Development and Characterization of an Allergic Asthma Rat Model for Interventional Studies

 , , ,
, , ,  ,
,  and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
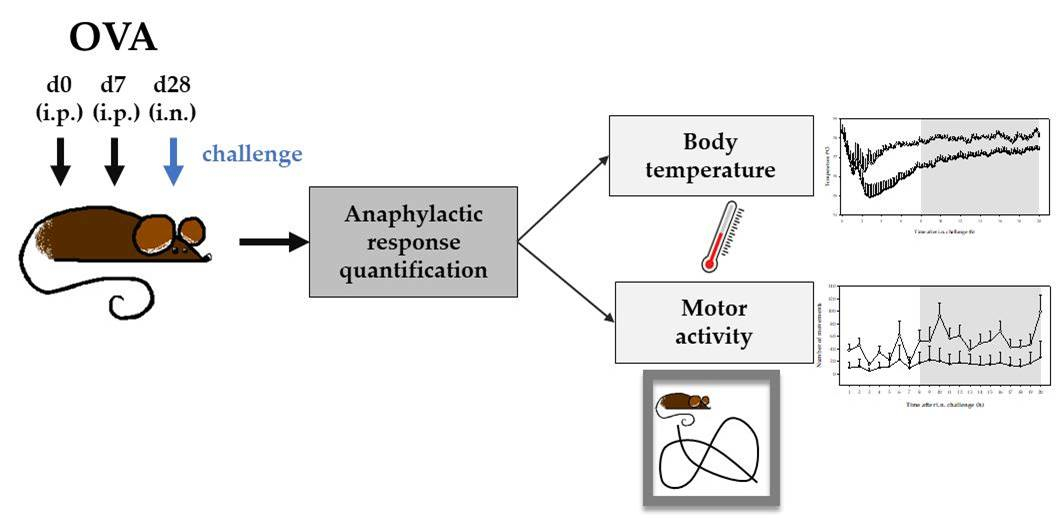
1. Introduction
2. Results
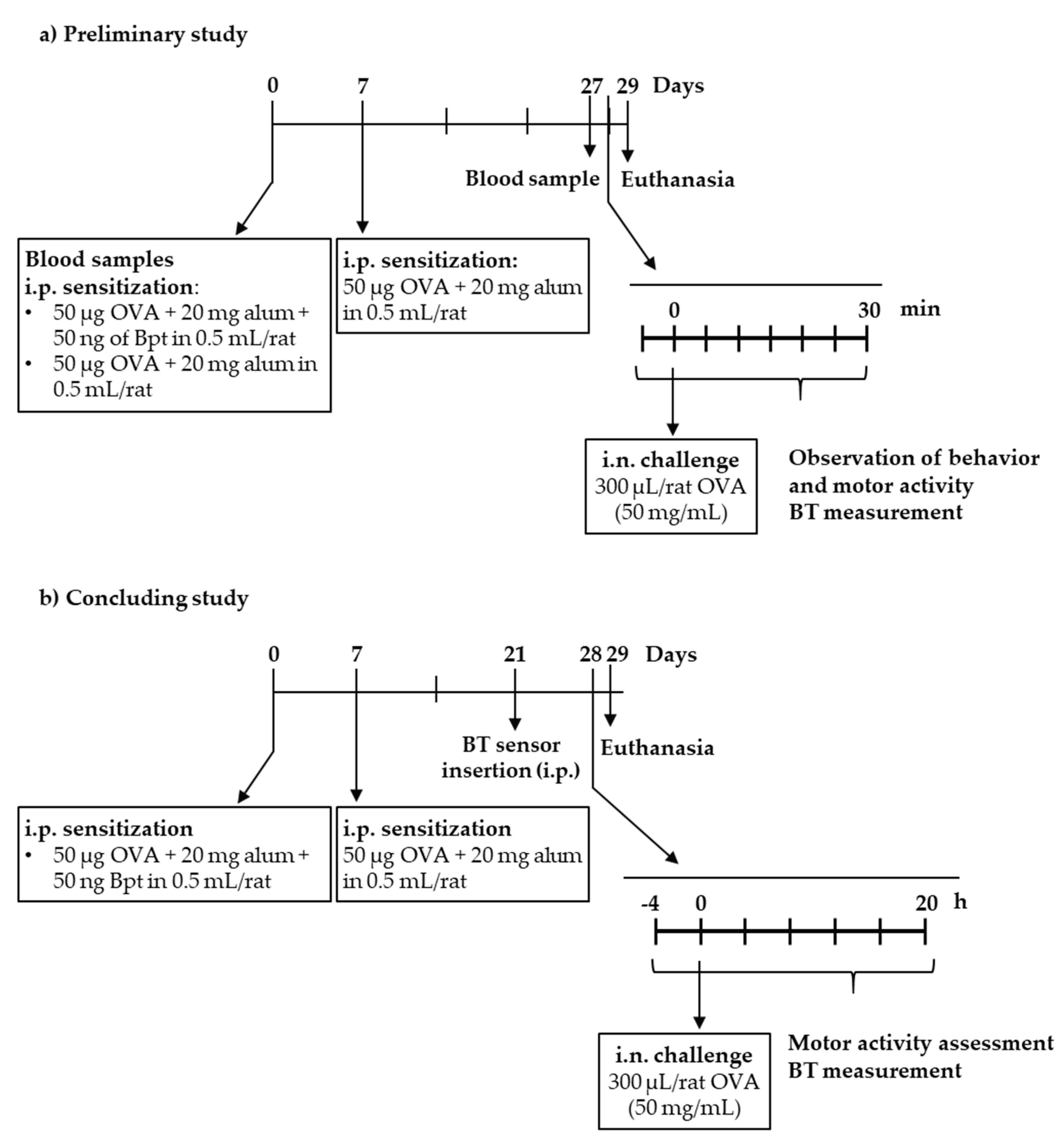
2.1. Preliminary Study
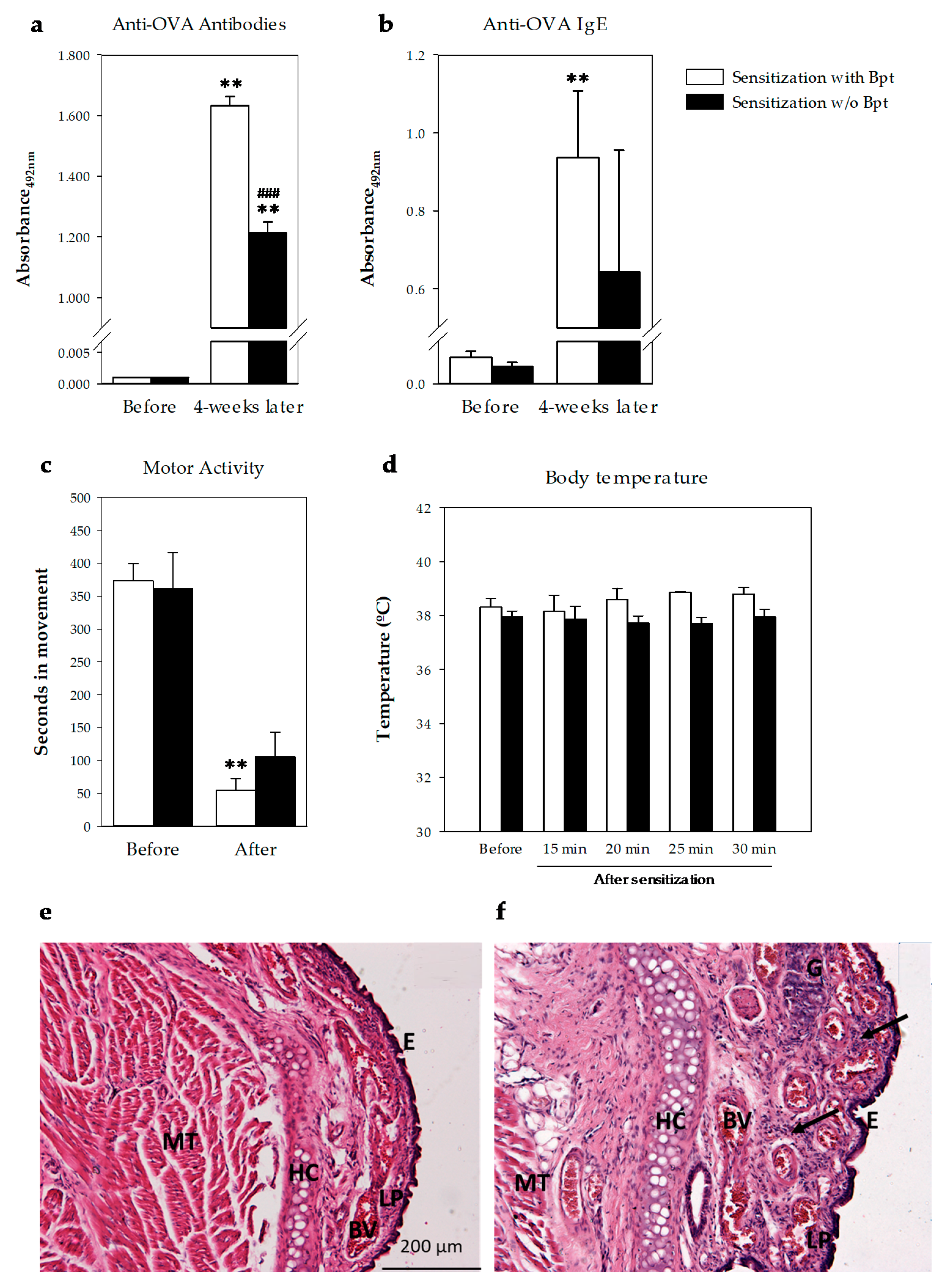
2.1.1. Serum Anti-OVA IgE Antibodies
2.1.2. Intranasal-Induced Shock Quantification
2.1.3. Respiratory Airway Histology
2.2. Definitive Study
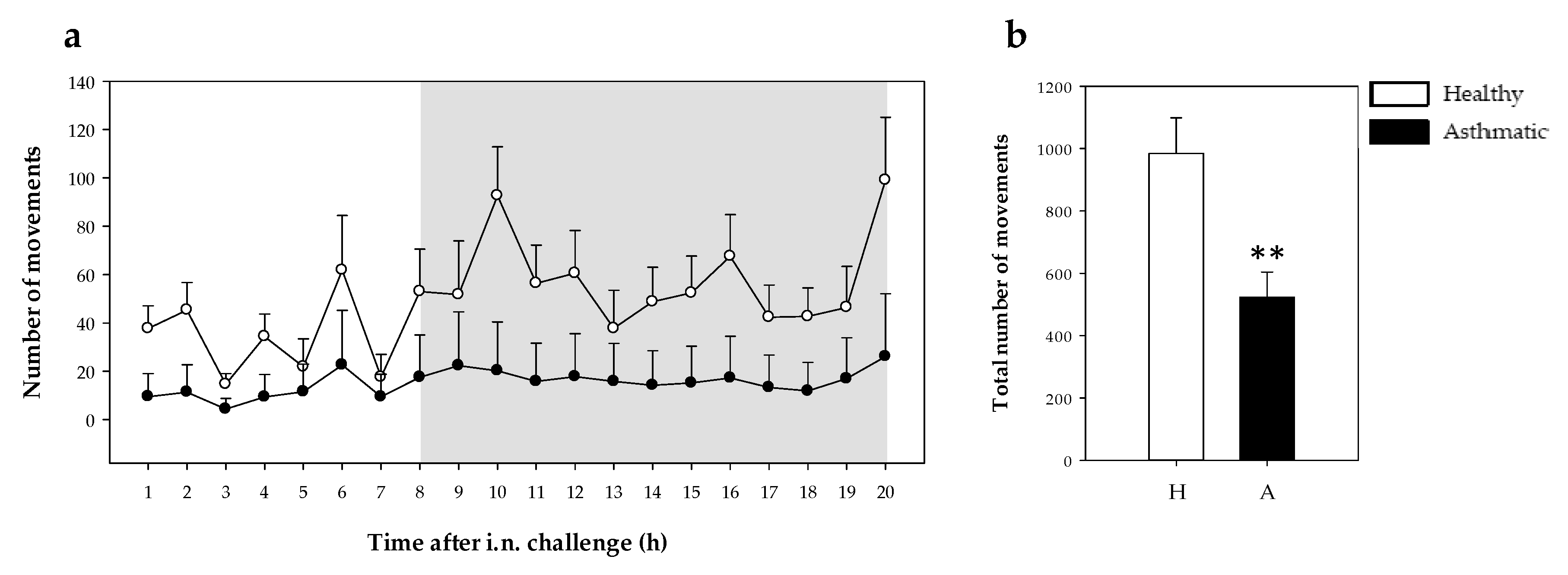
2.2.1. Motor Activity
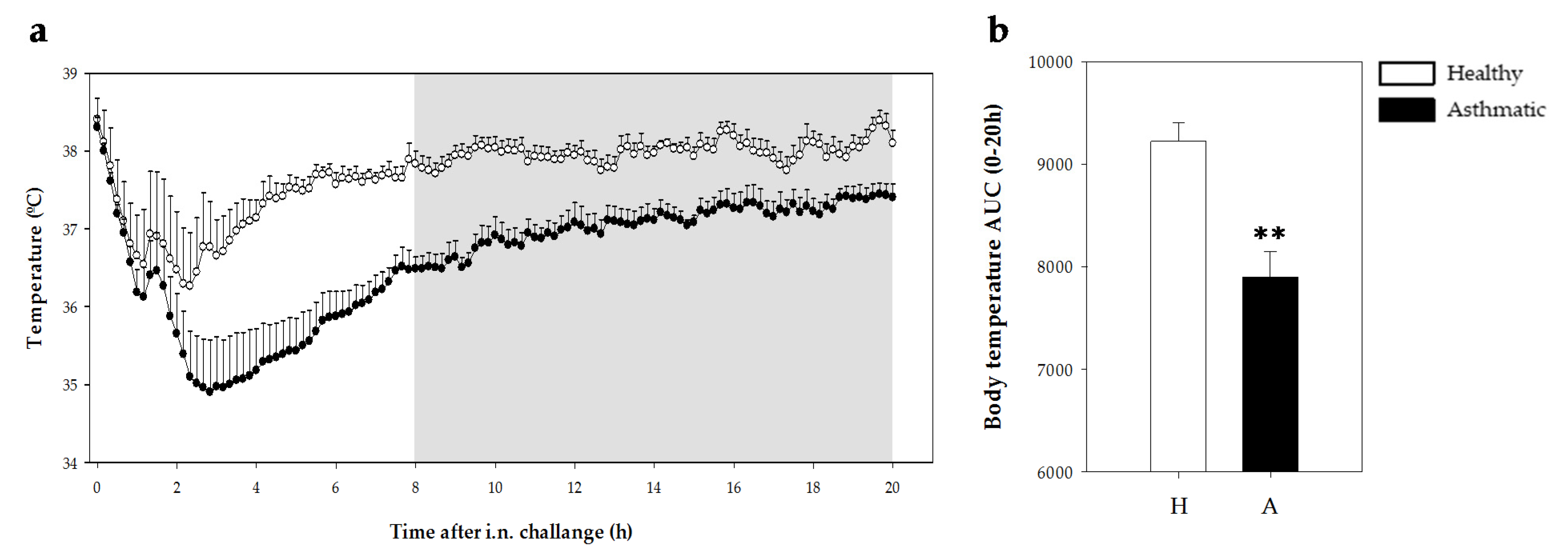
2.2.2. Temperature
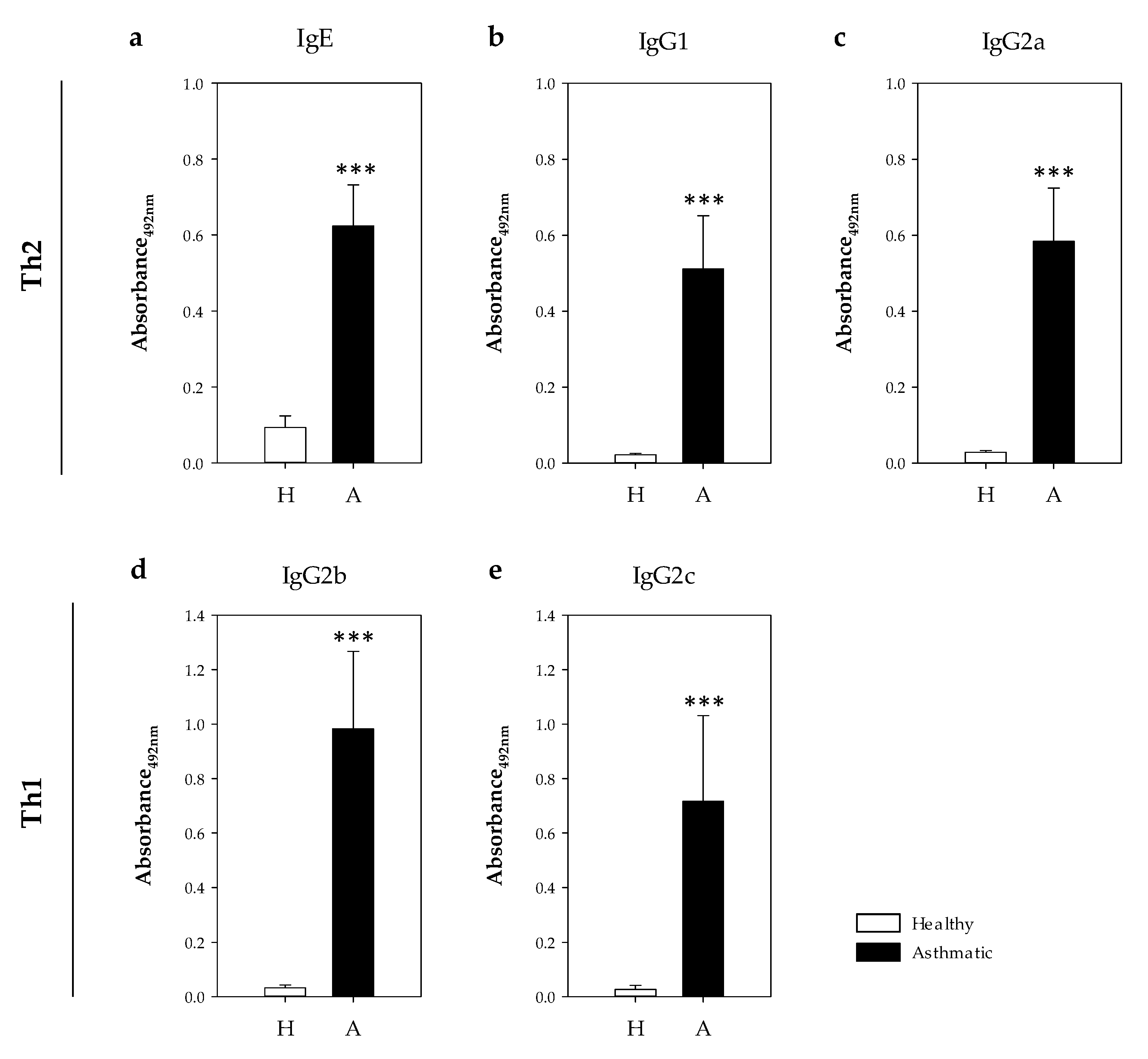
2.2.3. Serum Anti-OVA Antibodies.
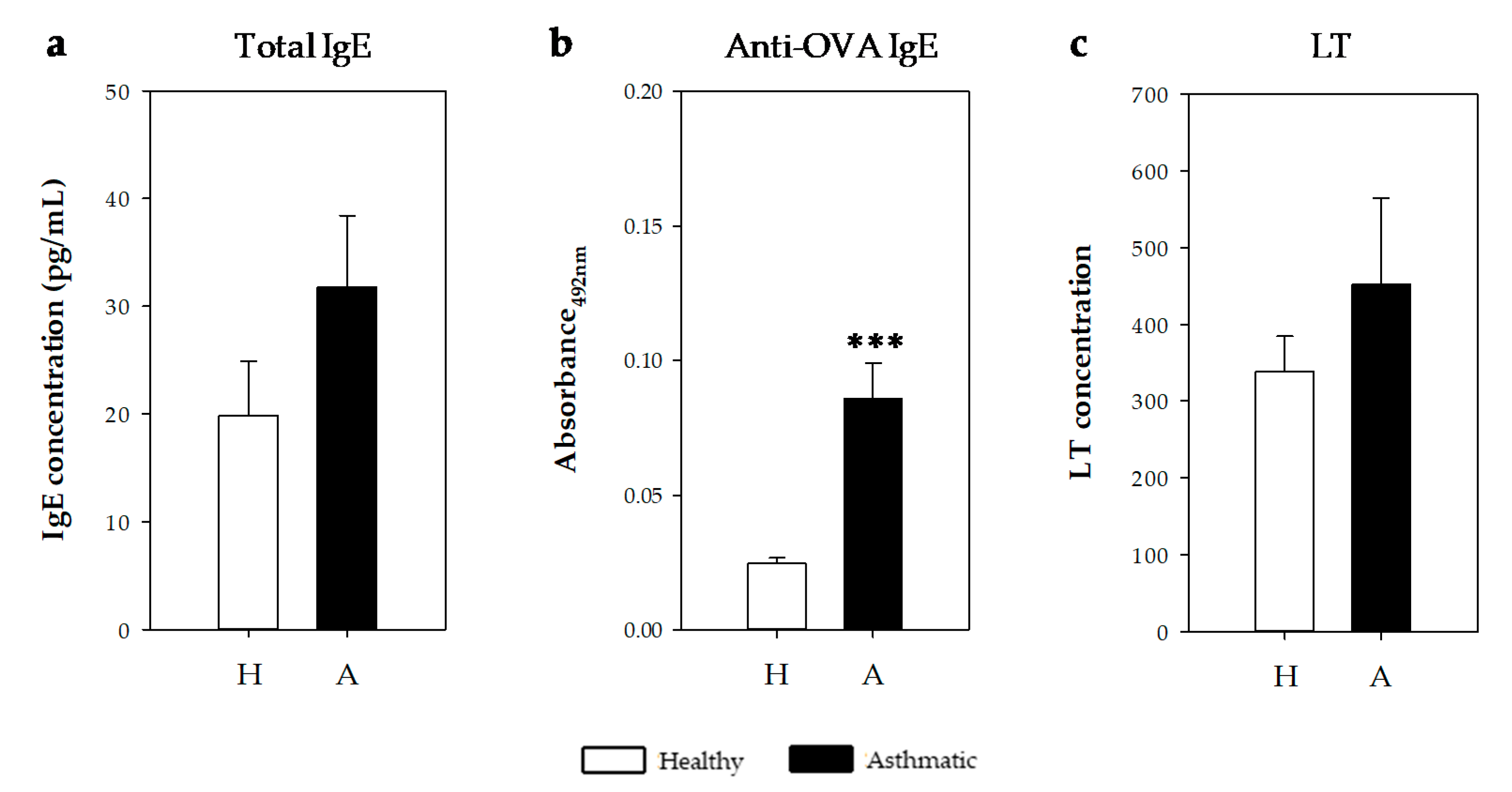
2.2.4. IgE Antibodies and Leukotrienes in Bronchoalveolar Lavage Fluid
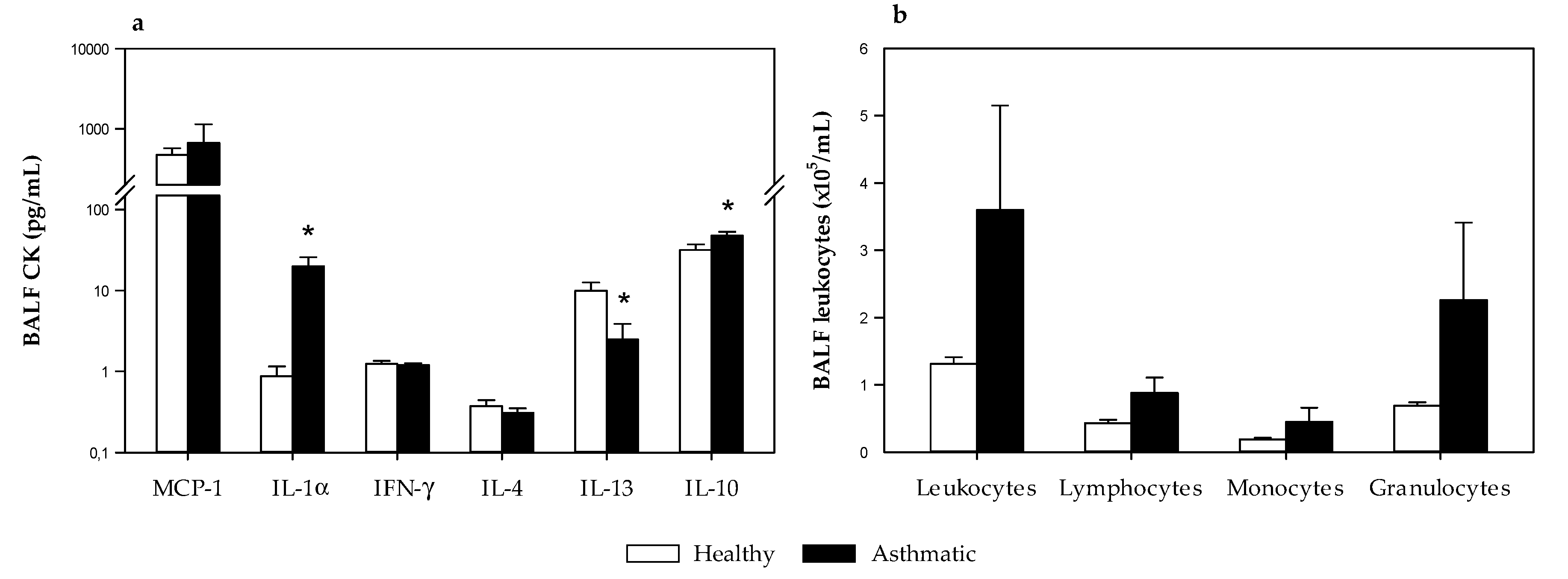
2.2.5. Cytokine and Leukocytes in Bronchoalveolar Lavage Fluid
3. Discussion
4. Materials and Methods
4.1. Animals and Ethical Issues
4.2. Experimental Designs
4.3. Motor Activity Assessment
4.4. Temperature Measurement
4.5. Blood Collection
4.6. BALF Collection
4.7. IgE and Anti-OVA Antibodies Quantification
4.8. Histological Study
4.9. Quantification of Cysteinyl Leukotriene (CysLT)
4.10. Quantification of Cytokines
4.11. Leukocytes in BALF
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AUC | Area under the curve |
| BALF | Bronchoalveolar lavage fluid |
| Bpt | Bordetella pertussis toxin |
| CysLT | Cysteinyl leukotriene |
