Patterns of Expansion and Expression Divergence of the Polygalacturonase Gene Family in Brassica oleracea



Abstract
:1. Introduction
2. Results
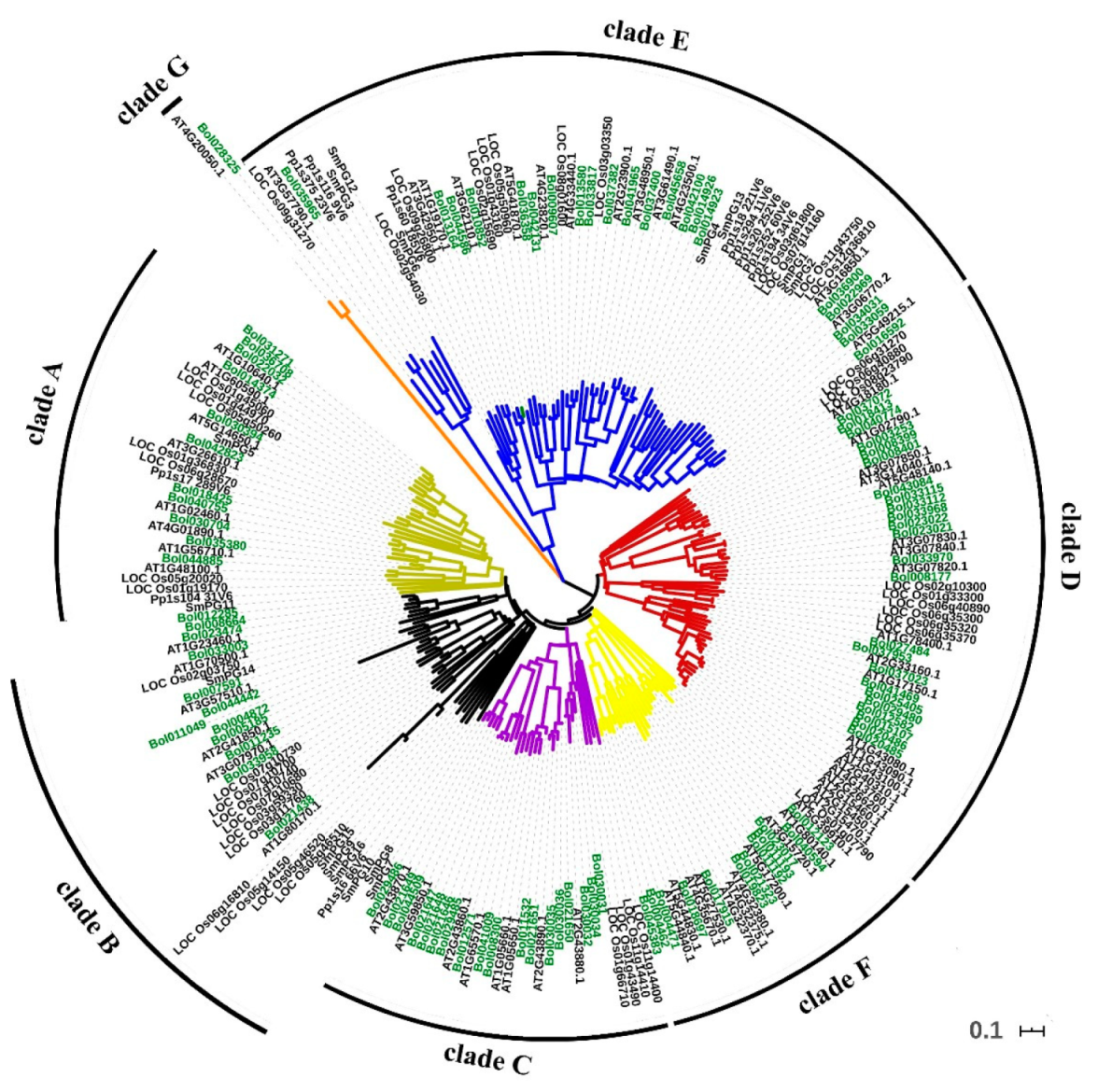
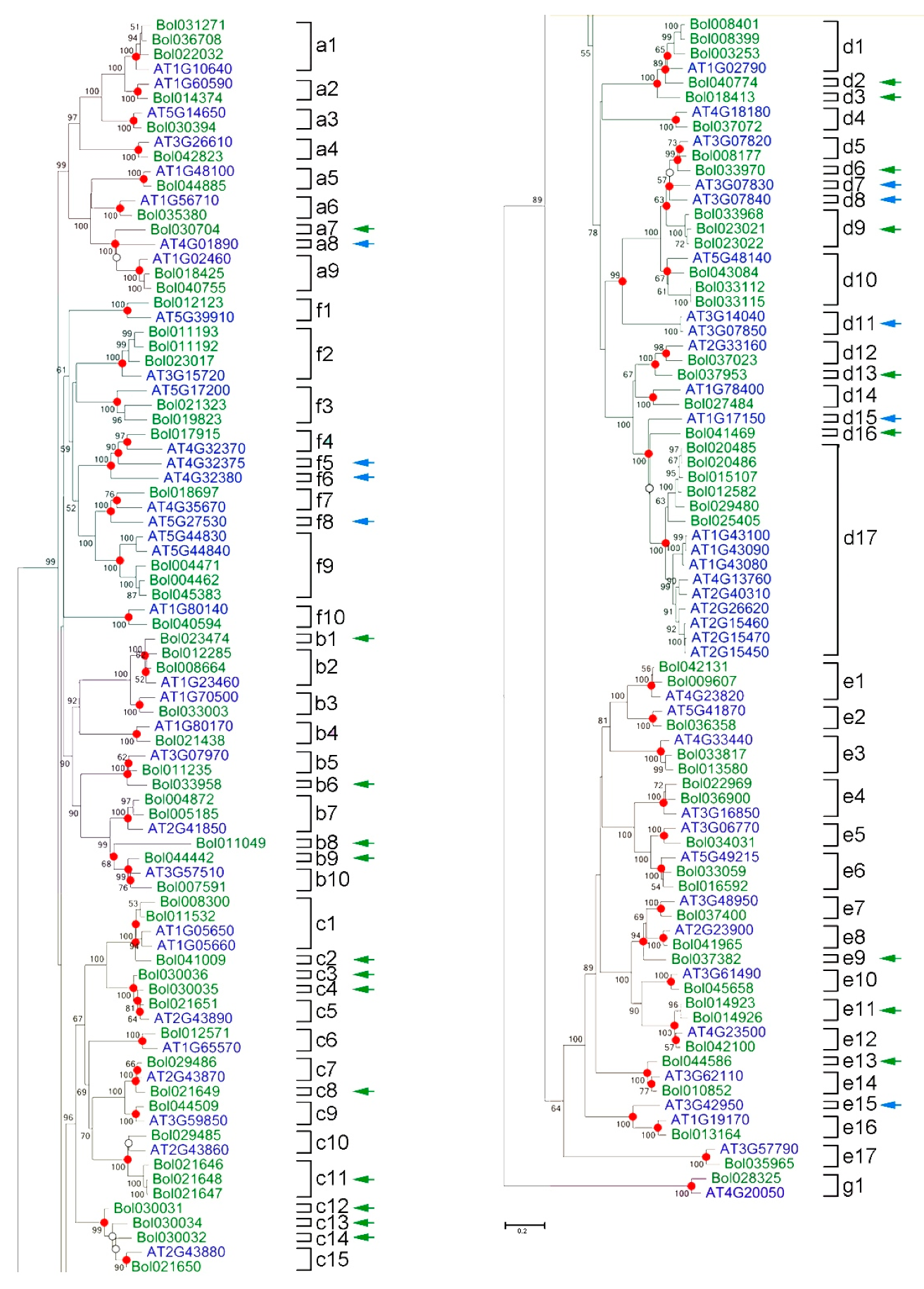
2.1. The Polygalacturonase Gene Family in B. oleracea Consists of 99 Members
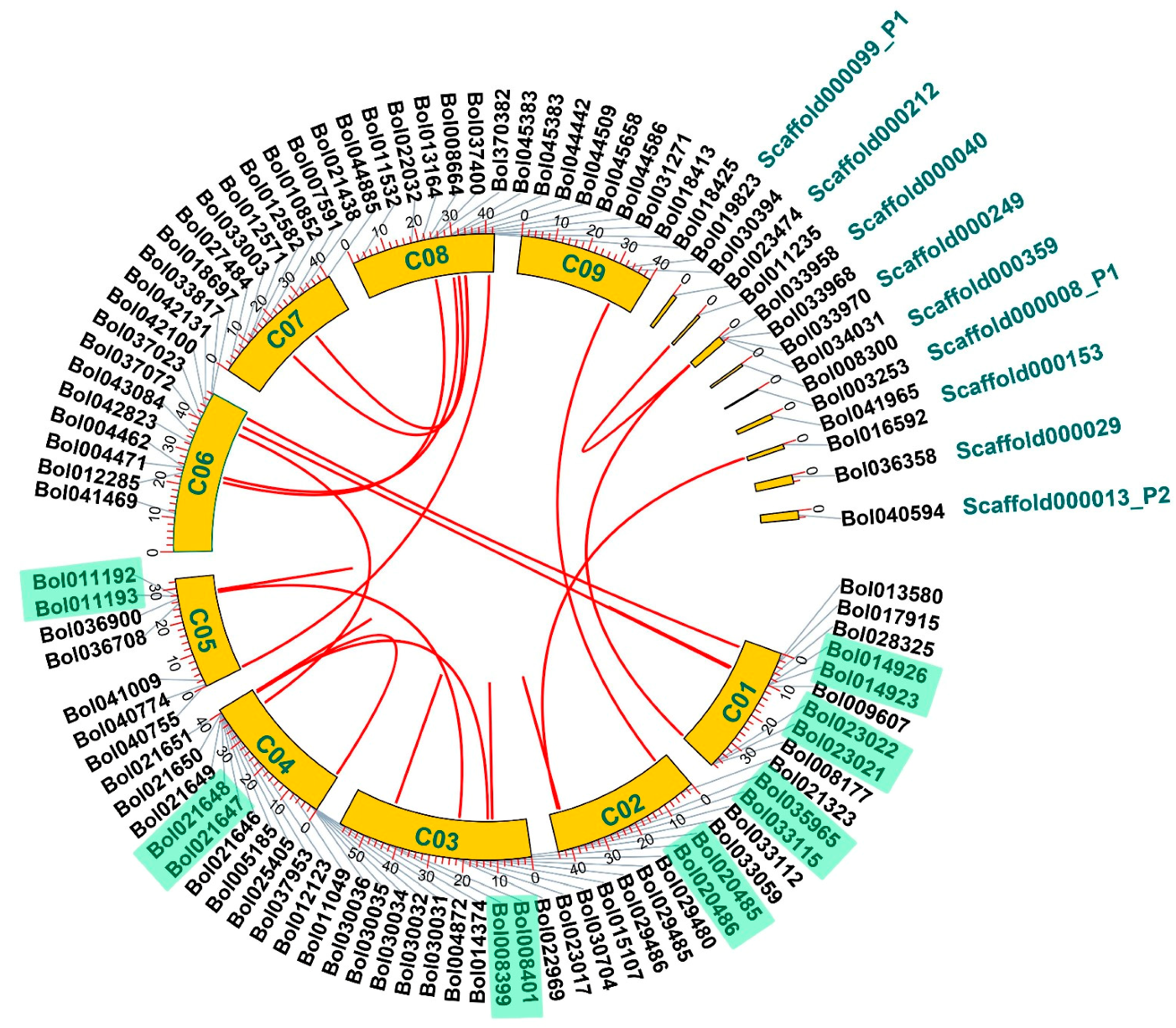
2.2. Expansion Mechanisms Accounting for the PG Family in B. oleracea
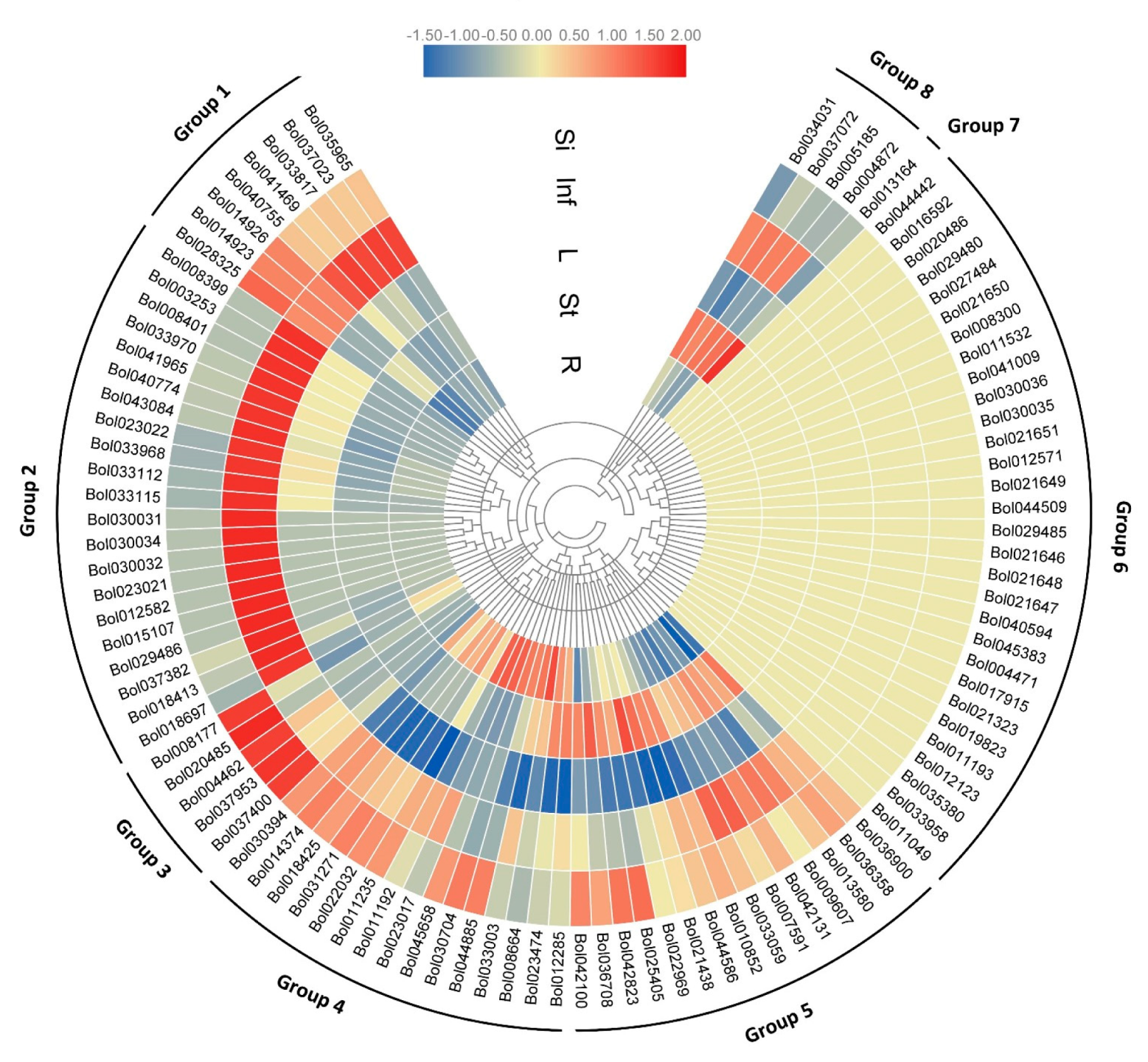
2.3. Expression Divergence Implies the Involvement of BoPGs in Different Tissues
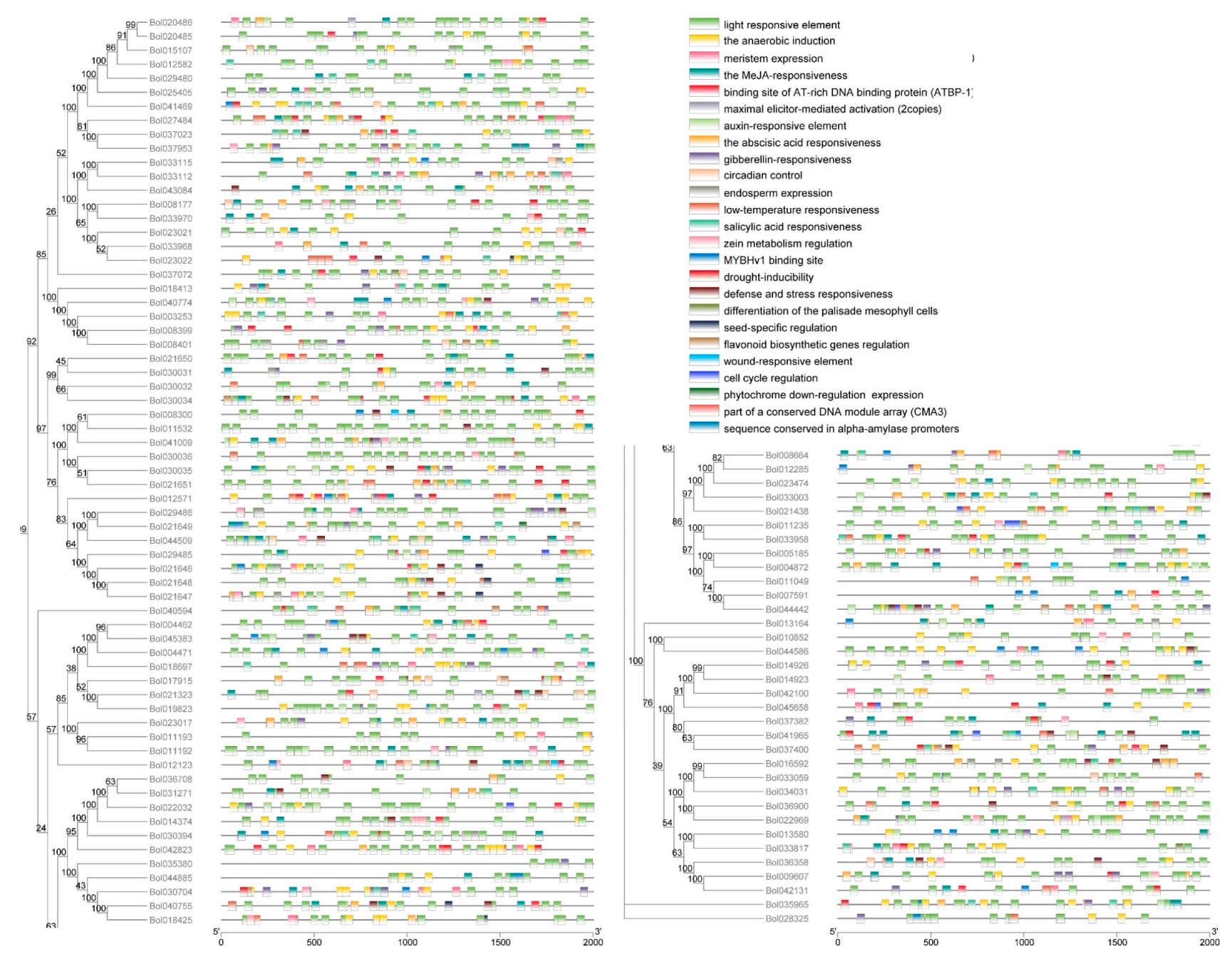
2.4. Cis-Elements Analysis of the BoPG Promoters
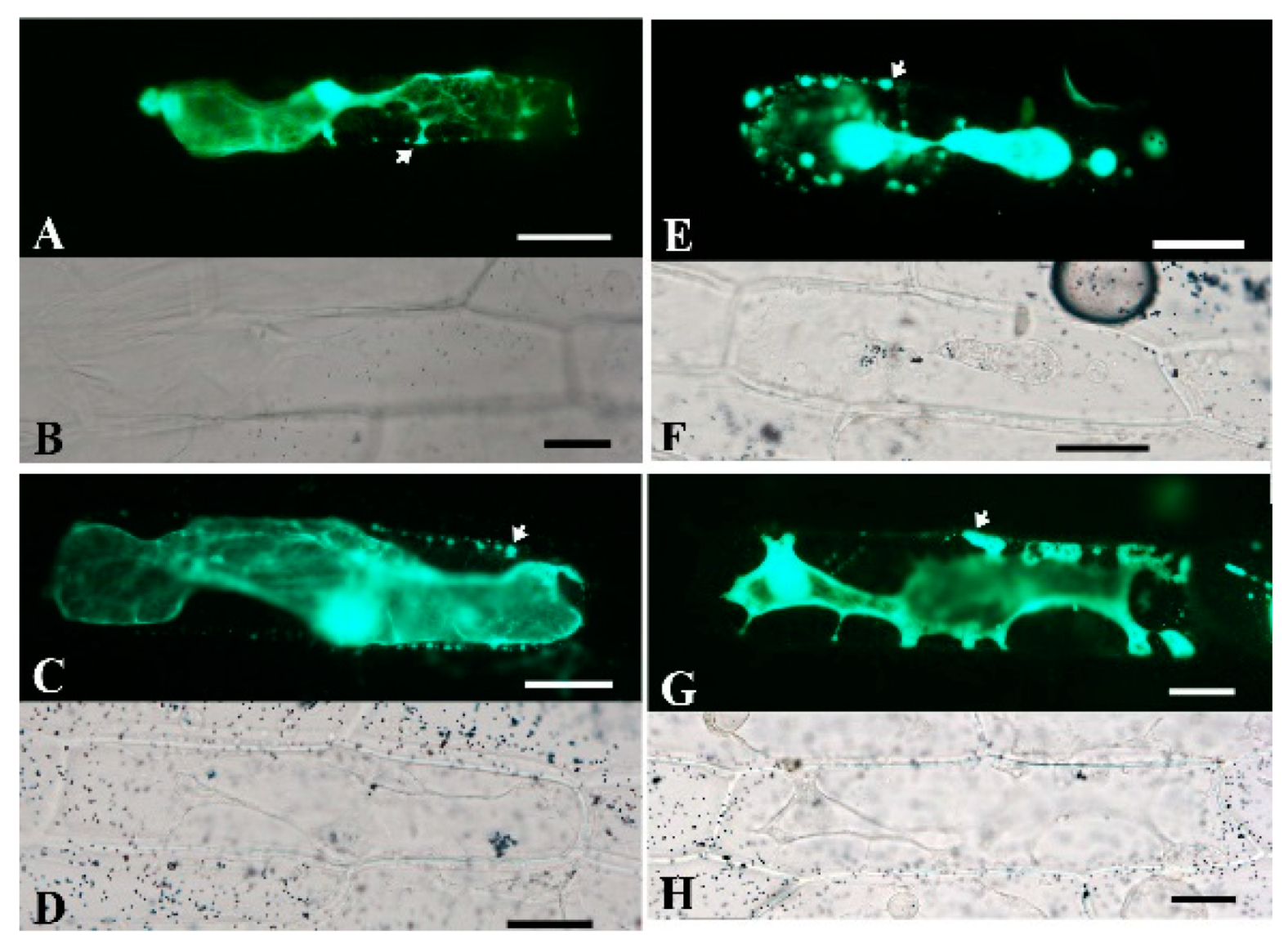
2.5. Subcellular Localization of BoPG Proteins
3. Discussion
4. Materials and Methods
4.1. Identification and Sequence Analysis
4.2. Phylogenetic and Molecular Evolution
4.3. Gene Expression Analysis
4.4. Subcellular Localization
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Demura, T.; Tashiro, G.; Horiguchi, G.; Kishimoto, N.; Kubo, M.; Matsuoka, N.; Minami, A.; Nagata-Hiwatashi, M.; Nakamura, K.; Okamura, Y.; et al. Visualization by comprehensive microarray analysis of gene expression programs during transdifferentiation of mesophyll cells into xylem cells. Proc. Natl. Acad. Sci. USA 2002, 99, 15794–15799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadfield, K.A.; Bennett, A.B. Polygalacturonases: Many genes in search of a function. Plant Physiol. 1998, 117, 337–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sander, L.; Child, R.; Ulvskov, P.; Albrechtsen, M.; Borkhardt, B. Analysis of a dehiscence zone endo-polygalacturonase in oilseed rape (Brassica napus) and Arabidopsis thaliana: Evidence for roles in cell separation in dehiscence and abscission zones, and in stylar tissues during pollen tube growth. Plant Mol. Biol. 2001, 46, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Sitrit, Y.; Hadfield, K.A.; Bennett, A.B.; Bradford, K.J.; Downie, A.B. Expression of a polygalacturonase associated with tomato seed germination. Plant Physiol. 