The Effects of Ozone on Atlantic Salmon Post-Smolt in Brackish Water—Establishing Welfare Indicators and Thresholds


 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Mortality
2.2. Blood
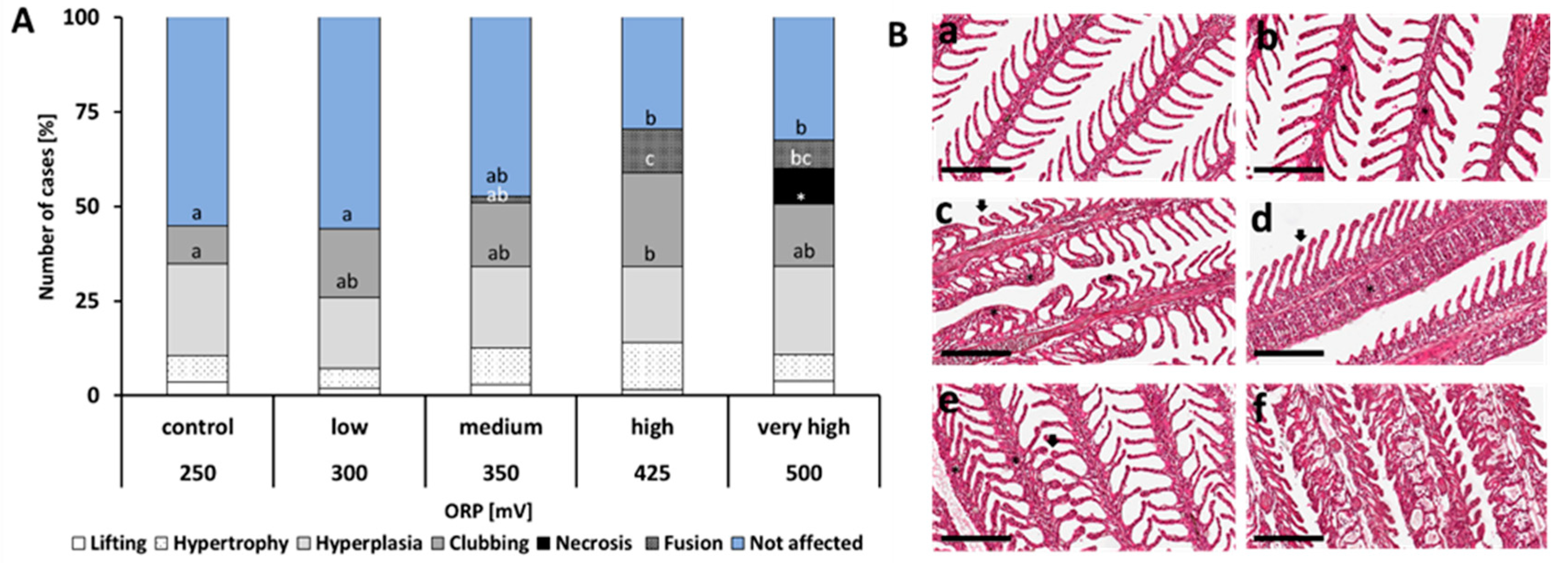
2.3. Histology
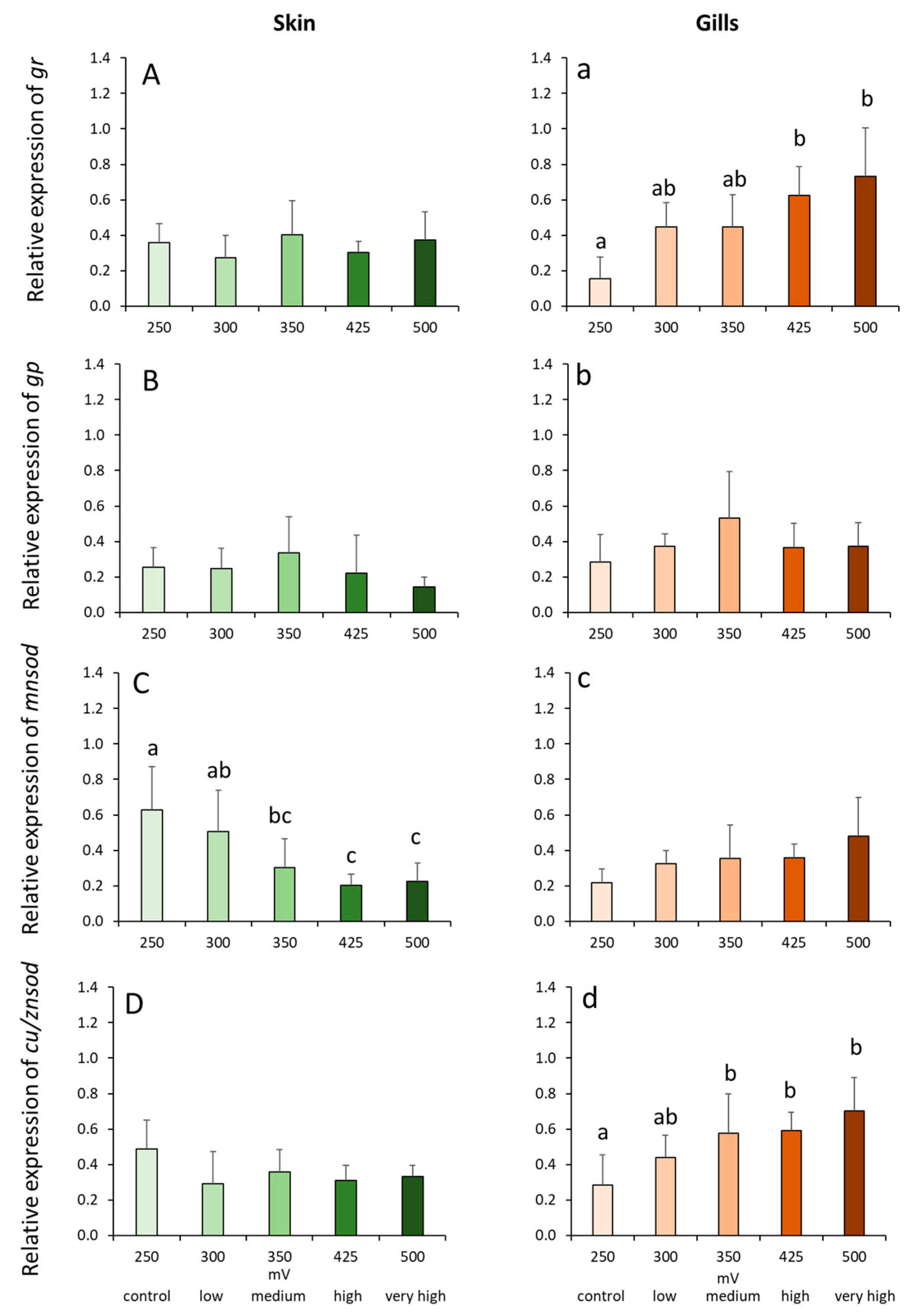
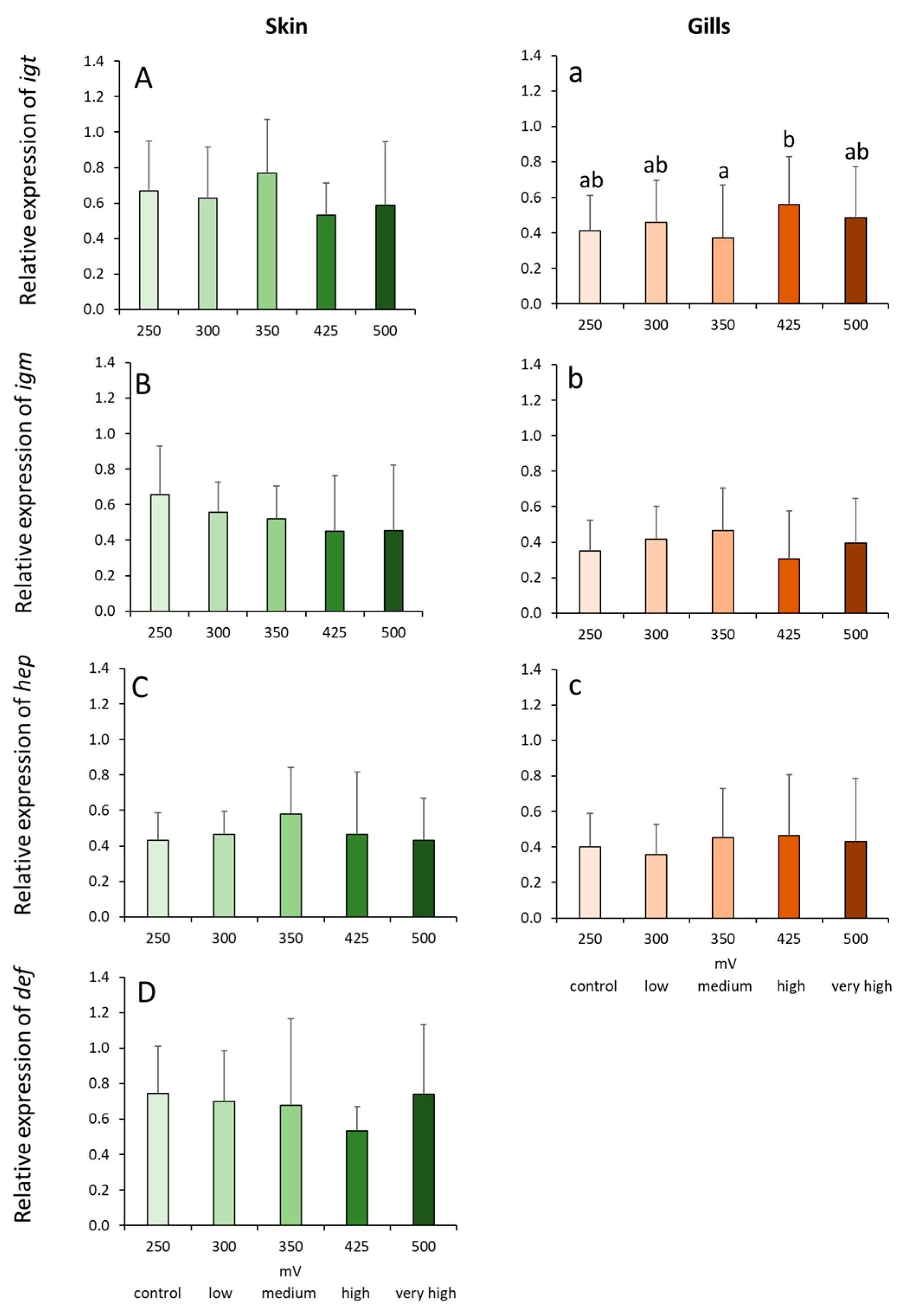
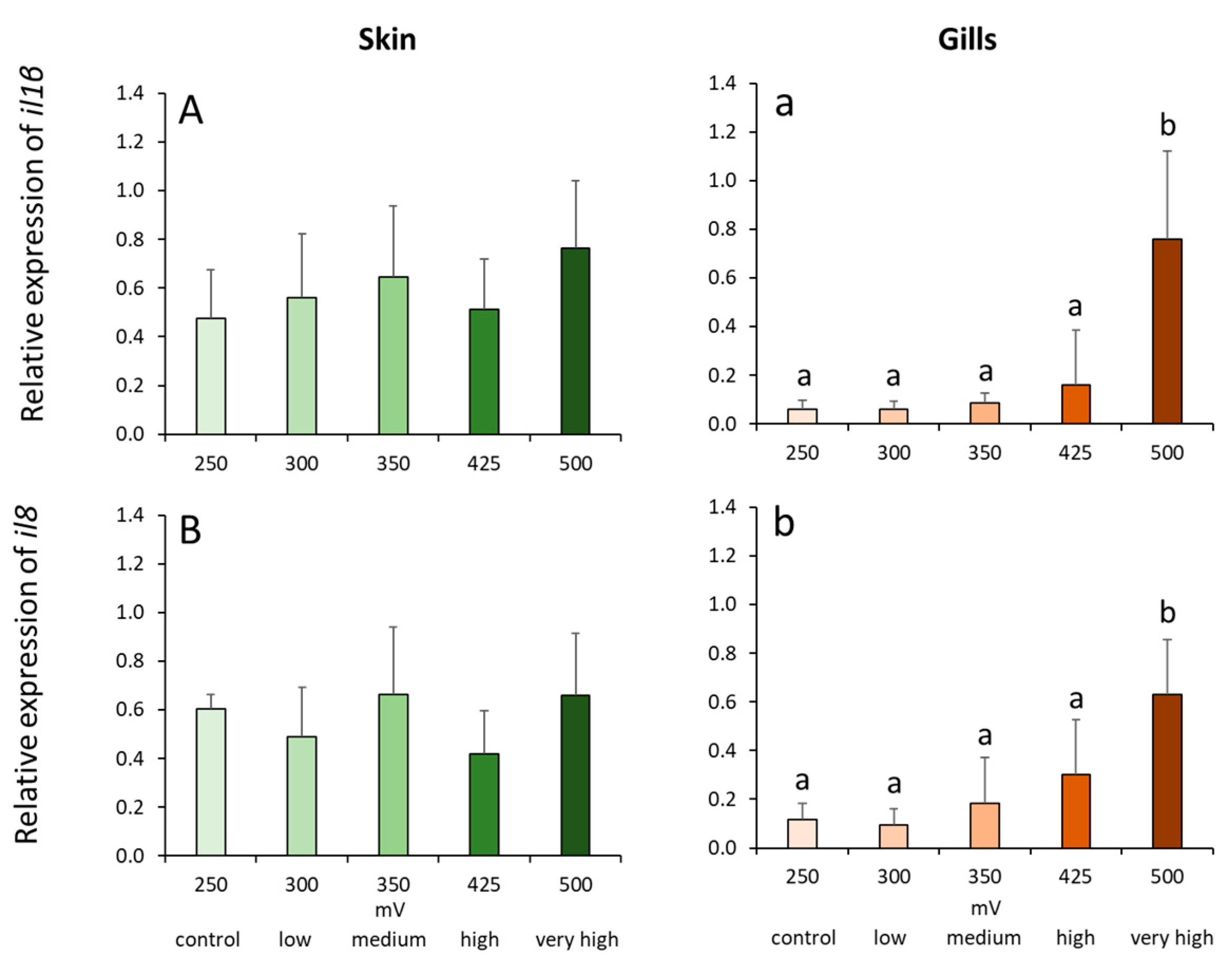
2.4. Gene Expression Profiles in the Skin and Gills
3. Discussion
4. Material and Methods
4.1. Fish Husbandry and Experimental Conditions
4.2. O3 Dosing and Treatments
4.3. Blood, Gill and Skin
4.4. Histology
4.5. RNA Extraction, cDNA Synthesis and RT-qPCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bergheim, A.; Drengstig, B.; Ulgenes, Y.; Fivelstad, S. Production of Atlantic salmon smolts in Europe—Current characteristics and future trends. Aquac. Eng. 2009, 41, 46–52. [Google Scholar] [CrossRef]
- Davidson, J.; Maya, T.; Gooda, C.; Waldrop, T.; Kenney, B.; Terjesen, B.F.; Summerfelt, S. Production of market-size North American strain Atlantic salmon Salmo salar in a land-based recirculation aquaculture system using freshwater. Aquac. Eng. 2016, 74, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ytrestøyl, T.; Takle, H.; Kolarevic, J.; Calabrese, S.; Timmerhaus, G.; Rosseland, B.O.; Teien, H.C.; Nilsen, T.O.; Handeland, S.O.; Stefansson, S.O. Performance and welfare of Atlantic salmon, Salmo salar L. post-smolts in recirculating aquaculture systems: Importance of salinity and water velocity. J. World Aquac. Soc. 2020. [Google Scholar] [CrossRef] [Green Version]
- Powell, A.; Scolding, J.W.S. Direct application of ozone in aquaculture systems. Rev. Aquac. 2018, 1010, 424–438. [Google Scholar] [CrossRef]
- Christensen, P.A.; Yonar, T.; Zakaria, K. The electrochemical generation of ozone: A review. Ozone Sci. Eng. 2013, 3535, 149–167. [Google Scholar] [CrossRef]
- Spiliotopoulou, A.; Martin, R.; Pedersen, L.-F.; Andersen, H.R. Use of fluorescence spectroscopy to control ozone dosage in recirculating aquaculture systems. Water Res. 2017, 111, 357–365. [Google Scholar] [CrossRef] [Green Version]
- Davidson, J.; Good, C.; Welsh, C.; Summerfelt, S. The effects of ozone and water exchange rates on water quality and rainbow trout Oncorhynchus mykiss performance in replicated water recirculating systems. Aquac. Eng. 2011, 4444, 80–96. [Google Scholar] [CrossRef] [Green Version]
