Short-Chain Fatty Acids (Except Hexanoic Acid) Lower NF-kB Transactivation, Which Rescues Inflammation-Induced Decreased Apolipoprotein A-I Transcription in HepG2 Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
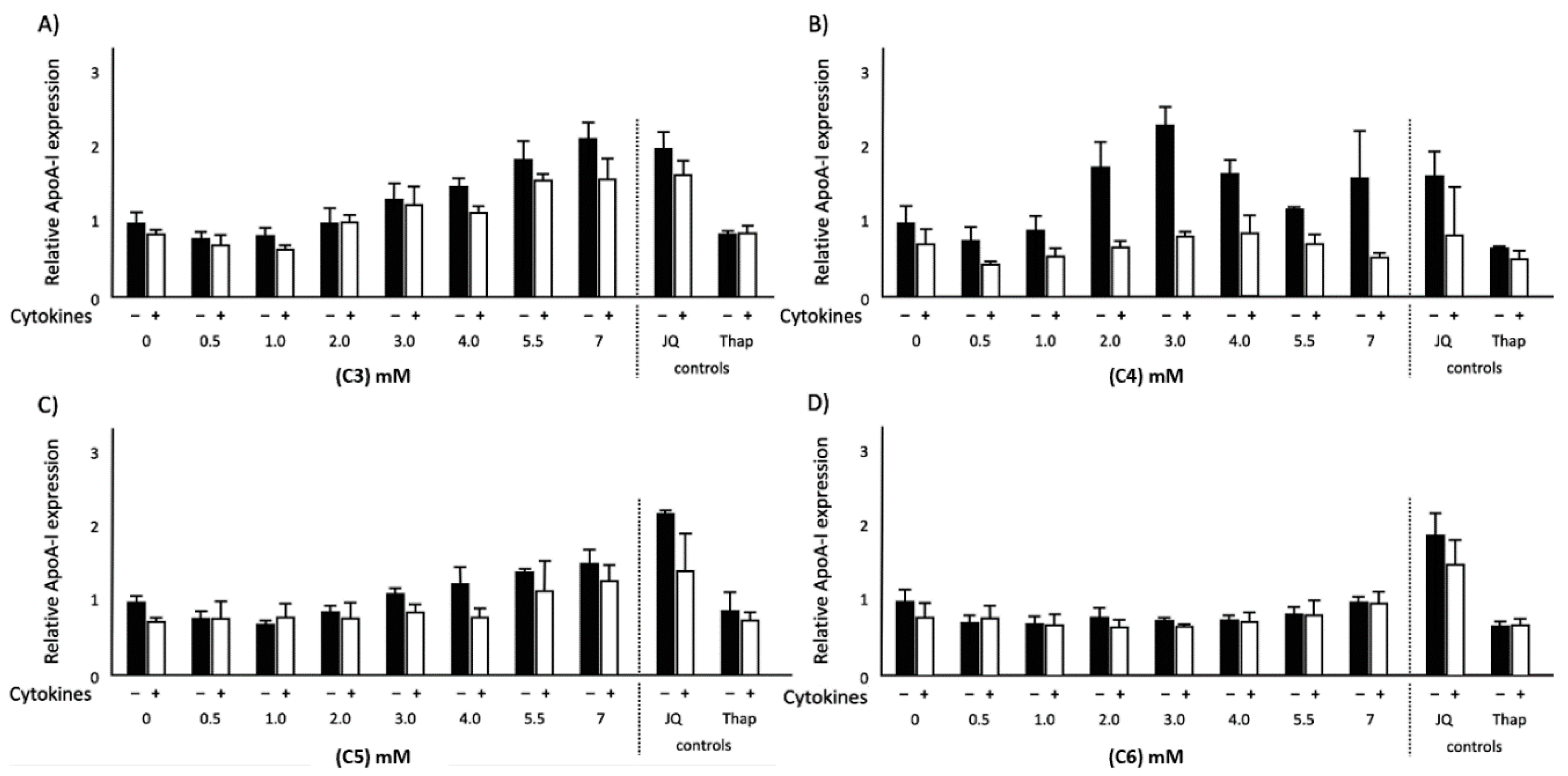
2.1. Effects of SCFAs on ApoA-I mRNA Expression in Normal and Inflammatory Conditions
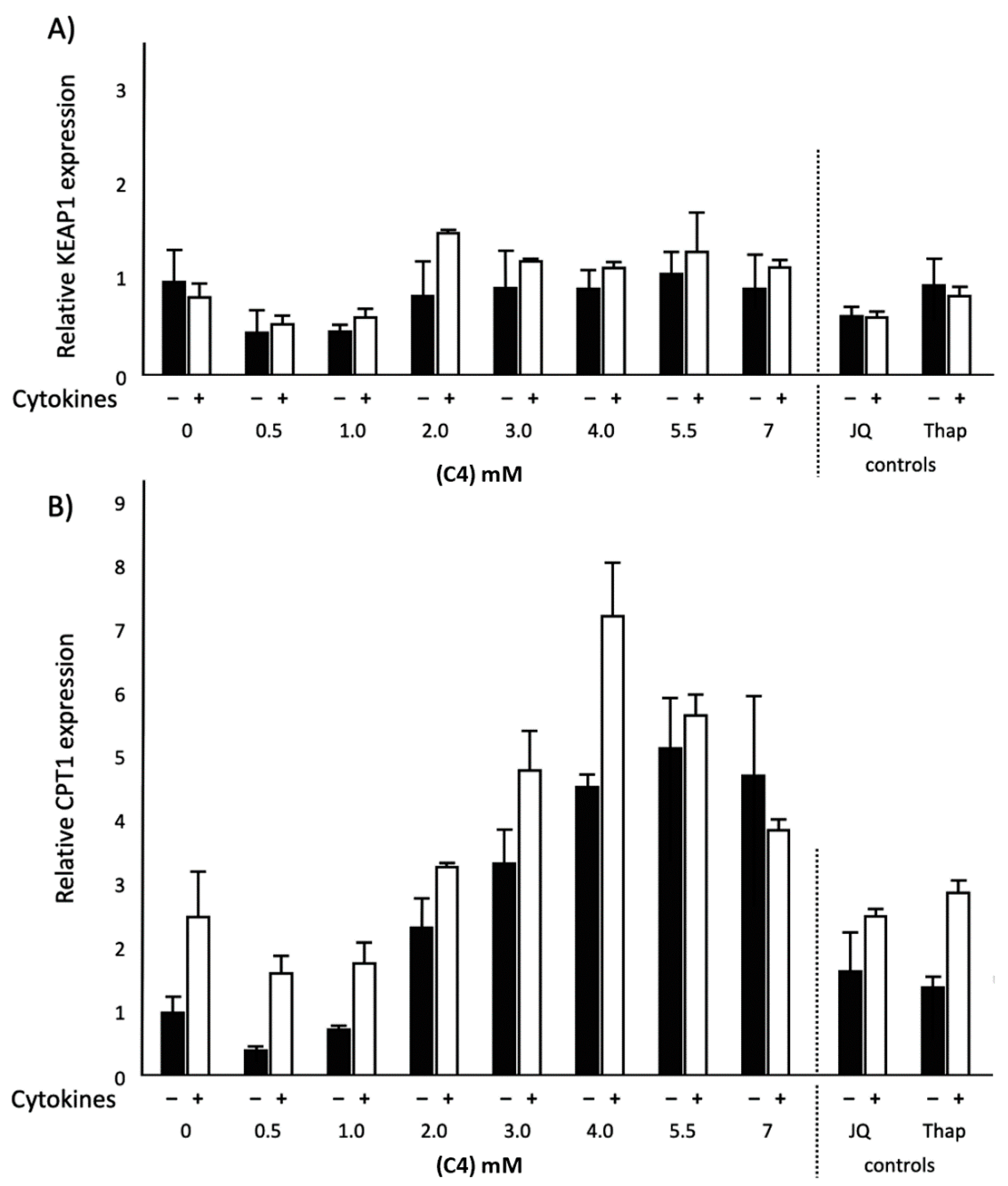
2.2. Effects of C4 on KEAP1 and CPT1 mRNA Expression and NF-κB Transactivation in Normal and Inflammatory Conditions
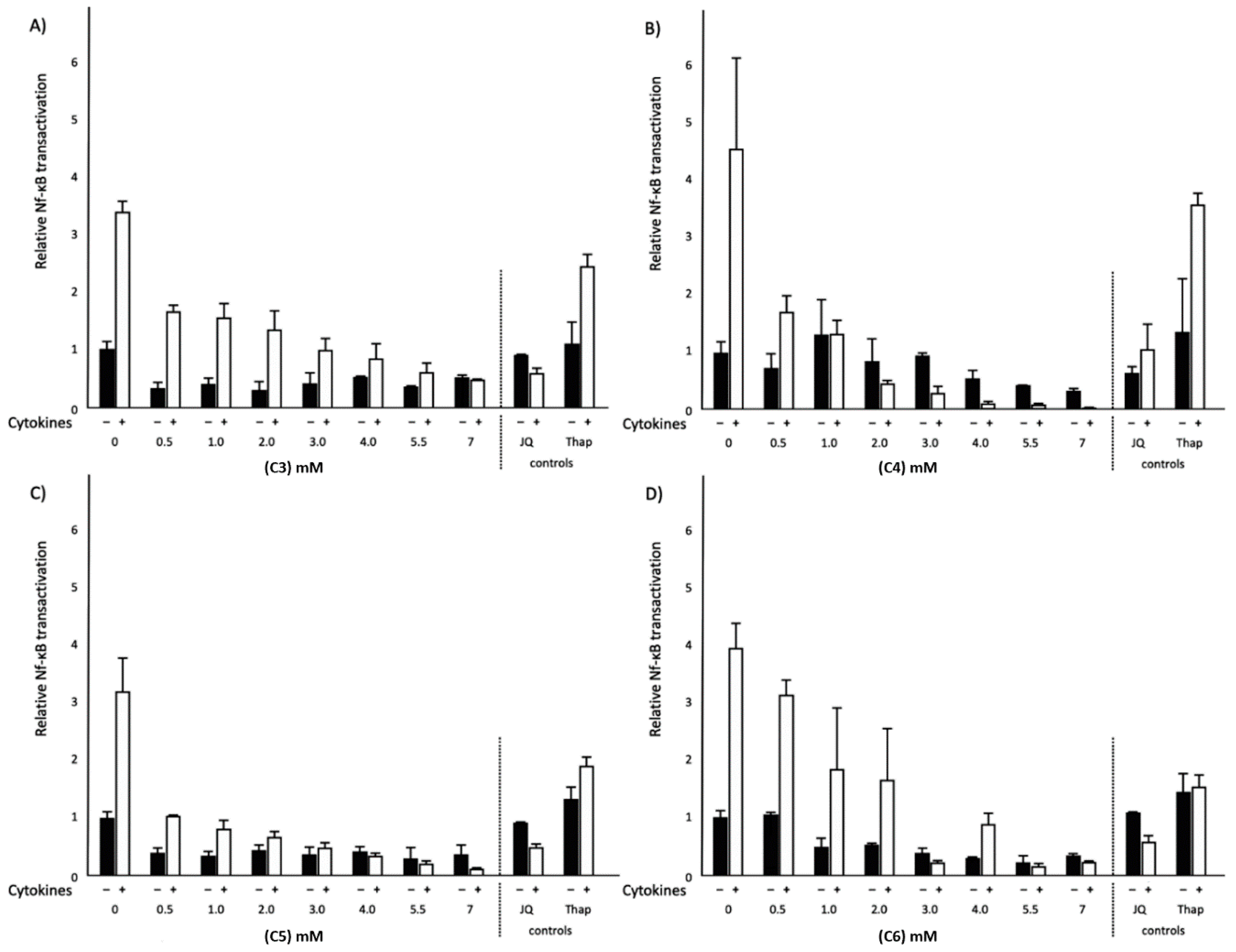
2.3. Effects of Different SCFAs on NF-κB Transactivation and IL-8 Secretion in Normal and Inflammatory Conditions
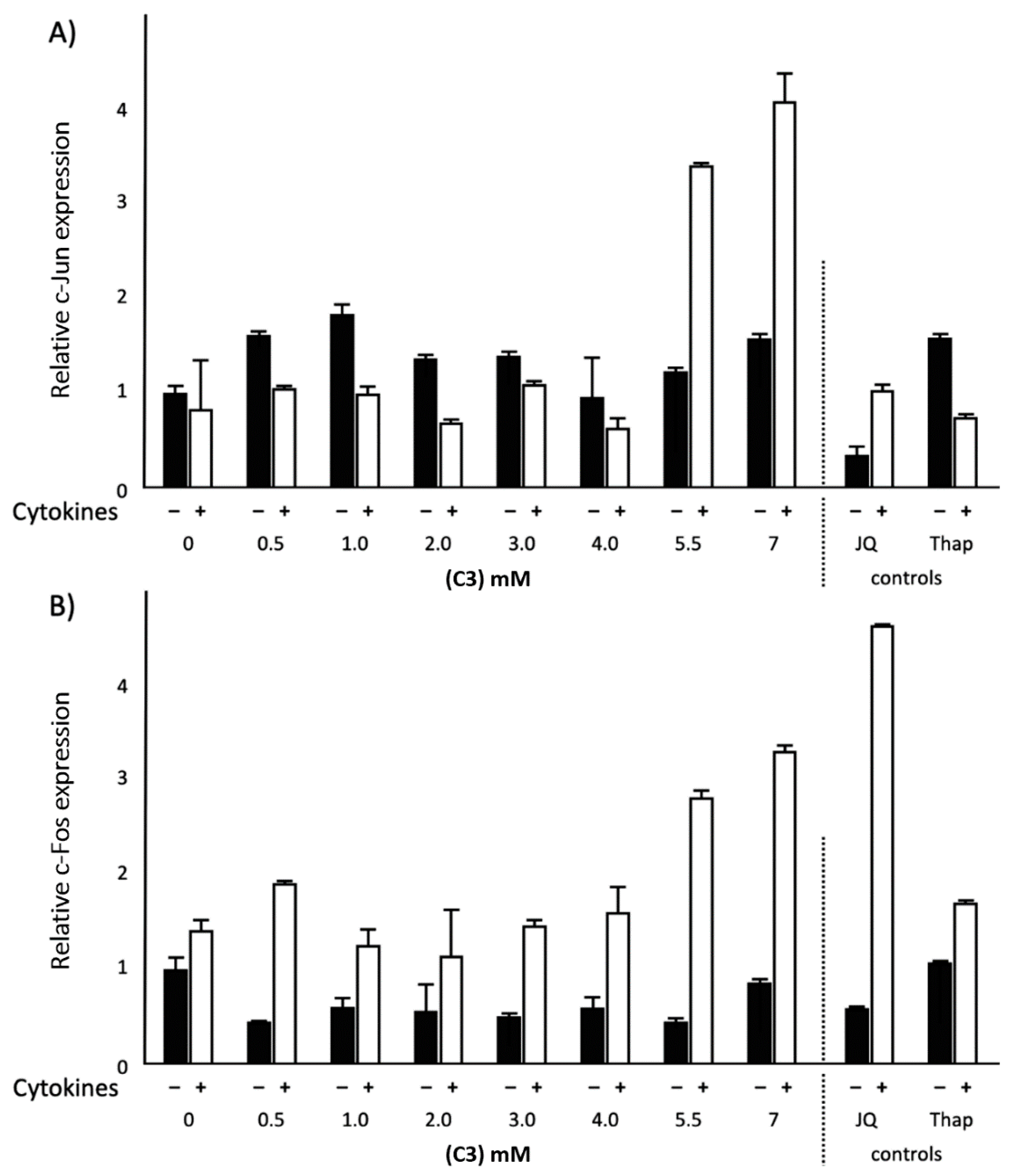
2.4. Effects of Different SCFAs on c-Jun and c-Fos mRNA Expression in Normal and Inflammatory Conditions
3. Discussion
4. Material and Methods
4.1. Materials
4.2. Cell Culture and SCFA Treatment
4.3. Quantification of Gene mRNA Transcription
4.4. Luciferase Assay
4.5. Quantification of IL-8 Secretion Levels in the Culture Medium
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Esteve, E.; Ricart, W.; Fernández-Real, J.M. Dyslipidemia and inflammation: An evolutionary conserved mechanism. Clin. Nutr. 2005, 24, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Kosmas, C.E.; Silverio, D.; Sourlas, A.; Montan, P.D.; Guzman, E. Dysfunctional high-density lipoprotein and atherogenesis. Vessel Plus. 2019, 3, 2. [Google Scholar] [CrossRef]
