Complement C4 Gene Copy Number Variation Genotyping by High Resolution Melting PCR


Abstract
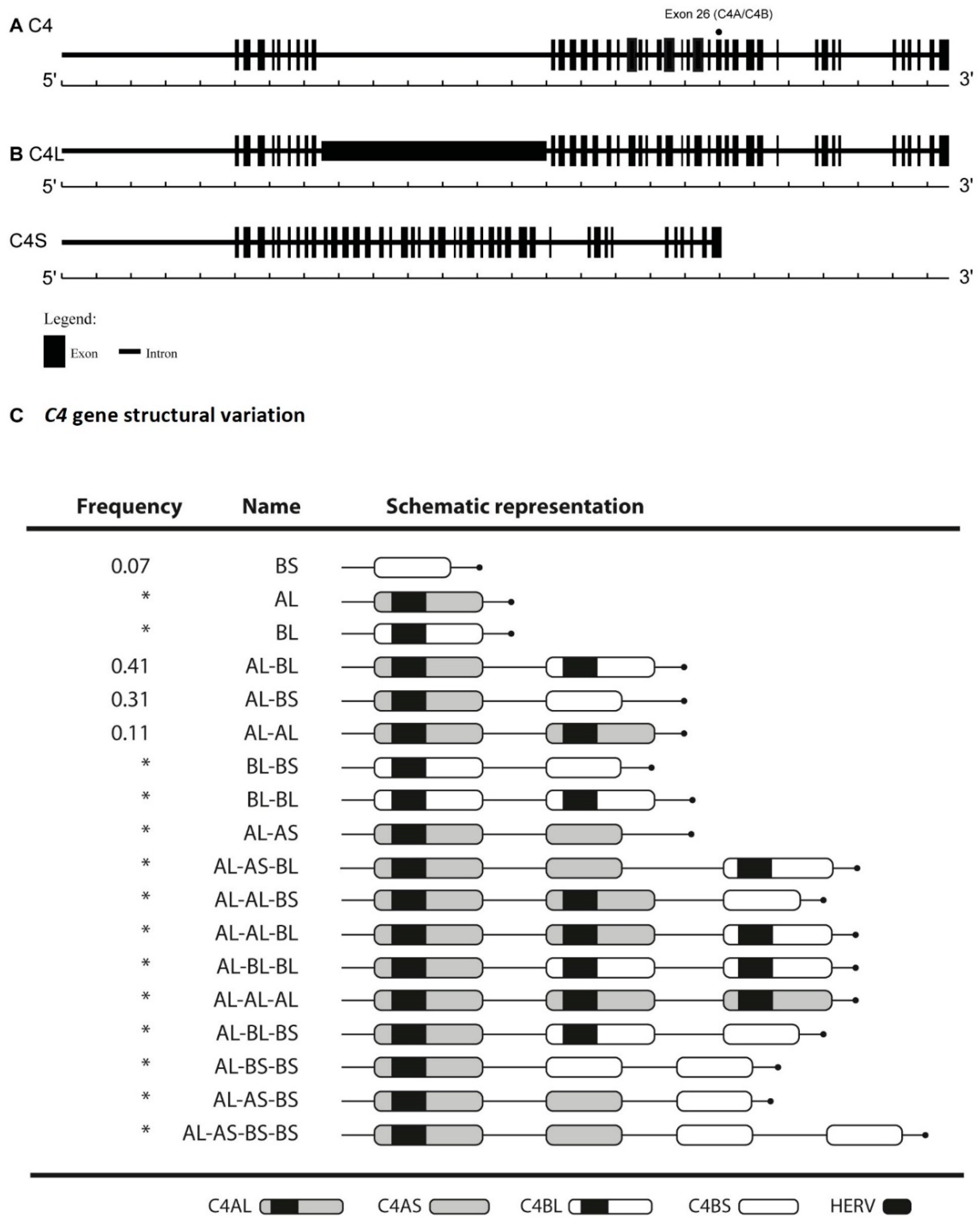
:1. Introduction
2. Results
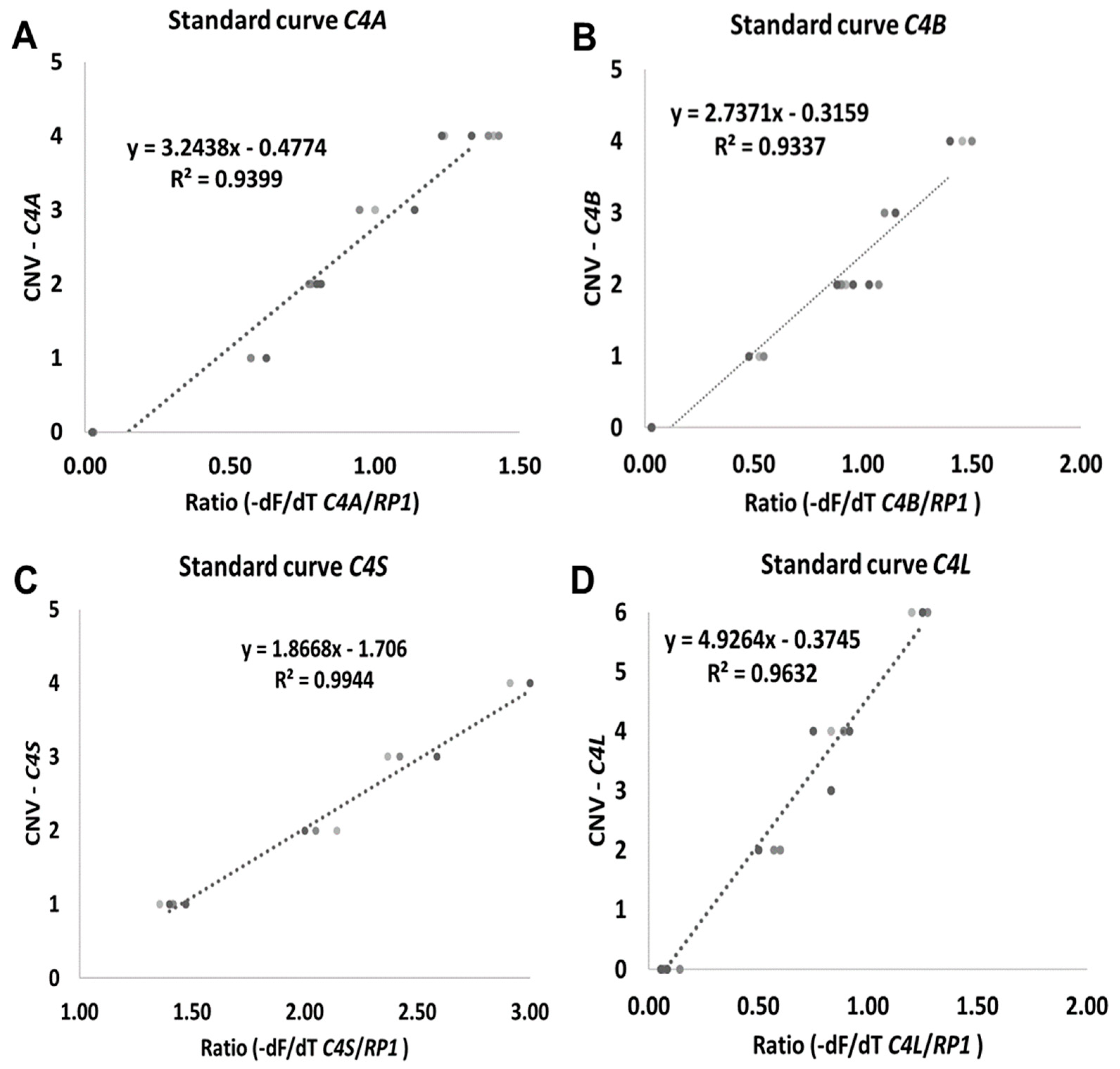
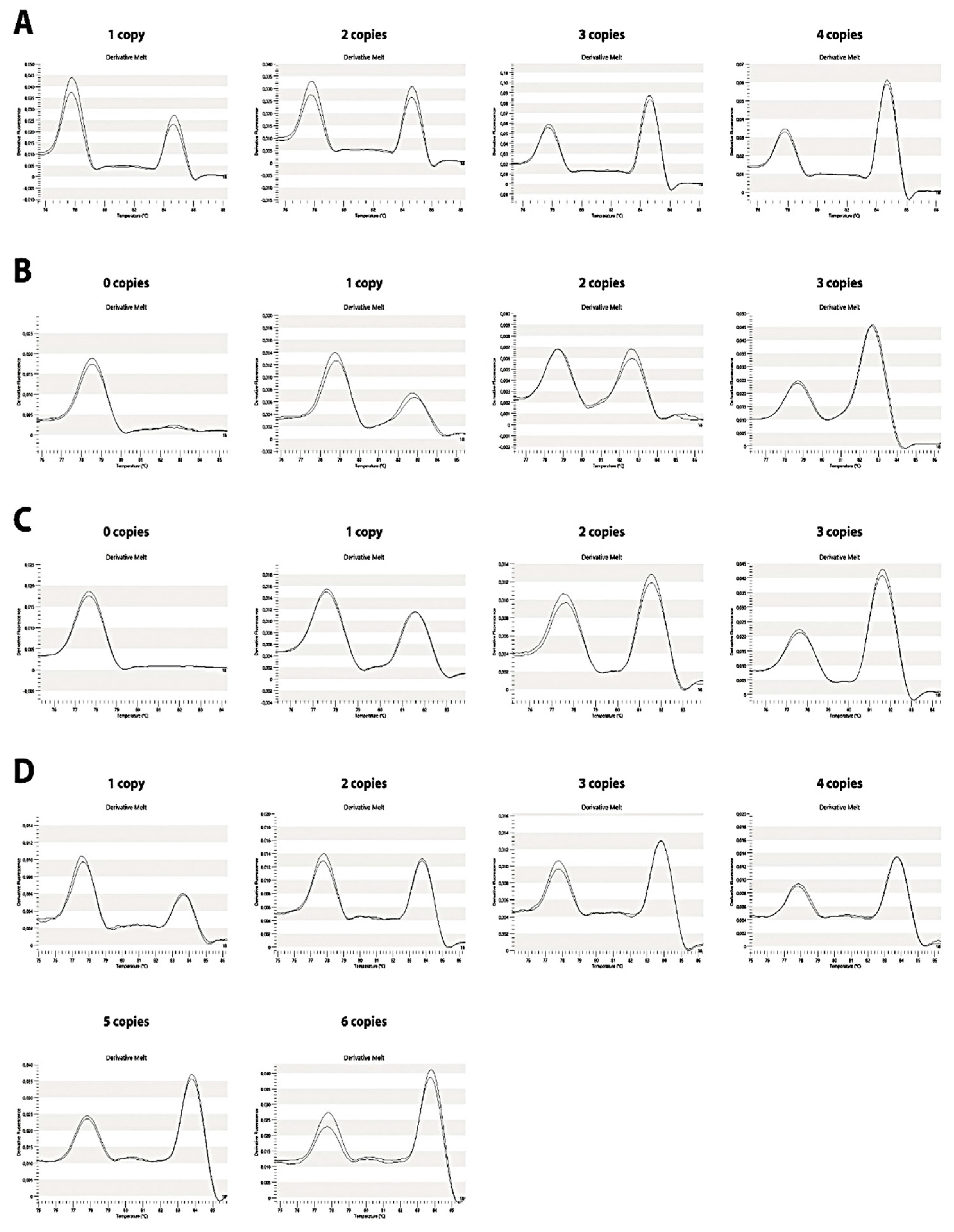
2.1. C4 CNV GRACE-PCR Design and Optimization
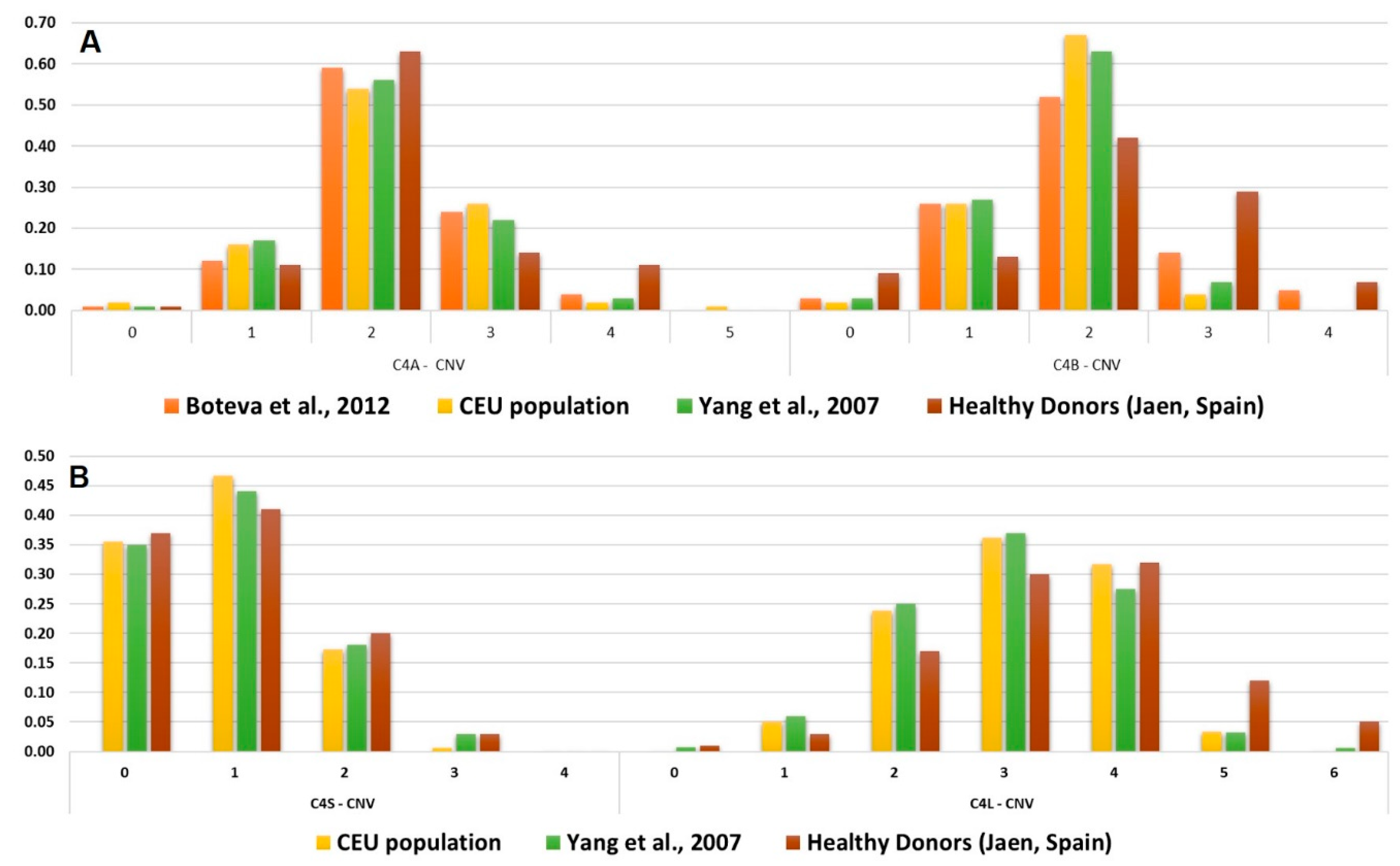
2.2. Validation of the C4 CNV Determined by HRM-PCR/GRACE-PCR Assay
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Blood DNA Extraction
4.3. DNA Quantification
4.4. qPCR Oligonucleotides
4.5. Number of PCR Cycles
4.6. Optimization of HRM-PCR/GRACE-PCR Assay
4.7. Standard Curve
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| C4A | Complement component C4A |
| C4B | Complement component C4B |
| C4L | long C4 gene |
| C4S | short C4 gene |
| CNV | Copy Number Variation |
| GRACE | Gene Ratio Analysis Copy Enumeration |
| HERV-K | Polymorphic endogenous retrovirus insertions |
| HRM | High Resolution Melting |
| STK19 | serine/threonine kinase 19 (STK19), also known as HLA-RP1, here referred to as RP1 |
References
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.Y.; Chung, E.K.; Yang, Y.; Blanchong, C.A.; Jacobsen, N.; Saxena, K.; Yang, Z.; Miller, W.; Varga, L.; Fust, G. Dancing with Complement C4 and the RP-C4-CYP21-TNX (RCCX) Modules of the Major Histocompatibility Complex. Prog. Nucleic Acid Res. Mol. Bio. 2003, 75, 217–292. [Google Scholar] [CrossRef]
