Early Cellular Responses Induced by Sedimentary Calcite-Processed Particles in Bright Yellow 2 Tobacco Cultured Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
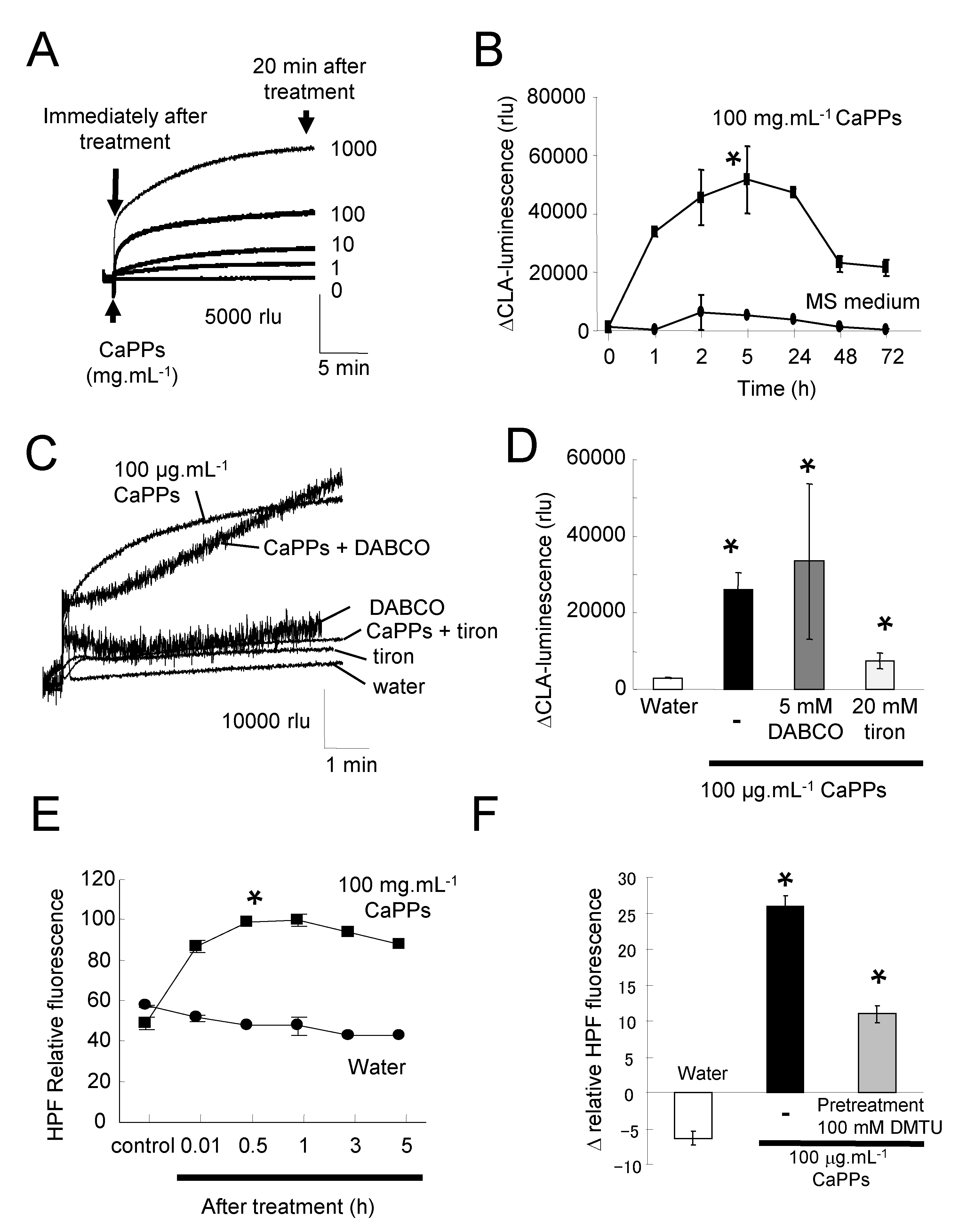
2.1. Non-Biological ROS Production by CaPPs
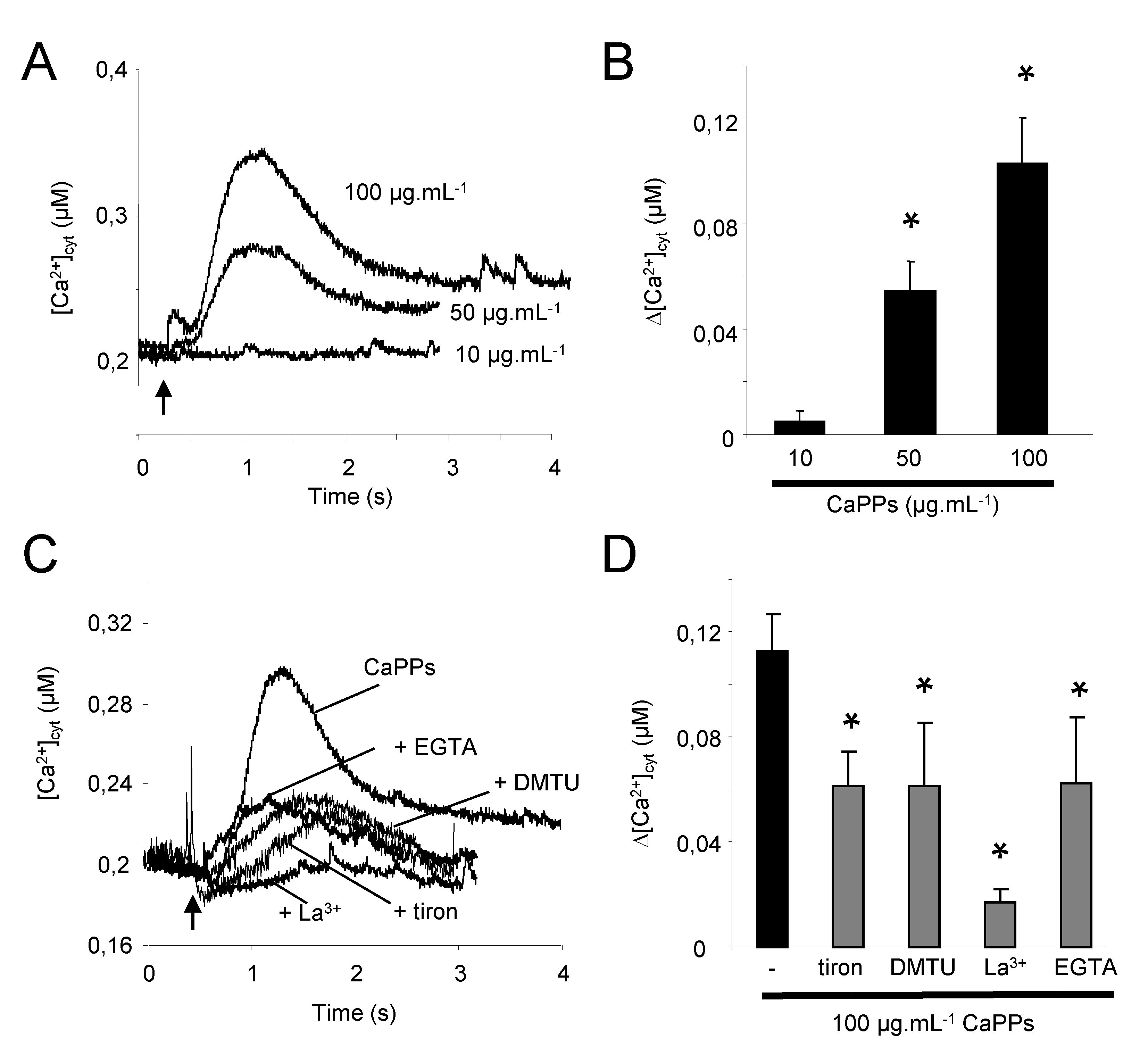
2.2. CaPP Particles Induced Cytosolic Calcium Variation in Tobacco BY-2 Cells
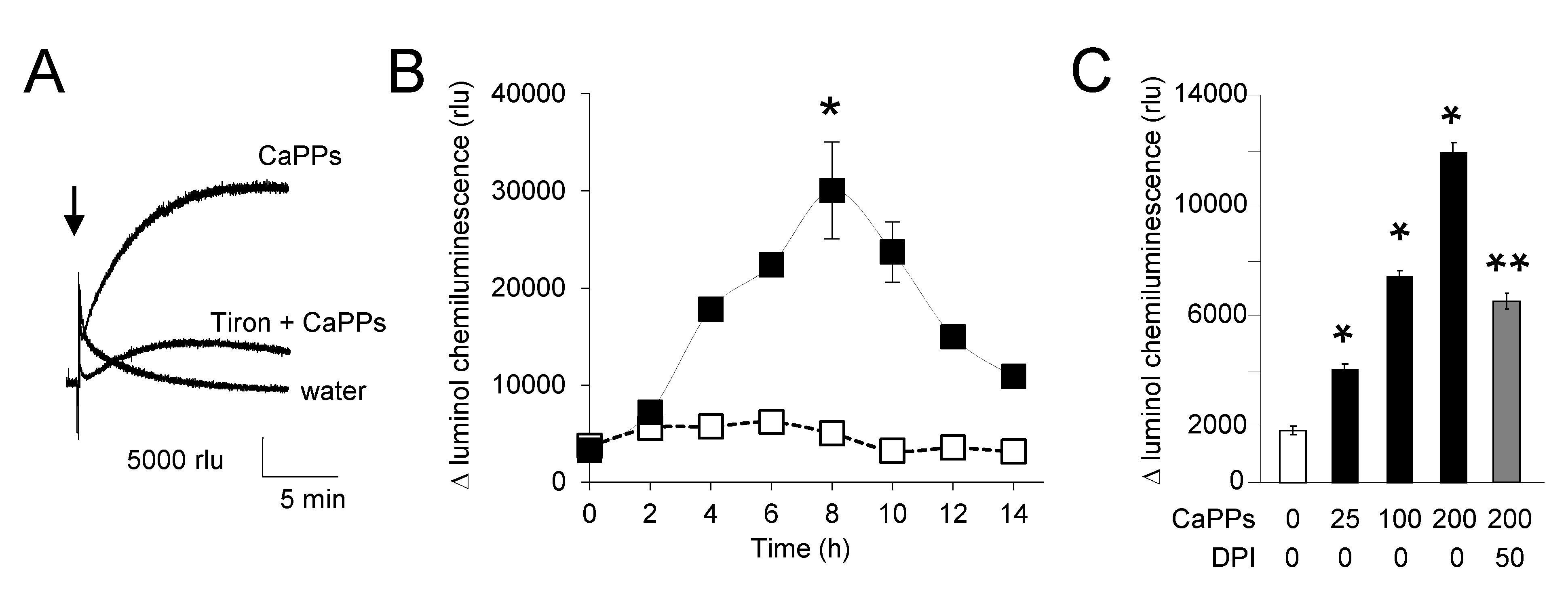
2.3. CaPPs Induced a NADPH Oxidase-Dependent ROS Production
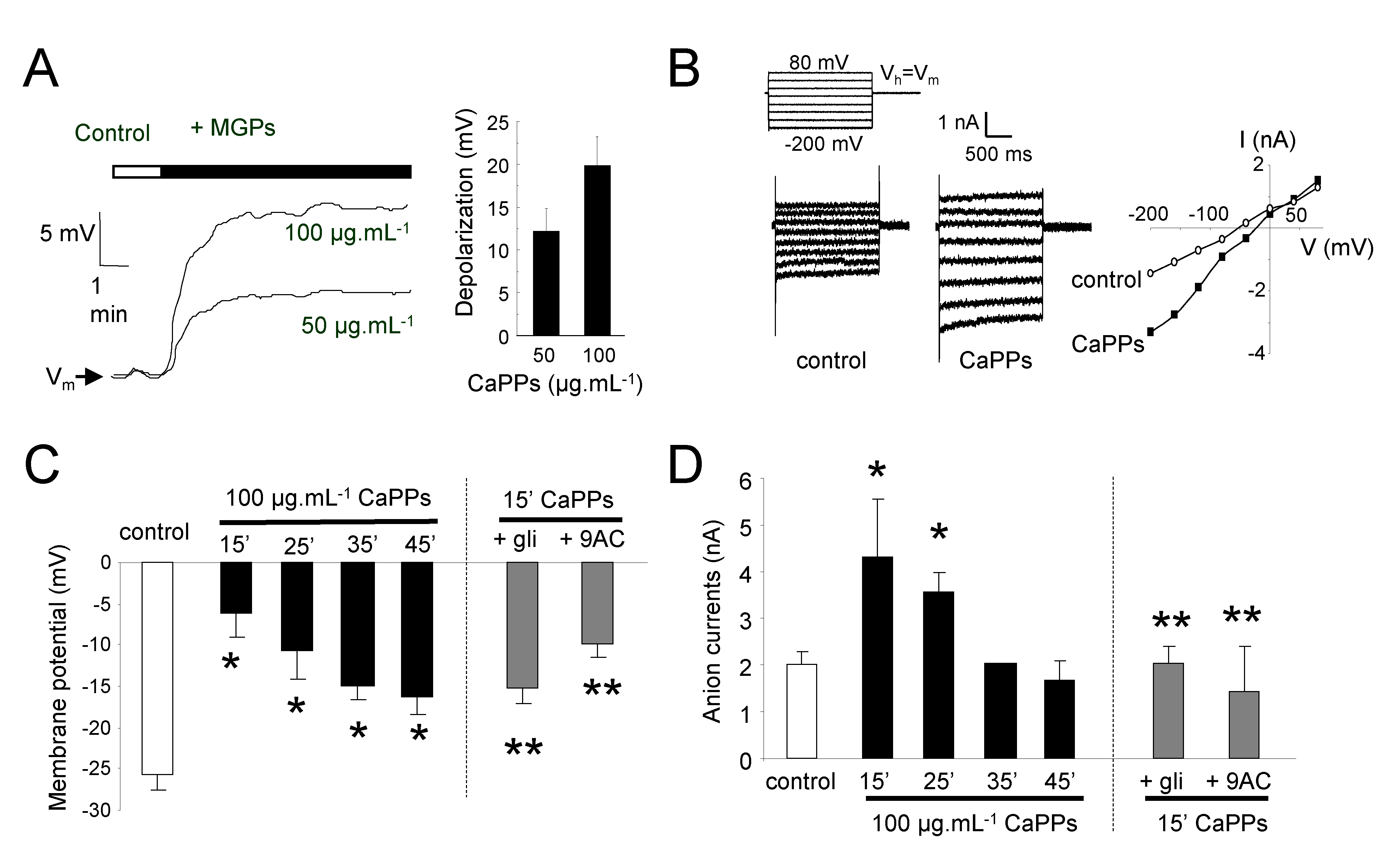
2.4. CaPPs Induce a Depolarization of Plasma Membrane Due to Anion Channel Activation
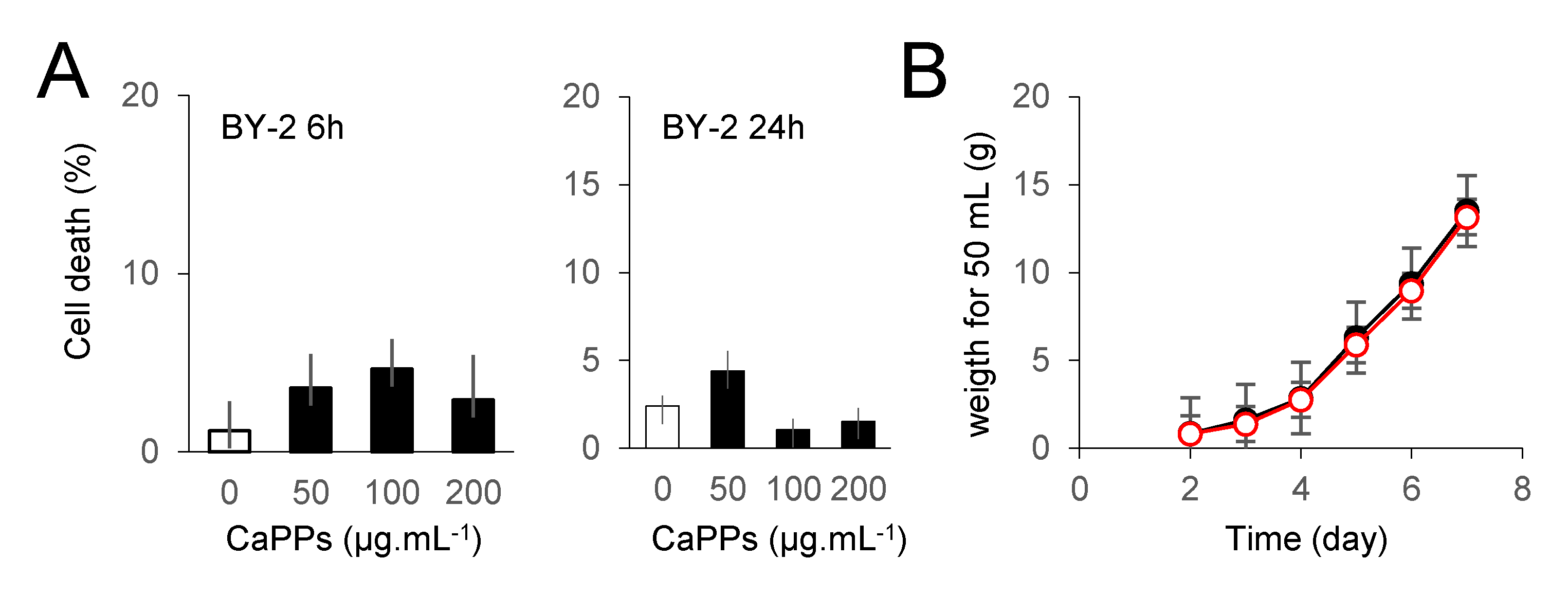
2.5. CaPPs Toxicity?
3. Discussion
4. Materials and Methods
4.1. CaPP Particles
4.2. Plant Cell Culture Conditions
4.3. Monitoring of ROS Production
4.4. Aequorin Luminescence Measurements
4.5. Electrophysiology
4.6. Cell Viability Assays
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Straaten, P. Farming with Rocks and Minerals: Challenges and Opportunities. An. Acad. Bras. Ciênc. 2006, 78. [Google Scholar] [CrossRef] [Green Version]
- Device for Micronizing Materials. EU Patent WO/2000/064586, 2000.
- Attia, F.; Martinez, L.; Lamaze, T. Foliar Application of Processed Calcite Particles Improved Leaf Photosynthsis of Potted Vitis vinifera Grown Under Water Deficit. OENO ONE 2014, 48, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Khot, L.R.; Sankaran, S.; Maja, J.M.; Eshani, R.; Schuster, E.W. Applications of Nanomaterials in Agricultural Production and Crop Protection. Crop Prot. 2012, 35, 64–70. [Google Scholar] [CrossRef]
- Parisi, C.; Vigani, M.; Rodríguez-Cerezo, E. Agricultural Nanotechnologies: What Are the Current Possibilities? Nano Today 2015, 10, 124–127. [Google Scholar] [CrossRef]