- Chyu, K.-Y.; Peter, A.; Shah, P.K. Progress in HDL-based therapies for atherosclerosis. Curr. Atheroscler. Rep. 2011, 13, 405. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.; Reinacher, D.; Li, J.; Wong, N.; Mooradian, A. Regulation of apoA1 gene expression with acidosis: Requirement for a transcriptional repressor. J. Mol. Endocrinol. 2001, 27, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Wang, L.; Hong, B. High-density lipoprotein-based drug discovery for treatment of atherosclerosis. Expert. Opin. Drug. Dis. 2015, 10, 841–855. [Google Scholar] [CrossRef]
- Gerster, R.; Eloranta, J.J.; Hausmann, M.; Ruiz, P.A.; Cosin-Roger, J.; Terhalle, A. Anti-inflammatory function of high-density lipoproteins via autophagy of IκB kinase. Cell. Mol. Gastroenterol. 2015, 1, 171–187. [Google Scholar] [CrossRef] [Green Version]
- Oslakovic, C.; Krisinger, M.; Jauhiainen, M.; Ehnholm, C.; Dahlbäck, B. 614 anticoagulant properties of apolipoprotein ai and phospholipid transfer protein. Atheroscler. Suppl. 2009, 10, 138. [Google Scholar] [CrossRef]
- Georgila, K.; Vyrla, D.; Drakos, E. Apolipoprotein AI (ApoA-I), Immunity, Inflammation and Cancer. Cancers 2019, 11, 1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohira, H.; Tsutsui, W.; Fujioka, Y. Are short chain fatty acids in gut microbiota defensive players for inflammation and atherosclerosis? J. Atheroscler. Thromb. 2017, 24, 660–672. [Google Scholar] [CrossRef] [Green Version]