| ELISA | Enzyme linked immunosorbent assay |
| IL | Interleukin |
| i.n. | Intranasal |
| IFN | Interferon |
| i.p. | Intraperitoneal |
| MCP-1 | Monocyte chemoattractant protein 1 |
| OVA | Ovalbumin |
| Th | T helper |
References
- World Health Organization. Prevention of Allergy and Allergic Asthma; World Health Organization: Geneva, Switzerland, 2003; Available online: https://apps.who.int/iris/handle/10665/68361 (accessed on 3 March 2020).
- Papi, A.; Brightling, C.; Pedersen, S.E.; Reddel, H.K. Asthma. Lancet 2018, 391, 783–800. [Google Scholar] [CrossRef]
- Agache, I. Severe asthma phenotypes and endotypes. Semin. Immunol. 2019, 46, 101301. [Google Scholar] [CrossRef] [PubMed]
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 Diseases and Injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- Rehman, A.; Amin, F.; Sadeeqa, S. Prevalence of asthma and its management: A review. J. Pak. Med. Assoc. 2018, 68, 1823–1827. [Google Scholar] [PubMed]
- Shin, Y.S.; Takeda, K.; Gelfand, E.W. Understanding asthma using animal models. Allergy Asthma Immunol. Res. 2009, 1, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucharewicz, I.; Bodzenta-łukaszyk, A.; Bucko, W. Experimental asthma in rats. Pharmacol. Rep. 2008, 60, 783–788. [Google Scholar]
- Rosenberg, H.F.; Druey, K.M. Modeling asthma: Pitfalls, promises, and the road ahead. J. Leukoc. Biol. 2018, 104, 41–48. [Google Scholar] [CrossRef]