- Wang, P.; Lombi, E.; Zhao, F.J.; Kopittke, P.M. Nanotechnology: A New Opportunity in Plant Sciences. Trends Plant Sci. 2016, 21, 699–712. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, S.; Pandey, R.; Singh, V.P.; Sharma, N.C.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. An Overview on Manufactured Nanoparticles in Plants: Uptake, Translocation, Accumulation and Phytotoxicity. Plant Physiol. Biochem. 2017, 110, 2–12. [Google Scholar] [CrossRef]
- Iavicoli, I.; Leso, V.; Beezhold, D.H.; Shvedova, A.A. Nanotechnology in Agriculture: Opportunities, Toxicological Implications, and Occupational risks. Toxicol. Appli. Pharmacol. 2017, 329, 96–111. [Google Scholar] [CrossRef]
- Arora, S.; Sharma, P.; Kumar, S.; Nayan, R.; Khanna, P.K.; Zaidi, M.G.H. Gold-Nanoparticles Induced Enhancement in Growth and Seed Yield of Brassica juncea. Plant Growth Regul. 2012, 66, 303–310. [Google Scholar] [CrossRef]
- Rai, M.; Ingle, A. Role of Nanotechnology in Agriculture with Special Reference to Management of Insect Pests. Appl. Microbiol. Biotechnol. 2012, 94, 287–293. [Google Scholar] [CrossRef]
- Wu, H.; Tito, N.; Juan, P.; Giraldo, J.P. Anionic Cerium Oxide Nanoparticles Protect Plant Photosynthesis from Abiotic Stress by Scavenging Reactive Oxygen Species. ACS Nano 2017, 11, 11283–11297. [Google Scholar] [CrossRef]
- Oberdöster, G.; Oberdöster, E.; Oberdöster, J. Nanotoxicology: An Emerging Discipline Evolving from Studies of Ultrafine Particles. Environ. Health Respect 2005, 113, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Voinov, M.A.; Sosa, J.A.; Morrison, P.E.; Smirnova, T.I.; Smirnov, A.I. Surface-Mediated Production of Hydroxyl Radicals as a Mechanism of