- Nakatani, M.; Inoue, R.; Tomonaga, S.; Fukuta, K.; Tsukahara, T. Production, Absorption, and Blood Flow Dynamics of Short-Chain Fatty Acids Produced by Fermentation in Piglet Hindgut during the Suckling–Weaning Period. Nutrients 2018, 10, 1220. [Google Scholar] [CrossRef] [Green Version]
- Tayyeb, J.Z.; Popeijus, H.E.; Mensink, R.P.; Konings, M.C.; Mulders, K.H.; Plat, J. The effects of short-chain fatty acids on the transcription and secretion of apolipoprotein A-I in human hepatocytes in vitro. J. Cell. Biochem. 2019, 120, 17219–17227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S. Short-chain fatty acid propionate protects from hypertensive cardiovascular damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; De Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Krieken, S.E.; Popeijus, H.E.; Mensink, R.P.; Plat, J. Link between ER-stress, PPAR-alpha activation, and BET inhibition in relation to apolipoprotein A-I transcription in HepG2 cells. J. Cell. Biochem. 2017, 118, 2161–2167. [Google Scholar] [CrossRef]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [Green Version]
- Haas, M.J.; Horani, M.; Mreyoud, A.; Plummer, B.; Wong, N.C.; Mooradian, A.D. Suppression of apolipoprotein AI gene expression in HepG2 cells by TNF α and IL-1β. Biochim. Biophys. Acta 2003, 1623, 120–128. [Google Scholar] [CrossRef]