1999, 121, 419–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, R.L.; Bennett, A.B. Role of cell-wall hydrolases in fruit ripening. Annu. Rev. Plant Physiol. Mol. Biol. 1991, 42, 675–703. [Google Scholar] [CrossRef]
- Kim, J.; Patterson, S.E. Expression divergence and functional redundancy of polygalacturonases in floral organ abscission. Plant Signal. Behav. 2006, 1, 281–283. [Google Scholar] [CrossRef] [Green Version]
- Roongsattham, P.; Morcillo, F.; Jantasuriyarat, C.; Pizot, M.; Moussu, S.; Jayaweera, D.; Collin, M.; Gonzalez-Carranza, Z.H.; Amblard, P.; Tregear, J.W.; et al. Temporal and spatial expression of polygalacturonase gene family members reveals divergent regulation during fleshy fruit ripening and abscission in the monocot species oil palm. BMC Plant Biol. 2012, 12. [Google Scholar] [CrossRef] [Green Version]
- Ke, X.; Wang, H.; Li, Y.; Zhu, B.; Zang, Y.; He, Y.; Cao, J.; Zhu, Z.; Yu, Y. Genome-wide identification and analysis of polygalacturonase genes in Solanum lycopersicum. Int. J. Mol. Sci. 2018, 19, 2290. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Cao, J.; Zhang, A.; Ye, Y.; Zhang, Y.; Liu, T. The polygalacturonase gene BcMF2 from Brassica campestris is associated with intine development. J. Exp. Bot. 2009, 60, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Ye, Y.; Zhang, Y.; Zhang, A.; Liu, T.; Cao, J. BcMF9 a novel polygalacturonase gene, is required for both Brassica campestris intine and exine formation. Ann. Bot. 2009, 104, 1339–1351. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.; Qiu, L.; Huang, L.; Yu, X.; Lu, G.; Cao, J. Isolation and characterization of an anther-specific polygalacturonase gene, BcMF16, in Brassica campestris ssp. chinensis. Plant Mol. Biol. Rep. 2012, 30, 330–338. [Google Scholar] [CrossRef]