Iron Oxide Nanoparticle Biotoxicity. J. Am. Chem. Soc. 2011, 133, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.P.; Xia, Q.; Hwang, H.M.; Ray, C.P.; Yu, H. Mechanisms of Nanotoxicity: Generation of Reactive Oxygen Species. J. Food Drug Anal. 2014, 22, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, D.; Kadono, T.; Meimoun, P.; Kawano, T.; Bouteau, F. TiO2 Nanoparticles Induce ROS Generation and Cytosolic Ca2+ Increases on BY-2 Tobacco Cells: A Chemiluminescence Study. Luminescence 2010, 25, 140–142. [Google Scholar]
- Lecourieux, D.; Ranjeva, R.; Pugin, A. Calcium in Plant Defence-Signalling Pathways. New Phytol. 2006, 171, 249–269. [Google Scholar] [CrossRef] [PubMed]
- Batistič, O.; Kudla, J. Analysis of Calcium Signaling Pathways in Plants. Biochim. Biophys. Acta 2012, 1820, 1283–1293. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox Regulation in Photosynthetic Organisms: Signaling, Acclimation, and Practical Implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory Burst Oxidases: The Engines of ROS Signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, Y.; Wan, Y.; Zheng, J.; Zhang, X.; Wang, C.; Fang, X.; Lin, J. Study of the Inhibitory Effect of Water-Soluble Fullerenes on Plant Growth at the Cellular Level. ACS Nano 2011, 4, 5743–5748. [Google Scholar] [CrossRef]
- Kadono, T.; Tran, D.; Errakhi, R.; Hiramatsu, T.; Meimoun, P.; Briand, J.; Iwaya-Inoue, M.; Kawano, T.; Bouteau, F. Increased Anion Channel Activity Is an Unavoidable Event in Ozone-Induced Programmed Cell Death. PLoS ONE 2010, 5, e13373. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Sugioka, K.; Ushijima, Y.; Goto, T. Chemiluminescence Probe with Cypridina Luciferin Analog, 2-methyl-6-phenyl-3,7-dihydroimidazo[1,2-a]pyrazin-3-one, for Estimating the Ability of Human Granulocytes to Generate O2−. Anal. Biochem. 1986, 159, 363–369. [Google Scholar] [CrossRef]
