Epigallocatechin 3-Gallate Has a Neuroprotective Effect in Retinas of Rabbits with Ischemia/Reperfusion through the Activation of Nrf2/HO-1

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
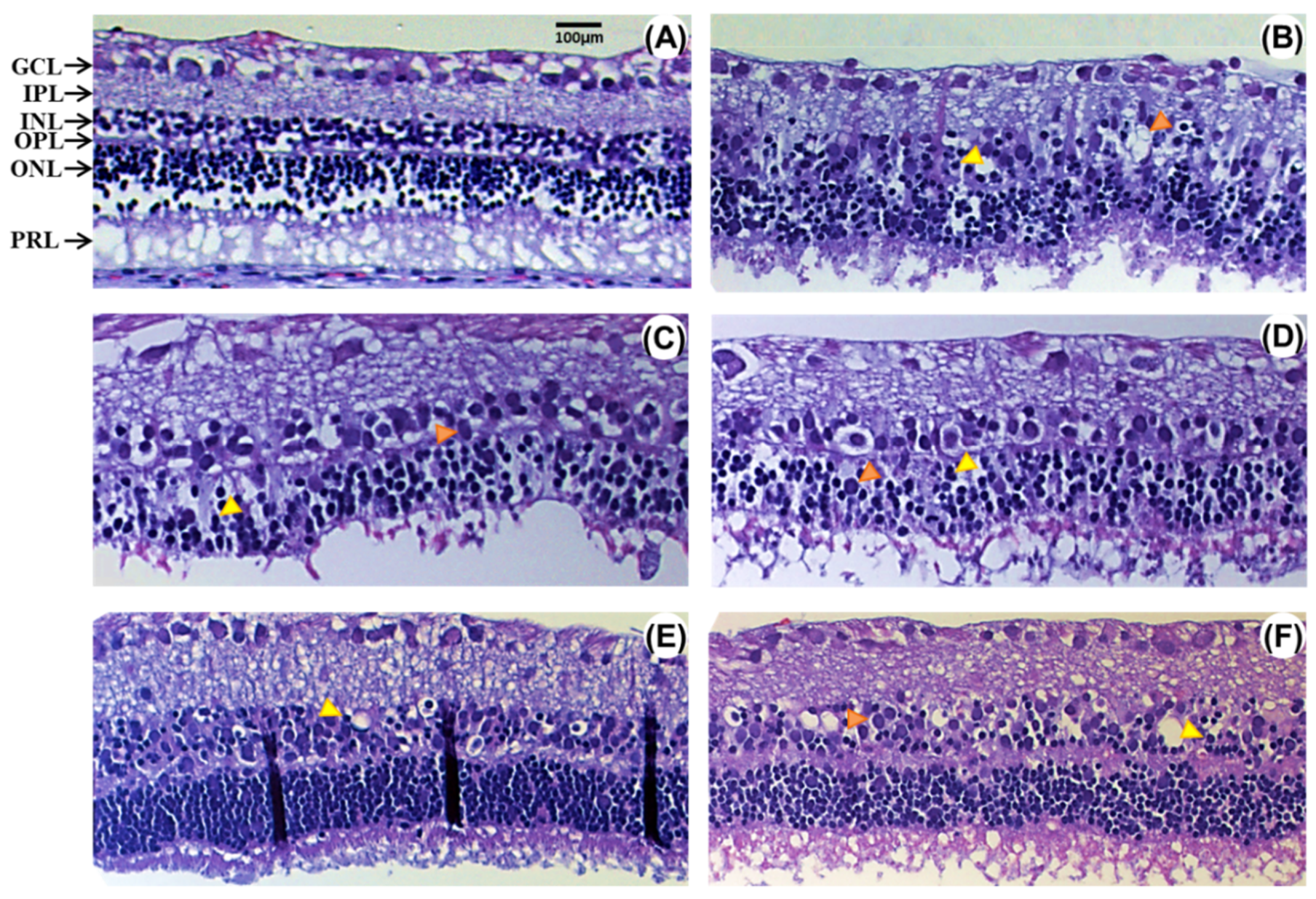
2.1. Effect of EGCG on Morphological Changes after Retinal Ischemia-Reperfusion (rI/R)
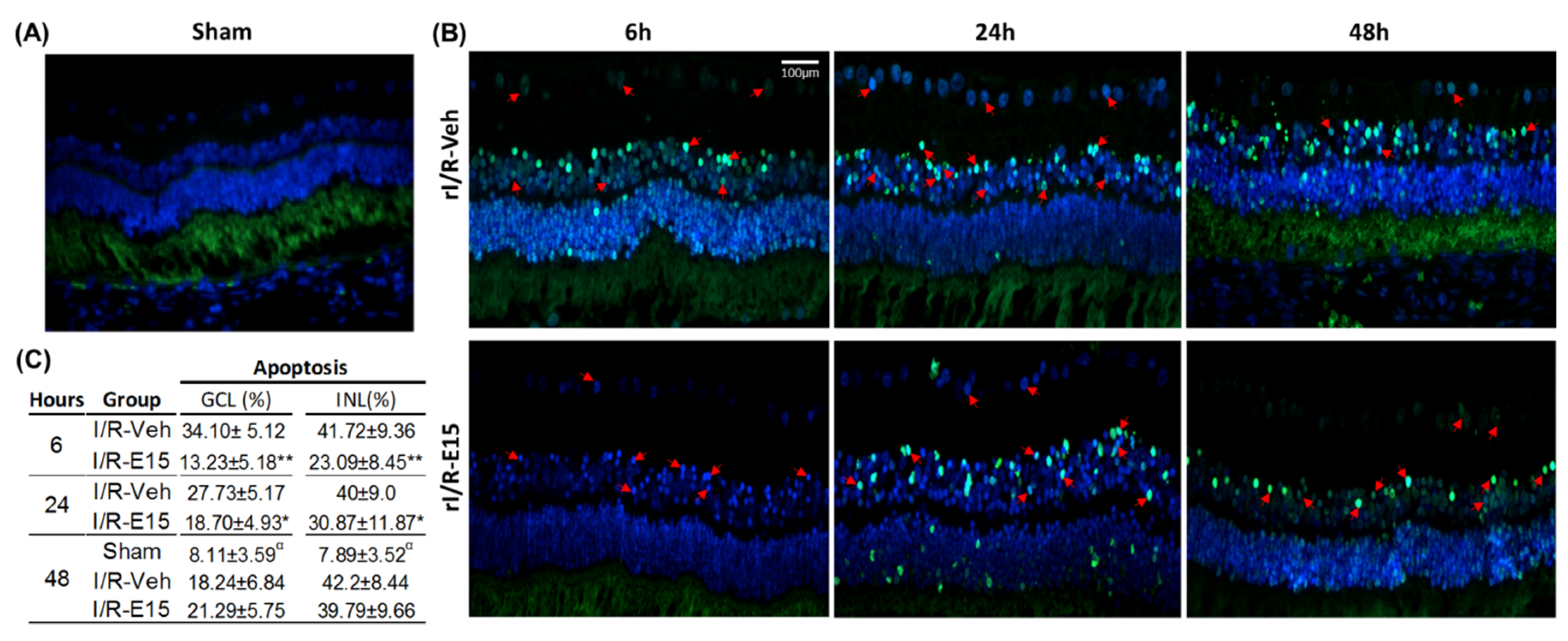
2.2. Neuroprotective Efficacy of EGCG at an Optimal Dose
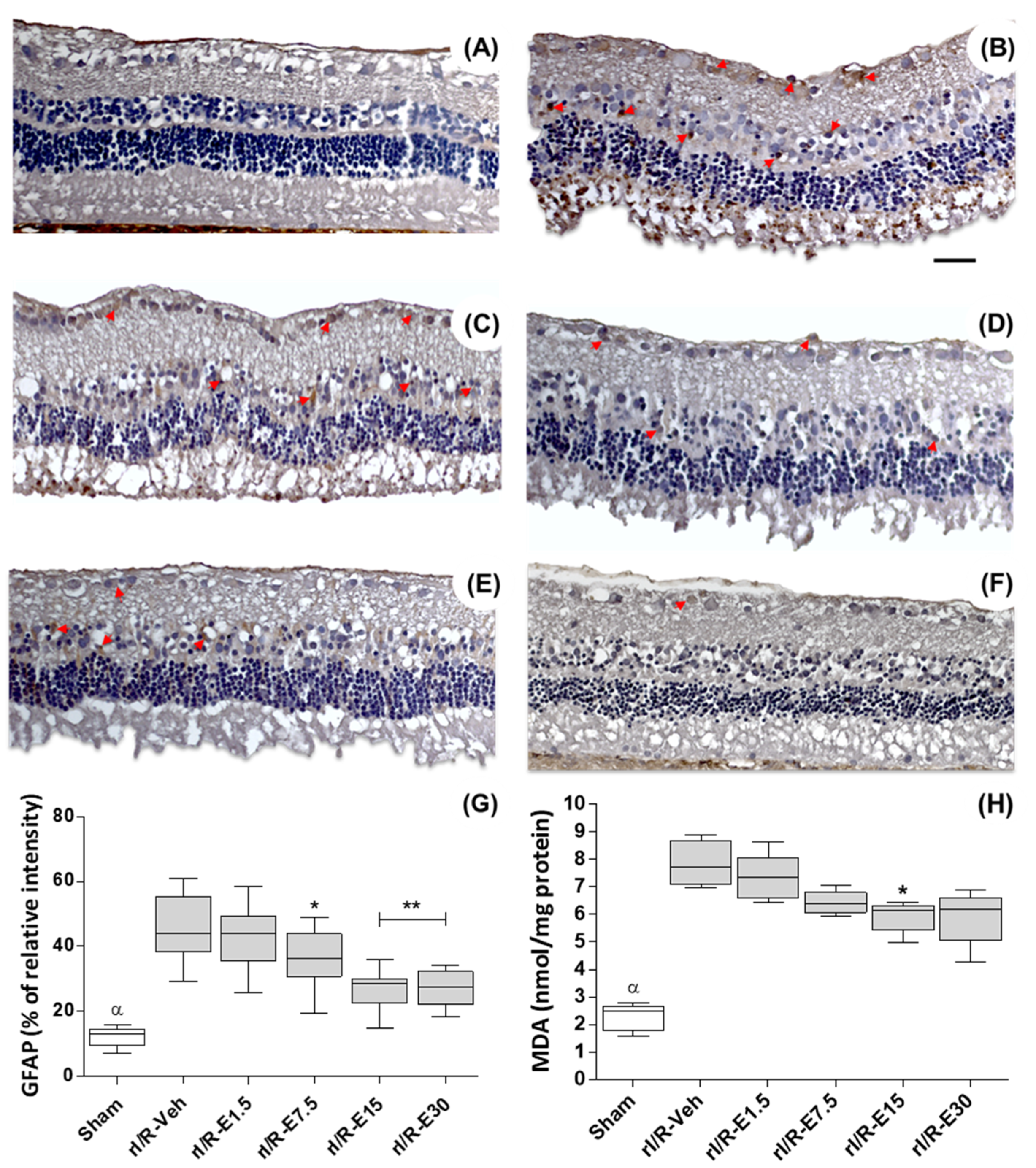
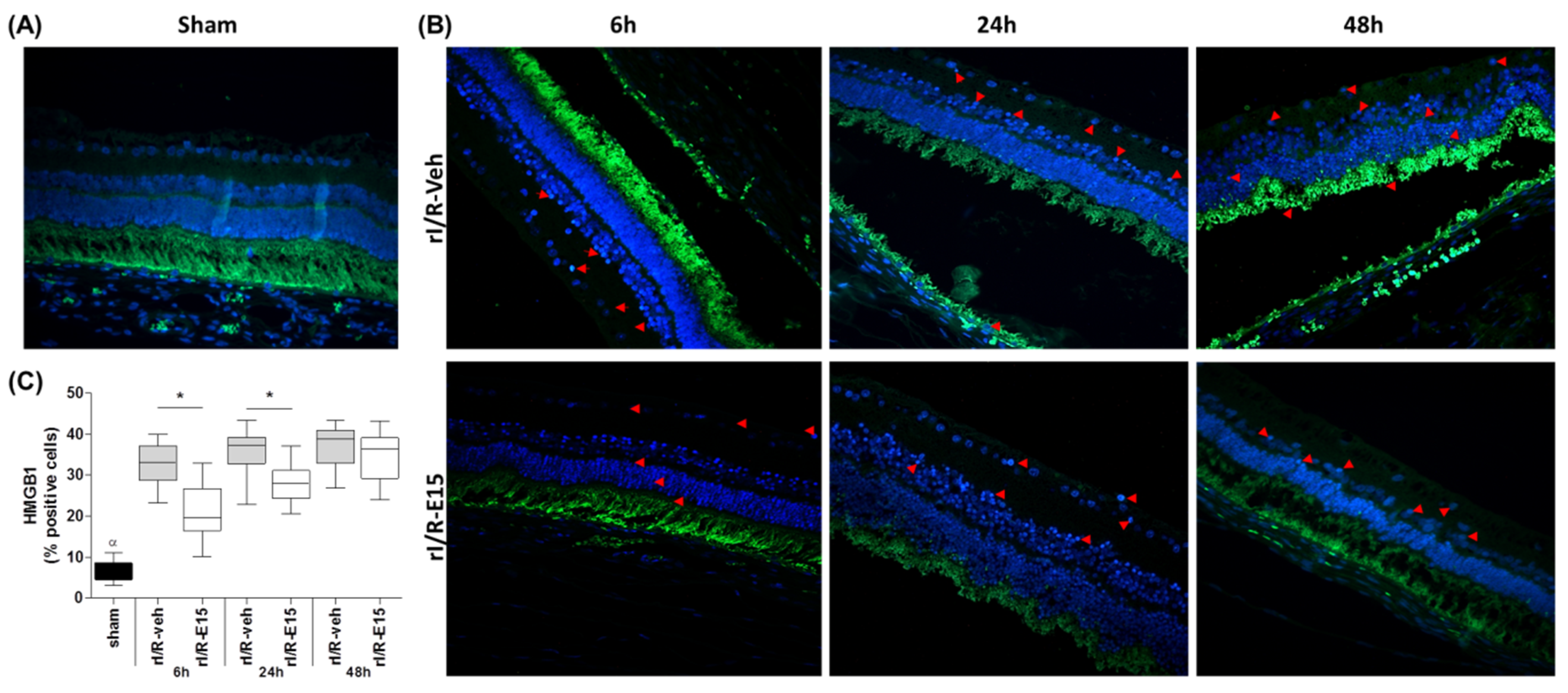
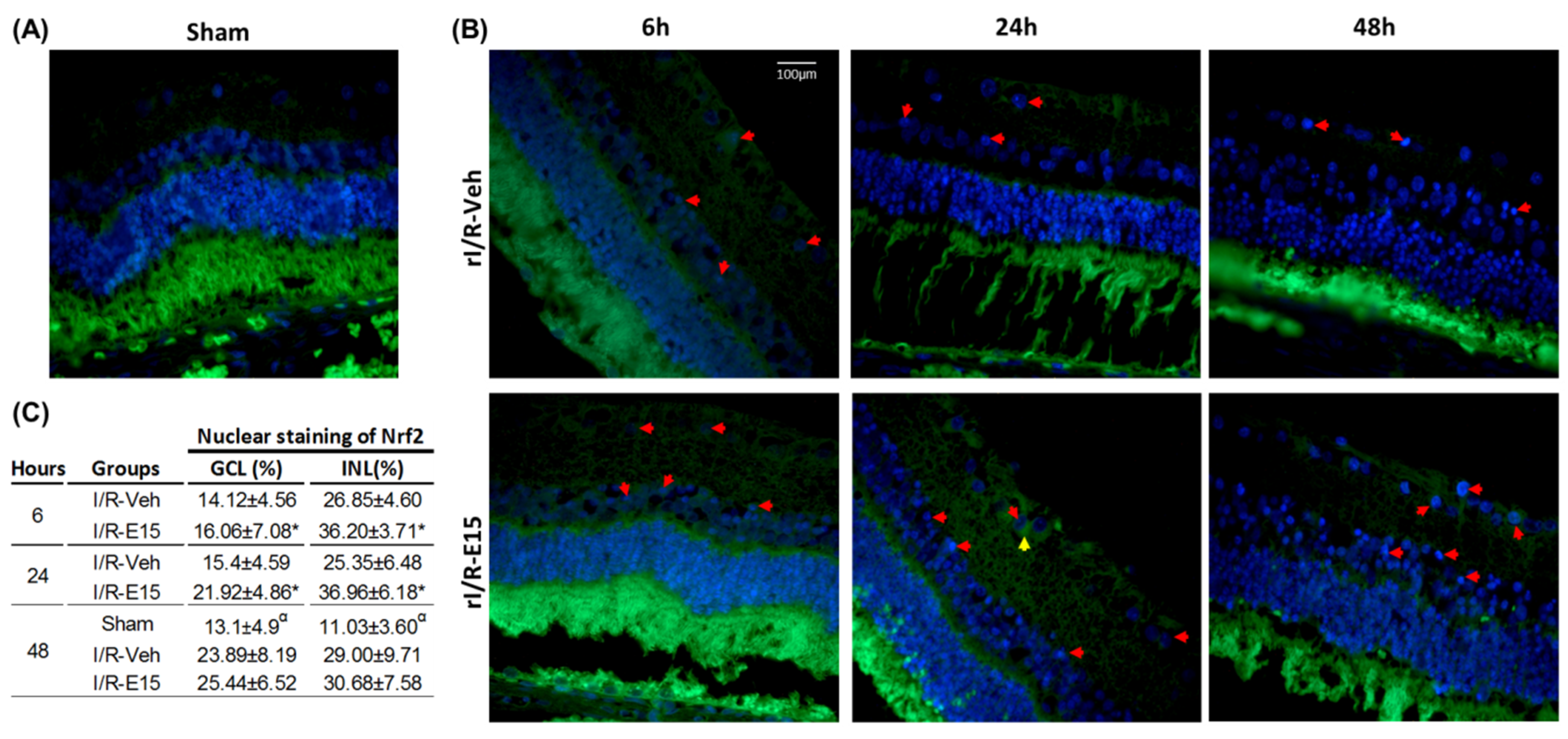
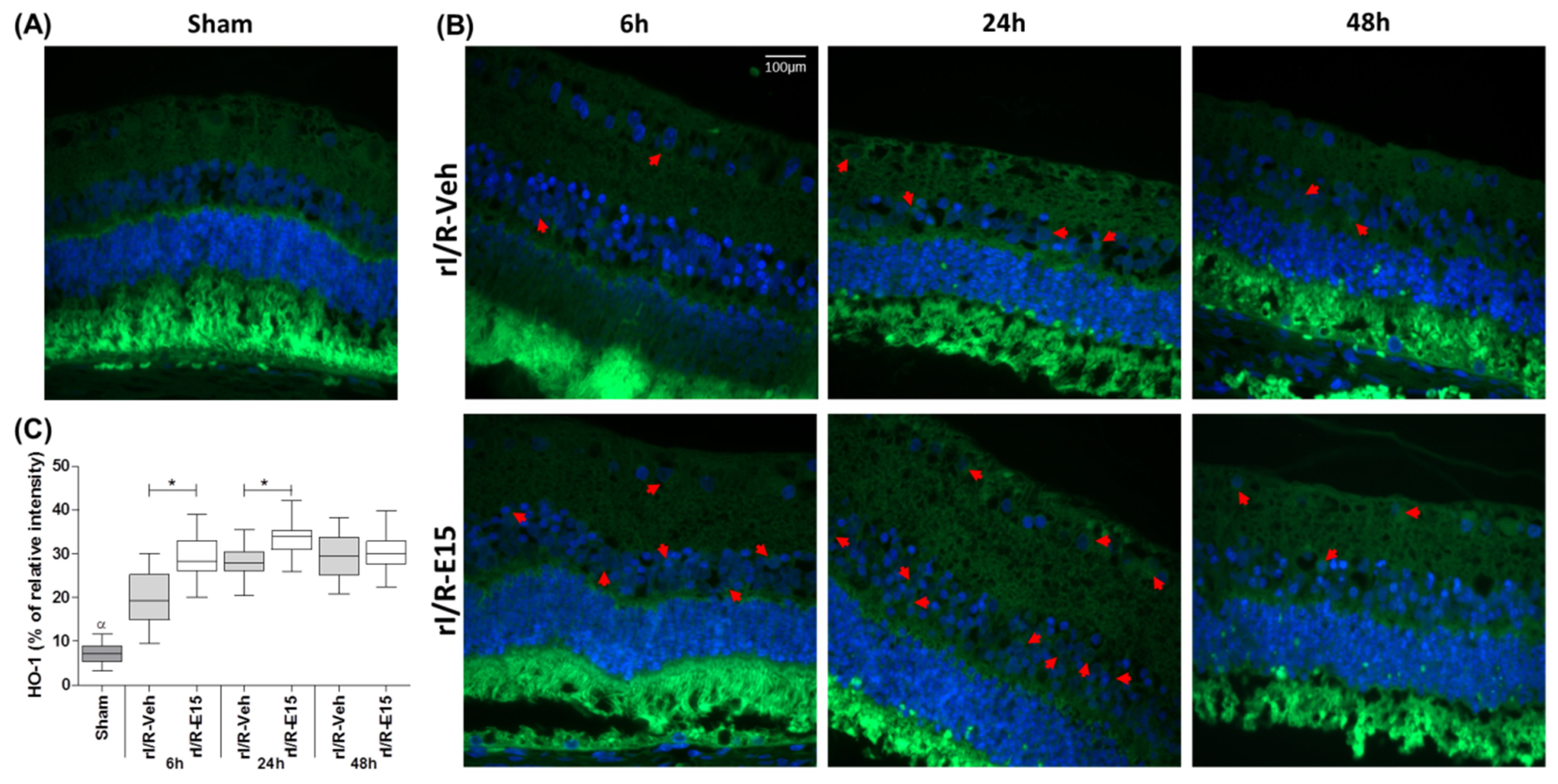
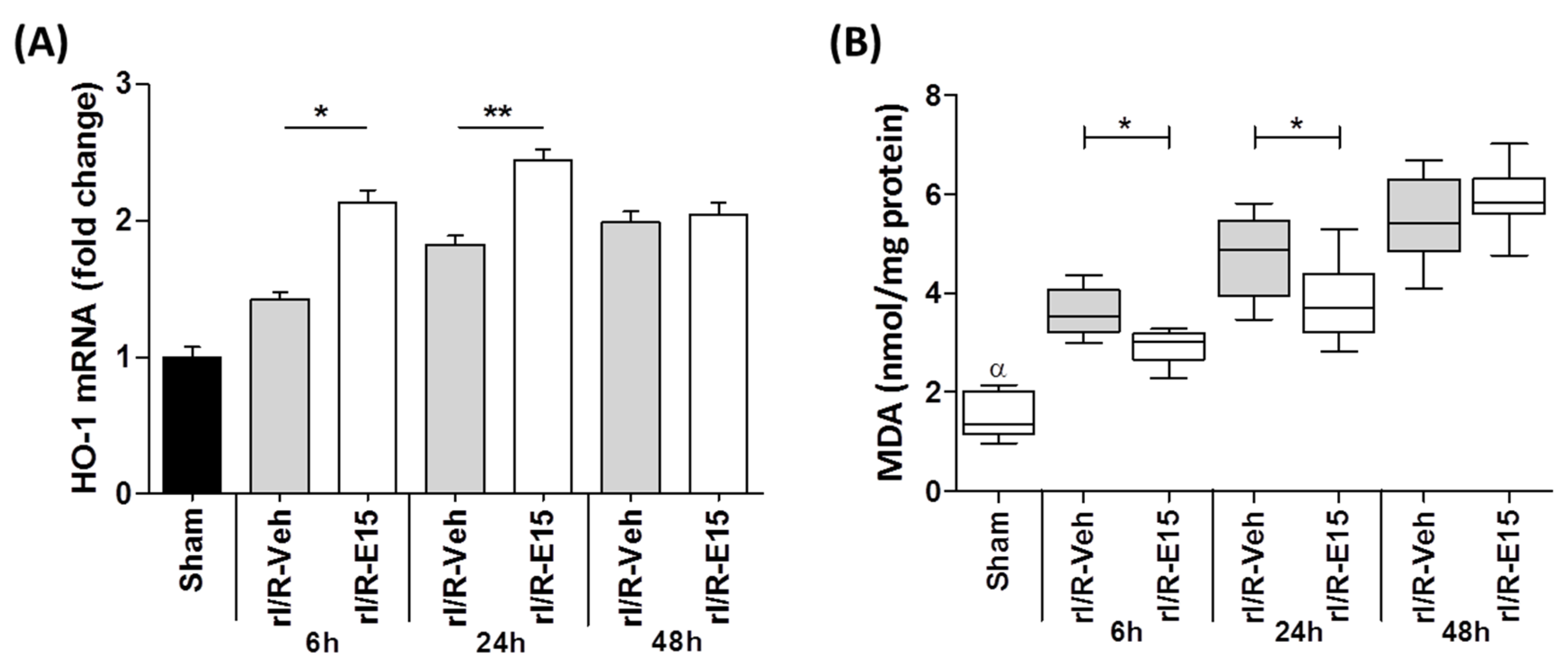
2.3. E15 Activates Nrf2/HO-1 Antioxidative Pathway in rI/R
2.4. Retinal Lipid Peroxidation in the Timeline
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Retinal Ischemia/Reperfusion (rI/R) Model
4.3. Histological Evaluation
4.4. Lipid Peroxidation Assay
4.5. Efficacy of EGCG Single-Dose Treatment on rI/R
4.6. Terminal Deoxynucleotidyl Transferase (TdT)-Mediated dUTP Nick End Labeling (TUNEL) Assay
4.7. Immunofluorescence
4.8. Semi-Quantitative PCR of Ho-1 mRNA
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| rI/R | Retinal ischemia/reperfusion |
| EGCG | Epigallocatechin-3-gallate |
| HMGB1 | High-mobility group Box 1 |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| HO-1 | Heme oxygenase-1 |
| GFAP | Glial fibrillary acidic protein |
| GCL | Ganglion cell layer |
| OPL | Outer plexiform layer |
| INL | Internal nuclear layer |
| IPL | Internal plexiform layer |
| ONL | Outer nuclear layer |
| PRL | Photoreceptor layer |
References
- Osborne, N.N.; Casson, R.J.; Wood, J.P.; Chidlow, G.; Graham, M.; Melena, J. Retinal ischemia: Mechanisms of damage and potential therapeutic strategies. Prog. Retin. Eye Res. 2004, 23, 91–147. [Google Scholar] [CrossRef] [PubMed]