- Zhang, A.; Chen, Q.; Huang, L.; Qiu, L.; Cao, J. Cloning and expression of an anther-abundant polygalacturonase gene BcMF17 from Brassica campestris ssp chinensis. Plant Mol. Biol. Rep. 2011, 29, 943–951. [Google Scholar] [CrossRef]
- Yu, Y.; Lv, M.; Liang, Y.; Xiong, X.; Cao, J. Molecular cloning and characterization of a novel polygalacturonase gene, BcMF24, involved in pollen development of Brassica campestris ssp chinensis. Plant Mol. Biol. Rep. 2014, 32, 476–486. [Google Scholar] [CrossRef]
- Lyu, M.; Liang, Y.; Yu, Y.; Ma, Z.; Song, L.; Yue, X.; Cao, J. Identification and expression analysis of BoMF25, a novel polygalacturonase gene involved in pollen development of Brassica oleracea. Plant Reprod. 2015, 28, 121–132. [Google Scholar] [CrossRef]
- He, Y.; Karre, S.; Johal, G.S.; Christensen, S.A.; Balint-Kurti, P. Amaize polygalacturonase functions as a suppressor of programmed cell death in plants. BMC Plant Biol. 2019, 19, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torki, M.; Mandaron, P.; Mache, R.; Falconet, D. Characterization of a ubiquitous expressed gene family encoding polygalacturonase in Arabidopsis thaliana. Gene 2000, 242, 427–436. [Google Scholar] [CrossRef]
- Park, K.C.; Kwon, S.J.; Kim, P.H.; Bureau, T.; Kim, N.S. Gene structure dynamics and divergence of the polygalacturonase gene family of plants and fungus. Genome 2008, 51, 30–40. [Google Scholar] [CrossRef]
- Park, K.C.; Kwon, S.J.; Kim, N.S. Intron loss mediated structural dynamics and functional differentiation of the polygalacturonase gene family in land plants. Genes Genom. 2010, 32, 570–577. [Google Scholar] [CrossRef]
