Parity Attenuates Intraepithelial Corneal Sensory Nerve Loss in Female Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Age-Related Loss of Axon Density Observed In Naturally Aged Female Nulliparous Mice Is Attenuated in Multiparous Mice
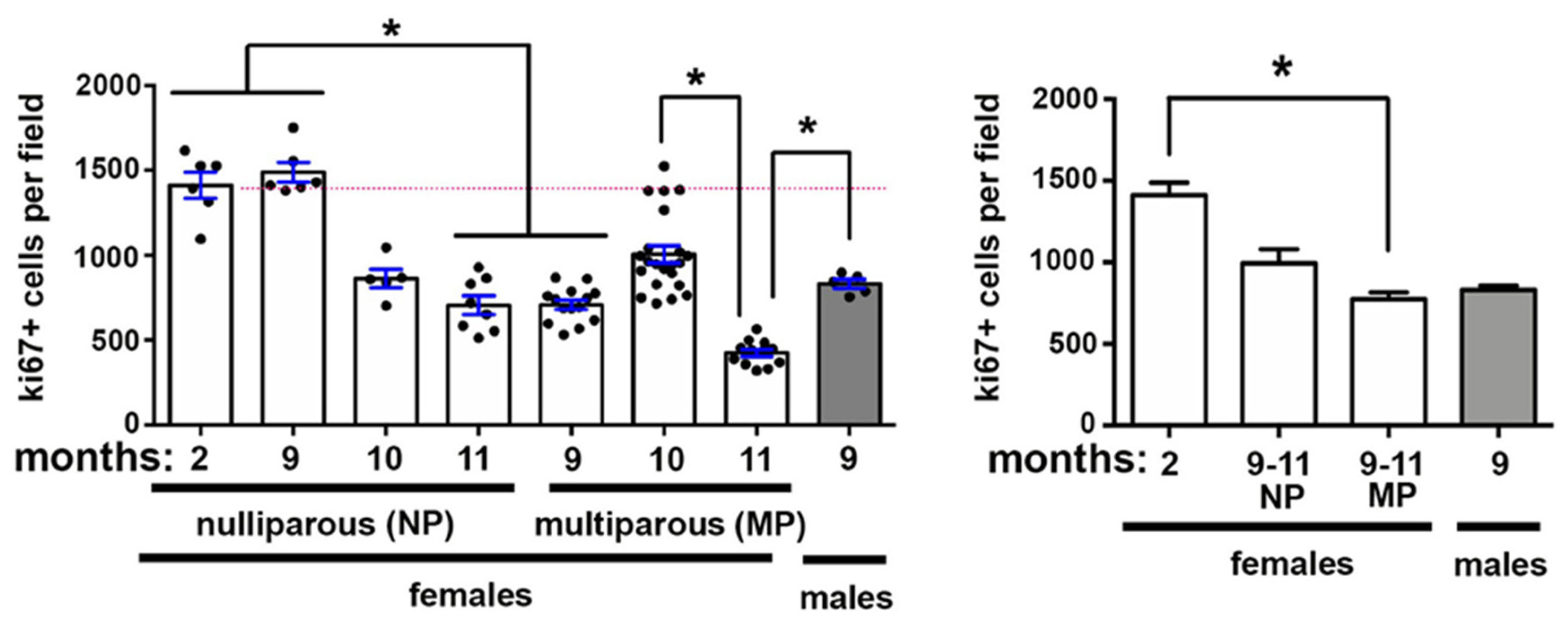
2.2. Corneal Epithelial Cell Proliferation Changes Also Occur With Aging and Pregnancy
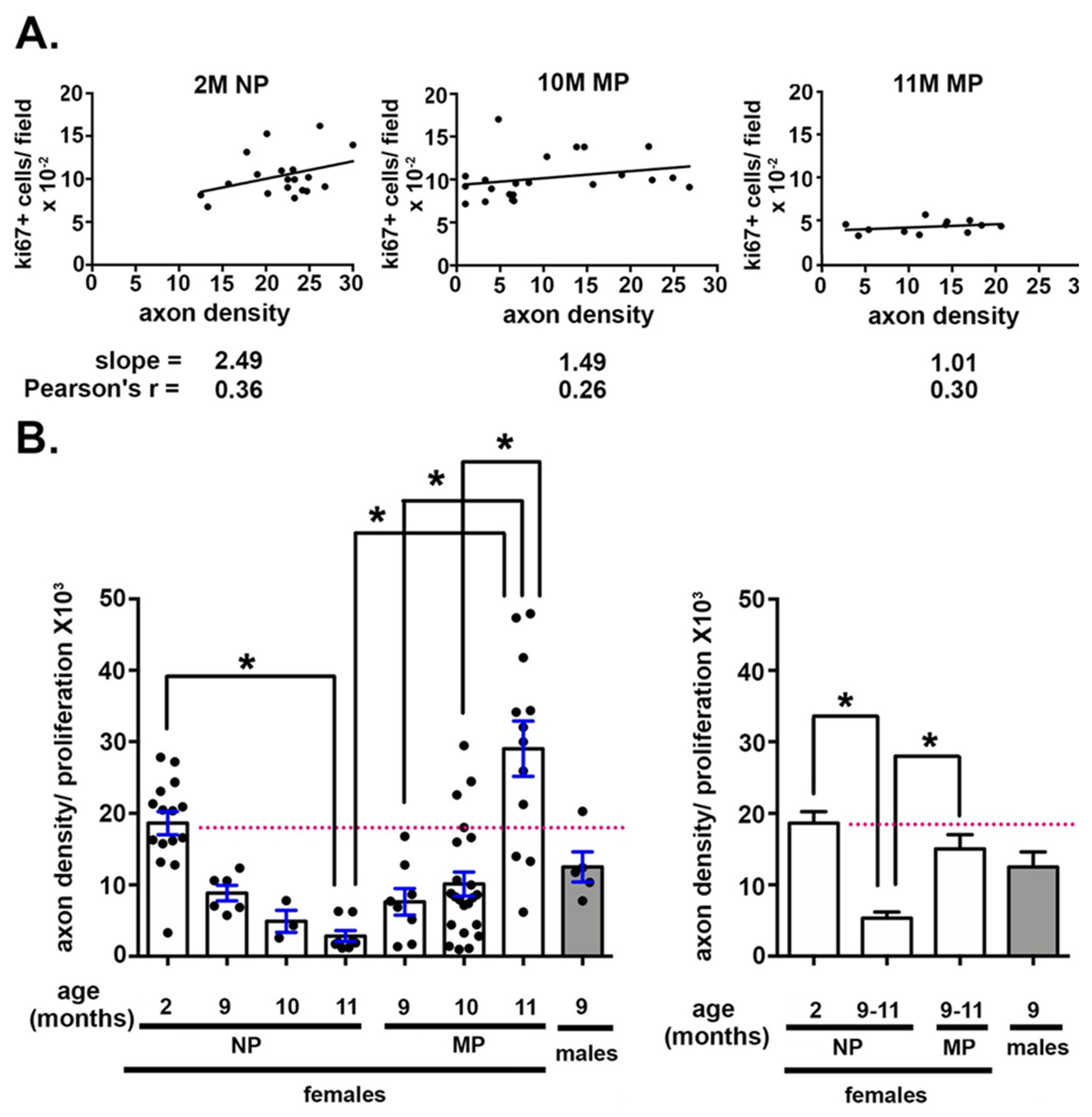
2.3. The Impact of Axon Density on Corneal Epithelial Cell Proliferation Varies With Age and Parity
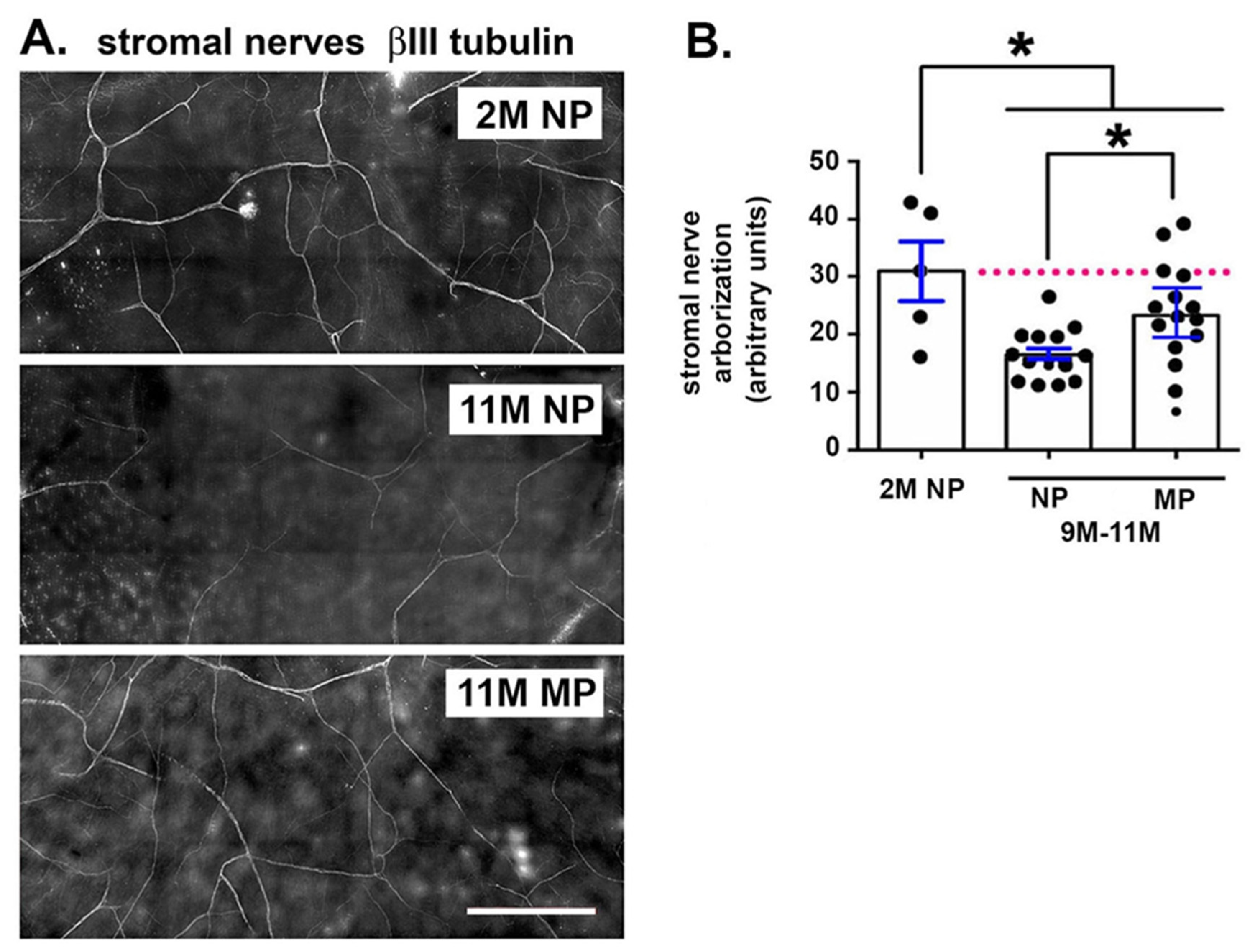
2.4. Stromal Nerve Arborization Is Reduced With Aging in NP and MP Mice
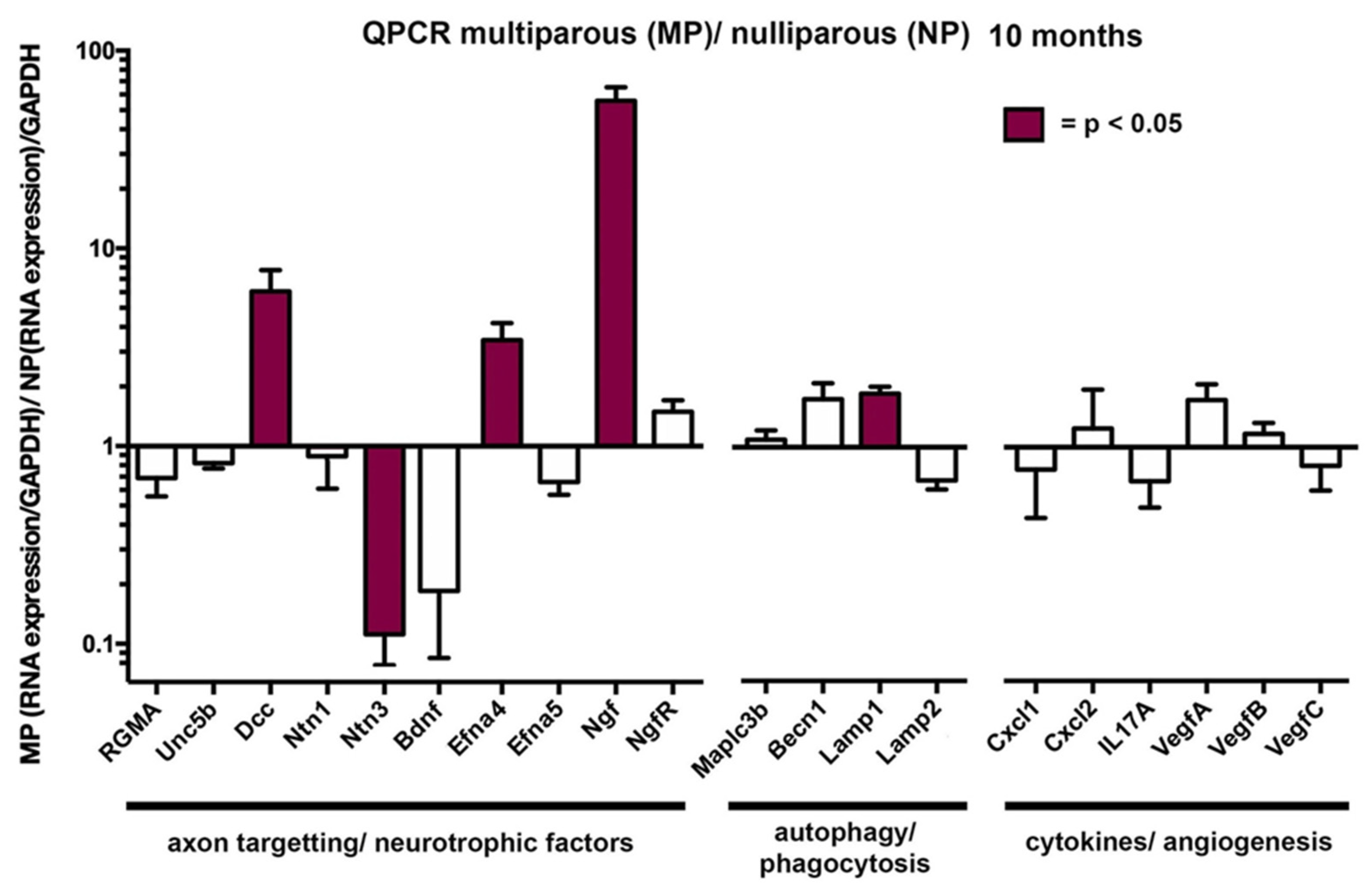
2.5. The Attenuation of the Age-Related Loss of Axon Density Seen in Aged MP Mice Is Associated With Elevated Levels of Mrnas for Several Neurotrophic Factors
2.6. Age-Related Loss of Corneal Sensitivity Is Attenuated in 9M MP Mice
2.7. Longer Parallel Intraepithelial Corneal Nerve Terminals (Picnts) Accompany the Enhanced Corneal Sensitivity at 9M in MP Mice
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ICNs | intraepithelial corneal nerves |
| ICBNs | intraepithelial corneal basal nerves |
| ICNTs | intraepithelial corneal nerve terminals |
| NP | nulliparous (virgin female) |
| MP | multiparous (breeding female) |
| PRL | prolactin |
| NGF | nerve growth factor |
| DMSO | dimethyl sulfoxide |
| PBS | phosphate buffered saline |
| M | months |
| DAPI | 4,6-diamidino-2- phenylindole |
| qPCR | quantitative polymerase chain reaction |
References
- Galea, L.A.; Wainwright, S.R.; Roes, M.M.; Duarte-Guterman, P.; Chow, C.; Hamson, D.K. Sex, hormones and neurogenesis in the hippocampus: Hormonal modulation of neurogenesis and potential functional implications. J. Neuroendocrinol. 2013, 25, 1039–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridges, R.S. Long-term alterations in neural and endocrine processes induced by motherhood in mammals. Horm. Behav. 