- Setsukinai, K.; Urano, Y.; Kakinuma, K.; Majima, H.J.; Nagano, T. Development of Novel Fluorescence Probes That Can Reliably Detect Reactive Oxygen Species and Distinguish Specific Species. J. Biol. Chem. 2003, 278, 170–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.; et al. Reactive Oxygen Species Produced by NADPH Oxidase Regulate Plant Cell Growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef]
- Knight, M.R.; Campbell, A.K.; Smith, S.M.; Trewavas, A.J. Transgenic Plant Aequorin Reports the Effects of Touch and Cold-Shock and Elicitors on Cytoplasmic Calcium. Nature 1991, 352, 524–526. [Google Scholar] [CrossRef]
- Monetti, E.; Kadono, T.; Tran, D.; Azzarello, E.; Arbelet-Bonnin, D.; Biligui, B.; Briand, J.; Kawano, T.; Mancuso, S.; Bouteau, F. Deciphering in Early Events Involved in Hyperosmotic Stress-Induced Programmed Cell Death in Tobacco BY-2 Cells. J. Exp. Bot. 2014, 65, 1361–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, T.M.; Auh, C.K. The Superoxide Synthases of Plasma Membrane Preparations from Cultured Rose Cells. Plant Physiol. 1996, 110, 621–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gestelen, P.; Asard, H.; Caubergs, R.J. Solubilization and Separation of a Plant Plasma Membrane NADPH–O2- Synthase from Other NADPH Oxidoreductases. Plant Physiol. 1997, 115, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, A.; Lamotte, O.; Reboutier, D.; Bouteau, F.; Pugin, A.; Wendehenne, D. Cryptogein-Induced Anion Effluxes: Electrophysiological Properties and Analysis of the Mechanisms Through Which They Contribute to the Elicitor-Triggered Cell Death. Plant Signal Behav. 