- Carpintero, R.; Pineiro, M.; Andres, M.; Iturralde, M.; Alava, M.; Heegaard, P.M. The concentration of apolipoprotein AI decreases during experimentally induced acute-phase processes in pigs. Infect. Immun. 2005, 73, 3184–3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chait, A.; Han, C.Y.; Oram, J.F.; Heinecke, J.W. Lipoprotein-associated inflammatory proteins: Markers or mediators of cardiovascular disease. J. Lipid Res. 2005, 46, 389–403. [Google Scholar] [CrossRef] [Green Version]
- Morishima, A.; Ohkubo, N.; Maeda, N.; Miki, T.; Mitsuda, N. NFκB regulates plasma apolipoprotein AI and high density lipoprotein cholesterol through inhibition of peroxisome proliferator-activated receptor α. J. Biol. 2003, 278, 38188–38193. [Google Scholar]
- Trusca, V.G.; Dumitrescu, M.; Fenyo, I.M.; Tudorache, I.F.; Simionescu, M.; Gafencu, A.V. The Mechanism of Bisphenol A Atherogenicity Involves Apolipoprotein AI Downregulation through NF-κB Activation. Int. J. Mol. Sci. 2019, 20, 6281. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.-L.; Qian, J.-M.; Wang, F.-R.; Ma, Z.-Y.; Wang, Q.-W. Butyrate protects liver against ischemia reperfusion injury by inhibiting nuclear factor kappa B activation in Kupffer cells. J. Surg. Res. 2014, 187, 653–659. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.C.; Henricks, P.A.; Garssen, J.; Folkerts, G. Time and concentration dependent effects of short chain fatty acids on lipopolysaccharide-or tumor necrosis factor α-induced endothelial activation. Front. Pharmacol. 