1. Introduction
Cancer metastasis is associated with a high mortality rate and is the primary cause of cancer morbidity. The spread of tumor cells from their primary origin occurs mainly via blood and lymphatic vessels [
1]. The 3-dimensional (3D) fast growth of tumors creates a dynamic microenvironment with elevated demands for nutrients and oxygen exchange as well as metabolic waste disposal which are accommodated by the expedited angiogenesis process [
2,
3].
Angiogenesis is regulated by a dynamic balance between inducer and inhibitor molecules. Tumor angiogenesis is a multistep process triggered by diverse factors secreted by the hypoxic tumor as well as by the host cells [
4]. This process is characterized by a formation of new blood vessels from pre-existing vessels [
2,
3].
Similar to angiogenesis, the growth and formation of new lymphatic vessels, known as lymphangiogenesis, occurs in response to multiple triggers [
5]. Lymphangiogenesis was shown to play a critical role in tumor progression and metastasis [
6]. Unlike hematologic vessels, lymphatic vessels are thin and consist of discontinuous membranes that enable the penetration of tumor cells into the lymphatic circulation and the adjacent capillary lumens. In response to secreted signals, both lymphatic and vascular endothelial cells proliferate and migrate toward the stimulus, assembling into tube-like structures [
1].
Several pathways were identified as promoting angiogenesis and lymphangiogenesis, the most studied and dominant one being the vascular endothelial growth factor (VEGF) pathway. VEGF, which is overexpressed in most cancers and functions as a crucial regulator in angiogenesis and lymphangiogenesis [
3,
7,
8,
9], has become an important target for inhibiting tumor metastasis. Among the various VEGF family members, VEGF-C and VEGF-D are secreted by the tumor and stromal cells, playing a dominant role in mediating tumor-related lymphangiogenesis [
6,
10,
11,
12].
However, there are many additional mediators that are involved in capillary formation and remodeling. Among these mediators is MetAp2, which belongs to a family of post-translational modification enzymes, acting downstream to VEGF. There are two forms of MetAp in eukaryotes: MetAp1 and MetAp2, the latter having received more interest. This is because the induction of Metap2 was shown to mediate cell proliferation and lead to G
1 arrest when inhibited [
13,
14,
15,
16,
17], since it affects protein synthesis via the post-translation removal of methionine [
18,
19,
20,
21].
Given MetAp2’s major role in protein synthesis in activated vascular endothelial cells, we hypothesize that MetAp2 also affects lymphangiogenesis by mediating the lymphatic endothelium formation via shared mechanisms. In patient-derived breast cancer tissues, we found that MetAp2 is expressed and co-localized with lymphatic vessels. At the cellular level, we found that the basal expressions of MetAp2 and its activities in human lymphatic endothelial cells (LECs) versus vascular endothelial cells (VECs) were comparable, with slightly higher levels in VECs. In functionality assays including cell proliferation, adhesion, and tube formation, LECs showed suppression upon exposure to TNP-470, a MetAp2 inhibitor. In vivo, in a non-cancerous context using the zebrafish developmental model, the specific inhibition of MetAp2 led to a significant reduction in the development of lymphatic vessels. Parachordal cells (PACs), which make up the lymphatic vessels, were observed in only 0.76 out of 3 segments after treatment with 500 µM of the MetAp2 inhibitor. Moreover, in a subcutaneous (S.C.) murine model using B16/F10 melanoma tumors overexpressing VEGF-C, systemic treatment with 30 mg/kg q.o.d induced over a 50% reduction in the tumors’ volume. A histological examination of the tumors suggested an associated dual reduction of angiogenesis and lymphangiogenesis induced by MetAp2 inhibition.
Taken together, our data suggest that MetAp2 is a mediator of both angiogenesis and lymphangiogenesis and that its inhibition may slow cancer progression and metastasis.
3. Discussion
Tumor metastasis is the leading cause of mortality worldwide and is the main cause for cancer-related complications. Most current treatments for invasive cancers are not sufficiently effective [
22,
23]. Despite the initial promise of anti-angiogenic therapies, the clinical successes of monotherapies have only been partial [
3,
24].
There is a critical need for complimentary approaches that improve clinical outcomes and lead to higher survival rates in cancer patients. Intervening in the molecular pathways that are involved in lymphangiogenesis may offer a potential therapy target for combating tumor progression and metastasis. While ample research has focused on MetAp2’s involvement in angiogenesis and endothelial cell biology, its role in lymphangiogenesis and in lymphatic endothelial cell biology has not been sufficiently studied.