- Blanchong, C.A.; Chung, E.K.; Rupert, K.L.; Yang, Y.; Yang, Z.; Zhou, B.; Moulds, M.; Yu, C.Y. Genetic, structural and functional diversities of human complement components C4A and C4B and their mouse homologues, Slp and C4. Int. Immunopharmacol. 2001, 1, 365–392. [Google Scholar] [CrossRef]
- Fernando, M.M.; Boteva, L.; Morris, D.L.; Zhou, B.; Wu, Y.L.; Lokki, M.-L.; Yu, C.Y.; Rioux, J.D.; Hollox, E.J.; Vyse, T.J. Assessment of complement C4 gene copy number using the paralog ratio test. Hum. Mutat. 2010, 31, 866–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleynen, I.; Konings, P.; Robberecht, C.; Amininejad, L.; Machiels, K.; Rutgeerts, P.; Louis, E.; Franchimont, D.; Laukens, D.; Théâtre, E.; et al. Genome-Wide Copy Number Variation Scan Identifies Complement Component C4 as Novel Susceptibility Gene for Crohnʼs Disease. Inflamm. Bowel Dis. 2016, 22, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Sekar, A.; So, H.-C.; Bialas, A.R.; De Rivera, H.; Davis, A.; Hammond, T.R.; Kamitaki, N.; Tooley, K.; Presumey, J.; Buxbaum, J.D.; et al. Schizophrenia risk from complex variation of complement component 4. Nature 2016, 530, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Flachsbart, F.; Caliebe, A.; Heinsen, F.-A.; Hemming-Karlsen, T.; Schreiber, S.; Franke, A.; Nebel, A. Investigation of Complement Component C4 Copy Number Variation in Human Longevity. PLoS ONE 2014, 9, e86188. [Google Scholar] [CrossRef] [Green Version]