- Spiliotopoulou, A.; Rojas-Tirado, P.; Chhetri, R.K.; Kaarsholm, K.M.S.; Martin, R.; Pedersen, P.B.; Pedersen, L.-F.; Andersen, H.-R. Ozonation control and effects of ozone on water quality in recirculating aquaculture systems. Water Res. 2018, 133, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Summerfelt, S.T.; Hochheimer, J.N. Review of ozone processes and applications as an oxidizing agent in aquaculture. Progress. Fish Cult. 1997, 59, 94–105. [Google Scholar] [CrossRef]
- Glavas, S.; Toby, S. Reaction between ozone and hydrogen sulfide. J. Phys. Chem. 1975, 79, 779–782. [Google Scholar] [CrossRef]
- Summerfelt, S.T.; Sharrer, M.J.; Tsukuda, S.M.; Gearheart, M. Process requirements for achieving full-flow disinfection of recirculating water using ozonation and UV irradiation. Aquac. Eng. 2009, 40, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Reiser, S.; Schroeder, J.P.; Wuertz, S.; Kloas, W.; Hanel, R. Histological and physiological alterations in juvenile turbot (Psetta maxima, L.) exposed to sublethal concentrations of ozone-produced oxidants in ozonated seawater. Aquaculture 2010, 307, 157–164. [Google Scholar] [CrossRef]
- Summerfelt, S.T. Ozonation and UV irradiation—An introduction and examples of current applications. Aquac. Eng. 2003, 28, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Buchan, K.A.; Martin–Robichaud, D.J.; Benfey, T.J. Measurement of dissolved ozone in sea water: A comparison of methods. Aquac. Eng. 2005, 33, 225–231. [Google Scholar] [CrossRef]
- Schröder, J. Potential and Limitations of Ozone in Marine Recirculating Aquaculture Systems-Guidelines and Thresholds for a Safe Application. Ph.D. Thesis, Christian-Albrechts-Universität, Kiel, Germany, 2011. [Google Scholar]
- Fisher, D.J.; Burton, D.T.; Yonkos, L.T.; Turley, S.D.; Ziegler, G.P. The relative acute toxicity of continuous and intermittent exposures of chlorine and bromine to aquatic organisms in the presence and absence of ammonia. Water Res. 1999, 33, 760–768. [Google Scholar] [CrossRef]
