Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications

 , , and
, , and
Abstract
:1. Obesity: Definition
2. Epidemiology of Obesity
3. Adipose Tissue
4. Adipokines and Metabolic Disorders
5. Obesity, Cytokines and Inflammation
6. Obesity, Adipokines and Chronic Complications
7. Obesity, Adipokines and Psoriasis
8. Obesity, Adipokines and Diabetic Foot
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| WHO | World Health Organization |
| WOF | World Obesity Federation |
| ICD-11 | International Classification of Diseases, Eleventh Revision |
| WAT | White adipose tissue |
| BAT | Brown adipose tissue |
| BMI | Body mass index |
| VAI | Visceral adiposity index |
| T2DM | Type 2 diabetes mellitus |
| T1DM | Type 1 diabetes mellitus |
| CAD | Coronary artery disease |
| IDF | International Diabetes Federation |
| PCOS | Polycystic ovary syndrome |
| CRP | C-reactive protein |
| hsCRP | high sensitivity C-reactive protein |
| L-LR | Leptin–leptin receptor |
| IL1β | Interleukin 1β |
| TNFα | Tumour necrosis factor alpha |
| IL6 | Interleukin 6 |
| IL34 | Interleukin 34 |
| NF-Κb | Nuclear factor-κB |
| UCP-1 | Uncoupling protein-1 |
| PDR | Proliferative diabetic retinopathy |
References
- World Health Organization. Obesity: Preventing and Managing the Global Epidemic: Report of a WHO Consultation on Obesity; World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Bray, G.; Kim, K.; Wilding, J.P.H.; on behalf of the World Obesity Federation. Obesity: A chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ICD-11-Mortality and Morbidity Statistics. Available online: https://icd.who.int/browse11/l-m/en (accessed on 30 April 2020).
- Bozeman, S.R.; Hoaglin, D.C.; Burton, T.; Pashos, C.; Ben-Joseph, R.H.; Hollenbeak, C.S. Predicting waist circumference from body mass index. BMC Med. Res. Methodol. 2012, 12, 115. [Google Scholar] [CrossRef] [Green Version]
- Patry-Parisien, J.; Shields, M.; Bryan, S. Comparison of waist circumference using the World Health Organization and National Institutes of Health protocols. Health Rep. 2012, 23, 10. [Google Scholar]
- Bermúdez, V.; Rojas, M.; Salazar, J.; Añez, R.; Toledo, A.; Bello, L.; Apruzzese, V.; González, R.; Chacín, M.; Cabrera, M.; et al. Sensitivity and Specificity Improvement in Abdominal Obesity Diagnosis Using Cluster Analysis during Waist Circumference Cut-Off Point Selection. J. Diabetes Res. 2015, 2015, 750265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalvøy, H.; Sauter, A. Detection of intraneural needle-placement with multiple frequency bioimpedance monitoring: A novel method. J. Clin. Monit. 2015, 30, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, A.; Judd, S.E.; Allison, D.; Oza-Frank, R.; Affuso, O.; Safford, M.M.; Howard, V.J.; Howard, G. The geographic distribution of obesity in the US and the potential regional differences in misreporting of obesity. Obesity 2013, 22, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drewa, A.; Zorena, K. Prevention of overweight and obesity in children and adolescents in European countries. Pediatr. Endocrinol. Diabetes Metab. 2017, 23, 152–158. [Google Scholar] [CrossRef]
- Spinelli, A.; Buoncristiano, M.; Kovacs, V.A.; Yngve, A.; Spiroski, I.; Obreja, G.; Starc, G.; Pérez, N.; Rito, A.; Kunešová, M.; et al. Prevalence of Severe Obesity among Primary School Children in 21 European Countries. Obes. Facts 2019, 12, 244–258. [Google Scholar] [CrossRef]
- Koletzko, B.; Fishbein, M.; Lee, W.S.; Moreno, L.; Mouane, N.; Mouzaki, M.; Verduci, E. Prevention of Childhood Obesity. J. Pediatr. Gastroenterol. Nutr. 2020, 70, 702–710. [Google Scholar] [CrossRef]
- Lin, T.K.; Teymourian, Y.; Tursini, M.S. The effect of sugar and processed food imports on the prevalence of overweight and obesity in 172 countries. Glob. Health 2018, 14, 35. [Google Scholar] [CrossRef]
- Adams, J.; White, M. Characterisation of UK diets according to degree of food processing and associations with socio-demographics and obesity: Cross-sectional analysis of UK National Diet and Nutrition Survey. Int. J. Behav. Nutr. Phys. Act. 2015, 12, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawal, K.; Patel, T.P.; Purohit, K.M.; Israni, K.; Kataria, V.; Bhatt, H.; Gupta, S. Influence of obese phenotype on metabolic profile, inflammatory mediators and stemness of hADSC in adipose tissue. Clin. Nutr. 2020, 0261561420300960. [Google Scholar] [CrossRef] [PubMed]