- Toda, N.; Nakanishi-Toda, M. Nitric oxide: Ocular blood flow, glaucoma, and diabetic retinopathy. Prog. Retin. Eye Res. 2007, 26, 205–238. [Google Scholar] [CrossRef] [PubMed]
- Batliwala, S.; Xavier, C.; Liu, Y.; Wu, H.; Pang, I.H. Involvement of Nrf2 in Ocular Diseases. Oxid. Med. Cell. Longev. 2017, 2017, 1703810. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.G.; Xie, K.L.; Han, H.Z.; Wang, W.N.; Liu, D.Q.; Wang, G.L.; Yu, Y.H. Heme oxygenase-1 mediates the anti-inflammatory effect of molecular hydrogen in LPS-stimulated RAW 264.7 macrophages. Int. J. Surg. 2013, 11, 1060–1066. [Google Scholar] [CrossRef] [Green Version]
- Hanus, J.; Kolkin, A.; Chimienti, J.; Botsay, S.; Wang, S. 4-Acetoxyphenol Prevents RPE Oxidative Stress-Induced Necrosis by Functioning as an NRF2 Stabilizer. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5048–5059. [Google Scholar] [CrossRef] [PubMed]
- Paine, A.; Eiz-Vesper, B.; Blasczyk, R.; Immenschuh, S. Signaling to heme oxygenase-1 and its anti-inflammatory therapeutic potential. Biochem. Pharmacol. 2010, 80, 1895–1903. [Google Scholar] [CrossRef] [Green Version]
- Lambros, M.L.; Plafker, S.M. Oxidative Stress and the Nrf2 Anti-Oxidant Transcription Factor in Age-Related Macular Degeneration. Adv. Exp. Med. Biol. 2016, 854, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Gong, J.; Yoshida, T.; Eberhart, C.G.; Xu, Z.; Kombairaju, P.; Sporn, M.B.; Handa, J.T.; Duh, E.J. Nrf2 has a protective role against neuronal and capillary degeneration in retinal ischemia-reperfusion injury. Free Radic. Biol. Med. 2011, 51, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Peng, P.H.; Ko, M.L.; Chen, C.F. Epigallocatechin-3-gallate reduces retinal ischemia/reperfusion injury by attenuating neuronal nitric oxide synthase expression and activity. Exp. Eye Res. 2008, 86, 637–646. [Google Scholar] [CrossRef]
