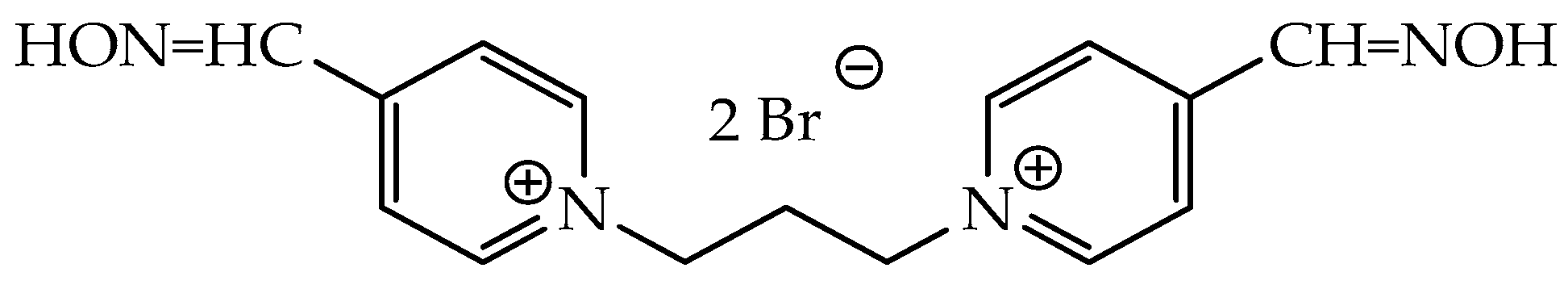
Understanding the Interaction Modes and Reactivity of Trimedoxime toward MmAChE Inhibited by Nerve Agents: Theoretical and Experimental Aspects

 ,
, 

Abstract
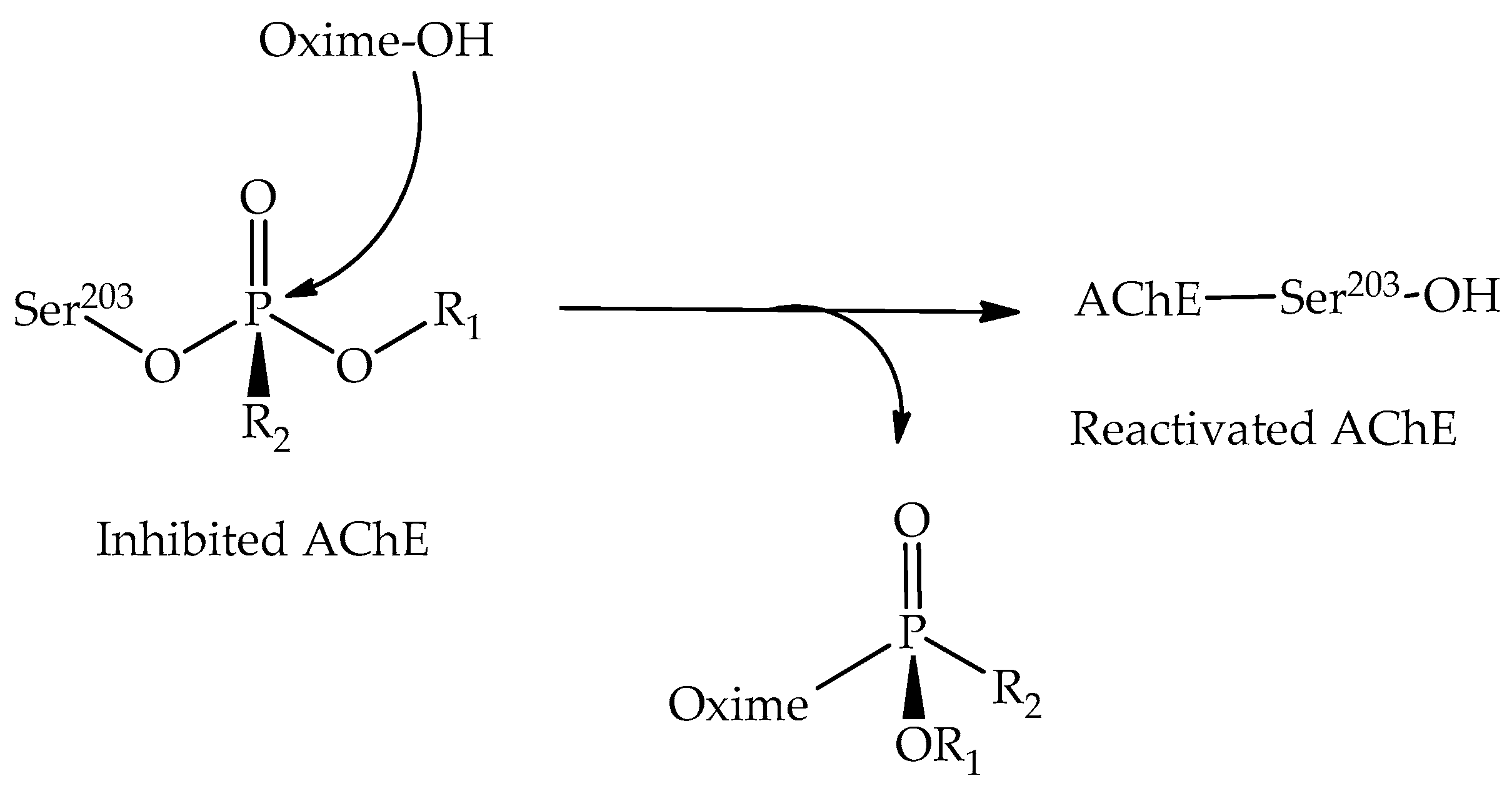
:1. Introduction
2. Results and Discussion
2.1. In Vitro Test: Experimental Results
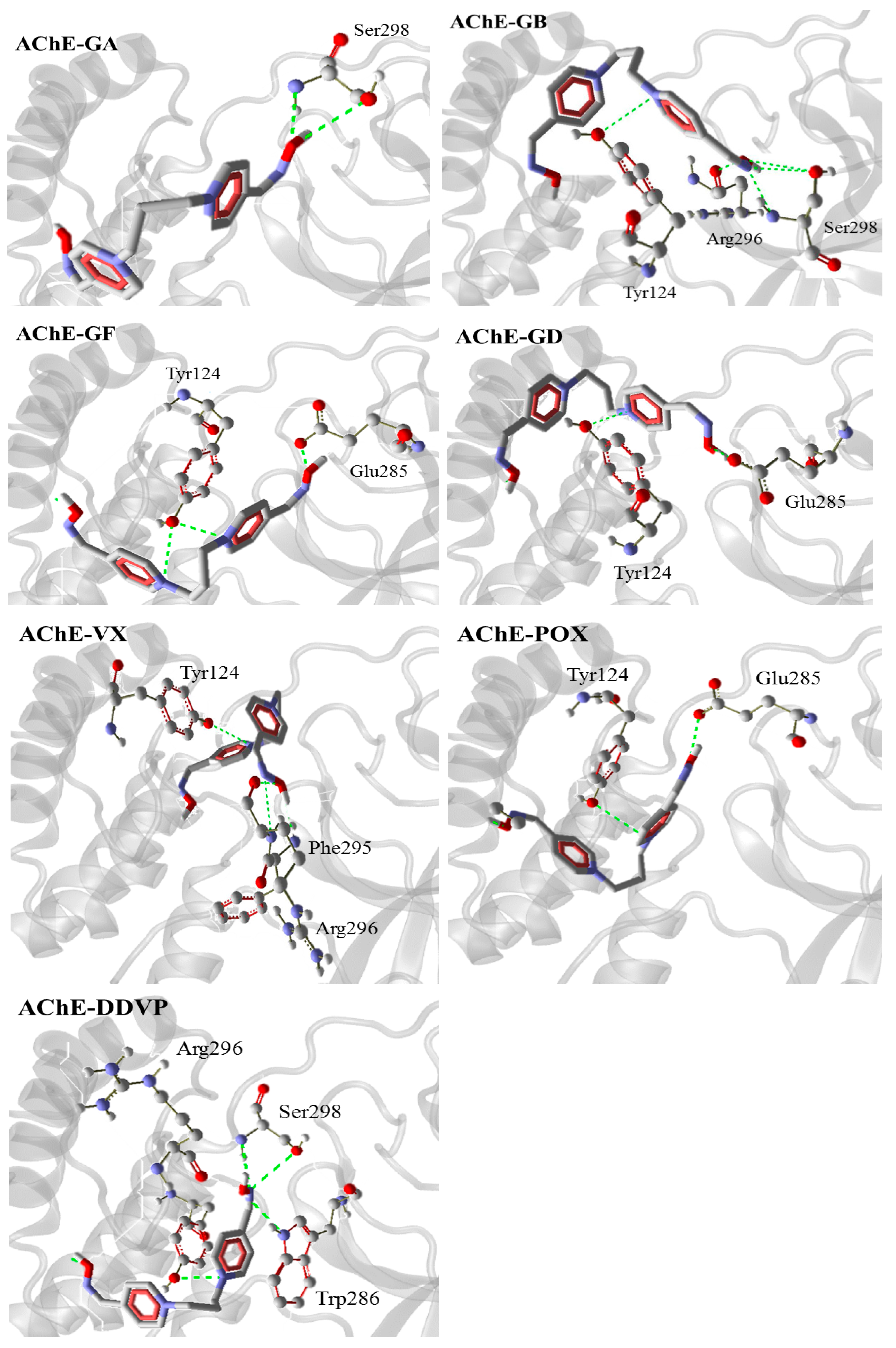
2.2. Affinity and Thermodynamics: Docking Results
2.3. Investigating Kinetic Parameters for Biological Activity: Mechanistic Studies
3. Materials and Methods
3.1. Experimental Details
3.2. Docking Procedure
3.3. QM/MM Procedure
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AChE | Acetylcholinesterase |
| MmAChE | Mus musculus Acetylcholinesterase |
| OP | Organophosphorus compounds |
| ACh | Acetylcholine |
| BBB | Blood Brain Barrier |
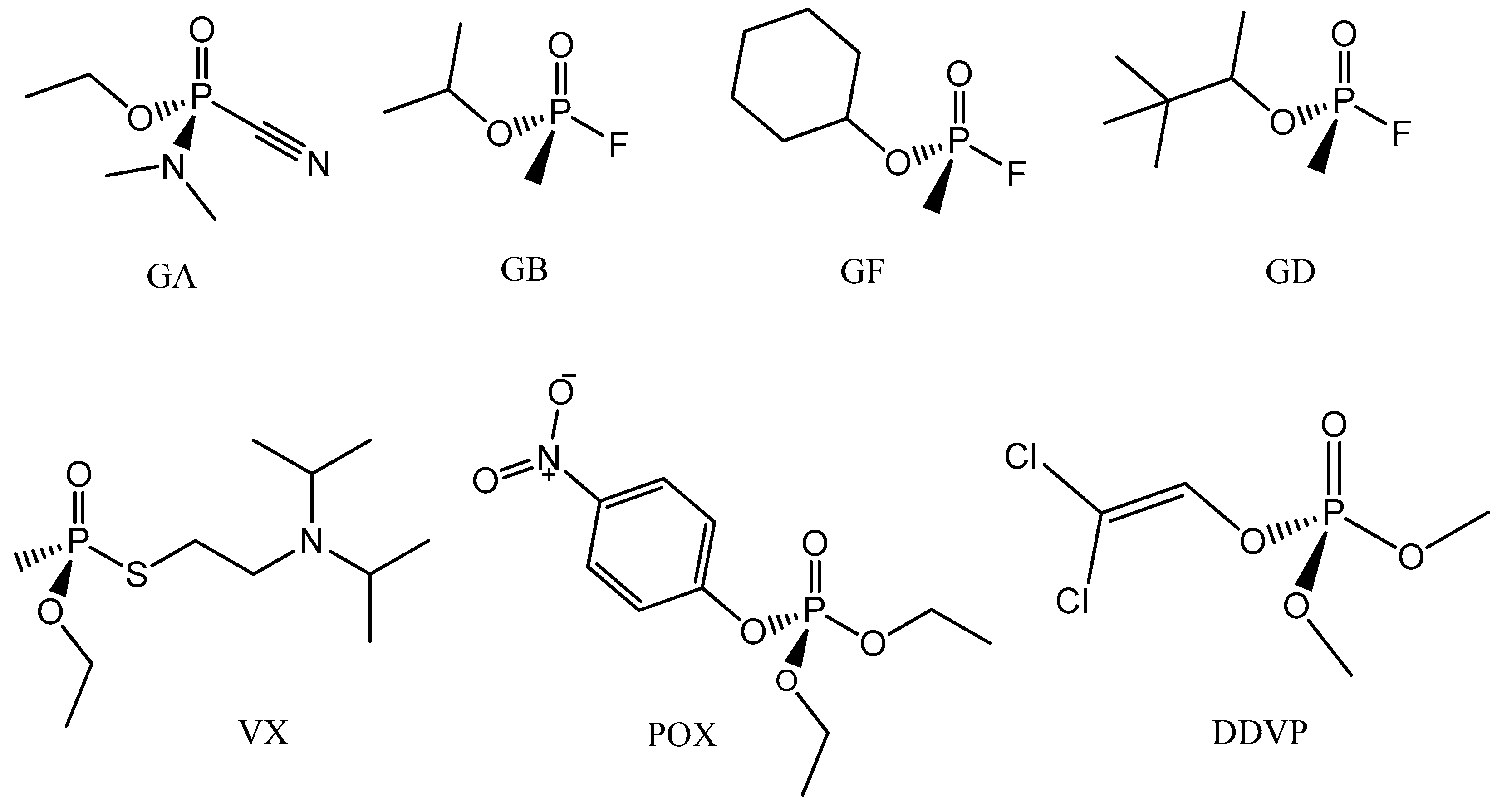
| GA | Tabun |
| GB | Sarin |
| GF | Cyclosarin |
| GD | Soman |
| POX | Paraoxon |
| DDVP | Dichlorvos |
| SER | Serine |
| Tyr | Tyrosine |
| ARG | Arginine |
| GLU | Glutamic acid |
| PHE | Phenylalanine |
| TRP | Tryptophan |
| QM/MM | Quantum Mechanics/Molecular Mechanics |
| MLR | Multiple Linear Regression |
| TLC | Thin Layer Chromatography |