- Yang, Z.L.; Liu, H.J.; Wang, X.R.; Zeng, Q.Y. Molecular evolution and expression divergence of the Populus polygalacturonase supergene family shed light on the evolution of increasingly complex organs in plants. New Phytol. 2013, 197, 1353–1365. [Google Scholar] [CrossRef]
- Yu, Y.; Liang, Y.; Lv, M.; Wu, J.; Lu, G.; Cao, J. Genome-wide identification and characterization of polygalacturonase genes in Cucumis sativus and Citrullus lanatus. Plant Physiol. Biochem. 2014, 74, 263–275. [Google Scholar] [CrossRef]
- Liang, Y.; Yu, Y.; Shen, X.; Dong, H.; Lyu, M.; Xu, L.; Ma, Z.; Liu, T.; Cao, J. Dissecting the complex molecular evolution and expression of polygalacturonase gene family in Brassica rapa ssp chinensis. Plant Mol. Biol. 2015, 89, 629–646. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shao, H.; Fan, S.; Ma, J.; Zhang, D.; Han, M. Identification and phylogenetic analysis of the polygalacturonase gene family in Apple. Hortic. Plant J. 2016, 2, 241–252. [Google Scholar] [CrossRef]
- Thompson, P.; Patel, G. Compensatory regulation of two closely related hemoglobin loci in Chironomus tentans. Genetics 1972, 70, 275–290. [Google Scholar] [PubMed]
- Kassahn, K.S.; Dang, V.T.; Wilkins, S.J.; Perkins, A.C.; Ragan, M.A. Evolution of gene function and regulatory control after whole-genome duplication: Comparative analyses in vertebrates. Genome Res. 2009, 19, 1404–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, G.; Wolfe, K.H. Functional divergence of duplicated genes formed by polyploidy during Arabidopsis evolution. Plant Cell 2004, 16, 1679–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, J.M.; Cui, L.Y.; Wall, P.K.; Zhang, Q.; Zhang, X.H.; Leebens-Mack, J.; Ma, H.; Altman, N.; de Pamphilis, C.W. Expression pattern shifts following duplication indicative of subfunctionalization and neofunctionalization in regulatory genes of Arabidopsis. Mol. Biol. Evol. 2006, 23, 469–478. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon-intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Lysak, M.A.; Koch, M.A.; Pecinka, A.; Schubert, I. Chromosome triplication found across the tribe Brassiceae. Genome Res. 2005, 15, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Town, C.D.; Cheung, F.; Maiti, R.; Crabtree, J.; Haas, B.J.; Wortman, J.R.; Hine, E.E.; Althoff, R.; Arbogast, T.S.; Tallon, L.J.; et al. Comparative genomics of Brassica oleracea and Arabidopsis thaliana reveal gene loss, fragmentation, and dispersal after polyploidy. PMC Plant Cell 2006, 18, 1348–1359. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Shiu, S.H.; Thoma, S.; Li, W.H.; Patterson, S.E. Patterns of expansion and expression divergence in the plant polygalacturonase gene family. Genome Biol. 2006, 7, R87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wray, G.A.; Hahn, M.W.; Abouheif, E.; Balhoff, J.P.; Pizer, M.; Rockman, M.V.; Romano, L.A. The evolution of transcriptional regulation in eukaryotes. Mol. Biol. Evol. 2003, 20, 1377–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, M.F.; Xu, T.; Chen, W.Z.; Li, T.L. Ultrastructural localization of polygalacturonase in ethylene-stimulated abscission of tomato pedicel explants. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellapenna, D.; Bennett, A.B. In vitro synthesis and processing of tomato fruit polygalacturonase. Plant Physiol. 1998, 86, 1057–1063. [Google Scholar] [CrossRef] [Green Version]
- Chaw, S.M.; Chang, C.C.; Chen, H.L.; Li, W.H. Dating the monocot-dicot divergence and the origin of core eudicots using whole chloroplast genomes. J. Mol. Evol. 2004, 58, 424–441. [Google Scholar]
- Li, W.H.; Yang, J.; Gu, X. Expression divergence between duplicate genes. Trends Genet. 2005, 21, 602–607. [Google Scholar] [CrossRef]