2016, 77, 193–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawluski, J.L.; Charlier, T.D.; Lieblich, S.E.; Hammond, G.L.; Galea, L.A. Reproductive experience alters corticosterone and CBG levels in the rat dam. Physiol. Behav. 2009, 96, 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritzel, R.M.; Patel, A.R.; Spychala, M.; Verma, R.; Crapser, J.; Koellhoffer, E.C.; Schrecengost, A.; Jellison, E.R.; Zhu, L.; Venna, V.R.; et al. Multiparity improves outcomes after cerebral ischemia in female mice despite features of increased metabovascular risk. Proc. Natl. Acad. Sci. USA 2017, 114, E5673–E5682. [Google Scholar] [CrossRef] [Green Version]
- Costanza, M.; Binart, N.; Steinman, L.; Pedotti, R. Prolactin: A versatile regulator of inflammation and autoimmune pathology. Autoimmun. Rev. 2015, 14, 223–230. [Google Scholar] [CrossRef]
- Carvalho-Freitas, M.I.; Anselmo-Franci, J.A.; Teodorov, E.; Nasello, A.G.; Palermo-Neto, J.; Felicio, L.F. Reproductive experience modifies dopaminergic function, serum levels of prolactin, and macrophage activity in female rats. Life Sci. 2007, 81, 128–136. [Google Scholar] [CrossRef]
- Purves-Tyson, T.D.; Keast, J.R. Rapid actions of estradiol on cyclic amp response-element binding protein phosphorylation in dorsal root ganglion neurons. Neuroscience 2004, 129, 629–637. [Google Scholar] [CrossRef]
- Frank, P.; Barrientos, G.; Tirado-Gonzalez, I.; Cohen, M.; Moschansky, P.; Peters, E.M.; Klapp, B.F.; Rose, M.; Tometten, M.; Blois, S.M. Balanced levels of nerve growth factor are required for normal pregnancy progression. Reproduction 2014, 148, 179–189. [Google Scholar] [CrossRef]
- Wong, A.W.; Yeung, J.K.P.; Payne, S.C.; Keast, J.R.; Osborne, P.B. Neurite outgrowth in normal and injured primary sensory neurons reveals different regulation by nerve growth factor (NGF) and artemin. Mol. Cell Neurosci. 2015, 65, 125–134. [Google Scholar] [CrossRef]