| HPLC | High-performance liquid chromatography |
| NMR | Nuclear Magnetic Resonance |
| MVD | Molegro Virtual Docker |
References
- Sadik, O.A.; Land, W.H.; Wang, J. Targeting Chemical and Biological Warfare Agents at the Molecular Level. Electroanal. 2003, 15, 1149–1159. [Google Scholar] [CrossRef]
- Ramalho, T.C.; de Castro, A.A.; Silva, D.R.; Silva, M.C.; Franca, T.C.C.; Bennion, B.J.; Kuca, K. Computational Enzymology and Organophosphorus Degrading Enzymes: Promising Approaches toward Remediation Technologies of Warfare Agents and Pesticides. Curr. Med. Chem. 2016, 23, 1041–1061. [Google Scholar] [CrossRef] [PubMed]
- Spiers, J. New perspectives on vulnerability using emic and etic approaches. J. Adv. Nurs. 2000, 31, 715–721. [Google Scholar] [CrossRef] [PubMed]
- França, T.C.C.; Silva, G.R.; Castro, A.T. De Chemical Defense: A new subject in the Chemical Teaching. Rev. Virtual Quím. 2010, 2, 84–104. [Google Scholar]
- Chauhan, S.; Chauhan, S.; D’Cruz, R.; Faruqi, S.; Singh, K.K.; Varma, S.; Singh, M.; Karthik, V. Chemical warfare agents. Environ. Toxicol. Pharmacol. 2008, 26, 113–122. [Google Scholar] [CrossRef]