- Reiser, S.; Wuertz, S.; Schroeder, J.P.; Kloas, W.; Hanel, R. Risks of seawater ozonation in recirculation aquaculture—Effects of oxidative stress on animal welfare of juvenile turbot (Psetta maxima, L.). Aquat. Toxicol. 2011, 105, 508–517. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Jean-paul, B.; Triplet, S. Effect of oxidation–reduction potential on performance of European sea bass (Dicentrarchus labrax) in recirculating aquaculture systems. Aquac. Int. 2014, 22, 1263–1282. [Google Scholar] [CrossRef] [Green Version]
- Tango, M.S.; Gagnon, G.A. Impact of ozonation on water quality in marine recirculation systems. Aquac. Eng. 2003, 29, 125–137. [Google Scholar] [CrossRef]
- Scolding, J.W.; Powell, A.; Boothroyd, D.P.; Shields, R.J. The effect of ozonation on the survival, growth and microbiology of the European lobster (Homarus gammarus). Aquaculture 2012, 364, 217–223. [Google Scholar] [CrossRef]
- Cabillon, N.A.R.; Lazado, C.C. Mucosal barrier functions of fish under changing environmental conditions. Fishes 2019, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Good, C.; Davidson, J.; Welsh, C.; Snekvik, K.; Summerfelt, S. The effects of ozonation on performance, health and welfare of rainbow trout Oncorhynchus mykiss in low-exchange water recirculation aquaculture systems. Aquac. Eng. 2011, 44, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Richardson, L.B.; Burton, D.T.; Block, R.M.; Stavola, A.M. Lethal and sublethal exposure and recovery effects of ozone-produced oxidants on adult white perch (Morone americana Gmelin). Water Res. 1983, 17, 205–213. [Google Scholar] [CrossRef]
- Richards, J.G.; Farrell, A.P.; Brauner, C.J. Fish Physiology: Hypoxia; Academic Press: San Diego, CA, USA, 2009. [Google Scholar]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef] [PubMed]
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- Ritola, O.; Livingstone, D.R.; Peters, L.D.; Lindström-Seppä, P. Antioxidant processes are affected in juvenile rainbow trout (Oncorhynchus mykiss Walbaum) exposed to ozone and oxygen supersaturated water. Aquaculture 2002, 210, 1–19. [Google Scholar] [CrossRef]
- Ritola, O.; Livingstone, D.R.; Peters, L.D.; Lindström-Seppä, P. Glutathione-dependent defence system and monooxygenase enzyme activities in Arctic charr Salvelinus alpinus (L.) exposed to ozone. Aquaculture 2000, 185, 219–233. [Google Scholar] [CrossRef]