- Schaumberg, D.A.; Sullivan, D.A.; Buring, J.E.; Dana, M.R. Prevalence of dry eye syndrome among US women. Am. J. Ophthalmol. 2003, 136, 318–326. [Google Scholar] [CrossRef]
- Schaumberg, D.A.; Dana, R.; Buring, J.E.; Sullivan, D.A. Prevalence of dry eye disease among US men: Estimates from the Physicians' Health Studies. Arch. Ophthalmol. 2009, 127, 763–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, S.E.; Klein, R.; Klein, B.E. Incidence of dry eye in an older population. Arch. Ophthalmol. 2004, 122, 369–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrand, K.F.; Fridman, M.; Stillman, I.O.; Schaumberg, D.A. Prevalence of Diagnosed Dry Eye Disease in the United States Among Adults Aged 18 Years and Older. Am. J. Ophthalmol. 2017, 182, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matossian, C.; McDonald, M.; Donaldson, K.E.; Nichols, K.K.; MacIver, S.; Gupta, P.K. Dry Eye Disease: Consideration for Women's Health. J. Womens Health (Larchmt) 2019, 28, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Pflugfelder, S.C.; de Paiva, C.S. The Pathophysiology of Dry Eye Disease: What We Know and Future Directions for Research. Ophthalmology 2017, 124, S4–S13. [Google Scholar] [CrossRef]
- Stepp, M.A.; Pal-Ghosh, S.; Tadvalkar, G.; Williams, A.; Pflugfelder, S.C.; de Paiva, C.S. Reduced intraepithelial corneal nerve density and sensitivity accompany desiccating stress and aging in C57BL/6 mice. Exp. Eye Res. 2018, 169, 91–98. [Google Scholar] [CrossRef]
- Stepp, M.A.; Pal-Ghosh, S.; Downie, L.E.; Zhang, A.C.; Chinnery, H.R.; Machet, J.; Di Girolamo, N. Corneal Epithelial "Neuromas": A Case of Mistaken Identity? Cornea 2020. [Google Scholar] [CrossRef]
- Barrat, F.; Lesourd, B.; Boulouis, H.J.; Thibault, D.; Vincent-Naulleau, S.; Gjata, B.; Louise, A.; Neway, T.; Pilet, C. Sex and parity modulate cytokine production during murine ageing. Clin. Exp. Immunol. 1997, 109, 562–568. [Google Scholar] [CrossRef]