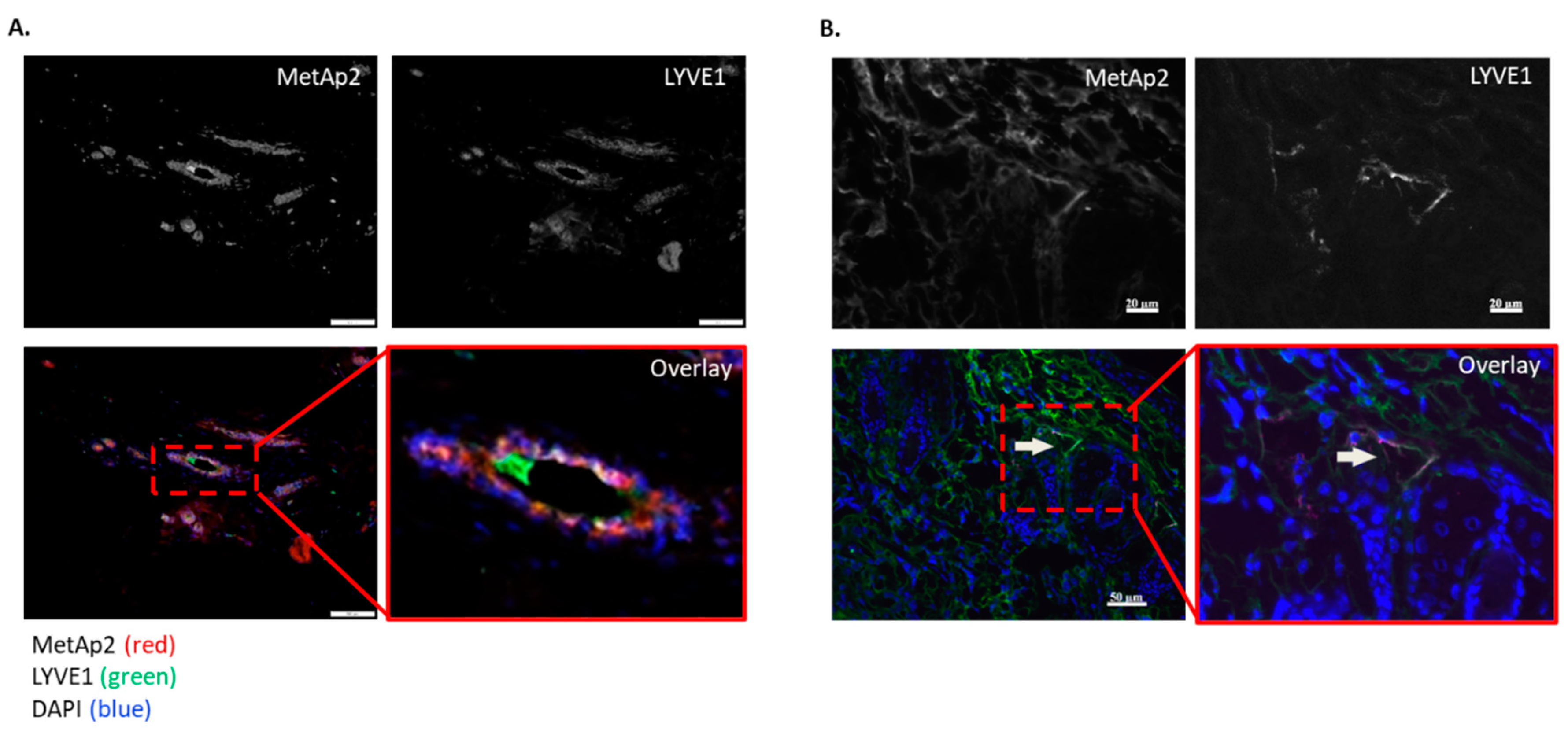
Here we demonstrated with a series of experiments the effect of MetAp2 inhibition on LECs’ cellular functions and, more specifically, on lymphangiogenesis. Histological samples of human breast cancers showed a high expression of MetAp2 and the co-localization of MetAp2 with lymphatic vasculature (
Figure 1). Breast cancer treatment often involves the removal of the axillary lymph nodes where tumor metastasis is usually found [
25]. Co-localization may suggest that the lymphatic development is linked with higher MetAp2 levels.