- Guilherme, S.; Crespo, R.; Azevedo, D.; Marques, A.; Santos, M.A.; Serradeiro, R.; Pacheco, M. DNA and chromosomal damage in Senegalese sole (Solea senegalensis) as side effects of ozone-based water treatment-Contribution to optimization of fish-farming practices. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 219, 68–76. [Google Scholar] [CrossRef]
- Biller, J.D.; Takahashi, L.S. Oxidative stress and fish immune system: Phagocytosis and leukocyte respiratory burst activity. An. Acad. Bras. Ciências 2018, 90, 3403–3414. [Google Scholar] [CrossRef] [Green Version]
- Fazio, F.; Piccione, G.; Saoca, C.; Caputo, A.R. Assessment of oxidative stress in Flathead mullet (Mugil cephalus) and Gilthead sea bream (Sparus aurata). Vet. Med. 2015, 60. [Google Scholar] [CrossRef] [Green Version]
- Wedemeyer, G. Physiology of Fish in Intensive Culture Systems; Springer Science & Business Media: Boston, MA, USA , 1996. [Google Scholar]
- Hermann, B.T.; Wuertz, S.; Vanselow, K.H.; Schulz, C.; Stiller, K.T. Divergent gene expression in the gills of juvenile turbot (Psetta maxima) exposed to chronic severe hypercapnia indicates dose-dependent increase in intracellular oxidative stress and hypoxia. Aquat. Toxicol. 2019, 206, 72–80. [Google Scholar] [CrossRef]
- Stiller, K.T.; Vanselow, K.H.; Moran, D.; Bojens, G. The effect of carbon dioxide on growth and metabolism in juvenile turbot Scophthalmus maximus L. Aquaculture 2015, 444, 143–150. [Google Scholar] [CrossRef]
- Stiller, K.T.; Vanselow, K.H.; Moran, D.; Riesen, G.; Koppe, W.; Dietz, C.; Schulz, C. The effect of diet, temperature and intermittent low oxygen on the metabolism of rainbow trout. Br. J. Nutr. 2017, 117, 784–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandnes, K.; Lie, Ø.; Waagbø, R. Normal ranges of some blood chemistry parameters in adult farmed Atlantic salmon, Salmo salar. J. Fish Biol. 1988, 32, 129–136. [Google Scholar] [CrossRef]
- Wuertz, S.; Schulze, S.G.E.; Eberhardt, U.; Schulz, C.; Schroeder, J.P. Acute and chronic nitrite toxicity in juvenile pike-perch (Sander lucioperca) and its compensation by chloride. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2013, 157, 352–360. [Google Scholar] [CrossRef]
- Mallatt, J. Fish gill structural changes induced by toxicants and other irritants: A statistical review. Can. J. Fish. Aquat. Sci. 1985, 42, 630–648. [Google Scholar] [CrossRef]