- Csongrádi, É.; Káplár, M.; Nagy, B.; Koch, C.; Juhász, A.; Bajnok, L.; Varga, Z.; Seres, I.; Karányi, Z.; Magyar, M.; et al. Adipokines as atherothrombotic risk factors in obese subjects: Associations with haemostatic markers and common carotid wall thickness. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Szepietowska, B.; Polonsky, B.; Sherazi, S.; Biton, Y.; Kutyifa, V.; McNitt, S.; Aktas, M.K.; Moss, A.J.; Zareba, W. Effect of obesity on the effectiveness of cardiac resynchronization to reduce the risk of first and recurrent ventricular tachyarrhythmia events. Cardiovasc. Diabetol. 2016, 15, 93. [Google Scholar] [CrossRef] [Green Version]
- Vistisen, D.; Witte, D.; Tabak, A.G.; Herder, C.; Brunner, E.J.; Kivimaki, M.; Færch, K. Patterns of Obesity Development before the Diagnosis of Type 2 Diabetes: The Whitehall II Cohort Study. PLoS Med. 2014, 11, e1001602. [Google Scholar] [CrossRef] [Green Version]
- Chandra, A.; Neeland, I.J.; Berry, J.D.; Ayers, C.R.; Rohatgi, A.; Das, S.R.; Khera, A.; McGuire, D.K.; De Lemos, J.A.; Turer, A.T. The Relationship of Body Mass and Fat Distribution With Incident Hypertension. J. Am. Coll. Cardiol. 2014, 64, 997–1002. [Google Scholar] [CrossRef] [Green Version]
- Cheong, K.C.; Ghazali, S.M.; Hock, L.K.; Subenthiran, S.; Huey, T.C.; Kuay, L.K.; Mustapha, F.; Yusoff, A.F.; Mustafa, A.N. The discriminative ability of waist circumference, body mass index and waist-to-hip ratio in identifying metabolic syndrome: Variations by age, sex and race. Diabetes Metab. Syndr. Clin. Res. Rev. 2015, 9, 74–78. [Google Scholar] [CrossRef]
- Kelishadi, R. Health impacts of Obesity. Pak. J. Med. Sci. 2014, 31, 239–242. [Google Scholar] [CrossRef]
- O’Doherty, M.G.; Jørgensen, T.; Borglykke, A.; Brenner, H.; Schöttker, B.; Wilsgaard, T.; Siganos, G.; Kavousi, M.; Hughes, M.; Müezzinler, A.; et al. Repeated measures of body mass index and C-reactive protein in relation to all-cause mortality and cardiovascular disease: Results from the consortium on health and ageing network of cohorts in Europe and the United States. Eur. J. Epidemiol. 2014, 29, 887–897. [Google Scholar] [CrossRef]
- Shantsila, A.; A Lane, D.; Beevers, D.G.; Lip, G.Y. Does obesity influence target organ damage and outcomes in patients with malignant phase hypertension? The West Birmingham Malignant Hypertension Project. Hypertens. Res. 2013, 36, 546–549. [Google Scholar] [CrossRef] [Green Version]
- Duncan, M.E.; Goldacre, M.J. Certification of deaths from diabetes mellitus and obesity in England: Trends into the twenty-first century. J. Public Health 2012, 35, 293–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, T.; Fontaine, K.R.; Keith, S.W.; Bangalore, S.S.; Campos, G.D.L.; Bartolucci, A.; Pajewski, N.M.; Allison, D. Obesity and mortality: Are the risks declining? Evidence from multiple prospective studies in the United States. Obes. Rev. 2014, 15, 619–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Kane, G. What is the real cost of our food? Implications for the environment, society and public health nutrition. Public Health Nutr. 2011, 15, 268–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, N.; Ahlawat, S.S.; Sharma, D.P.; Dabur, R. Novel trends in development of dietary fiber rich meat products—A critical review. J. Food Sci. Technol. 2013, 52, 633–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.-W.; Lee, M.; Oh, K.-J. Adipose Tissue-Derived Signatures for Obesity and Type 2 Diabetes: Adipokines, Batokines and MicroRNAs. J. Clin. Med. 2019, 8, 854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flehmig, G.; Scholz, M.; Klöting, N.; Fasshauer, M.; Tonjes, A.; Stumvoll, M.; Youn, B.-S.; Blüher, M. Identification of Adipokine Clusters Related to Parameters of Fat Mass, Insulin Sensitivity and Inflammation. PLoS ONE 2014, 9, e99785. [Google Scholar] [CrossRef]
- Cinti, S. Adipose Organ Development and Remodeling. Compr. Physiol. 2018, 8, 1357–1431. [Google Scholar]
- Froy, O.; Garaulet, M. The Circadian Clock in White and Brown Adipose Tissue: Mechanistic, Endocrine, and Clinical Aspects. Endocr. Rev. 2018, 39, 261–273. [Google Scholar] [CrossRef]
- Herz, C.T.; Kiefer, F.W. Adipose tissue browning in mice and humans. J. Endocrinol. 2019, 241, 97–109. [Google Scholar] [CrossRef]
- Tews, D.; Pula, T.; Funcke, J.; Jastroch, M.; Keuper, M.; Debatin, K.-M.; Wabitsch, M.; Fischer-Posovszky, P. Elevated UCP1 levels are sufficient to improve glucose uptake in human white adipocytes. Redox Boil. 2019, 26, 101286. [Google Scholar] [CrossRef]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.H.; Doria, A.; et al. Identification and importance of brown adipose tissue in adult humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cinti, S. White, brown, beige and pink: A rainbow in the adipose organ. Curr. Opin. Endocr. Metab. Res. 2019, 4, 29–36. [Google Scholar] [CrossRef]