- Russjan, E.; Kaczyńska, K. Murine models of hapten-induced asthma. Toxicology 2018, 410, 41–48. [Google Scholar] [CrossRef]
- Zosky, G.R.; Sly, P.D. Animal models of asthma. Clin. Exp. Allergy 2007, 37, 973–988. [Google Scholar] [CrossRef]
- Aun, M.V.; Bonamichi-Santos, R.; Arantes-Costa, F.M.; Kalil, J.; Giavina-Bianchi, P. Animal models of asthma: Utility and limitations. J. Asthma Allergy 2017, 10, 293–301. [Google Scholar] [CrossRef] [Green Version]
- Kianmehr, M.; Ghorani, V.; Boskabady, M.H. Animal model of asthma, various methods and measured parameters, a methodological review. Iran. J. Allergy Asthma Immunol. 2016, 15, 445–465. [Google Scholar]
- Pauluhn, J.; Mohr, U. Experimental approaches to evaluate respiratory allergy in animal models. Exp. Toxicol. Pathol. 2005, 56, 203–234. [Google Scholar] [CrossRef] [PubMed]
- Elias, J.A.; Lee, C.G.; Zheng, T.; Ma, B.; Homer, R.J.; Zhu, Z. New insights into the pathogenesis of asthma. J. Clin. Investig. 2003, 111, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Camps-Bossacoma, M.; Abril-Gil, M.; Franch, A.; Perez-Cano, F.; Castell, M. Induction of an oral sensitization model in rats. Clin. Immunol. Endocr. Metab. Drugs 2015, 1, 89–101. [Google Scholar] [CrossRef]
- Li, Y.; Chen, L.; Guo, F.; Cao, Y.; Hu, W.; Shi, Y.; Lin, X.; Hou, J.; Li, L.; Ding, X.; et al. Biochemical and biophysical research communications effects of epigallocatechin-3-gallate on the HMGB1/RAGE pathway in PM 2.5-exposed asthmatic rats. Biochem. Biophys. Res. Commun. 2019, 513, 898–903. [Google Scholar] [CrossRef]