- Koppang, E.O.; Kvellestad, A.; Fischer, U. Fish Mucosal Immunity: Gill. In Mucosal Health in Aquaculture; Elsevier: Amsterdam, The Netherlands, 2015; pp. 93–133. [Google Scholar]
- Streit, B. Bioaccumulation of contaminants in Fish. In Fish Ecotoxicology; Birkhäuser: Basel, Switzerland, 1998; pp. 353–387. [Google Scholar]
- Lazado, C.C.; Voldvik, V. Temporal control of responses to chemically induced oxidative stress in the gill mucosa of Atlantic salmon (Salmo salar). J. Photochem. Photobiol. B Biol. 2020, 205, 111851. [Google Scholar] [CrossRef]
- Fukunaga, K.; Nakazono, N.; Suzuki, T.; Takama, K. Mechanism of Oxidative Damage to Fish Red Blood Cells by Ozone. IUBMB Life 1999, 48, 631–634. [Google Scholar] [CrossRef]
- Ritola, O.; Nakazono, N.; Suzuki, T.; Takama, K. Effects of in vitro exposure to ozone and/or hyperoxia on superoxide dismutase, catalase, glutathione and lipid peroxidation in red blood cells and plasma of rainbow trout, Oncorhynchus mykiss (Walbaum). Aquac. Res. 2002, 33, 165–175. [Google Scholar] [CrossRef]
- Ryu, H.S.; Han, J.H.; Song, J.A.; Choi, C.Y. Effects of Total Residual Oxidant on Oxidative Stress in Juvenile Olive Flounder Paralichthys Olivaceus. Ozone Sci. Eng. 2019, 42, 277–285. [Google Scholar] [CrossRef]
- Martínez-Álvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant defenses in fish: Biotic and abiotic factors. Rev. Fish Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Koren, H.S.; Devlin, R.B.; Graham, D.E.; Mann, R.; McGee, M.P.; Horstman, D.H.; Kozumbo, W.J.; Becker, S.L.; House, D.E.; McDonnell, W.F. Ozone-induced inflammation in the lower airways of human subjects. Am. Rev. Respir. Dis. 1989, 139, 407–415. [Google Scholar] [CrossRef]
- Fry, R.C.; Rager, J.E.; Zhou, H.; Zou, B.; Brickey, J.W.; Ting, J.; Lay, J.C.; Peden, D.B.; Alexis, N.E. Individuals with increased inflammatory response to ozone demonstrate muted signaling of immune cell trafficking pathways. Respir. Res. 2012, 13, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller Yury, I.; Choi, S.-H.; Wiesner, P.; Fang, L.; Harkewicz, R.; Hartvigsen, K.; Boullier, A.; Gonen, A.; Diehl, C.J.; Que, X. Oxidation-specific epitopes are danger-associated molecular patterns recognized by pattern recognition receptors of innate immunity. Circ. Res. 2011, 108, 235–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Secombes, C.J. The Function of Fish Cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef]