- Rexová-Benková, L. Evidence for the role of carboxyl groups in activity of endopolygalacturonase of Aspergillus niger. Chemical modification by carbodiimide reagent. Collect. Czech. Chem. Commun. 1990, 55, 1389–1395. [Google Scholar] [CrossRef]
- Rao, M.N.; Kembhavi, A.A.; Pant, A. Implication of tryptophan and histidine in the active site of endo-polygalacturonase from Aspergillus ustus: Elucidation of the reaction mechanism. Biochim. Biophys. Acta 1996, 1296, 167–173. [Google Scholar]
- Bussink, H.J.; Buxton, F.P.; Visser, J. Expression and sequence comparison of the Aspergillus niger and Aspergillus tubigensis genes encoding polygalacturonase II. Curr. Genet. 1991, 19, 467–474. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.P.; Zhao, M.; Ma, J.; Yu, J.; Huang, S. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef]
- Zou, C.; Lehti-Shiu, M.D.; Thibaud-Nissen, F.; Prakash, T.; Buell, C.R.; Shiu, S.H. Evolutionary and expression signatures of pseudogenes in Arabidopsis and Rice. Plant Physiol. 2009, 151, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.L.; Postlethwait, J. Preservation of duplicate genes by complementary, degenerative mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [PubMed]
- Lyu, M.; Yu, Y.; Jiang, J.; Song, L.; Liang, Y.; Ma, Z.; Xiong, X.; Cao, J. BcMF26a and BcMF26b are duplicated polygalacturonase genes with divergent expression patterns and functions in pollen development and pollen tube formation in Brassica campestris. PLoS ONE 2015, 10, e0131173. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, F.; Zhao, Y.; Shi, L.; Zhu, X. Cloning and expression analysis of polygalacturonase and pectin methylesterase genes during softening in apricot (Prunus armeniaca L.) fruit. Sci. Hortic. 2019, 256, 108607. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, Y.; Lin, H.; Lin, M.; Li, H.; Yuan, F.; Chen, Y.; Xiao, J. Effects of paper containing 1-MCP postharvest treatment on the disassembly of cell wall polysaccharides and softening in Younai plum fruit during storage. Food Chem. 2018, 264, 1–8. [Google Scholar] [CrossRef]