- Urzua, U.; Chacon, C.; Lizama, L.; Sarmiento, S.; Villalobos, P.; Kroxato, B.; Marcelain, K.; Gonzalez, M.J. Parity History Determines a Systemic Inflammatory Response to Spread of Ovarian Cancer in Naturally Aged Mice. Aging Dis. 2017, 8, 546–557. [Google Scholar] [CrossRef] [Green Version]
- Deems, N.P.; Leuner, B. Pregnancy, postpartum and parity: Resilience and vulnerability in brain health and disease. Front. Neuroendocrinol. 2020, 57, 100820. [Google Scholar] [CrossRef]
- Stepp, M.A.; Pal-Ghosh, S.; Tadvalkar, G.; Li, L.; Brooks, S.R.; Morasso, M.I. Molecular basis of Mitomycin C enhanced corneal sensory nerve repair after debridement wounding. Sci. Rep. 2018, 8, 16960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.; Levi-Montalcini, R. Purification and properties of a nerve growth-promoting factor isolated from mouse sarcoma 180. Cancer Res. 1957, 17, 15–20. [Google Scholar] [PubMed]
- Levi-Montalcini, R. The nerve growth factor thirty-five years later. In Vitro Cell Dev. Biol. 1987, 23, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Lambiase, A.; Manni, L.; Rama, P.; Bonini, S. Clinical application of nerve growth factor on human corneal ulcer. Arch. Ital. Biol. 2003, 141, 141–148. [Google Scholar] [PubMed]
- Lambiase, A.; Sacchetti, M.; Bonini, S. Nerve growth factor therapy for corneal disease. Curr. Opin. Ophthalmol. 2012, 23, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Marfurt, C.F. Corneal Nerves: Anatomy; Elsevier: New York, NY, USA, 2010; Volume set 1. [Google Scholar]
- Sacchetti, M.; Lambiase, A.; Schmidl, D.; Schmetterer, L.; Ferrari, M.; Mantelli, F.; Allegretti, M.; Garhoefer, G. Effect of recombinant human nerve growth factor eye drops in patients with dry eye: A phase IIa, open label, multiple-dose study. Br. J. Ophthalmol. 2020, 104, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Pflugfelder, S.C.; Massaro-Giordano, M.; Perez, V.L.; Hamrah, P.; Deng, S.X.; Espandar, L.; Foster, C.S.; Affeldt, J.; Seedor, J.A.; Afshari, N.A.; et al. Topical Recombinant Human Nerve Growth Factor (Cenegermin) for Neurotrophic Keratopathy: A Multicenter Randomized Vehicle-Controlled Pivotal Trial. Ophthalmology 2020, 127, 14–26. [Google Scholar] [CrossRef] [Green Version]