- Ingerslev, H.-C.; Rønneseth, A.; Pettersen, E.F.; Wergeland, H.I. Differential Expression of Immune Genes in Atlantic Salmon (Salmo salar L.) Challenged Intraperitoneally or by Cohabitation with IPNV. Scand. J. Immunol. 2009, 69, 90–98. [Google Scholar] [CrossRef]
- AnvariFar, H.; Keramat, A.; Miandare, H.P.-P.S.K.; Ouraji, H. Apoptosis in fish: Environmental factors and programmed cell death. Cell Tissue Res. 2017, 368, 425–439. [Google Scholar] [CrossRef]
- Igata, E.; Inoue, T.; Ohtani-Fujita, N.; Sowa, Y.; Tsujimoto, Y.; Sakai, T. Molecular cloning and functional analysis of the murine bax gene promoter. Gene 1999, 238, 407–415. [Google Scholar] [CrossRef]
- Kannan, K.; Jain, S.K. Oxidative stress and apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef]
- Roberts, R.J.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef]
- Beckmann, R.; Mizzen, L.; Welch, W. Interaction of Hsp 70 with newly synthesized proteins: Implications for protein folding and assembly. Science 1990, 248, 850–854. [Google Scholar] [CrossRef]
- Noble, C.; Gismervik, S.; Iversen, M.; Kolarevic, J. Welfare Indicators for Farmed Atlantic Salmon: Tools for Assessing Fish Welfare; Nofima: Tromsø, Norway, 2018. [Google Scholar]
- Sveen, L.R. Aquaculture Relevant Stressors and Their Impacts on Skin and Wound Healing in Post-Smolt Atlantic Salmon (Salmo Salar L.). Ph.D. Thesis, The University of Bergen, Bergen, Norway, 2018. [Google Scholar]
- Lazado, C.C.; Haddeland, S.; Timmerhaus, G.; Berg, R.S.; Merkin, G.; Pittman, K.; Pedersen, L.-F. Morphomolecular alterations in the skin mucosa of Atlantic salmon (Salmo salar) after exposure to peracetic acid-based disinfectant. Aquac. Rep. 2020, 17, 100368. [Google Scholar] [CrossRef]
- Nagasawa, K.; Lazado, C.; Fernandes, J.M. Validation of endogenous reference genes for qPCR quantification of muscle transcripts in atlantic cod subjected to different photoperiod regimes. In Aquaculture; IntechOpen: London, UK, 2012. [Google Scholar]