- Nowakowska, B. Clinical interpretation of copy number variants in the human genome. J. Appl. Genet. 2017, 58, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhang, J.; Liao, D.; Yang, L.; Wang, Y.; Hou, S. Association between C4, C4A, and C4B copy number variations and susceptibility to autoimmune diseases: A meta-analysis. Sci. Rep. 2017, 7, 42628. [Google Scholar] [CrossRef] [Green Version]
- Gilliam, B.E.; Wolff, A.E.; Moore, T.L. Partial C4 Deficiency in Juvenile Idiopathic Arthritis Patients. J. Clin. Rheumatol. 2007, 13, 256–260. [Google Scholar] [CrossRef]
- Zorzetto, M.; Datturi, F.; Divizia, L.; Pistono, C.; Campo, I.; De Silvestri, A.; Cuccia, M.; Ricevuti, G. Complement C4A and C4B gene copy number study in Alzheimer’s disease patients. Curr. Alzheimer Res. 2016, 13, 1. [Google Scholar] [CrossRef]
- Milich, D.R. Influence of C4A deficiency on nonresponse to HBsAg vaccination: A new immune response gene. J. Hepatol. 2002, 37, 396–399. [Google Scholar] [CrossRef]
- Höhler, T.; Stradmann-Bellinghausen, B.; Starke, R.; Sänger, R.; Victor, A.; Rittner, C.; Schneider, P.M. C4A deficiency and nonresponse to hepatitis B vaccination. J. Hepatol. 2002, 37, 387–392. [Google Scholar] [CrossRef]