- Shen, C.; Chen, L.; Jiang, L.; Lai, T.Y. Neuroprotective effect of epigallocatechin-3-gallate in a mouse model of chronic glaucoma. Neurosci. Lett. 2015, 600, 132–136. [Google Scholar] [CrossRef]
- Xie, J.; Jiang, L.; Zhang, T.; Jin, Y.; Yang, D.; Chen, F. Neuroprotective effects of Epigallocatechin-3-gallate (EGCG) in optic nerve crush model in rats. Neurosci. Lett. 2010, 479, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Safa, R.; Rusciano, D.; Osborne, N.N. Epigallocatechin gallate, an active ingredient from green tea, attenuates damaging influences to the retina caused by ischemia/reperfusion. Brain Res. 2007, 1159, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.O.; Chan, K.P.; Yang, Y.P.; Qin, Y.J.; Li, W.Y.; Chan, S.O.; Wang, C.C.; Pang, C.P. Effects of EGCG content in green tea extract on pharmacokinetics, oxidative status and expression of inflammatory and apoptotic genes in the rat ocular tissues. J. Nutr. Biochem. 2015, 26, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Blanch, R.J.; Ahmed, Z.; Berry, M.; Scott, R.A.; Logan, A. Animal models of retinal injury. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2913–2920. [Google Scholar] [CrossRef] [Green Version]
- Heuss, N.D.; Pierson, M.J.; Roehrich, H.; McPherson, S.W.; Gram, A.L.; Li, L.; Gregerson, D.S. Optic nerve as a source of activated retinal microglia post-injury. Acta Neuropathol. Commun. 2018, 6, 66. [Google Scholar] [CrossRef] [Green Version]