- Corrêa, L.H.; Heyn, G.S.; Magalhaes, K.G. The Impact of the Adipose Organ Plasticity on Inflammation and Cancer Progression. Cells 2019, 8, 662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cinti, S. Pink Adipocytes. Trends Endocrinol. Metab. 2018, 29, 651–666. [Google Scholar] [CrossRef]
- De Rosa, G.; Catena, G.; Gaudio, G.; D’Angelo, A.; Maffioli, P. Adipose tissue dysfunction and metabolic disorders: Is it possible to predict who will develop type 2 diabetes mellitus? Role of markErs in the progreSsion of dIabeteS in obese paTIeNts. Cytokine 2020, 127, 154947. [Google Scholar] [CrossRef] [PubMed]
- Landecho, M.F.; Tuero, C.; Valenti, V.; Bilbao, I.; De La Higuera, M.; Frühbeck, G. Relevance of Leptin and Other Adipokines in Obesity-Associated Cardiovascular Risk. Nutriens 2019, 11, 2664. [Google Scholar] [CrossRef] [Green Version]
- Grewal, T.; Enrich, C.; Rentero, C.; Buechler, C. Annexins in Adipose Tissue: Novel Players in Obesity. Int. J. Mol. Sci. 2019, 20, 3449. [Google Scholar] [CrossRef] [Green Version]
- Buechler, C.; Feder, S.; Haberl, E.; Aslanidis, C. Chemerin Isoforms and Activity in Obesity. Int. J. Mol. Sci. 2019, 20, 1128. [Google Scholar] [CrossRef] [Green Version]
- Lima, L.C.F.; Braga, V.D.A.; Silva, M.D.S.D.F.; Cruz, J.D.C.; Santos, S.H.S.; Monteiro, M.M.D.O.; Balarini, C.M. Adipokines, diabetes and atherosclerosis: An inflammatory association. Front. Physiol. 2015, 6, 1. [Google Scholar] [CrossRef]
- Okla, M.; Zaher, W.; Alfayez, M.; Chung, S. Inhibitory Effects of Toll-Like Receptor 4, NLRP3 Inflammasome, and Interleukin-1β on White Adipocyte Browning. Inflammation 2018, 41, 626–642. [Google Scholar] [CrossRef]
- Xu, X.; Xu, J. Effects of different obesity-related adipokines on the occurrence of obstructive sleep apnea. Endocr. J. 2020, 20–36. [Google Scholar] [CrossRef] [Green Version]
- Pivtorak, K.V.; Shevchuk, N.A.; Pivtorak, N.A.; Fedzhaga, I.V. Correction of adipocyte secretion disorders in patients with non-alcoholic fatty liver disease with overweight and obesity. Wiad. Lek. 2019, 72, 1477–1480. [Google Scholar] [PubMed]
- Martín, M.S.; Guillén, J.R.O.; Cano, A.P.; Ángel, J.M.R. Metabolic syndrome, non-alcoholic fatty liver disease and hepatocarcinoma. Rev. Esp. Enferm. Dige. 2020, 112, 131–136. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Mottillo, E.P.; Granneman, J.G. Adipose tissue plasticity from WAT to BAT and in between. Biochim. Biophys. Acta Bioenerg. 2013, 1842, 358–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarroya, F.; Cereijo, R.; Villarroya, J.; Giralt, M. Brown adipose tissue as a secretory organ. Nat. Rev. Endocrinol. 2016, 13, 26–35. [Google Scholar] [CrossRef]
- Ma, X.; Lee, P.; Chisholm, D.J.; James, D.E. Control of Adipocyte Differentiation in Different Fat Depots; Implications for Pathophysiology or Therapy. Front. Endocrinol. 2015, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Żelechowska, P.; Kozłowska, E.; Pastwińska, J.; Agier, J.; Brzezińska–Błaszczyk, E. Adipocytokine Involvement in Innate Immune Mechanisms. J. Interf. Cytokine Res. 2018, 38, 527–538. [Google Scholar] [CrossRef]
- Gomez-Hernandez, A.; Beneit, N.; Díaz-Castroverde, S.; Escribano, O. Differential Role of Adipose Tissues in Obesity and Related Metabolic and Vascular Complications. Int. J. Endocrinol. 2016, 2016, 1216783. [Google Scholar] [CrossRef] [Green Version]
- Malaguarnera, M.; Vacante, M.; Russo, C.; Malaguarnera, G.; Antic, T.; Malaguarnera, L.; Bella, R.; Pennisi, G.; Galvano, F.; Frigiola, A. Lipoprotein(a) in Cardiovascular Diseases. BioMed Res. Int. 2013, 2013, 650989. [Google Scholar] [CrossRef] [Green Version]
- Jachimowicz-Duda, O.; Wąż, P.; Zorena, K. Interleukin-34 concentration is dependent on LDL cholesterol, HbA1c, CRP and WHR in patients with type 2 diabetes. In Proceedings of the Crossroads of Lipid Metabolism and Diabetes 2015, Copenhagen, Denmark, 19–24 April 2015; p. 52. [Google Scholar]
- Zorena, K.; Jachimowicz-Duda, O.; Wąż, P. The cut-off value for interleukin 34 as an additional potential inflammatory biomarker for the prediction of the risk of diabetic complications. Biomarkers 2016, 21, 1–7. [Google Scholar] [CrossRef]