It is well established that MetAp2 is overexpressed in activated endothelium, but its level in primary LECs and its specific role in lymphangiogenesis is yet to be determined [
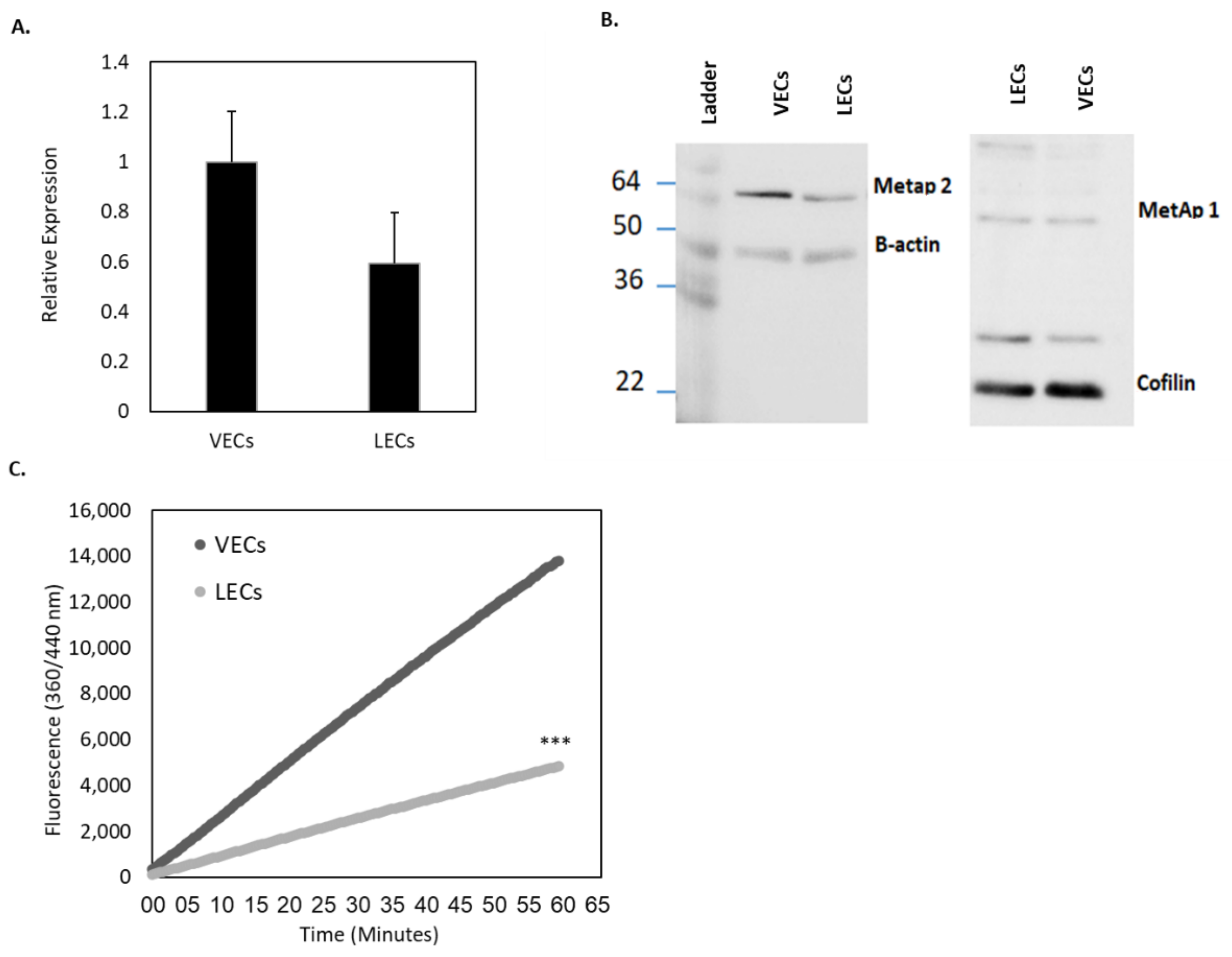
26]. We found that MetAp2 mRNA and the enzyme itself were highly expressed in active LECs (
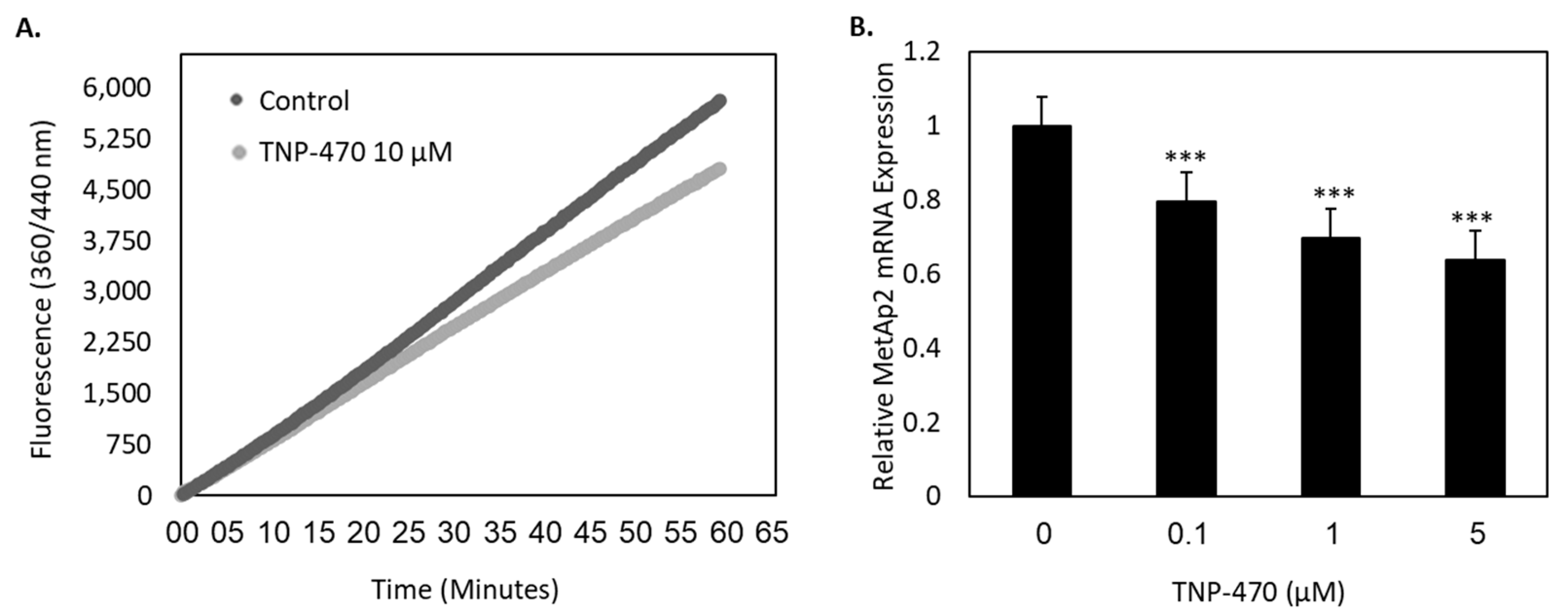
Figure 2). The enzymatic expression and the proteolytic activity were also found to be susceptible to biochemical inhibition by TNP-470 (
Figure 3).