2007, 2, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Brault, M.; Amiar, Z.; Pennarun, A.M.; Monestiez, M.; Zhang, Z.; Cornel, D.; Dellis, O.; Knight, H.; Bouteau, F.; Rona, J.P. Plasma Membrane Depolarization Induced by Abscisic Acid in Arabidopsis Suspension Cells Involves Reduction of Proton Pumping in Addition to Anion Channel Activation, Which Are Both Ca2+ Dependent. Plant Physiol. 2004, 135, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reboutier, D.; Bianchi, M.; Brault, M.; Roux, C.; Dauphin, A.; Rona, J.P.; Legue, V.; Lapeyrie, F.; Bouteau, F. The Indolic Compound Hypaphorine Produced by Ectomycorrhizal Fungus Interferes with Auxin Action and Evokes Early Responses in Non-Host Arabidopsis Thaliana. Mol. Plant Microbe Interac. 2002, 15, 932–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, J.I.; Keller, B.U. Two Types of Anion Channel Currents in Guard Cells with Distinct Voltage Regulation. Proc. Natl. Acad. Sci. USA 1992, 89, 5025–5029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedrich, R.; Busch, H.; Raschke, K. Ca2+ and Nucleotide Dependent Regulation of Voltage Dependent Anion Channels in the Plasma Membrane of Guard Cells. EMBO J. 1990, 9, 3889–3892. [Google Scholar] [CrossRef]
- Shen, C.X.; Zhang, Q.F.; Li, J.; Bi, F.C.; Yao, N. Induction of Programmed Cell Death in Arabidopsis and Rice by Single-Wall Carbon Nanotubes. Am. J. Bot. 2010, 97, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Deepa, M.; Sudhakar, P.; Venkata, K.; Kota, N.; Reddy, B.; Krishna, T.G.; Krishna, N.V.; Prasad, V. First Evidence on Phloem Transport of Nanoscale Calcium Oxide in Groundnut Using Solution Culture Technique. Appl Nanosci. 