positive men. J. Med. Virol. 2007, 79, 1446–1454. [Google Scholar] [CrossRef] [PubMed]
- Grijsen, M.L.; Vrouenraets, S.M.; Steingrover, R.; Lips, P.; Reiss, P.; Wit, F.W.; Prins, J.M. High prevalence of reduced bone mineral density in primary HIV-1-infected men. Aids 2010, 24, 2233–2238. [Google Scholar] [CrossRef] [Green Version]
- Titanji, K.; Vunnava, A.; Sheth, A.N.; Delille, C.; Lennox, J.L.; Sanford, S.E.; Foster, A.; Knezevic, A.; Easley, K.A.; Weitzmann, M.N.; et al. Dysregulated B cell expression of RANK-L and OPG correlates with loss of bone mineral density in HIV infection. PLoS Pathog. 2014, 10, e1004497. [Google Scholar] [CrossRef]
- Beaupere, C.; Garcia, M.; Larghero, J.; Fève, B.; Capeau, J.; Lagathu, C. The HIV proteins Tat and Nef promote human bone marrow mesenchymal stem cell senescence and alter osteoblastic differentiation. Aging Cell 2015, 14, 534–546. [Google Scholar] [CrossRef]
- Butler, J.S.; Dunning, E.C.; Murray, D.W.; Doran, P.P.; O’Byrne, J.M. HIV-1 protein induced modulation of primary human osteoblast differentiation and function via a Wnt/β-catenin-dependent mechanism. J. Orthop. Res. 2013, 31, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Gibellini, D.; De Crignis, E.; Ponti, C.; Cimatti, L.; Borderi, M.; Tschon, M.; Giardino, R.; Re, M.C. HIV-1 triggers apoptosis in primary osteoblasts and HOBIT cells through TNFalpha activation. J. Med. Virol. 2008, 80, 1507–1514. [Google Scholar] [CrossRef]
- Jacome-Galarza, C.E.; Percin, G.I.; Muller, J.T.; Mass, E.; Lazarov, T.; Eitler, J.; Rauner, M.; Yadav, V.K.; Crozet, L.; Bohm, M.; et al. Developmental origin, functional maintenance and genetic rescue of osteoclasts. Nature 2019, 568, 541–545. [Google Scholar] [CrossRef]
- Kotani, M.; Kikuta, J.; Klauschen, F.; Chino, T.; Kobayashi, Y.; Yasuda, H.; Tamai, K.; Miyawaki, A.; Kanagawa, O.; Tomura, M.; et al. Systemic circulation and bone recruitment of osteoclast precursors tracked by using fluorescent imaging techniques. J. Immunol. 2013, 190, 605–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teitelbaum, S.L. The osteoclast and its unique cytoskeleton. Ann. NY Acad. Sci. 2011, 1240, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Georgess, D.; Machuca-Gayet, I.; Blangy, A.; Jurdic, P. Podosome organization drives osteoclast-mediated bone resorption. Cell Adh. Migr. 2014, 8, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, S. RANK-L-Independent Osteoclastogenesis: A Long-Standing Controversy. J. Bone Miner. Res. 2017, 32, 431–433. [Google Scholar] [CrossRef] [Green Version]
- Wiesner, C.; Le-Cabec, V.; El Azzouzi, K.; Maridonneau-Parini, I.; Linder, S. Podosomes in space: Macrophage migration and matrix degradation in 2D and 3D settings. Cell Adh. Migr. 2014, 8, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Luxenburg, C.; Addadi, L.; Geiger, B. The molecular dynamics of osteoclast adhesions. Eur. J. Cell Biol. 2006, 85, 203–211. [Google Scholar]
- Georgess, D.; Mazzorana, M.; Terrado, J.; Delprat, C.; Chamot, C.; Guasch, R.M.; Perez-Roger, I.; Jurdic, P.; Machuca-Gayet, I. Comparative transcriptomics reveals RhoE as a novel regulator of actin dynamics in bone-resorbing osteoclasts. Mol. Biol. Cell 2014, 25, 380–396. [Google Scholar] [CrossRef]