2018, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- Usami, M.; Kishimoto, K.; Ohata, A.; Miyoshi, M.; Aoyama, M.; Fueda, Y. Butyrate and trichostatin A attenuate nuclear factor kappaB activation and tumor necrosis factor alpha secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells. Nutr. Res. 2008, 28, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Elliott, C.; Allport, V.; Loudon, J.; Wu, G.; Bennett, P. Nuclear factor-kappa B is essential for up-regulation of interleukin-8 expression in human amnion and cervical epithelial cells. Mol. Hum. Reprod. 2001, 7, 787–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, G.; Duez, H.; Blanquart, C.; Berezowski, V.; Poulain, P.; Fruchart, J.-C. Statin-induced inhibition of the Rho-signaling pathway activates PPARα and induces HDL apoA-I. J. Clin. Investig. 2001, 107, 1423–1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nepelska, M.; Cultrone, A.; Beguet-Crespel, F.; Le Roux, K.; Dore, J.; Arulampalam, V. Butyrate produced by commensal bacteria potentiates phorbol esters induced AP-1 response in human intestinal epithelial cells. PLoS ONE 2012, 7, e52869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Wei, Y.; Wu, L.; Wei, J.; Wang, X.; Bai, Y. C/EBP β mediates endoplasmic reticulum stress regulated inflammatory response and extracellular matrix degradation in LPS-stimulated human periodontal ligament cells. Int. J. Mol. Sci. 2016, 17, 385. [Google Scholar] [CrossRef]