- Schulte-Herbruggen, O.; Litzke, J.; Hornych, K.; Zingler, C.; Hoppner, J.; Virchow, J.C.; Hellweg, R.; Lommatzsch, M. Maternal nerve growth factor serum levels in the perinatal period. J. Reprod. Immunol. 2007, 74, 170–173. [Google Scholar] [CrossRef]
- Schulz, K.H.; Gold, S.M.; Witte, J.; Bartsch, K.; Lang, U.E.; Hellweg, R.; Reer, R.; Braumann, K.M.; Heesen, C. Impact of aerobic training on immune-endocrine parameters, neurotrophic factors, quality of life and coordinative function in multiple sclerosis. J. Neurol. Sci. 2004, 225, 11–18. [Google Scholar] [CrossRef]
- Jang, Y.; Lee, B.; Kim, E.K.; Shim, W.S.; Yang, Y.D.; Kim, S.M. Involuntary swimming exercise in pregnant rats disturbs ERK1/2 signaling in embryonic neurons through increased cortisol in the amniotic fluid. Biochem. Biophys. Res. Commun. 2018, 495, 1208–1213. [Google Scholar] [CrossRef]
- Mazzoccoli, G. Circadian rhythmicity of lymphocyte subpopulation and relationship with neuro-endocrine system. J. Biol. Regul. Homeost. Agents 2010, 24, 341–350. [Google Scholar]
- Cermakian, N.; Lange, T.; Golombek, D.; Sarkar, D.; Nakao, A.; Shibata, S.; Mazzoccoli, G. Crosstalk between the circadian clock circuitry and the immune system. Chronobiol. Int. 2013, 30, 870–888. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gonzalez, O.; Bech, F.; Gallar, J.; Merayo-Lloves, J.; Belmonte, C. Functional Properties of Sensory Nerve Terminals of the Mouse Cornea. Investig. Ophthalmol. Vis. Sci. 2017, 58, 404–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O'Donnell, M.; Chance, R.K.; Bashaw, G.J. Axon growth and guidance: Receptor regulation and signal transduction. Annu. Rev. Neurosci. 2009, 32, 383–412. [Google Scholar] [CrossRef] [Green Version]