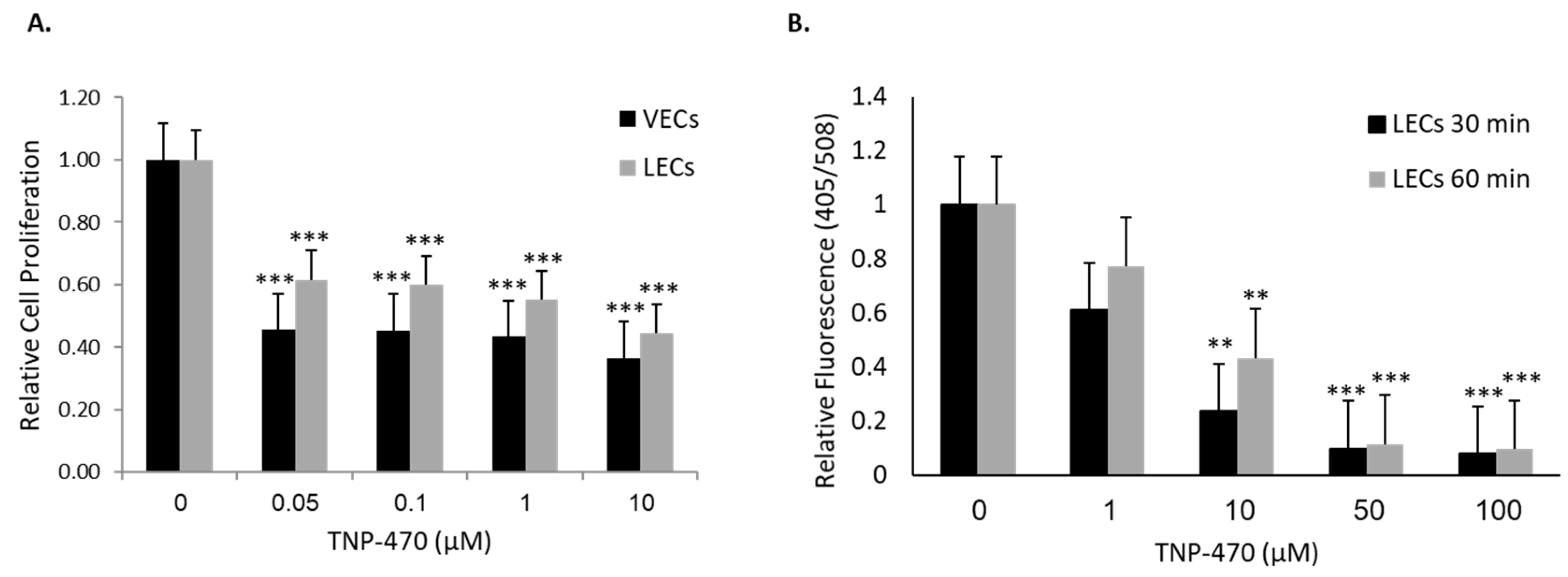
After establishing MetAp2’s basal expression and activity, we investigated its effect on cellular functionality in LECs. MetAp2 inhibition in the vascular endothelium is known to regulate cell proliferation through cell cycle arrest in the late G
1 phase [
27]. We found that HMVEC-dLyAd’s cell proliferation was impaired when MetAp2 was inhibited. This was observed when even low concentrations of the antagonist were used. The lowest concentration that was used, 0.05 µM, reduced proliferation by ~50%. These findings indicate the potential antagonistic effects of MetAp2 inhibitors on lymphatic vessel formation. Datta et al. suggested that
fumagillin and its analogs stabilize MetAp2 in a certain configuration, rather than inhibiting it, which in turn causes an elevation in its affinity for extracellular signal-regulated kinases (ERK1/2). This, consequently, leads to a reduction in MetAp2’s anti-inhibitory protection of eukaryotic initiation factor 2 (eIF2), which in turn decreases the total cellular protein synthesis, induces cell-cycle arrest and impairs the cell’s ability to function [
28,
29]. This corresponds with our results which showed that treatment with a fumagillin analogue had a relatively small effect on MetAp2 expression in contrast to the cell functionality which was substantially affected. It is probable that the differences in the enzyme levels in VECs, when compared to those in LECs, may explain the higher susceptibility of VECs to enzymatic inhibition. Moreover, the LECs that were used (HMVEC-dLyAd) might contain a small percentage of endothelial cells which, when purified, might show even more significant differences between the two cell lines.
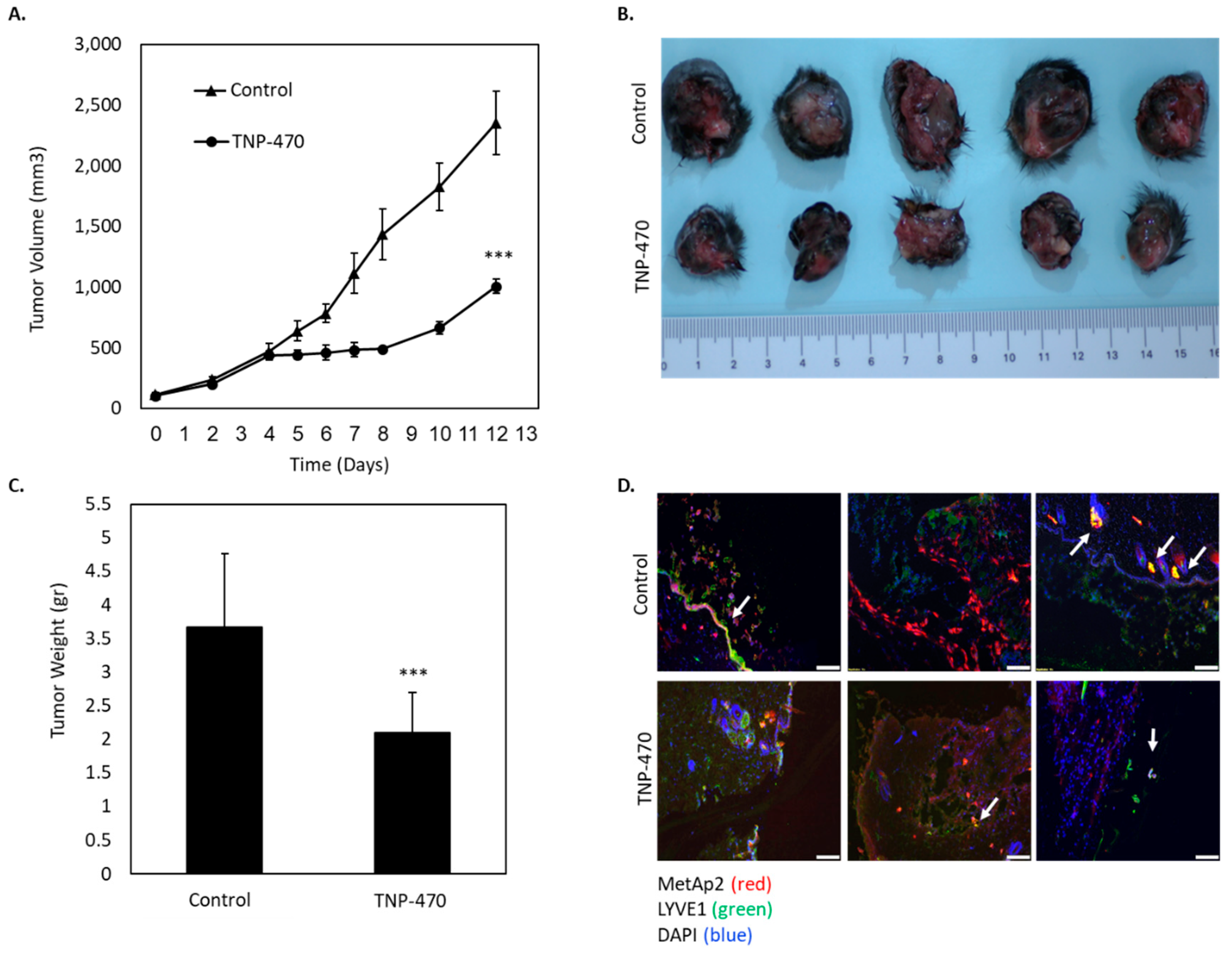
These observations correlate with our in vivo histological analyses which showed that MetAp2 inhibition affected LECs’ remodulation while maintaining relatively steady levels of the MetAp2 enzyme. The immunofluorescence of VEGF-C overexpressing murine melanoma tissue sections showed that in treated tissues, LYVE1+ cells were organized more sporadically and less collectively as vessels as compared with untreated samples. This may be a result of a disruption in the intercellular interactions of lymphatic cells, as confirmed in the tube formation assay, occurring without substantially lowering the MetAp2 levels in these affected cells.