- Abril-Gil, M.; Garcia-Just, A.; Pérez-Cano, F.J.; Franch, À.; Castell, M. Development and characterization of an effective food allergy model in Brown Norway rats. PLoS ONE 2015, 10, e0125314. [Google Scholar] [CrossRef] [Green Version]
- Zainal, Z.; Rahim, A.A.; Khaza’ai, H.; Chang, S.K. Effects of palm oil tocotrienol-rich fraction (TRF) and carotenes in ovalbumin (OVA)-challenged asthmatic brown Norway rats. Int. J. Mol. Sci. 2019, 20, 1764. [Google Scholar] [CrossRef] [Green Version]
- Knippels, L.M.J.; Penninks, A.H.; van Meeteren, M.; Houben, G.F. Humoral and cellular immune responses in different rat strains on oral exposure to ovalbumin. Food Chem. Toxicol. 1999, 37, 881–888. [Google Scholar] [CrossRef]
- Smole, U.; Schabussova, I.; Pickl, W.F.; Wiedermann, U. Murine models for mucosal tolerance in allergy. Semin. Immunol. 2017, 30, 12–27. [Google Scholar] [CrossRef]
- Sun, N.; Zhou, C.; Pu, Q.; Wang, J.; Huang, K.; Che, H. Allergic reactions compared between BN and Wistar rats after oral exposure to ovalbumin. J. Immunotoxicol. 2013, 10, 67–74. [Google Scholar] [CrossRef]
- Lindholm Bøgh, K.; Barkholt, V.; Bernhard Madsen, C. The sensitising capacity of intact β-lactoglobulin is reduced by Co-administration with digested β-lactoglobulin. Int. Arch. Allergy Immunol. 2013, 161, 21–36. [Google Scholar] [CrossRef]