- Lara-Nunez, A.; Ventura-Gallegos, J.; Luisa, A.A.; Cruz-Ortega, R. Phytotoxicity of sicyos deppei during tomato germination and its effects on the role of ABA and cell wall enzymes. Bot. Sci. 2015, 93, 771–781. [Google Scholar] [CrossRef]
- Ogawa, M.; Kay, P.; Wilson, S.; Swain, S.M. Arabidopsis dehiscence zone Polygalacturonase1 (ADPG1), ADPG2, and QUARTET2 are polygalacturonases required for cell separation during reproductive development in Arabidopsis. Plant Cell 2009, 21, 216–233. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.Y.; Osborne, E.; Poindexter, P.D.; Somerville, C.R. Microspore separation in the quartet 3 mutants of Arabidopsis is impaired by a defect in a developmentally regulated polygalacturonase required for pollen mother cell wall degradation. Plant Physiol. 2003, 133, 1170–1180. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Wu, J.; Fang, L.; Wang, X. Syntenic gene analysis between Brassica rapa and other Brassicaceae species. Front. Plant Sci. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Zhao, X.; Liu, T.; Dong, H.; Cao, J. Developmental characteristics of floral organs and pollen of Chinese cabbage (Brassica campestris L. ssp chinensis). Plant. Syst. Evol. 2010, 286, 103–115. [Google Scholar] [CrossRef]
- Suzuki, T.; Higgins, P.J.; Crawford, D.R. Control selection for RNA quantitation. Biotechniques 2000, 29, 332–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Paralogous Gene Pairs | Ka | Ks | Ka/Ks | Categories | |
|---|---|---|---|---|---|
| Bol020486 | Bol020485 | 0.0021 | 0.0264 | 0.0808 | IV |
| Bol037023 | Bol037953 | 0.1202 | 0.5223 | 0.2301 | II |
| Bol033115 | Bol033112 | 0 | 0 | —— | I |
| Bol008177 | Bol033970 | 0.0547 | 0.267 | 0.2047 | II |