- Malin, S.K.; Kirwan, J.P.; Sia, C.L.; González, F. Pancreatic β-cell dysfunction in polycystic ovary syndrome: Role of hyperglycemia-induced nuclear factor-κB activation and systemic inflammation. Am. J. Physiol. Metab. 2015, 308, 770–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomić, M.; Ljubić, S.; Kaštelan, S.; Antunica, A.G.; Jazbec, A.; Poljičanin, T. Inflammation, Haemostatic Disturbance, and Obesity: Possible Link to Pathogenesis of Diabetic Retinopathy in Type 2 Diabetes. Mediat. Inflamm. 2013, 2013, 818671. [Google Scholar] [CrossRef] [PubMed]
- Richardson, V.R.; Smith, K.; Carter, A.M. Adipose tissue inflammation: Feeding the development of type 2 diabetes mellitus. Immunobiology 2013, 218, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Daniele, G.; Mendoza, R.G.; Winnier, D.; Fiorentino, T.V.; Pengou, Z.; Cornell, J.; Andreozzi, F.; Jenkinson, C.; Cersosimo, E.; Federici, M.; et al. The inflammatory status score including IL-6, TNF-α, osteopontin, fractalkine, MCP-1 and adiponectin underlies whole-body insulin resistance and hyperglycemia in type 2 diabetes mellitus. Acta Diabetol. 2013, 51, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Schöttker, B.; Herder, C.; Rothenbacher, D.; Roden, M.; Kolb, H.; Müller, H.; Brenner, H. Proinflammatory Cytokines, Adiponectin, and Increased Risk of Primary Cardiovascular Events in Diabetic Patients With or Without Renal Dysfunction. Diabetes Care 2013, 36, 1703–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monda, V.; Polito, R.; Lovino, A.; Finaldi, A.; Valenzano, A.; Nigro, E.; Corso, G.; Sessa, F.; Asmundo, A.; Di Nunno, N.; et al. Short-Term Physiological Effects of a Very Low-Calorie Ketogenic Diet: Effects on Adiponectin Levels and Inflammatory States. Int. J. Mol. Sci. 2020, 21, 3228. [Google Scholar] [CrossRef]
- Koska, J.; Ortega, E.; Bogardus, C.; Krakoff, J.; Bunt, J.C. The effect of insulin on net lipid oxidation predicts worsening of insulin resistance and development of type 2 diabetes mellitus. Am. J. Physiol. Metab. 2007, 293, 264–269. [Google Scholar] [CrossRef]
- Li, K.; Li, L.; Yang, G.Y.; Liu, H.; Li, S.B.; Boden, G. Effect of short hairpin RNA-mediated adiponectin/Acrp30 down-regulation on insulin signaling and glucose uptake in the 3T3-L1 adipocytes. J. Endocrinol. Investig. 2009, 33, 96–102. [Google Scholar] [CrossRef]
- Pal, G.K.; Adithan, C.; Ananthanarayanan, P.H.; Pal, P.; Nanda, N.; Durgadevi, T.; Lalitha, V.; Syamsunder, A.N.; Dutta, T.K. Sympathovagal Imbalance Contributes to Prehypertension Status and Cardiovascular Risks Attributed by Insulin Resistance, Inflammation, Dyslipidemia and Oxidative Stress in First Degree Relatives of Type 2 Diabetics. PLoS ONE 2013, 8, e78072. [Google Scholar] [CrossRef]
- Pandey, G.; Shihabudeen, M.S.; David, H.P.; Thirumurugan, K.; Thirumurugan, K. Association between hyperleptinemia and oxidative stress in obese diabetic subjects. J. Diabetes Metab. Disord. 2015, 14, 24. [Google Scholar] [CrossRef] [Green Version]
- Halcox, J.; Roy, C.; Tubach, F.; Banegas, J.R.; Dallongeville, J.; De Backer, G.; Guallar, E.; Sazova, O.; Medina, J.; Perk, J.; et al. C-reactive protein levels in patients at cardiovascular risk: EURIKA study. BMC Cardiovasc. Disord. 2014, 14, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Shukaili, A.; Al-Ghafri, S.; Al-Marhoobi, S.; Al-Abri, S.; Al-Lawati, J.; Al-Maskari, M. Analysis of Inflammatory Mediators in Type 2 Diabetes Patients. Int. J. Endocrinol. 2013, 2013, 976810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, F.J.D.C.; Fernandez-Perez, C.; Ruiz, I.M.; Pérez-Jáuregui, C.D.G.; Rodríguez, B.S.; Losada, T.G.; Galindo, A.S. Microvascular complications and risk factors in patients with type 2 diabetes. Endocrinol. Nutr. 2011, 58, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.-J.; Lee, S.K.; Song, Y.S.; Jang, Y.J.; Park, H.S.; Hong, J.P.; Ko, A.R.; Kim, D.Y.; Kim, J.-H.; Lee, Y.J.; et al. IL-34 Is Associated with Obesity, Chronic Inflammation, and Insulin Resistance. J. Clin. Endocrinol. Metab. 2014, 99, 1263–1271. [Google Scholar] [CrossRef] [Green Version]
- Nieto-Vazquez, I.; Fernández-Veledo, S.; Krämer, D.K.; Vila, R.; García, L.; Lorenzo, M. Insulin resistance associated to obesity: The link TNF-alpha. Arch. Physiol. Biochem. 2008, 114, 183–194. [Google Scholar] [CrossRef]
- Sookoian, S.; Garciía, S.I.; Gianotti, T.F.; Dieuzeide, G.; Gonzalez, C.; Pirola, C. The G-308A Promoter Variant of the Tumor Necrosis Factor-α Gene Is Associated With Hypertension in Adolescents Harboring the Metabolic Syndrome. Am. J. Hypertens. 2005, 18, 1271–1275. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.; Sun, J.; Huang, J.; He, Y.; Yu, A.; Yu, X.; Yang, Z. TNF-α Reduces G0s2 Expression and Stimulates Lipolysis through PPAR-γ Inhibition in 3T3-L1 Adipocytes. Cytokine 2014, 69, 196–205. [Google Scholar] [CrossRef]