Forming new lymphatic capillaries depends on the cells’ ability to migrate, form cell-cell interactions and adhere to the extracellular matrix (ECM) [
30,
31]. MetAp2 inhibition resulted in a significant impairment in the cells’ adherence capabilities to the ECM. Assembly of cells into vessels requires cell adhesion and proliferation. These two processes were impaired when MetAp2 was inhibited (
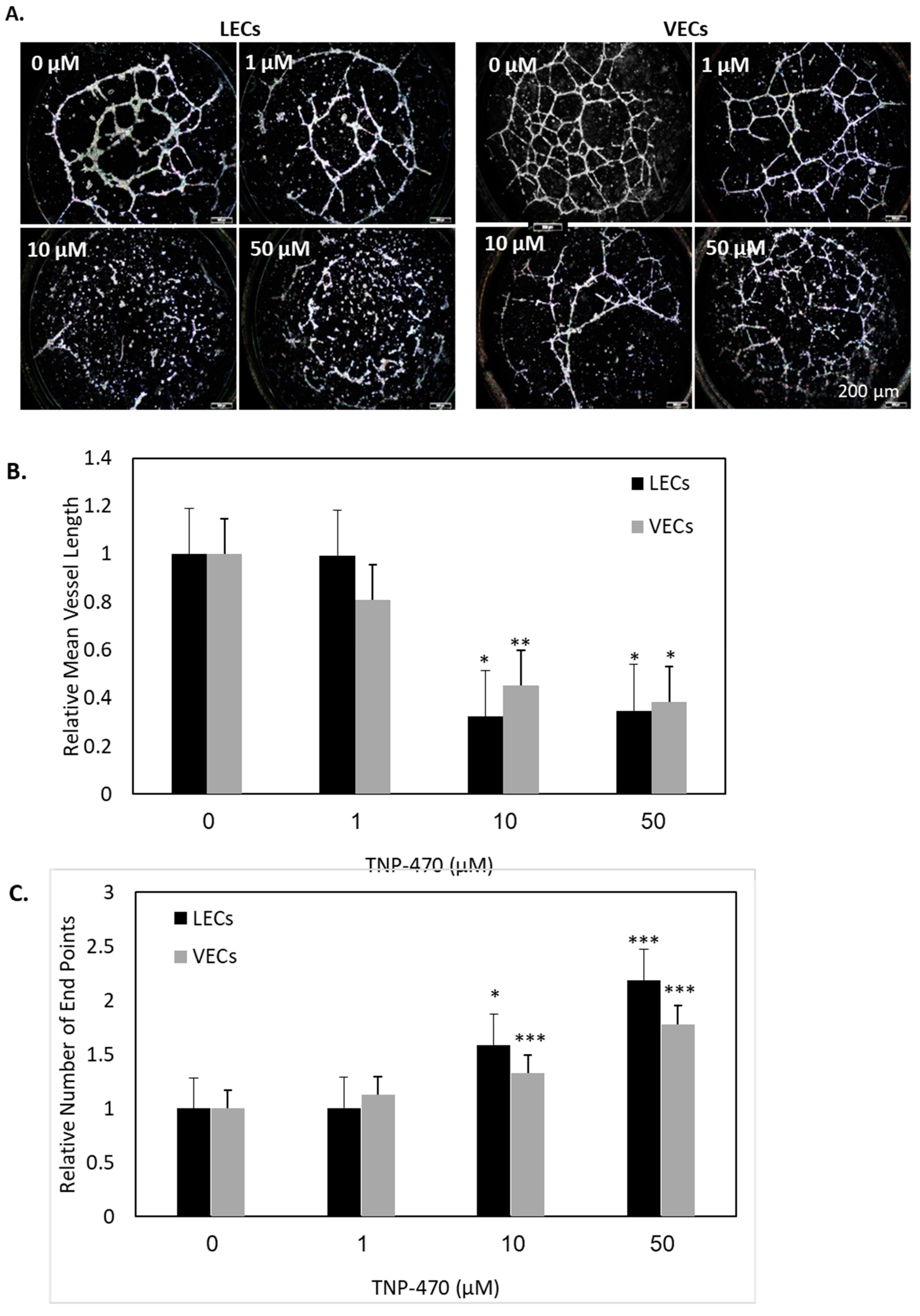
Figure 4). In line with this, HMVEC-dLyAd cells’ morphogenesis was remarkably affected, as demonstrated by the tube formation assay (
Figure 5). The cells’ ability to interact was notably reduced, as measured by the high number of vessel ends.