- Kainulainen, L.; Peltola, V.; Seppänen, M.; Viander, M.; He, Q.; Lokki, M.-L.; Ruuskanen, O. C4A deficiency in children and adolescents with recurrent respiratory infections. Hum. Immunol. 2012, 73, 498–501. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.L.; Savelli, S.L.; Yang, Y.; Zhou, B.; Rovin, B.H.; Birmingham, D.J.; Nagaraja, H.N.; Hebert, L.A.; Yu, C.Y. Sensitive and specific real-time polymerase chain reaction assays to accurately determine copy number variations (CNVs) of human complement C4A, C4B, C4-long, C4-short, and RCCX modules: Elucidation of C4 CNVs in 50 consanguineous subjects with defined HLA genotypes. J. Immunol. 2007, 179, 3012–3025. [Google Scholar] [CrossRef] [Green Version]
- Paakkanen, R.; Vauhkonen, H.; Eronen, K.T.; Järvinen, A.; Seppänen, M.; Lokki, M.-L. Copy Number Analysis of Complement C4A, C4B and C4A Silencing Mutation by Real-Time Quantitative Polymerase Chain Reaction. PLoS ONE 2012, 7, e38813. [Google Scholar] [CrossRef]
- Grassmann, F.; Cantsilieris, S.; Schulz-Kuhnt, A.-S.; White, S.J.; Richardson, A.J.; Hewitt, A.W.; Vote, B.J.; Schmied, D.; Guymer, R.; Weber, B.H.; et al. Multiallelic copy number variation in the complement component 4A (C4A) gene is associated with late-stage age-related macular degeneration (AMD). J. Neuroinflamm. 2016, 13, 81. [Google Scholar] [CrossRef] [Green Version]
- Hollox, E.J.; Hoh, B.-P. Human gene copy number variation and infectious disease. Hum. Genet. 2014, 133, 1217–1233. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Jeon, J.-T. Methods to detect and analyze copy number variations at the genome-wide and locus-specific levels. Cytogenet. Genome Res. 2008, 123, 333–342. [Google Scholar] [CrossRef]