- Shivanna, M.; Srinivas, S.P. Elevated cAMP opposes (TNF-α)-induced loss in the barrier integrity of corneal endothelium. Mol. Vis. 2010, 16, 1781–1790. [Google Scholar]
- Kadłubowska, J.; Malaguarnera, L.; Wąż, P.; Zorena, K. Neurodegeneration and Neuroinflammation in Diabetic Retinopathy: Potential Approaches to Delay Neuronal Loss. Curr. Neuropharmacol. 2016, 14, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Mrugacz, M.; Bryl, A.; Bossowski, A. Neuroretinal Apoptosis as a Vascular Dysfunction in Diabetic Patients. Curr. Neuropharmacol. 2016, 14, 826–830. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.-S.; Wu, C.-S.; Chang, J.-F.; Lin, W.-N. Leptin Promotes cPLA2 Gene Expression through Activation of the MAPK/NF-κB/p300 Cascade. Int. J. Mol. Sci. 2015, 16, 27640–27658. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.-M.; Yu, H.; Van Der Ploeg, L.H. Evidence of altered hypothalamic pro-opiomelanocortin/neuropeptide Y mRNA expression in tubby mice. Mol. Brain Res. 1998, 59, 273–279. [Google Scholar] [CrossRef]
- Ge, T.T.; Yao, X.X.; Zhao, F.L.; Zou, X.H.; Yang, W.; Cui, R.J.; Li, B.J. Role of leptin in the regulation of food intake in fasted mice. J. Cell. Mol. Med. 2020, 24, 4524–4532. [Google Scholar] [CrossRef]
- Pérez, A.P.; Vilariño-García, T.; Fernandez-Riejos, P.; Martín-González, J.; Segura-Egea, J.J.; Sánchez-Margalet, V. Role of leptin as a link between metabolism and the immune system. Cytokine Growth Factor Rev. 2017, 35, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Elfassy, Y.; McAvoy, C.; Fellahi, S.; Dupont, J.; Feve, B.; Levy, R.; Bastard, J.-P. Seminal plasma adipokines: Involvement in human reproductive functions. Eur. Cytokine Netw. 2017, 28, 141–150. [Google Scholar] [CrossRef]
- Nemer, L.B.; Shi, H.; Carr, B.R.; Word, R.A.; Bukulmez, O. Effect of Body Weight on Metabolic Hormones and Fatty Acid Metabolism in Follicular Fluid of Women Undergoing In Vitro Fertilization: A Pilot Study. Reprod. Sci. 2018, 26, 404–411. [Google Scholar] [CrossRef]
- Morioka, T.; Emoto, M.; Yamazaki, Y.; Kawano, N.; Imamura, S.; Numaguchi, R.; Urata, H.; Motoyama, K.; Mori, K.; Fukumoto, S.; et al. Leptin is associated with vascular endothelial function in overweight patients with type 2 diabetes. Cardiovasc. Diabetol. 2014, 13, 10. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.-H.; Kim, B.-Y.; Mok, J.-O.; Kang, S.-K.; Kim, C.-H. Association between serum adipocytokine levels and microangiopathies in patients with type 2 diabetes mellitus. J. Diabetes Investig. 2013, 5, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, S.; Arefhosseini, S.R.; Ebrahimi-Mamaeghani, M.; Fallah, P.; Bazi, Z. Adiponectin as a potential biomarker of vascular disease. Vasc. Health Risk Manag. 2015, 11, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Mihalopoulos, N.L.; Yap, J.T.; Beardmore, B.; Holubkov, R.; Nanjee, M.N.; Hoffman, J.M. Cold-Activated Brown Adipose Tissue is Associated with Less Cardiometabolic Dysfunction in Young Adults with Obesity. Obesity 2020, 28, 916–923. [Google Scholar] [CrossRef]
- Moradi, N.; Fadaei, R.; Rashidbeygi, E.; Kargasheh, F.B.; Malek, M.; Nahrkhalaji, A.S.; Fallah, S. Evaluation of changing the pattern of CTRP5 and inflammatory markers levels in patients with coronary artery disease and type 2 diabetes mellitus. Arch. Physiol. Biochem. 2020, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kumada, M.; Kihara, S.; Ouchi, N.; Kobayashi, H.; Okamoto, Y.; Ohashi, K.; Maeda, K.; Nagaretani, H.; Kishida, K.; Maeda, N.; et al. Adiponectin Specifically Increased Tissue Inhibitor of Metalloproteinase-1 Through Interleukin-10 Expression in Human Macrophages. Circulation 2004, 109, 2046–2049. [Google Scholar] [CrossRef] [Green Version]
- Zha, N.; Wu, X.; Gao, P. Adiponectin and Its Receptors in Diabetic Kidney Disease: Molecular Mechanisms and Clinical Potential. Endocrinology 2017, 158, 2022–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alnaggar, A.R.L.; Sayed, M.; El-Deena, K.E.; Gomaa, M.; Hamed, Y. Evaluation of serum adiponectin levels in diabetic nephropathy. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 128–131. [Google Scholar] [CrossRef] [PubMed]