- Oz, O.; Gurelik, G.; Akyurek, N.; Cinel, L.; Hondur, A. A short duration transient ischemia induces apoptosis in retinal layers: An experimental study in rabbits. Eur. J. Ophthalmol. 2005, 15, 233–238. [Google Scholar] [CrossRef]
- Polat, N.; Ozer, M.A.; Parlakpinar, H.; Vardi, N.; Aksungur, Z.; Ozhan, O.; Yildiz, A.; Turkoz, Y. Effects of molsidomine on retinal ischemia/reperfusion injury in rabbits. Biotech. Histochem. 2018, 93, 188–197. [Google Scholar] [CrossRef]
- Shen, J.; Yang, Q.; Yu, D.; Wu, J.; Zhu, Y.; Guo, W. Vulnerability study of myelinated and unmyelinated nerve fibers in acute ocular hypertension in rabbit. Mol. Med. Rep. 2017, 16, 6794–6802. [Google Scholar] [CrossRef] [Green Version]
- Uckermann, O.; Uhlmann, S.; Pannicke, T.; Francke, M.; Gamsalijew, R.; Makarov, F.; Ulbricht, E.; Wiedemann, P.; Reichenbach, A.; Osborne, N.N.; et al. Ischemia-reperfusion causes exudative detachment of the rabbit retina. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2592–2600. [Google Scholar] [CrossRef]
- Ren, J.; Fan, C.; Chen, N.; Huang, J.; Yang, Q. Resveratrol pretreatment attenuates cerebral ischemic injury by upregulating expression of transcription factor Nrf2 and HO-1 in rats. Neurochem. Res. 2011, 36, 2352–2362. [Google Scholar] [CrossRef]
- Chen, F.; Jiang, L.; Shen, C.; Wan, H.; Xu, L.; Wang, N.; Jonas, J.B. Neuroprotective effect of epigallocatechin-3-gallate against N-methyl-D-aspartate-induced excitotoxicity in the adult rat retina. Acta Ophthalmol. 2012, 90, e609–e615. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, M.; Jing, X.; Shi, H.; Ren, M.; Lou, H. (-)-Epigallocatechin gallate protects against cerebral ischemia-induced oxidative stress via Nrf2/ARE signaling. Neurochem. Res. 2014, 39, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Cheng-Chung Wei, J.; Huang, H.C.; Chen, W.J.; Huang, C.N.; Peng, C.H.; Lin, C.L. Epigallocatechin gallate attenuates amyloid beta-induced inflammation and neurotoxicity in EOC 13.31 microglia. Eur. J. Pharmacol. 