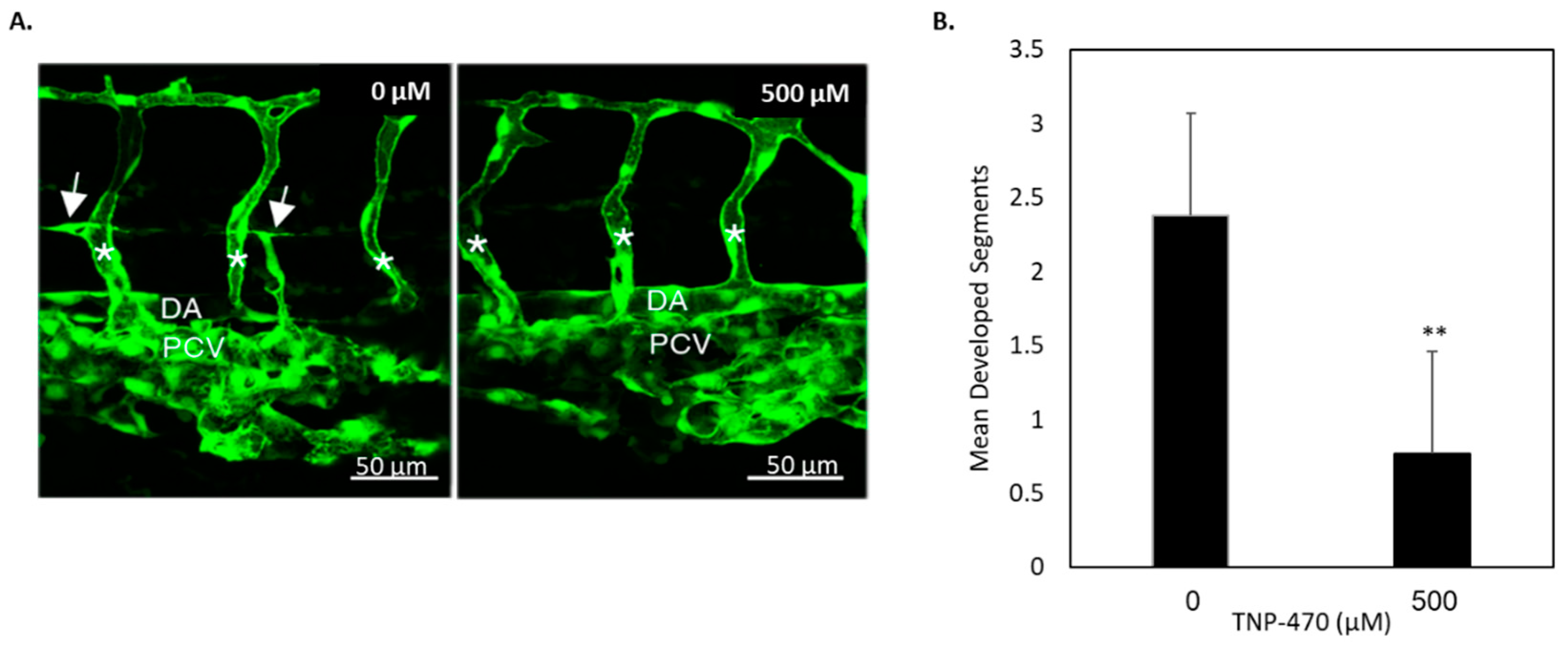
To determine whether our in vitro findings are also present in vivo, cancer and non-cancer animal models were utilized to investigate MetAp2’s activity. The zebrafish model provides a convenient way for the real-time visualization of developmental processes, especially the
Tg(
fli1:EGFP)y1 model which enables one to monitor vasculogenesis. In the zebrafish model, which is similar to mammalian development, we found MetAp2 to be highly involved in lymphatic capillary formation and sprouting. While trunk endothelial vessels developed by 29 hpf, PAVs developed at about 48 hpf, creating an ideal window of time for the inhibition of lymphatic vessel formation without the impairment of blood vasculature development. When zebrafish with a developed blood vasculature were treated with a MetAp2 inhibitor, a significant reduction in lymphatic development was observed (
Figure 6), implying MetAp2’s significance in embryonic lymphangiogenesis.
Several studies showed that tumor cells overexpressing VEGF-C created a richer lymphangiogenic microenvironment when compared with wild type tumors [
6,
10]. In order to enhance lymphangiogenesis in tumors, we injected C57/BL6J mice with melanoma B16/F10 cells overexpressing VEGF-C. Treatment with a MetAp2 inhibitor led to a more than 50% decrease in the tumors’ size, associated with reduced lymphatic vascularization (
Figure 7). It should be noted that B16/F10-VEGF-C cells showed a significant increase in proliferation when compared with B16/F10 cells, which might explain the first cells’ high sensitivity to the MatAp2 inhibitor (
Figure S2).
Taken together, our data demonstrate that MetAp2 has a dual role in blood and lymphatic vascularization, with specific significant effects on cell recruitment and endothelium remodeling. In our previous studies, we showed the preventative effect of an orally available formulated TNP-470 in melanoma liver metastases in an intrasplenic injection model [
32]. These significant results were mainly attributed to its anti-angiogenic activity. However, our current observations suggest the involvement of MetAp2 in lymphangiogenesis and the potential to prevent a lymphatic pro-metastatic process by inhibiting MetAp2. This is of particularly high clinical relevance since lymphatic metastasis is common in different cancers and serves as a prognosis indicator of a disease’s aggressiveness. MetAp2 may prove to be an effective target for both angiogenesis and lymphangiogenesis, and its inhibition may be used one day to supplement current methods for cancer treatment.