- Curtis, J.L. Cell-mediated adaptive immune defense of the lungs. Proc. Am. Thorac. Soc. 2005, 2, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Abril-Gil, M.; Massot-Cladera, M.; Pérez-Cano, F.J.; Castellote, C.; Franch, A.; Castell, M. A diet enriched with cocoa prevents IgE synthesis in a rat allergy model. Pharmacol. Res. 2012, 65, 603–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knippels, L.M.J.; Penninks, A.H. Recent advances using rodent models for predicting human allergenicity. Toxicol. Appl. Pharmacol. 2005, 207, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, N.; Xiao, J.; Wang, Y.H.; Dong, R. The amygdala via the paraventricular nucleus regulates asthma attack in rats. CNS Neurosci. Ther. 2020, 1–11. [Google Scholar] [CrossRef]
- Guan, Y.; Shen, H.; Shen, J.; Jia, Y.; Dong, X.; Jin, Y.; Xie, Q. Anti-allergic activities of 5,7-dimethoxy-3,4’-dihydroxyflavone via inhalation in rat allergic models. Eur. J. Pharmacol. 2019, 848, 55–61. [Google Scholar] [CrossRef]
- Kucharewicz, I.; Kasacka, I.; Pawlak, D.; Tankiewicz-Kwedlo, A.; Mroczko, B.; Buczko, W.; Bodzenta-Lukaszyk, A. The concentration of kynurenine in rat model of asthma. Folia Histochem. Cytobiol. 2008, 46, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, P.; Pathak, M.P.; Patowary, P.; Chakrabarti, S.; Goyary, D.; Karmakar, S.; Dwivedi, S.K. Synthesized atropine nanoparticles ameliorate airway hyperreactivity and remodeling in a murine model of chronic asthma. J. Drug Deliv. Sci. Technol. 2020, 56, 101507. [Google Scholar] [CrossRef]