2015, 5, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Sofo, A.; Scopa, A.; Manfra, M.; De Nisco, M.; Tenore, G.C.; Nuzzo, V. Different Water and Light Regimes Affect Ionome Composition in Grapevine (Vitis Vinifera L.). Vitis 2013, 52, 13–20. [Google Scholar]
- Balážová, Ľ.; Babula, P.; Baláž, M.; Bačkorová, M.; Bujňáková, Z.; Briančin, J.; Kurmanbayeva, A.; Sagi, M. Zinc Oxide Nanoparticles Phytotoxicity on Halophyte from Genus Salicornia. Plant Physiol Biochem. 2018, 130, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Vainonen, J.P.; Kangasjärvi, J. Plant Signalling in Acute Ozone Exposure. Plant Cell Environ. 2015, 38, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Kadono, T.; Yamaguchi, Y.; Furuichi, T.; Hirono, M.; Garrec, J.P. Ozone-Induced Cell Death Mediated with Oxidative and Calcium Signaling Pathways in Tobacco Bel-w3 and Bel-B Cell Suspension Cultures. Plant Signal. Behav. 2006, 1, 312–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, D.; El-Maarouf-Bouteau, H.; Rossi, M.; Biligui, B.; Briand, J.; Kawano, T.; Mancuso, S.; Bouteau, F. Post-Transcriptional Regulation of GORK Channels by Superoxide Anion Contributes Towards Increases in Outward Rectifying K+ Currents. New Phytol. 2013, 198, 1039–1048. [Google Scholar] [CrossRef]
- Tran, D.; Molas, M.L.; Kadono, T.; Errakhi, R.; Briand, J.; Biligui, B.; Kawano, T.; Bouteau, F. A Role for Oxalic Acid Generation in Ozone-Induced Programmed Cell Death. Plant Cell Environ. 2013, 36, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.; Rossi, M.; Biligui, B.; Kawano, T.; Mancuso, S.; Bouteau, F. Ozone-Induced Caspase-Like Activities Are Dependent on Early Ion Channel Regulations and ROS Generation in Arabidopsis Thaliana Cells. Plant Signal. Behav. 