4. Materials and Methods
4.1. Reagents
TNP-470 (O-(Chloroacetylcarbamoyl)fumagillol), also known as AGM-1470, was purchased from MedChem Partners (Lexington, MA, USA).
4.2. Human Breast Carcinoma Histological Staining
The study complies with the Declaration of Helsinki. The study protocol was approved by the Institutional Ethics Committee (0346-12, 04-02-2013). Paraffin-embedded sections were prepared and used for immunofluorescence staining. Paraffin-embedded sections were deparaffinized and rehydrated. Antigen retrieval was performed by immersing the samples in TRIS-EDTA buffer and heating in a microwave oven for 15 min. PBS with 0.1% v/v Tween 20 was used to wash the samples, following each step of the procedure. After 15 min of permeabilization with 0.1% Triton, slides were incubated for 1.5 h at room temperature (RT) with a blocking reagent (3% serum in PBS) in order to reduce non-specific antibody (Ab) binding. Sections were incubated overnight at 4 °C with primary Abs in a blocking solution. The primary Abs included: anti-MetAp2 and anti-LYVE1 for lymphatic microvessel (LV) staining. LV and MetAp2 were detected by Alexa Flour® 488 and Alexa Flour® 647 secondary Abs, respectively, and nuclei were detected by DAPI staining.
4.3. Cell Culture
HUVECs (VECs) and HMVEC-dLyAd cells (LECs) were purchased from Lonza (Walkersville, MD, USA). All cells were characterized before use, mycoplasma-free, using an EZ-PCR Mycoplasma Test Kit (Biological Industries), and were used for the experiments up to passage 12. All cells were kept in a humidified incubator at 37 °C with 5% CO2. HUVECs were maintained in a specific medium supplemented with the PeproGrow-MacroV kit (ENDO-BM & GS-MacroV, PeproTech) and penicillin/streptomycin. For HMVEC-dLyAd cells, an EGMTM-2 MV Microvascular Endothelial Cell Growth Medium (Lonza, Walkersville, MD, USA) supplemented with penicillin/streptomycin was used. For starvation, primary cells were cultivated in their appropriate medium without supplementation. B16/F10 murine melanoma cell lines were obtained from ATTC (Manassas, VA, USA) and cultivated in DMEM supplemented with 10% FCS and penicillin/streptomycin.
4.4. Western Blot
Cells were lysed with RIPA buffer in a protease inhibitor cocktail (Sigma, S8820) for 30 min on ice. Lysates were centrifuged, and the supernatant was collected. Protein content was determined according to the BCA Protein Assay kit (PierceTM, Thermo Fisher Scientific, Cambridge, MA, USA). Proteins (15 μg protein) were separated by a 12.5% Tris-glycine SDS-PAGE and transferred onto a Polyvinylidene difluoride membrane (Millipore Corporation, Billerica, MA, USA). Membranes were blocked for 2 h and then incubated with anti-MetAp2 abs (Ab134124, Abcam, Cambridge, UK) overnight at 4 °C in TBST containing 5% BSA. Membranes were washed three times for 5 min in TBST, incubated with a 1:5000 dilution of goat anti-rabbit secondary ab conjugated to horseradish peroxidase for 1 h (Ab97080, Abcam). β-Actin (Ab49900, Abcam) was used as the loading control.
4.5. Total RNA/Protein Extraction and Expression Analysis
Total RNA was isolated from each sample using the RNeasy Kit (Qiagen, Valencia, CA, USA). 5 µg of total RNA from each sample was reverse transcribed with the Superscript First-strand cDNA synthesis kit (Invitrogen, Carlsbad, CA, USA) using a random primer. qRT-PCR for MetAp2 (Hs00199152_m1) and 18S rRNA (Hs03003631_g1) was performed in triplicate using TaqMan Gene Expression assays (Life Technologies, Grand Island, NY, USA). The relative level of each RNA sample was calculated using the ∆∆Ct method normalized to the corresponding 18S rRNA levels, n = 3.
4.6. Activity Assay
HUVECs and HMVEC-dLyAd cells were subcultivated using trypsin, then centrifuged and counted. To obtain a homogenate containing an equivalent cell number, cells were centrifuged again and re-suspended in an appropriate volume of cold RIPA buffer containing a protease inhibitor cocktail. Cells were disrupted using a probe sonicator on ice (Sonic Ruptor 400, OMNI International). Insoluble cellular components were removed by centrifugation at 15,000 RPM for 10 min at 4 °C. The protein content of the supernatant was determined according to the Bradford protein assay using BSA as the standard [
33]. The enzymatic activity was tested on 5 μg of protein using L-Met-AMC (Santa Cruz Biotechnology) as a substrate. The reaction was performed in an assay buffer (pH 7.5) containing 50 mM HEPES, 0.1 mM CoCl2, 100 mM NaCl and 1 mg/mL PEG 6000, in a final volume of 100 μL. Fluorescence was measured every 20 sec for 1 h at 25 °C using a plate reader (Synergy HT Multi-Mode Microplate Reader, BioTek),
n = 2.
4.7. Activity Assay Following MetAp2 Inhibition
HMVEC-dLyAd cells were grown to 80–90% confluency and were then washed with cold PBS and scraped on ice using a protease inhibitor-free RIPA buffer. A total of three HMVEC-dLyA dishes were scraped using the same buffer, which was transferred from one plate to the other. Insoluble cellular components were removed by centrifugation at 15,000 RPM for 10 min at 4 °C. The protein content of the supernatant was determined according to the Bradford protein assay using BSA as the standard. The enzymatic assay was performed using 5 µg of protein per sample, as described previously. To test the inhibitory effect of TNP-470, samples were incubated with the inhibitor for 15 min at RT before adding the substrate. An increase of fluorescence, due to substrate degradation during the enzymatic assay, was measured every 25 seconds for 1 h at 25 °C using a plate reader, as previously mentioned, n = 2.