2016, 770, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, B.; Jayavelu, S.; Murhekar, K.; Rajkumar, T. Repeated dose studies with pure Epigallocatechin-3-gallate demonstrated dose and route dependant hepatotoxicity with associated dyslipidemia. Toxicol. Rep. 2016, 3, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Nan, W.; Zhonghang, X.; Keyan, C.; Tongtong, L.; Wanshu, G.; Zhongxin, X. Epigallocatechin-3-Gallate Reduces Neuronal Apoptosis in Rats after Middle Cerebral Artery Occlusion Injury via PI3K/AKT/eNOS Signaling Pathway. BioMed Res. Int. 2018, 2018, 6473580. [Google Scholar] [CrossRef] [Green Version]
- Rahman, R.M.; Nair, S.M.; Helps, S.C.; Shaw, O.M.; Sims, N.R.; Rosengren, R.J.; Appleton, I. (-)-Epigallocatechin gallate as an intervention for the acute treatment of cerebral ischemia. Neurosci. Lett. 2005, 382, 227–230. [Google Scholar] [CrossRef]
- Peng, P.H.; Ko, M.L.; Chen, C.F.; Juan, S.H. Haem oxygenase-1 gene transfer protects retinal ganglion cells from ischaemia/reperfusion injury. Clin. Sci. (Lond.) 2008, 115, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Cho, H.; Hartsock, M.J.; Mitchell, K.L.; Gong, J.; Wu, L.; Wei, Y.; Wang, S.; Thimmulappa, R.K.; Sporn, M.B.; et al. Neuroprotective role of Nrf2 for retinal ganglion cells in ischemia-reperfusion. J. Neurochem. 2015, 133, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Liu, F.; Liu, X.; Huang, M.; He, L.; Ramakrishna, S.; Luo, H.; Hu, H.; Li, H.; Xu, Y. Methyl 3,4-dihydroxybenzoate protects retina in a mouse model of acute ocular hypertension through multiple pathways. Exp. Eye Res. 2019, 181, 15–24. [Google Scholar] [CrossRef]