- Steppan, C.M.; Lazar, M.A. The current biology of resistin. J. Intern. Med. 2004, 255, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Cebeci, E.; Çakan, C.; Gursu, M.; Uzun, S.; Karadağ, S.; Koldaş, M.; Çalhan, T.; Helvaci, S.A.; Öztürk, S. The Main Determinants of Serum Resistin Level in Type 2 Diabetic Patients are Renal Function and Inflammation not Presence of Microvascular Complication, Obesity and Insulin Resistance. Exp. Clin. Endocrinol. Diabetes 2018, 127, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.S.; Le, Y.Y. Role of resistin in inflammation and inflammation-related diseases. Cell. Mol. Immunol. 2006, 3, 29–34. [Google Scholar]
- Reilly, M. Resistin Is an Inflammatory Marker of Atherosclerosis in Humans. Circulation 2005, 111, 932–939. [Google Scholar] [CrossRef] [Green Version]
- Uslu, S.; Kebapçi, N.; Kara, M.; Bal, C. Relationship between adipocytokines and cardiovascular risk factors in patients with type 2 diabetes mellitus. Exp. Ther. Med. 2012, 4, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Pourshahidi, K.; Wallace, J.M.W.; Mulhern, M.S.; Horigan, G.; Strain, J.J.; McSorley, E.M.; Magee, P.J.; Bonham, M.P.; Livingstone, M.B.E. Indices of adiposity as predictors of cardiometabolic risk and inflammation in young adults. J. Hum. Nutr. Diet. 2015, 29, 26–37. [Google Scholar] [CrossRef]
- Osawa, H.; Ochi, M.; Kato, K.; Yamauchi, J.; Nishida, W.; Takata, Y.; Kawamura, R.; Onuma, H.; Takasuka, T.; Shimizu, I.; et al. Serum resistin is associated with the severity of microangiopathies in type 2 diabetes. Biochem. Biophys. Res. Commun. 2007, 355, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Gulcelik, N.E.; Usman, A.; Gürlek, A. Role of adipocytokines in predicting the development of diabetes and its late complications. Endocrinology 2009, 36, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Gulcelik, N.E.; Karakaya, J.; Gedik, A.; Usman, A.; Gurlek, A. Serum vaspin levels in type 2 diabetic women in relation to microvascular complications. Eur. J. Endocrinol. 2009, 160, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetta, H.F.; Ez-Eldeen, M.E.; Mohamed, G.A.; Gaber, M.A.; ElBadre, H.M.; Ahmed, E.A.; Abdellatief, R.B.; Abd-ElBaky, R.M.; Elkady, A.; Nafee, A.M.; et al. Visfatin Serum Levels in Obese Type 2 Diabetic Patients: Relation to Proinflammatory Cytokines and Insulin Resistance. Egypt J. Immunol. 2018, 25, 141–151. [Google Scholar]
- Yamawaki, H. Vascular effects of novel adipocytokines: Focus on vascular contractility and inflammatory responses. Boil. Pharm. Bull. 2011, 34, 307–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smitka, K.; Marešová, D. Adipose Tissue as an Endocrine Organ: An Update on Pro-inflammatory and Anti-inflammatory Microenvironment. Prague Med. Rep. 2015, 116, 87–111. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.S.; Lee, M.H.; Song, H.K.; Kim, J.E.; Ghee, J.Y.; Cha, J.J.; Lee, J.E.; Kim, H.; Han, J.-Y.; Cha, D.R. Chronic Administration of Visfatin Ameliorated Diabetic Nephropathy in Type 2 Diabetic Mice. Kidney Blood Press. Res. 2016, 41, 311–324. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.-D.; Xing, L.; Teng, J.-R.; Li, S.; Mi, N. Effects of basal insulin application on serum visfatin and adiponectin levels in type 2 diabetes. Exp. Ther. Med. 2015, 9, 2219–2224. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-P.; Chung, F.-M.; Chang, D.-M.; Tsai, J.C.-R.; Huang, H.-F.; Shin, S.-J.; Lee, Y.-J. Elevated Plasma Level of Visfatin/Pre-B Cell Colony-Enhancing Factor in Patients with Type 2 Diabetes Mellitus. J. Clin. Endocrinol. Metab. 2006, 91, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.-C.; Chang, T.-J.; Lee, W.-J.; Chuang, L.-M. The relationship of visfatin/pre–B-cell colony-enhancing factor/nicotinamide phosphoribosyltransferase in adipose tissue with inflammation, insulin resistance, and plasma lipids. Metabolism 2010, 59, 93–99. [Google Scholar] [CrossRef]
- Nway, N.C.; Sitticharoon, C.; Chatree, S.; Maikaew, P. Correlations between the expression of the insulin sensitizing hormones, adiponectin, visfatin, and omentin, and the appetite regulatory hormone, neuropeptide Y and its receptors in subcutaneous and visceral adipose tissues. Obes. Res. Clin. Pract. 2016, 10, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Ikmal, S.I.Q.S.; Huri, H.Z.; Vethakkan, S.R.; Ahmad, W.A.W. Potential Biomarkers of Insulin Resistance and Atherosclerosis in Type 2 Diabetes Mellitus Patients with Coronary Artery Disease. Int. J. Endocrinol. 2013, 2013, 698567. [Google Scholar] [CrossRef] [Green Version]