- Cassinari, K.; Quenez, O.; Joly-Hélas, G.; Beaussire, L.; Le Meur, N.; Castelain, M.; Goldenberg, A.; Guerrot, A.-M.; Brehin, A.-C.; Deleuze, J.-F.; et al. A Simple, Universal, and Cost-Efficient Digital PCR Method for the Targeted Analysis of Copy Number Variations. Clin. Chem. 2019, 65, 1153–1160. [Google Scholar] [CrossRef]
- Turner, A.; Sasse, J.; Váradi, A. Development and validation of a high throughput, closed tube method for the determination of haemoglobin alpha gene (HBA1 and HBA2) numbers by gene ratio assay copy enumeration-PCR (GRACE-PCR). BMC Med. Genet. 2015, 16, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrovic, A. High-Resolution Assessment of Copy Number Variation. Clin. Chem. 2015, 61, 684–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Chung, E.K.; Zhou, B.; Blanchong, C.A.; Yu, C.Y.; Füst, G.; Kovács, M.; Vatay, A.; Szalai, C.; Karadi, I.; et al. Diversity in intrinsic strengths of the human complement system: Serum C4 protein concentrations correlate with C4 gene size and polygenic variations, hemolytic activities, and body mass index. J. Immunol. 2003, 171, 2734–2745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.; Sasse, J.; Váradi, A. Rapid detection of pathological mutations and deletions of the haemoglobin beta gene (HBB) by High Resolution Melting (HRM) analysis and Gene Ratio Analysis Copy Enumeration PCR (GRACE-PCR). BMC Med. Genet. 