- De Menezes, E.G.; Machado, A.A.; Barbosa, F., Jr.; de Paula, F.J.; Navarro, A.M. Bone metabolism dysfunction mediated by the increase of proinflammatory cytokines in chronic HIV infection. J. Bone Miner. Metab 2017, 35, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Ofotokun, I.; McIntosh, E.; Weitzmann, M.N. HIV: Inflammation and bone. Curr. HIV/AIDS Rep. 2012, 9, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Titanji, K.; Vunnava, A.; Foster, A.; Sheth, A.N.; Lennox, J.L.; Knezevic, A.; Shenvi, N.; Easley, K.A.; Ofotokun, I.; Weitzmann, M.N. T-cell receptor activator of nuclear factor-kappaB ligand/osteoprotegerin imbalance is associated with HIV-induced bone loss in patients with higher CD4+ T-cell counts. AIDS 2018, 32, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Vikulina, T.; Fan, X.; Yamaguchi, M.; Roser-Page, S.; Zayzafoon, M.; Guidot, D.M.; Ofotokun, I.; Weitzmann, M.N. Alterations in the immuno-skeletal interface drive bone destruction in HIV-1 transgenic rats. Proc. Natl. Acad. Sci. USA 2010, 107, 13848–13853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gohda, J.; Ma, Y.; Huang, Y.; Zhang, Y.; Gu, L.; Han, Y.; Li, T.; Gao, B.; Gao, G.F.; Inoue, J.; et al. HIV-1 replicates in human osteoclasts and enhances their differentiation in vitro. Retrovirology 2015, 12, 12. [Google Scholar] [CrossRef] [Green Version]
- Ofotokun, I.; Titanji, K.; Vikulina, T.; Roser-Page, S.; Yamaguchi, M.; Zayzafoon, M.; Williams, I.R.; Weitzmann, M.N. Role of T-cell reconstitution in HIV-1 antiretroviral therapy-induced bone loss. Nat. Commun. 2015, 6, 8282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raynaud-Messina, B.; Bracq, L.; Dupont, M.; Souriant, S.; Usmani, S.M.; Proag, A.; Pingris, K.; Soldan, V.; Thibault, C.; Capilla, F.; et al. Bone degradation machinery of osteoclasts: An HIV-1 target that contributes to bone loss. Proc. Natl. Acad. Sci. USA 2018, 115, E2556–E2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boliar, S.; Gludish, D.W.; Jambo, K.C.; Kamng’ona, R.; Mvaya, L.; Mwandumba, H.C.; Russell, D.G. Inhibition of the lncRNA SAF drives activation of apoptotic effector caspases in HIV-1-infected human macrophages. Proc. Natl. Acad. Sci. USA 2019, 116, 7431–7438. [Google Scholar] [CrossRef] [Green Version]
- Clayton, K.L.; Collins, D.R.; Lengieza, J.; Ghebremichael, M.; Dotiwala, F.; Lieberman, J.; Walker, B.D. Resistance of HIV-infected macrophages to CD8(+) T lymphocyte-mediated killing drives activation of the immune system. Nat. Immunol. 2018, 19, 475–486. [Google Scholar] [CrossRef]
- Ganor, Y.; Real, F.; Sennepin, A.; Dutertre, C.A.; Prevedel, L.; Xu, L.; Tudor, D.; Charmeteau, B.; Couedel-Courteille, A.; Marion, S.; et al. HIV-1 reservoirs in urethral macrophages of patients under suppressive antiretroviral therapy. Nat. Microbiol. 2019, 4, 633–644. [Google Scholar] [CrossRef]
- Rodrigues, V.; Ruffin, N.; San-Roman, M.; Benaroch, P. Myeloid Cell Interaction with HIV: A Complex Relationship. Front. Immunol. 2017, 8, 1698. [Google Scholar] [CrossRef] [Green Version]
- Vérollet, C.; Souriant, S.; Bonnaud, E.; Jolicoeur, P.; Raynaud-Messina, B.; Kinnaer, C.; Fourquaux, I.; Imle, A.; Benichou, S.; Fackler, O.T.; et al. HIV-1 reprograms the migration of macrophages. Blood 2015, 125, 1611–1622. [Google Scholar] [CrossRef]
- Sattentau, Q.J.; Stevenson, M. Macrophages and HIV-1: An Unhealthy Constellation. Cell Host Microbe 2016, 19, 304–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batoon, L.; Millard, S.M.; Raggatt, L.J.; Pettit, A.R. Osteomacs and Bone Regeneration. Curr. Osteoporos Rep. 2017, 15, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Batoon, L.; Millard, S.M.; Wullschleger, M.E.; Preda, C.; Wu, A.C.; Kaur, S.; Tseng, H.W.; Hume, D.A.; Levesque, J.P.; Raggatt, L.J.; et al. CD169(+) macrophages are critical for osteoblast maintenance and promote intramembranous and endochondral ossification during bone repair. Biomaterials 2019, 196, 51–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozec, A.; Soulat, D. Latest perspectives on macrophages in bone homeostasis. Pflugers Arch. 2017, 469, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Culemann, S.; Grüneboom, A.; Nicolás-Ávila, J.Á.; Weidner, D.; Lämmle, K.F.; Rothe, T.; Quintana, J.A.; Kirchner, P.; Krljanac, B.; Eberhardt, M.; et al. Locally renewing resident synovial macrophages provide a protective barrier for the joint. Nature 2019, 572, 670–675. [Google Scholar] [CrossRef]