- Lai Wing Sun, K.; Correia, J.P.; Kennedy, T.E. Netrins: Versatile extracellular cues with diverse functions. Development 2011, 138, 2153–2169. [Google Scholar]
- Eberhart, J.; Swartz, M.E.; Koblar, S.A.; Pasquale, E.B.; Krull, C.E. EphA4 constitutes a population-specific guidance cue for motor neurons. Dev. Biol. 2002, 247, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Kania, A.; Klein, R. Mechanisms of ephrin-Eph signalling in development, physiology and disease. Nat. Rev. Mol. Cell Biol. 2016, 17, 240–256. [Google Scholar] [CrossRef]
- Croteau, L.P.; Kao, T.J.; Kania, A. Ephrin-A5 potentiates netrin-1 axon guidance by enhancing Neogenin availability. Sci. Rep. 2019, 9, 12009. [Google Scholar] [CrossRef] [Green Version]
- Pal-Ghosh, S.; Tadvalkar, G.; Karpinski, B.A.; Stepp, M.A. Diurnal control of sensory axon growth and shedding in the mouse cornea. Investig. Ophthalmol. Vis. Sci. undergoing minor revisions.
- Bahougne, T.; Kretz, M.; Angelopoulou, E.; Jeandidier, N.; Simonneaux, V. Impact of Circadian Disruption on Female Mice Reproductive Function. Endocrinology 2020, 161. [Google Scholar] [CrossRef]
- Takasu, N.N.; Nakamura, T.J.; Tokuda, I.T.; Todo, T.; Block, G.D.; Nakamura, W. Recovery from Age-Related Infertility under Environmental Light-Dark Cycles Adjusted to the Intrinsic Circadian Period. Cell Rep. 2015, 12, 1407–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blount, J.D.; Vitikainen, E.I.; Stott, I.; Cant, M.A. Oxidative shielding and the cost of reproduction. Biol. Rev. Camb. Philos. Soc. 2016, 91, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Costantini, D. Understanding diversity in oxidative status and oxidative stress: The opportunities and challenges ahead. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naviaux, R.K. Oxidative shielding or oxidative stress? J. Pharmacol. Exp. Ther. 2012, 342, 608–618. [Google Scholar] [CrossRef] [Green Version]