2013, 8, e25170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, J.; Gong, X.; Nahirney, D.; Duszyk, M.; Radomski, M. Polystyrene Nanoparticles Activate Ion Transport in Human Airway Epithelial Cells. Int. J. Nanomed. 2011, 6, 1343–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oukarroum, A.; Bras, S.; Perreault, F.; Popovic, R. Inhibitory Effects of Silver Nanoparticles in Two Green Algae, Chlorella Vulgaris and Dunaliella Tertiolecta. Ecotoxicol. Environ. Safety 2012, 78, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Powell, B.A.; Mortimer, M.; Ke, P.C. Adaptive Interactions between Zinc Oxide Nanoparticles and Chlorella sp. Environ. Sci. Technol. 2012, 46, 12178–12185. [Google Scholar] [CrossRef] [PubMed]
- Atha, D.H.; Wang, H.; Petersen, E.J.; Cleveland, D.; Holbrook, R.D.; Jaruga, P.; Dizdaroglu, M.; Xing, B.; Nelson, B.C. Copper Oxide Nanoparticle Mediated DNA Damage in Terrestrial Plant Models. Environ. Sci. Technol. 2012, 46, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Slomberg, D.L.; Schoenfisch, M.H. Silica Nanoparticle Phytotoxicity to Arabidopsis Thaliana. Environ. Sci. Technol. 2012, 46, 10247–10254. [Google Scholar] [PubMed]
- Khodakovskaya, M.V.; de Silva, K.; Biris, A.S.; Dervishi, E.; Villagarcia, H. Carbon Nanotubes Induce Growth Enhancement of Tobacco Cells. ACS Nano 2012, 6, 2128–2135. [Google Scholar] [CrossRef]
- Schroeder, J.I.; Kwak, J.M.; Allen, G.J. Guard Cell Abscisic Acid Signalling and Engineering of Drought Hardiness in Plants. Nature 2001, 410, 327–330. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bouizgarne, B.; El-Maarouf-Bouteau, H.; Frankart, C.; Reboutier, D.; Madiona, K.; Pennarun, A.M.; Monestiez, M.; Trouvier, J.; Amiar, Z.; Briand, J.; et al. Early Physiological Responses of Arabidopsis Thaliana Cells to Fusaric Acid: Toxic and Signaling Effects. New Phytol. 2006, 169, 209–218. [Google Scholar] [CrossRef]