- Jialal, I.; Devaraj, S.; Kaur, H.; Adams-Huet, B.; Bremer, A.A. Increased Chemerin and Decreased Omentin-1 in Both Adipose Tissue and Plasma in Nascent Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2013, 98, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Esteghamati, A.; Azizi, R.; Ebadi, M.; Noshad, S.; Mousavizadeh, K.; Afarideh, M.; Nakhjavani, M. The Comparative Effect of Pioglitazone and Metformin on Serum Osteoprotegerin, Adiponectin and Intercellular Adhesion Molecule Concentrations in Patients with Newly Diagnosed Type 2 Diabetes: A Randomized Clinical Trial. Exp. Clin. Endocrinol. Diabetes 2015, 123, 289–295. [Google Scholar] [CrossRef]
- El-Mesallamy, H.O.; El-Derany, M.O.; Hamdy, N.M. Serum Omentin-1 and Chemerin Levels Are Interrelated in Patients with Type 2 Diabetes Mellitus with or without Ischaemic Heart Disease: Omentin-1 and Chemerin Levels in Type 2 Diabetes with or without Heart Disease. Diabet. Med. 2011, 28, 1194–1200. [Google Scholar] [CrossRef]
- Zhuang, X.H.; Sun, F.D.; Chen, S.H.; Liu, Y.T.; Liu, W.; Li, X.B.; Pan, Z.; Lou, N.J. Circulating Chemerin Levels are Increased in First-Degree Relatives of Type 2 Diabetic Patients. Clin. Lab. 2014, 60, 983–988. [Google Scholar] [CrossRef]
- Coimbra, S.; Oliveira, H.; Reis, F.; Belo, L.; Rocha, S.; Quintanilha, A.; Figueiredo, A.; Teixeira, F.; Castro, E.; Rocha-Pereira, P.; et al. Circulating adipokine levels in Portuguese patients with psoriasis vulgaris according to body mass index, severity and therapy. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 1386–1394. [Google Scholar] [CrossRef]
- Rodríguez-Cerdeira, C.; Cordeiro-Rodríguez, M.; Gregorio, M.C.; López-Barcenas, A.; Martínez-Herrera, E.; Fabbrocini, G.; Sinani, A.; Arenas, R.; González-Cespón, J.L. Biomarkers of Inflammation in Obesity-Psoriatic Patients. Mediat. Inflamm. 2019, 2019, 7353420. [Google Scholar] [CrossRef]
- Fernández-Armenteros, J.; Gómez-Arbonés, X.; Buti-Soler, M.; Betriu-Bars, A.; Sanmartin-Novell, V.; Ortega-Bravo, M.; Martínez-Alonso, M.; Gari, E.; Portero-Otín, M.; Santamaria-Babi, L.; et al. Psoriasis, metabolic syndrome and cardiovascular risk factors. A population-based study. J. Eur. Acad. Dermatol. Venereol. 2018, 33, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Bavoso, N.C.; Pinto, J.M.; Soares, M.M.S.; Diniz, M.D.S.; Júnior, A.L.T. Psoriasis in obesity: Comparison of serum levels of leptin and adiponectin in obese subjects-cases and controls. An. Bras. Dermatol. 2019, 94, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Kyriakou, A.; Patsatsi, A.; Sotiriadis, D.; Goulis, D.G. Serum Leptin, Resistin, and Adiponectin Concentrations in Psoriasis: A Meta-Analysis of Observational Studies. Dermatology 2017, 233, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, Y.; Zhang, X.; Zhu, S.; He, H. Prevalence of peripheral neuropathy in patients with diabetes: A systematic review and meta-analysis. Prim. Care Diabetes 2020. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Sloan, G.; Ye, Y.; Wang, S.; Duan, B.; Tesfaye, S.; Gao, L. New Perspective in Diabetic Neuropathy: From the Periphery to the Brain, a Call for Early Detection, and Precision Medicine. Front. Endocrinol. 2020, 10, 929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuttolomondo, A.; Maida, C.; Pinto, A. Diabetic foot syndrome: Immune-inflammatory features as possible cardiovascular markers in diabetes. World J. Orthop. 2015, 6, 62–76. [Google Scholar] [CrossRef] [Green Version]
- Bruhn-Olszewska, B.; Korzon-Burakowska, A.; Węgrzyn, G.; Jakóbkiewicz-Banecka, J. Prevalence of polymorphisms in OPG, Rankl and Rank as potential markers for Charcot arthropathy development. Sci. Rep. 2017, 7, 501. [Google Scholar] [CrossRef] [Green Version]
- Małecki, R.; Rosiński, K.; Adamiec, R. Etiological Factors of Infections in Diabetic Foot Syndrome–Attempt to Define Optimal Empirical Therapy. Adv. Clin. Exp. Med. 2014, 23, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Jachomowicz-Duda, O.; Wąż, P.; Zorena, K. The relationship between interleukin-34 level and selected biochemical and clinical parameters in patients with type 2 diabetes. In Proceedings of the Synergy Conference: “Autoimmunity in 2017-Where Are We Now?”, New York, NY, USA, 28–29 April 2017. [Google Scholar]
- Viswanathan, V.; Dhamodharan, U.; Srinivasan, V.; Rajaram, R.; Aravindhan, V. Single nucleotide polymorphisms in cytokine/chemokine genes are associated with severe infection, ulcer grade and amputation in diabetic foot ulcer. Int. J. Boil. Macromol. 2018, 118, 1995–2000. [Google Scholar] [CrossRef]