- Naviaux, R.K. Metabolic features of the cell danger response. Mitochondrion 2014, 16, 7–17. [Google Scholar] [CrossRef]
- Alonso Alvarez, C. The oxidative cost of reproduction. Bioscience 2017, 283, 258–270. [Google Scholar] [CrossRef]
- Harris, B.N. Stress hypothesis overload: 131 hypotheses exploring the role of stress in tradeoffs, transitions, and health. Gen. Comp. Endocrinol. 2020, 288, 113355. [Google Scholar] [CrossRef]
- Stepp, M.A.; Pal-Ghosh, S.; Tadvalkar, G.; Williams, A.R.; Pflugfelder, S.C.; de Paiva, C.S. Reduced Corneal Innervation in the CD25 Null Model of Sjogren Syndrome. Int. J. Mol. Sci 2018, 19, 3821. [Google Scholar] [CrossRef] [Green Version]
- Labbe, A.; Alalwani, H.; Van Went, C.; Brasnu, E.; Georgescu, D.; Baudouin, C. The relationship between subbasal nerve morphology and corneal sensation in ocular surface disease. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4926–4931. [Google Scholar] [CrossRef]
- Bron, A.J.; de Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef]
- Niederkorn, J.Y.; Stern, M.E.; Pflugfelder, S.C.; De Paiva, C.S.; Corrales, R.M.; Gao, J.; Siemasko, K. Desiccating stress induces T cell-mediated Sjogren's Syndrome-like lacrimal keratoconjunctivitis. J. Immunol. 2006, 176, 3950–3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, F.; Xiao, Y.; Barbosa, F.L.; de Souza, R.G.; Hernandez, H.; Yu, Z.; Pflugfelder, S.C.; de Paiva, C.S. Age-associated antigen-presenting cell alterations promote dry-eye inducing Th1 cells. Mucosal. Immunol. 2019, 12, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Schein, O.D.; Hochberg, M.C.; Munoz, B.; Tielsch, J.M.; Bandeen-Roche, K.; Provost, T.; Anhalt, G.J.; West, S. Dry eye and dry mouth in the elderly: A population-based assessment. Arch. Intern. Med. 1999, 159, 1359–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, M.A. Dry eye in the elderly. Drugs Aging 2001, 18, 101–107. [Google Scholar] [CrossRef]