- Rivollier, A.; Mazzorana, M.; Tebib, J.; Piperno, M.; Aitsiselmi, T.; Rabourdin-Combe, C.; Jurdic, P.; Servet-Delprat, C. Immature dendritic cell transdifferentiation into osteoclasts: A novel pathway sustained by the rheumatoid arthritis microenvironment. Blood 2004, 104, 4029–4037. [Google Scholar] [CrossRef]
- Wakkach, A.; Mansour, A.; Dacquin, R.; Coste, E.; Jurdic, P.; Carle, G.F.; Blin-Wakkach, C. Bone marrow microenvironment controls the in vivo differentiation of murine dendritic cells into osteoclasts. Blood 2008, 112, 5074–5083. [Google Scholar] [CrossRef]
- Laperine, O.; Blin-Wakkach, C.; Guicheux, J.; Beck-Cormier, S.; Lesclous, P. Dendritic-cell-derived osteoclasts: A new game changer in bone-resorption-associated diseases. Drug Discov. Today 2016, 21, 1345–1354. [Google Scholar] [CrossRef]
- Vérollet, C.; Zhang, Y.M.; Le Cabec, V.; Mazzolini, J.; Charrière, G.; Labrousse, A.; Bouchet, J.; Medina, I.; Biessen, E.; Niedergang, F.; et al. HIV-1 Nef triggers macrophage fusion in a p61Hck- and protease-dependent manner. J. Immunol. 2010, 184, 7030–7039. [Google Scholar] [CrossRef] [Green Version]
- Kadiu, I.; Gendelman, H.E. Human immunodeficiency virus type 1 endocytic trafficking through macrophage bridging conduits facilitates spread of infection. J. Neuroimmune Pharmacol. 2011, 6, 658–675. [Google Scholar] [CrossRef] [Green Version]
- Baxter, A.E.; Russell, R.A.; Duncan, C.J.; Moore, M.D.; Willberg, C.B.; Pablos, J.L.; Finzi, A.; Kaufmann, D.E.; Ochsenbauer, C.; Kappes, J.C.; et al. Macrophage Infection via Selective Capture of HIV-1-Infected CD4(+) T Cells. Cell Host Microbe 2014, 16, 711–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurdic, P.; Saltel, F.; Chabadel, A.; Destaing, O. Podosome and sealing zone: Specificity of the osteoclast model. Eur. J. Cell Biol. 2006, 85, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Soe, K.; Delaisse, J.M. Time-lapse reveals that osteoclasts can move across the bone surface while resorbing. J. Cell Sci. 2017, 130, 2026–2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blavier, L.; Delaisse, J.M. Matrix metalloproteinases are obligatory for the migration of preosteoclasts to the developing marrow cavity of primitive long bones. J. Cell Sci. 1995, 108 Pt 12, 3649–3659. [Google Scholar]
- Vérollet, C.; Gallois, A.; Dacquin, R.; Lastrucci, C.; Pandruvada, S.N.; Ortega, N.; Poincloux, R.; Behar, A.; Cougoule, C.; Lowell, C.; et al. Hck contributes to bone homeostasis by controlling the recruitment of osteoclast precursors. FASEB J. 2013, 27, 3608–3618. [Google Scholar] [CrossRef] [Green Version]
- Gui, P.; Ben-Neji, M.; Belozertseva, E.; Dalenc, F.; Franchet, C.; Gilhodes, J.; Labrousse, A.; Bellard, E.; Golzio, M.; Poincloux, R.; et al. The Protease-Dependent Mesenchymal Migration of Tumor-Associated Macrophages as a Target in Cancer Immunotherapy. Cancer Immunol. Res. 2018, 6, 1337–1351. [Google Scholar] [CrossRef] [Green Version]
- Van Goethem, E.; Poincloux, R.; Gauffre, F.; Maridonneau-Parini, I.; Le Cabec, V. Matrix Architecture Dictates Three-Dimensional Migration Modes of Human Macrophages: Differential Involvement of Proteases and Podosome-Like Structures. J. Immunol. 2010, 184, 1049–1061. [Google Scholar] [CrossRef]
- Michalski, M.N.; McCauley, L.K. Macrophages and skeletal health. Pharmacol. Ther. 2017, 174, 43–54. [Google Scholar] [CrossRef]
- Pettit, A.R.; Chang, M.K.; Hume, D.A.; Raggatt, L.J. Osteal macrophages: A new twist on coupling during bone dynamics. Bone 2008, 43, 976–982. [Google Scholar] [CrossRef]
- Cougoule, C.; Carreno, S.; Castandet, J.; Labrousse, A.; Astarie-Dequeker, C.; Poincloux, R.; Le Cabec, V.; Maridonneau-Parini, I. Activation of the lysosome-associated p61Hck isoform triggers the biogenesis of podosomes. Traffic 2005, 6, 682–694. [Google Scholar] [CrossRef]