- Tuttolomondo, A.; La Placa, S.; Di Raimondo, D.; Bellia, C.; Caruso, A.; Sasso, B.L.; Guercio, G.; Diana, G.; Ciaccio, M.; Licata, G.; et al. Adiponectin, resistin and IL-6 plasma levels in subjects with diabetic foot and possible correlations with clinical variables and cardiovascular co-morbidity. Cardiovasc. Diabetol. 2010, 9, 50. [Google Scholar] [CrossRef] [Green Version]
- Herder, C.; Bongaerts, B.W.C.; Ouwens, D.M.; Rathmann, W.; Heier, M.; Carstensen-Kirberg, M.; Koenig, W.; Thorand, B.; Roden, M.; Meisinger, C.; et al. Low serum omentin levels in the elderly population with Type 2 diabetes and polyneuropathy. Diabet. Med. 2015, 32, 1479–1483. [Google Scholar] [CrossRef]
- Ahmad, J.; Zubair, M.; Malik, A. Plasma adiponectin, IL-6, hsCRP, and TNF-α levels in subject with diabetic foot and their correlation with clinical variables in a North Indian tertiary care hospital. Indian J. Endocrinol. Metab. 2012, 16, 769–776. [Google Scholar] [CrossRef]



{kind=link}
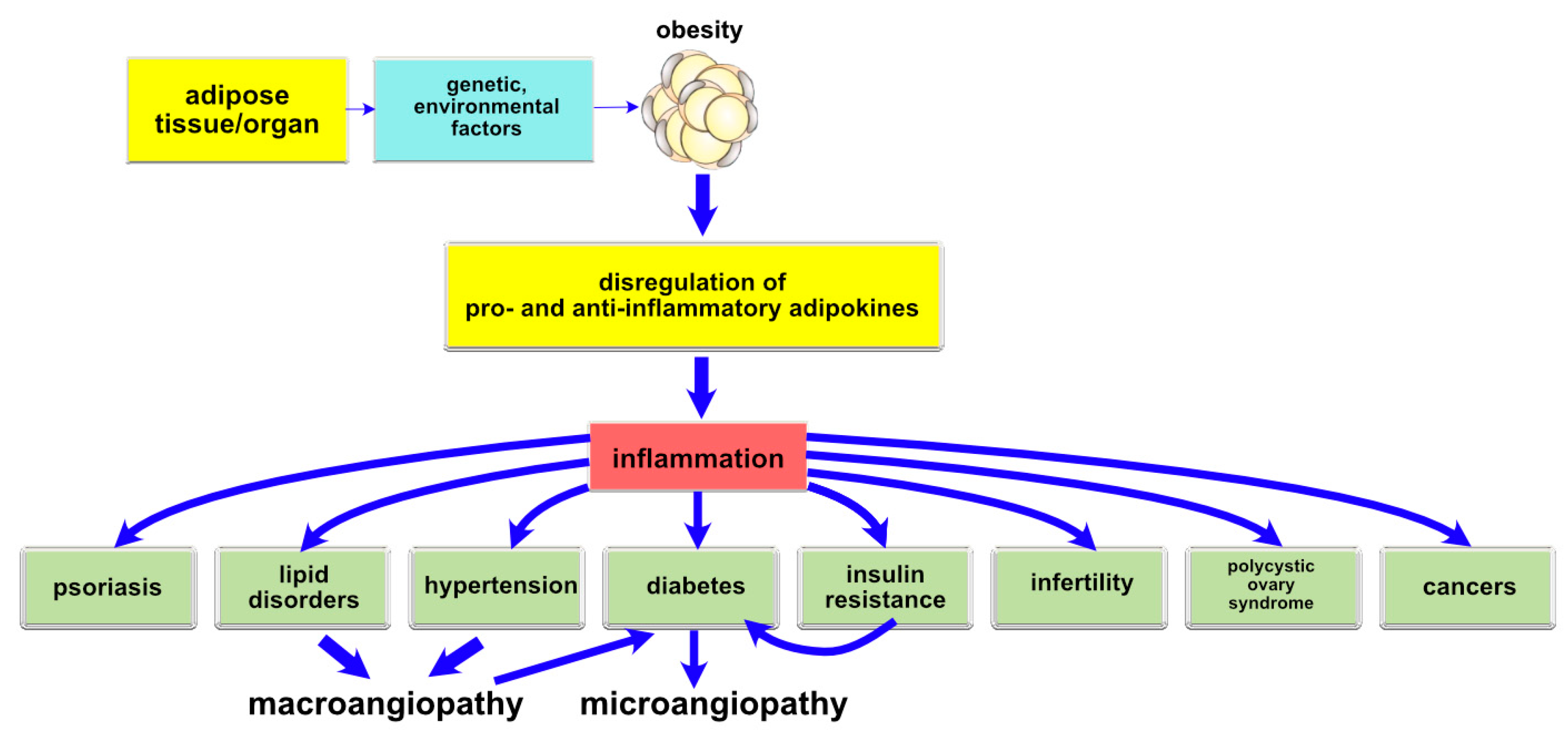{kind=link}
| Authors | Biochemical Factors | Concentration of Pro-Inflammatory and Anti-Inflammatory adipokines | Complications |
|---|---|---|---|
| Chandra et al. [18] | leptin adiponectin IL6 | higher concentration of leptin and lower concentration of adiponectin | hypertension |
| Jachimowicz-Duda et al. [52] | IL34 | higher concentration of IL34 | lipid disorders, macroangiopathy, T2DM |
| Zorena et al. [53] | IL34 | higher concentration of IL34 | microangiophaty, macroangiopathy, T2DM |
| Malin et al. [54] | TNFα | higher concentration of TNFα | insulin resistance |
| Daniele et al. [57] | adiponectin TNFα, IL6, MCP1 osteopontin, fractalkine | higher concentration of adiponectin, TNFα, IL6, MCP1, osteopontin and fractalkine | hyperglycemia, insulin resistance, T2DM |
| Chang, et al. [67] | IL34 | higher concentration of IL34 | atherosclerosis, insulin resistance, blood pressure |
| Shivanna et al. [71] | TNFα | higher concentration of TNFα | blood pressure, insulin resistance, atherosclerosis |
| Elfassy et al. [78] | leptin | increased concentration of leptin | reduced fertility in obese men |
| Bou Nemer et al. [79] | leptin | increased concentration of leptin in follicular fluid of in obese women undergoing in vitro fertilization compared to follicular fluid from nonobese (normal weight and overweight) women | reduced fertility in obese women |
| Jung et al. [81] | leptin | increased concentration of leptin | neuropathy, T2DM |
| Alnaggar et al. [87] | adiponectin | increased serum and urinary of adiponectin | T2DM, nephropathy, hypertension |
| Reilly et al. [91] | resistin | increased concentration of resistin | inflammation, metabolic factors and atherosclerosis |
| Osawa et al. [94] | resistin | increased concentration of resistin | diabetic retinopathy, nephropathy and neuropathy |
| El-Mesallamy et al. [108] | chemerin omentin-1 | increased concentration of serum chemerin decreased level of serum omentin-1 | T2DM, ischaemic heart disease |
| Zhuang et al. [109] | chemerin | increased of chemerin concentration in healthy subjects but with first-degree relatives (FDRs) of T2DM patients | insulin resistance |
| Coimbra et al. [110] | TNFα, IL6, leptin resistin adiponectin | higher concentration of leptin, resistin, TNFα, IL6 and significantly lower concentration of adiponectin | psoriasis, overweight/obesity |
| Bavoso et al. [113] | leptin adiponectin | increased concentration of lepton and lower of adiponectin | disregulation of the immune system, inflammation, psoriasis, obesity |
| Tuttolomondo et al. [122] | adiponectin IL6 | lower concentration of adiponectin and higher IL6 | T2DM diabetic foot |
| Herder et al. [123] | omentin | lower concentration of omentin | diabetic sensorimotor polyneuropathy |
| Ahmad et al. [124] | adiponectin IL6 TNFα | lower concentration of adiponectin higher concentration of IL6 higher TNFα | T2DM diabetic foot retinopathy nephropathy neuropathy |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103570
Zorena K, Jachimowicz-Duda O, Ślęzak D, Robakowska M, Mrugacz M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. International Journal of Molecular Sciences. 2020; 21(10):3570. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103570
Chicago/Turabian StyleZorena, Katarzyna, Olga Jachimowicz-Duda, Daniel Ślęzak, Marlena Robakowska, and Małgorzata Mrugacz. 2020. "Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications" International Journal of Molecular Sciences 21, no. 10: 3570. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103570