- Gravett, M.R.; Hopkins, F.B.; Self, A.J.; Webb, A.J.; Timperley, C.M.; Baker, M.J. Evidence of VX nerve agent use from contaminated white mustard plants. Proc. R. Soc. A Math. Phys. Eng. Sci. 2014, 470, 20140076. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, K.; Raza, S.; Vijayaraghavan, R. Chemical warfare agents. J. Pharm. Bioallied Sci. 2010, 2, 166–178. [Google Scholar] [CrossRef]
- El-Ebiary, A.A.; Elsharkawy, R.E.; Soliman, N.A.; Soliman, M.A.; Hashem, A.A. N-acetylcysteine in Acute Organophosphorus Pesticide Poisoning: A Randomized, Clinical Trial. Basic Clin. Pharmacol. Toxicol. 2016, 119, 222–227. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, V.M.R.; Donnici, C.L.; DaCosta, J.B.N.; Caixeiro, J.M.R. Organophosphorus pentavalent compounds: History, synthetic methods of preparation and application as insecticides and antitumor agents. Quim. Nova 2007, 30, 159–170. [Google Scholar]
- Black, R.M.; Read, R.W. Biological markers of exposure to organophosphorus nerve agents. Arch. Toxicol. 2013, 87, 421–437. [Google Scholar] [CrossRef]
- Dong, H.; Weng, Y.B.; Zhen, G.S.; Li, F.J.; Jin, A.C.; Liu, J.; Pany, S. Clinical emergency treatment of 68 critical patients with severe organophosphorus poisoning and prognosis