- Lazado, C.C.; Kumaratunga, H.P.S.; Nagasawa, K.; Babiak, l.; Giannetto, A.; Fernandes, J.M.O. Daily rhythmicity of clock gene transcripts in Atlantic cod fast skeletal muscle. PLoS ONE 2014, 9, e99172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solberg, M.F.; Kvamme, B.O.; Nilsen, F.; Glover, K.A. Effects of environmental stress on mRNA expression levels of seven genes related to oxidative stress and growth in Atlantic salmon Salmo salar L. of farmed, hybrid and wild origin. BMC Res. Notes 2012, 5, 672. [Google Scholar]
- Holm, H.J.; Skugor, S.; Bjelland, A.K.; Radunovic, S.; Wadsworth, S.; Koppang, E.O.; Evensen, O. Contrasting expression of immune genes in scaled and scaleless skin of Atlantic salmon infected with young stages of Lepeophtheirus salmonis. Dev. Comp. Immunol. 2017, 67, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Østbye, T.K.; KJÆR, M.A.; RØRÅ, A.M.B.; Torstensen, B.; Ruyter, B. High n-3 HUFA levels in the diet of Atlantic salmon affect muscle and mitochondrial membrane lipids and their susceptibility to oxidative stress. Rev. Fish Biol. Fish. 2011, 17, 177–190. [Google Scholar] [CrossRef]
- De la serrana, D.G.; Johnston, I.A. Expression of Heat Shock Protein (Hsp90) Paralogues Is Regulated by Amino Acids in Skeletal Muscle of Atlantic Salmon. PLoS ONE 2013, 8, e74295. [Google Scholar] [CrossRef] [Green Version]
- Sanden, M.; Olsvik, P.A.J.B.P. Intestinal cellular localization of PCNA protein and CYP1A mRNA in Atlantic salmon Salmo salar L. exposed to a model toxicant. BMC Physiol. 2009, 9, 3. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | ORP (mV) | Hct | Cl− | Na+ | Mg2+ |
|---|---|---|---|---|---|
| very high * | 500 | 52.9 ± 5.9 a | 121.0 ± 8.1 b | 143.5 ± 26.7 | 1.4 ± 0.2 b |
| high | 425 | 60.7 ± 5.6 b | 127.3 ± 3.3 ab | 159.3 ± 2.4 | 1.2 ± 0.1 ab |
| medium | 350 | 56.1 ± 6.0 ab | 128.2 ± 1.2 a | 161.5 ± 2.9 | 1.1 ± 0.1 a |
| low | 300 | 51.6 ± 4.7 a | 127.0 ± 1.7 ab | 162.7 ± 2.3 | 1.1 ± 0.1 a |
| control | 250 | 53.7 ± 3.2 a | 128.0 ± 1.9 a | 161.8 ± 1.2 | 1.1 ± 0.2 a |
| Gene name | Abbreviation | Sequence (5′–3′) | Reference |
|---|---|---|---|
| glutathione reductase | gr-F | CCAGTGATGGCTTTTTTGAACTT | [61] |
| gr-R | CCGGCCCCCACTATGAC | ||
| cu/zn superoxide dismutase | cu/znsod-F | CCACGTCCATGCCTTTGG | |
| cu/znsod-R | TCAGCTGCTGCAGTCACGTT | ||
| mn superoxide dismutase | mnsod-F | GTTTCTCTCCAGCCTGCTCTAAG | |
| mnsod-R | CCGCTCTCCTTGTCGAAGC | ||
| glutathione peroxidase | gp-F | GATTCGTTCCAAACTTCCTGCTA | |
| gp-R | GCTCCCAGAACAGCCTGTTG | ||