- Destaing, O.; Sanjay, A.; Itzstein, C.; Horne, W.C.; Toomre, D.; De Camilli, P.; Baron, R. The tyrosine kinase activity of c-Src regulates actin dynamics and organization of podosomes in osteoclasts. Mol. Biol. Cell 2008, 19, 394–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luxenburg, C.; Parsons, J.T.; Addadi, L.; Geiger, B. Involvement of the Src-cortactin pathway in podosome formation and turnover during polarization of cultured osteoclasts. J. Cell Sci. 2006, 119, 4878–4888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayanagi, H. The role of NFAT in osteoclast formation. Ann. NY Acad. Sci. 2007, 1116, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANK-L signaling in terminal differentiation of osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Matsuyama, T.; Hosokawa, Y.; Makihira, S.; Seki, M.; Karimbux, N.Y.; Goncalves, R.B.; Valverde, P.; Dibart, S.; Li, Y.P.; et al. B and T lymphocytes are the primary sources of RANK-L in the bone resorptive lesion of periodontal disease. Am. J. Pathol. 2006, 169, 987–998. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Wu, W.; Sun, W.; Zhang, Q.; Yan, F.; Xiao, Y. RANK-L expression in periodontal disease: Where does RANK-L come from? Biomed. Res. Int. 2014, 2014, 731039. [Google Scholar]
- Liu, D.; Xu, J.K.; Figliomeni, L.; Huang, L.; Pavlos, N.J.; Rogers, M.; Tan, A.; Price, P.; Zheng, M.H. Expression of RANK-L and OPG mRNA in periodontal disease: Possible involvement in bone destruction. Int. J. Mol. Med. 2003, 11, 17–21. [Google Scholar]
- Hikita, A.; Tanaka, S. Ectodomain shedding of receptor activator of NF-kappaB ligand. Adv. Exp. Med. Biol. 2007, 602, 15–21. [Google Scholar]
- Xu, Y.; Kulkosky, J.; Acheampong, E.; Nunnari, G.; Sullivan, J.; Pomerantz, R.J. HIV-1-mediated apoptosis of neuronal cells: Proximal molecular mechanisms of HIV-1-induced encephalopathy. Proc. Natl. Acad. Sci. USA 2004, 101, 7070–7075. [Google Scholar] [CrossRef] [Green Version]
- Souriant, S.; Balboa, L.; Dupont, M.; Pingris, K.; Kviatcovsky, D.; Cougoule, C.; Lastrucci, C.; Bah, A.; Gasser, R.; Poincloux, R.; et al. Tuberculosis Exacerbates HIV-1 Infection through IL-10/STAT3-Dependent Tunneling Nanotube Formation in Macrophages. Cell Rep. 2019, 26, 3586–3599 e3587. [Google Scholar] [CrossRef] [Green Version]
- Bouissou, A.; Proag, A.; Bourg, N.; Pingris, K.; Cabriel, C.; Balor, S.; Mangeat, T.; Thibault, C.; Vieu, C.; Dupuis, G.; et al. Podosome Force Generation Machinery: A Local Balance between Protrusion at the Core and Traction at the Ring. ACS Nano 2017, 11, 4028–4040. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mascarau, R.; Bertrand, F.; Labrousse, A.; Gennero, I.; Poincloux, R.; Maridonneau-Parini, I.; Raynaud-Messina, B.; Vérollet, C. HIV-1-Infected Human Macrophages, by Secreting RANK-L, Contribute to Enhanced Osteoclast Recruitment. Int. J. Mol. Sci. 2020, 21, 3154. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093154
Mascarau R, Bertrand F, Labrousse A, Gennero I, Poincloux R, Maridonneau-Parini I, Raynaud-Messina B, Vérollet C. HIV-1-Infected Human Macrophages, by Secreting RANK-L, Contribute to Enhanced Osteoclast Recruitment. International Journal of Molecular Sciences. 2020; 21(9):3154. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093154
Chicago/Turabian StyleMascarau, Rémi, Florent Bertrand, Arnaud Labrousse, Isabelle Gennero, Renaud Poincloux, Isabelle Maridonneau-Parini, Brigitte Raynaud-Messina, and Christel Vérollet. 2020. "HIV-1-Infected Human Macrophages, by Secreting RANK-L, Contribute to Enhanced Osteoclast Recruitment" International Journal of Molecular Sciences 21, no. 9: 3154. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093154