- Kaveh, M.; Eftekhar, N.; Boskabady, M.H. The effect of alpha linolenic acid on tracheal responsiveness, lung inflammation and immune markers in sensitized rats. Iran. J. Basic Med. Sci. 2019, 22, 255–261. [Google Scholar]
- Hanna, D.A.; Khalaf, M.M.; Abo-Saif, A.A. Polydatin protects against ovalbumin-induced bronchial asthma in rats; involvement of urocortin and surfactant-D expression. Immunopharmacol. Immunotoxicol. 2019, 41, 403–412. [Google Scholar] [CrossRef]
- Thakur, V.R.; Khuman, V.; Beladiya, J.V.; Chaudagar, K.K.; Mehta, A.A. An experimental model of asthma in rats using ovalbumin and lipopolysaccharide allergens. Heliyon 2019, 5, e02864. [Google Scholar] [CrossRef]
- Zemmouri, H.; Sekiou, O.; Ammar, S.; El Feki, A.; Bouaziz, M.; Messarah, M.; Boumendjel, A. Urtica dioica attenuates ovalbumin-induced inflammation and lipid peroxidation of lung tissues in rat asthma model. Pharm. Biol. 2017, 55, 1561–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlajković, S.; Dugandzija-Novaković, S.; Milanović, S.; Janković, B.D. Brain self-stimulation and immunity: Effect on humoral and cell-mediated immune responses. Int. J. Neurosci. 1993, 69, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Michael, K.C.; Bonneau, R.H.; Bourne, R.A.; Godbolt, L.D.; Caruso, M.J.; Hohmann, C.; Cavigelli, S.A. Divergent immune responses in behaviorally-inhibited vs. non-inhibited male rats. Physiol. Behav. 2020, 213, 112693. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, L.H.; Katz, M.; Kovacsics, C.E.; Gould, T.D.; Joppy, B.; Hoshino, A.; Hoffman, G.; Komarow, H.; Postolache, T.T. Allergic rhinitis induces anxiety-like behavior and altered social interaction in rodents. Brain Behav. Immun. 2009, 23, 784–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.M.; Serebrisky, D.; Lee, S.Y.; Huang, C.K.; Bardina, L.; Schofield, B.H.; Stanley, J.S.; Burks, A.W.; Bannon, G.A.; Sampson, H.A. A murine model of peanut anaphylaxis: T- and B-cell responses to a major peanut allergen mimic human responses. J. Allergy Clin. Immunol. 2000, 106, 150–158. [Google Scholar] [CrossRef]
- Abril-Gil, M.; Garcia-Just, A.; Cambras, T.; Pérez-Cano, F.J.; Castellote, C.; Franch, À.; Castell, M. Motor activity as an unbiased variable to assess anaphylaxis in allergic rats. Exp. Biol. Med. 2015, 240, 1373–1377. [Google Scholar] [CrossRef] [Green Version]
- Giménez-Rivera, V.; Metz, M.; Siebenhaar, F. Mast cell-mediated reactions in vivo. In Methods in Molecular Biology (Methods and Protocols); Gibbs, B., Falcone, F., Eds.; Humana Press: New York, NY, USA, 2014; Volume 1192, pp. 239–247. [Google Scholar]
- Cambras, T.; Castejón, L.; Díez-Noguera, A. Social interaction with a rhythmic rat enhances the circadian pattern of the motor activity and temperature of LL-induced arrhythmic rats. Physiol. Behav. 2012, 105, 835–840. [Google Scholar] [CrossRef]