- Moss, S.E.; Klein, R.; Klein, B.E. Long-term incidence of dry eye in an older population. Optom. Vis. Sci. 2008, 85, 668–674. [Google Scholar] [CrossRef] [Green Version]
- Tellefsen, S.; Morthen, M.K.; Richards, S.M.; Lieberman, S.M.; Rahimi Darabad, R.; Kam, W.R.; Sullivan, D.A. Sex Effects on Gene Expression in Lacrimal Glands of Mouse Models of Sjogren Syndrome. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5599–5614. [Google Scholar] [CrossRef] [Green Version]
- Rahimi Darabad, R.; Suzuki, T.; Richards, S.M.; Jakobiec, F.A.; Zakka, F.R.; Barabino, S.; Sullivan, D.A. Does estrogen deficiency cause lacrimal gland inflammation and aqueous-deficient dry eye in mice? Exp. Eye Res. 2014, 127, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, D.A.; Belanger, A.; Cermak, J.M.; Berube, R.; Papas, A.S.; Sullivan, R.M.; Yamagami, H.; Dana, M.R.; Labrie, F. Are women with Sjogren's syndrome androgen-deficient? J. Rheumatol. 2003, 30, 2413–2419. [Google Scholar]
- Piccinni, M.P.; Lombardelli, L.; Logiodice, F.; Kullolli, O.; Parronchi, P.; Romagnani, S. How pregnancy can affect autoimmune diseases progression? Clin. Mol. Allergy 2016, 14, 11. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stepp, M.A.; Pal-Ghosh, S.; Tadvalkar, G.; de Paiva, C.S. Parity Attenuates Intraepithelial Corneal Sensory Nerve Loss in Female Mice. Int. J. Mol. Sci. 2020, 21, 5172. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145172
Stepp MA, Pal-Ghosh S, Tadvalkar G, de Paiva CS. Parity Attenuates Intraepithelial Corneal Sensory Nerve Loss in Female Mice. International Journal of Molecular Sciences. 2020; 21(14):5172. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145172
Chicago/Turabian StyleStepp, Mary Ann, Sonali Pal-Ghosh, Gauri Tadvalkar, and Cintia S. de Paiva. 2020. "Parity Attenuates Intraepithelial Corneal Sensory Nerve Loss in Female Mice" International Journal of Molecular Sciences 21, no. 14: 5172. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145172