- Nakagami, Y.; Masuda, K.; Hatano, E.; Inoue, T.; Matsuyama, T.; Iizuka, M.; Ono, Y.; Ohnuki, T.; Murakami, Y.; Iwasaki, M.; et al. Novel Nrf2 activators from microbial transformation products inhibit blood-retinal barrier permeability in rabbits. Br. J. Pharmacol. 2015, 172, 1237–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.; He, M.; Liu, R.; Brecha, N.C.; Yu, A.C.; Pu, M. Sulforaphane protects rodent retinas against ischemia-reperfusion injury through the activation of the Nrf2/HO-1 antioxidant pathway. PLoS ONE 2014, 9, e114186. [Google Scholar] [CrossRef] [PubMed]
- Panickar, K.S.; Qin, B.; Anderson, R.A. Ischemia-induced endothelial cell swelling and mitochondrial dysfunction are attenuated by cinnamtannin D1, green tea extract, and resveratrol in vitro. Nutr. Neurosci. 2015, 18, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Anavy, N.D.; Chow, H.H. Contribution of presystemic hepatic extraction to the low oral bioavailability of green tea catechins in rats. Drug Metab. Dispos. 2002, 30, 1246–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swezey, R.R.; Aldridge, D.E.; LeValley, S.E.; Crowell, J.A.; Hara, Y.; Green, C.E. Absorption, tissue distribution and elimination of 4-[(3)h]-epigallocatechin gallate in beagle dogs. Int. J. Toxicol. 2003, 22, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, U.; Haller, J.; Decourt, J.D.; Girault, J.; Spitzer, V.; Weber, P. Plasma-kinetic characteristics of purified and isolated green tea catechin epigallocatechin gallate (EGCG) after 10 days repeated dosing in healthy volunteers. Int. J. Vitam. Nutr. Res. 2004, 74, 269–278. [Google Scholar] [CrossRef]