4.8. Proliferation Assay
HUVECs and HMVEC-dLyAd cells seeded in 96-well plates (2000 cells/well) were exposed to a range of TNP-470 concentrations (0–10 μM) for 72 h. After incubation, MTT (Sigma Aldrich, St. Louis City, MO, USA) was added (0.5 mg/mL) into each well for viability detection and incubated at 37 °C and 5% CO2 for 3 h. The absorbance was measured at 570 nm using a plate reader, n = 5.
4.9. Cell Adhesion Assay
HMVEC-dLyAd cells were stained with a lipophilic tracer, DiO (5 μL/1 × 106 cells), and seeded on a gelatin-coated 96-well plate (12,000 cells/well). Cells were treated with TNP-470 at various concentrations (0–100 μM) and incubated for 30 and 60 min. Unadhered cells were removed, and the relative fluorescence of each cell population was quantified using a plate reader, as previously described, n = 4.
4.10. Tube Formation Assay
The morphogenic potential of HUVECs and HMVEC-dLyAd cells to form capillary-like structures in vitro was evaluated by seeding cells on Matrigel, a basement membrane matrix (Corning, NY, USA). 10 μL of Matrigel was dispensed into each well in a µ-Plate Angiogenesis 96-well (ibidi GmbH Martinsried, Germany) and allowed to polymerize for 1 h at RT. HMVEC-dLyAd cells and HUVECs (12,000 cells/well) were synchronized by starvation in a serum-free medium for 3 h prior to seeding on the polymerized gel. 1.5 h after seeding, TNP-470 in various concentrations (0–50 μM) was added and left to incubate overnight. Images were taken using an inverted fluorescence microscope (Nikon ECLIPSE Ni-E). The mean vessel length and the total number of end points were quantified using the AngioTool image analysis software, n = 8.
4.11. Animal Models
Animal experiments were approved by the Institutional Animal Care and Use Committee of the Faculty of Medicine of the Hebrew University and followed the guidelines on the use of laboratory animals. 7–8-week-old male C57BL/6J mice were purchased from Harlan. TNP-470 was dissolved in 70% ethanol to obtain a stock solution of 15 mg/mL. Prior to injecting the mice I.P., the stock solution was diluted at 1:5 using sterile saline.
4.12. Zebrafish Analysis
Tg(
fli1:EGFP)y1 zebrafish were used in this study [
34]. To block pigmentation, embryos were raised from 22 hpf in the presence of 0.003% N-Phenylthiourea (PTU; Sigma-Aldrich #P7629). 29 hpf embryos were placed in each well of a 24-well plate in 1 mL egg water containing 0.1% DMSO (control) or 500 µM TNP-470. Embryos were kept at 28.5 °C until imaging took place. For imaging, live embryos were anaesthetized using Tricaine mesylate and were mounted in 0.5% low-melting-point agarose (SeaPlaque, Lonza). Images were acquired using a Zeiss LSM700 confocal microscope and an Axio Imager M2 compound microscope equipped with a 40 × 1.0 NA water objective. Images were exported as TIFF files using ZEN 2009 LE software (Zeiss), and figures were assembled using the Adobe Photoshop CS4 software,
n = 17.
4.13. B16/F10-VEGF-C Plasmid Amplification and Cell Transfection
B16/F10 cells were transfected with VEGF-C plasmids as follows: competent E. coli (DH5α, Biolab, Jerusalem, Israel) were transformed by heat shock using 1 µg of VEGF-C plasmid (#EX- K2961-M03) or control plasmid (#EX-NEG-M03) (Gene Copoeia Inc., Rockville, MD, USA), respectively. Cells were then transferred to 1 mL of LB medium and incubated for 1 h at 37 °C under constant shaking. The transformed E. coli were spread on LB-Agar plates containing Ampicillin (100 µg/mL) for selection. The petri dishes were incubated overnight at 37 °C, and a selected colony of each plasmid was transferred to 5 mL of Ampicillin LB medium for culture, and an amplification step was performed. Bacteria were centrifuged at 5000× g for 10 min, and plasmid purification was performed using the GenElute HP plasmid kit (Sigma-Aldrich, St Louis, MO, USA). The DNA concentration and its purity were determined using a microplate reader. Cells were seeded at 200,000 cells/well in a 6-well plate. After 48 h, cells reached a 90% confluency and were then transfected with 4 µg of plasmid using 10 µL/well of Lipofectamine 2000 (Thermo Fisher Scientific, Van Allen Way, Carlsbad, CA, USA). After an additional 24 h, the selection was performed using 3 mg/mL of Gentamycin (G418 Sulfate, Gold Biotechnology, St Louis, MO, USA).
4.14. Murine Melanoma Tumors
S.C. tumors were developed in the lower backs of C57BL/6J mice by injecting 1.5 × 106 B16/F10-VEGF-C cells or B16/F10-control cells (transfected with control plasmid) in 100 μL of PBS containing Matrigel (1:10). Tumors were transcutaneously measured every 1–2 days with a digital caliper, and volumes were calculated according to the ellipsoid formula (Length x Width2 × 0.52). Mice were weighted and observed daily throughout the experiment. Mice were treated I.P. q.o.d with 30 mg/kg of TNP-470. On day 12, the mice were sacrificed, and the tumors were surgically removed, weighted, frozen in OCT and cryo-sectioned with Leica CM-1950 for an immunofluorescence analysis, n = 8–10.
4.15. Statistical Analysis
The results are presented as the mean ± SEM. Studies containing two groups were assessed using the unpaired two-tailed Student’s t-test. Studies containing more than three groups were compared and analyzed using a one-way analysis of variance (ANOVA), and significant differences were detected using Tuckey’s multiple comparison post-test. Differences were considered statistically significant for p < 0.05.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