2016, 17, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szilagyi, A.; Blaskó, B.; Szilassy, D.; Füst, G.; Sasvari-Szekely, M.; Ronai, Z. Real-time PCR quantification of human complement C4A and C4B genes. BMC Genet. 2006, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Real, L.M.; Neukam, K.; Herrero, R.; Guardiola, J.M.; Reiberger, T.; Rivero-Juárez, A.; Salazar, J.; Mandorfer, M.; Merino, D.; Sorianob, V.; et al. IFNL4 ss469415590 Variant Shows Similar Performance to rs12979860 as Predictor of Response to Treatment against Hepatitis C Virus Genotype 1 or 4 in Caucasians. PLoS ONE 2014, 9, e95515. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line. | C4A | C4B | C4S | C4L | Haplotypes |
|---|---|---|---|---|---|
| COX | 0 | 2 | 2 | 0 | BS BS |
| WT51 | 4 | 0 | 0 | 4 | AL-AL AL–AL |
| MADURA | 2 | 2 | 4 | 0 | AS-AB AS-BS |
| CB6B | 4 | 4 | 2 | 6 | AL-AL-BL-BS AL-AL-BL-BS |
| WT8 | 1 | 2 | 0 | 3 | AL-BL BL |
| DAUDI | 2 | 1 | 1 | 2 | AL-BS AL |
| MANIKA | 2 | 3 | 3 | 2 | AL-BS-BS AL-BS |
| HOM2 | 3 | 2 | 1 | 4 | AL-AL-BS AL-BL |
| Target | Primers | Primer Sequence (5’ →3’) | Amplicon Size (bp) | Amplicon Tm (°C) |
|---|---|---|---|---|
| C4A | C4A-F | CCTTTGTGTTGAAGGTCCTGAGTT* | 141 | 84.8 |
| C4A-R | TCCTGTCTAACACTGGACAGGGGT* | |||
| C4B | C4B-F | TGCAGGAGACATCTAACTGGCTTCT* | 86 | 81.6 |