- Chi, W.; Chen, H.; Li, F.; Zhu, Y.; Yin, W.; Zhuo, Y. HMGB1 promotes the activation of NLRP3 and caspase-8 inflammasomes via NF-kappaB pathway in acute glaucoma. J. Neuroinflamm. 2015, 12, 137. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhu, S.; Li, J.; Assa, A.; Jundoria, A.; Xu, J.; Fan, S.; Eissa, N.T.; Tracey, K.J.; Sama, A.E.; et al. EGCG stimulates autophagy and reduces cytoplasmic HMGB1 levels in endotoxin-stimulated macrophages. Biochem. Pharmacol. 2011, 81, 1152–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Huang, X.; Shen, D.; Ming, Z.; Zheng, M.; Zhang, J. Polyphenol epigallocatechin-3-gallate inhibits hypoxia/reoxygenation-induced H9C2 cell apoptosis. Minerva Med. 2018, 109, 95–102. [Google Scholar] [CrossRef]
- Yang, G.Z.; Wang, Z.J.; Bai, F.; Qin, X.J.; Cao, J.; Lv, J.Y.; Zhang, M.S. Epigallocatechin-3-gallate protects HUVECs from PM2.5-induced oxidative stress injury by activating critical antioxidant pathways. Molecules 2015, 20, 6626–6639. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Yu, R.; Owuor, E.D.; Kong, A.N. Activation of antioxidant-response element (ARE), mitogen-activated protein kinases (MAPKs) and caspases by major green tea polyphenol components during cell survival and death. Arch. Pharm. Res. 2000, 23, 605–612. [Google Scholar] [CrossRef]
- Vestergaard, N.; Cehofski, L.J.; Honoré, B.; Aasbjerg, K.; Vorum, H. Animal Models Used to Simulate Retinal Artery Occlusion: A Comprehensive Review. Transl. Vis. Sci. Technol. 2019, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarrete Santos, A.; Tonack, S.; Kirstein, M.; Pantaleon, M.; Kaye, P.; Fischer, B. Insulin acts via mitogen-activated protein kinase phosphorylation in rabbit blastocysts. Reproduction 2004, 128, 517–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; He, Z.; Wu, L.; Fang, Y. Effects