| interleukin 1β | il1b-F | AGGACAAGGACCTGCTCAACT | [50] |
| il1b-R | CCGACTCCAACTCCAACACTA | ||
| interleukin 10 | il10-F | GGGTGTCACGCTATGGACAG | |
| il10-R | TGTTTCCGATGGAGTCGATG | ||
| immunoglobulin m | igm-F | TGAGGAGAACTGTGGGCTACACT | [62] |
| igm-R | TGTTAATGACCACTGAATGTGCAT | ||
| immunoglobulin t | igt-F | GGTGGTCATGGACGTACTATTT | |
| igt-R | CCTGTGCAGGCTCATATCTT | ||
| beta-defensin 1-like | defb1l-F | ATTTAGAAGACGTGGGCG | |
| defb1l-R | GGATGCTCAAACTACAGTGG | ||
| heat shock protein 70 | hsp70-F | CCCCTGTCCCTGGGTATTG | [61] |
| hsp70-R | CACCAGGCTGGTTGTCTGAGT | ||
| bcl-2 associated x protein | bax-F | TGACAGATTTCATCTACGAGCGGG | [63] |
| bax-R | GCCATCCAGCTCATCTCCAATCT | ||
| caspase 3a | casp3a-F | ACAGCAAAGAGCTAGAGGTCCAACAC | |
| casp3a-R | AAAGCCAGGAGAGTTTGACGCAG | ||
| caspase 7 | casp7-F | AGCGAGTGGGCAAGTGCATCA | |
| casp7-R | CGTCGAAGCCCAGGCTCTTAAA | ||
| elongation factor alpha-1 | ef1a-F | GAATCGGCTATGCCTGGTGAC | [64] |
| ef1a-R | GGATGATGACCTGAGCGGTG | ||
| ribosomal protein L13 | rpl13-F | CGCTCCAAGCTCATCCTCTTCCC | |
| rpl13-R | CCATCTTGAGTTCCTCCTCAGTGC | ||
| acidic ribosomal protein | arp-F | TCATCCAATTGCTGGATGACTATC | [65] |
| arp-R | CTTCCCACGCAAGGACAGA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stiller, K.T.; Kolarevic, J.; Lazado, C.C.; Gerwins, J.; Good, C.; Summerfelt, S.T.; Mota, V.C.; Espmark, Å.M.O. The Effects of Ozone on Atlantic Salmon Post-Smolt in Brackish Water—Establishing Welfare Indicators and Thresholds. Int. J. Mol. Sci. 2020, 21, 5109. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145109
Stiller KT, Kolarevic J, Lazado CC, Gerwins J, Good C, Summerfelt ST, Mota VC, Espmark ÅMO. The Effects of Ozone on Atlantic Salmon Post-Smolt in Brackish Water—Establishing Welfare Indicators and Thresholds. International Journal of Molecular Sciences. 2020; 21(14):5109. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145109
Chicago/Turabian StyleStiller, Kevin T., Jelena Kolarevic, Carlo C. Lazado, Jascha Gerwins, Christopher Good, Steven T. Summerfelt, Vasco C. Mota, and Åsa M. O. Espmark. 2020. "The Effects of Ozone on Atlantic Salmon Post-Smolt in Brackish Water—Establishing Welfare Indicators and Thresholds" International Journal of Molecular Sciences 21, no. 14: 5109. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145109