| Bol008399 | Bol008401 | 0.0221 | 0.0661 | 0.3347 | I |
| Bol029486 | Bol021649 | 0.0383 | 0.2782 | 0.1377 | IV |
| Bol021648 | Bol021647 | 0.0063 | 0.0072 | 0.8854 | I |
| Bol004462 | Bol045383 | 0.0316 | 0.1065 | 0.297 | IV |
| Bol021323 | Bol019823 | 0.1487 | 0.3771 | 0.3942 | I |
| Bol011193 | Bol011192 | 0.0504 | 0.1386 | 0.3636 | IV |
| Bol040755 | Bol018425 | 0.0359 | 0.2634 | 0.1361 | II |
| Bol008664 | Bol012285 | 0.0383 | 0.367 | 0.1044 | II |
| Bol011235 | Bol033958 | 0.0934 | 0.3071 | 0.3041 | IV |
| Bol005185 | Bol004872 | 0.0471 | 0.2669 | 0.1766 | I |
| Bol007591 | Bol044442 | 0.1061 | 0.323 | 0.3283 | IV |
| Bol010852 | Bol044586 | 0.0528 | 0.3528 | 0.1498 | I |
| Bol014926 | Bol014923 | 0.003 | 0.0152 | 0.201 | IV |
| Bol016592 | Bol033059 | 0.0526 | 0.3394 | 0.155 | IV |
| Bol036900 | Bol022969 | 0.0622 | 0.2988 | 0.2082 | II |
| Bol013580 | Bol033817 | 0.0427 | 0.3785 | 0.1129 | III |
| Bol009607 | Bol042131 | 0.0294 | 0.3163 | 0.0928 | II |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, M.; Iftikhar, J.; Guo, R.; Wu, B.; Cao, J. Patterns of Expansion and Expression Divergence of the Polygalacturonase Gene Family in Brassica oleracea. Int. J. Mol. Sci. 2020, 21, 5706. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165706
Lyu M, Iftikhar J, Guo R, Wu B, Cao J. Patterns of Expansion and Expression Divergence of the Polygalacturonase Gene Family in Brassica oleracea. International Journal of Molecular Sciences. 2020; 21(16):5706. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165706
Chicago/Turabian StyleLyu, Meiling, Junaid Iftikhar, Rongfang Guo, Binghua Wu, and Jiashu Cao. 2020. "Patterns of Expansion and Expression Divergence of the Polygalacturonase Gene Family in Brassica oleracea" International Journal of Molecular Sciences 21, no. 16: 5706. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165706