- Finkel, A.S.; Redman, S. Theory and Operation of a Single Microelectrode Voltage Clamp. J. Neurosci. Methods 1984, 11, 101–127. [Google Scholar] [CrossRef]
- Bouteau, F.; Tran, D. Plant response to stress: Microelectrode voltage clamp studies. In Plant Electrophysiology; Volkov, A.G., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 3, pp. 69–90. [Google Scholar]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, D.; Zhao, T.; Arbelet-Bonnin, D.; Kadono, T.; Meimoun, P.; Cangémi, S.; Kawano, T.; Errakhi, R.; Bouteau, F. Early Cellular Responses Induced by Sedimentary Calcite-Processed Particles in Bright Yellow 2 Tobacco Cultured Cells. Int. J. Mol. Sci. 2020, 21, 4279. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124279
Tran D, Zhao T, Arbelet-Bonnin D, Kadono T, Meimoun P, Cangémi S, Kawano T, Errakhi R, Bouteau F. Early Cellular Responses Induced by Sedimentary Calcite-Processed Particles in Bright Yellow 2 Tobacco Cultured Cells. International Journal of Molecular Sciences. 2020; 21(12):4279. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124279
Chicago/Turabian StyleTran, Daniel, Tingting Zhao, Delphine Arbelet-Bonnin, Takashi Kadono, Patrice Meimoun, Sylvie Cangémi, Tomonori Kawano, Rafik Errakhi, and François Bouteau. 2020. "Early Cellular Responses Induced by Sedimentary Calcite-Processed Particles in Bright Yellow 2 Tobacco Cultured Cells" International Journal of Molecular Sciences 21, no. 12: 4279. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124279