of induction/inhibition of endogenous heme oxygenase-1 on lipid metabolism, endothelial function, and atherosclerosis in rabbits on a high fat diet. J. Pharmacol. Sci. 2012, 118, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igarashi, T.; Ohsawa, I.; Kobayashi, M.; Igarashi, T.; Suzuki, H.; Iketani, M.; Takahashi, H. Hydrogen prevents corneal endothelial damage in phacoemulsification cataract surgery. Sci. Rep. 2016, 6, 31190. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Pérez, J.; Martínez-Rosas, M.; Conde-Castañón, C.A.; Toscano-Garibay, J.D.; Ruiz-Pérez, N.J.; Flores, P.L.; Mera Jiménez, E.; Flores-Estrada, J. Epigallocatechin 3-Gallate Has a Neuroprotective Effect in Retinas of Rabbits with Ischemia/Reperfusion through the Activation of Nrf2/HO-1. Int. J. Mol. Sci. 2020, 21, 3716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103716
Rivera-Pérez J, Martínez-Rosas M, Conde-Castañón CA, Toscano-Garibay JD, Ruiz-Pérez NJ, Flores PL, Mera Jiménez E, Flores-Estrada J. Epigallocatechin 3-Gallate Has a Neuroprotective Effect in Retinas of Rabbits with Ischemia/Reperfusion through the Activation of Nrf2/HO-1. International Journal of Molecular Sciences. 2020; 21(10):3716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103716
Chicago/Turabian StyleRivera-Pérez, Josué, Martín Martínez-Rosas, César A. Conde-Castañón, Julia D. Toscano-Garibay, Nancy J. Ruiz-Pérez, Pedro L. Flores, Elvia Mera Jiménez, and Javier Flores-Estrada. 2020. "Epigallocatechin 3-Gallate Has a Neuroprotective Effect in Retinas of Rabbits with Ischemia/Reperfusion through the Activation of Nrf2/HO-1" International Journal of Molecular Sciences 21, no. 10: 3716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103716