- Van der Krieken, S.E.; Popeijus, H.E.; Konings, M.; Dullens, S.P.; Mensink, R.P.; Plat, J. C/EBP-β Is Differentially Affected by PPARα Agonists Fenofibric Acid and GW7647, But Does Not Change Apolipoprotein A-I Production During ER-Stress and Inflammation. J. Cell. Biochem. 2017, 118, 754–763. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tayyeb, J.Z.; Popeijus, H.E.; Mensink, R.P.; Konings, M.C.J.M.; Mokhtar, F.B.A.; Plat, J. Short-Chain Fatty Acids (Except Hexanoic Acid) Lower NF-kB Transactivation, Which Rescues Inflammation-Induced Decreased Apolipoprotein A-I Transcription in HepG2 Cells. Int. J. Mol. Sci. 2020, 21, 5088. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145088
Tayyeb JZ, Popeijus HE, Mensink RP, Konings MCJM, Mokhtar FBA, Plat J. Short-Chain Fatty Acids (Except Hexanoic Acid) Lower NF-kB Transactivation, Which Rescues Inflammation-Induced Decreased Apolipoprotein A-I Transcription in HepG2 Cells. International Journal of Molecular Sciences. 2020; 21(14):5088. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145088
Chicago/Turabian StyleTayyeb, Jehad Z., Herman E. Popeijus, Ronald P. Mensink, Maurice C. J. M. Konings, Fatma B. A. Mokhtar, and Jogchum Plat. 2020. "Short-Chain Fatty Acids (Except Hexanoic Acid) Lower NF-kB Transactivation, Which Rescues Inflammation-Induced Decreased Apolipoprotein A-I Transcription in HepG2 Cells" International Journal of Molecular Sciences 21, no. 14: 5088. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145088