- Moura, C.T.M.; Bezerra, F.C.; De Moraes, I.M.I.; Magalhães, P.J.C.; Capaz, F.R. Increased responsiveness to 5-hydroxytryptamine after antigenic challenge is inhibited by nifedipine and niflumic acid in rat trachea in vitro. Clin. Exp. Pharmacol. Physiol. 2005, 32, 1119–1123. [Google Scholar] [CrossRef]
- Boonpiyathad, T.; Celebi Sözener, Z.; Satitsuksonoa, P.; Akdis, C.A. Immunological mechanisms in asthma. Semin. Immunol. 2019, 46, 101333. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Périz, M.; Pérez-Cano, F.J.; Rodríguez-Lagunas, M.J.; Cambras, T.; Pastor-Soplin, S.; Best, I.; Castell, M.; Massot-Cladera, M. Development and Characterization of an Allergic Asthma Rat Model for Interventional Studies. Int. J. Mol. Sci. 2020, 21, 3841. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113841
Périz M, Pérez-Cano FJ, Rodríguez-Lagunas MJ, Cambras T, Pastor-Soplin S, Best I, Castell M, Massot-Cladera M. Development and Characterization of an Allergic Asthma Rat Model for Interventional Studies. International Journal of Molecular Sciences. 2020; 21(11):3841. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113841
Chicago/Turabian StylePériz, Marta, Francisco J. Pérez-Cano, Maria J. Rodríguez-Lagunas, Trinitat Cambras, Santiago Pastor-Soplin, Iván Best, Margarida Castell, and Malén Massot-Cladera. 2020. "Development and Characterization of an Allergic Asthma Rat Model for Interventional Studies" International Journal of Molecular Sciences 21, no. 11: 3841. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113841