| C4B-R | CATGCTCCTATGTATCACTGGAGAGA* | |||
| C4S | C4S-F | TTGCTCGTTCTGCTCATTCCTT* | 103 | 81.5 |
| C4S-R | GGCGCAGGCTGCTGTATT* | |||
| C4L | C4L-F | TTGCTCGTTCTGCTCATTCCTT* | 133 | 83.7 |
| C4L-R | CCAATGGACTTCAGGAACCC | |||
| RP1 | RP1-F | GACCAAATGACACAGACCTTTGG* | 79 | 77.6 |
| RP1-R | GACTTTGGTTGGTTCCACAAGTC* |
| Gene | Final Concentration | Tube |
|---|---|---|
| C4A | 0.5 µM | Tube 1 |
| RP1 | 0.33 µM | |
| C4B | 0.21 µM | Tube 2 |
| RP1 | 0.16 µM | |
| C4S | 0.25 µM | Tube 3 |
| RP1 | 0.5 µM | |
| C4L | 0.5 µM | Tube 4 |
| RP1 | 0.25 µM |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaimes-Bernal, C.P.; Trujillo, M.; Márquez, F.J.; Caruz, A. Complement C4 Gene Copy Number Variation Genotyping by High Resolution Melting PCR. Int. J. Mol. Sci. 2020, 21, 6309. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176309
Jaimes-Bernal CP, Trujillo M, Márquez FJ, Caruz A. Complement C4 Gene Copy Number Variation Genotyping by High Resolution Melting PCR. International Journal of Molecular Sciences. 2020; 21(17):6309. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176309
Chicago/Turabian StyleJaimes-Bernal, Claudia P., Monte Trujillo, Francisco José Márquez, and Antonio Caruz. 2020. "Complement C4 Gene Copy Number Variation Genotyping by High Resolution Melting PCR" International Journal of Molecular Sciences 21, no. 17: 6309. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176309