analysis after rescue. Medicine 2017, 96, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Kassa, J.; Korabecny, J.; Nepovimova, E.; Jun, D. The influence of modulators of acetylcholinesterase on the resistance of mice against soman and on the effectiveness of antidotal treatment of soman poisoning in mice. J. Appl. Biomed. 2018, 16, 10–14. [Google Scholar] [CrossRef]
- Worek, F.; Thiermann, H.; Wille, T. Organophosphorus compounds and oximes: A critical review. Arch. Toxicol. 2020, 94, 2275–2292. [Google Scholar] [CrossRef] [PubMed]
- de Castro, A.A.; Assis, L.C.; Soares, F.V.; Kuca, K.; Polisel, D.A.; da Cunha, E.F.F.; Ramalho, T.C. Trends in the Recent Patent Literature on Cholinesterase Reactivators (2016–2019). Biomolecules 2020, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Chambers, J.E.; Meek, E.C. Novel centrally active oxime reactivators of acetylcholinesterase inhibited by surrogates of sarin and VX. Neurobiol. Dis. 2020, 133, 104487. [Google Scholar] [CrossRef]
- Lorke, D.E.; Petroianu, G.A. The Experimental Oxime K027-A Promising Protector From Organophosphate Pesticide Poisoning. A Review Comparing K027, K048, Pralidoxime, and Obidoxime. Front. Neurosci. 2019, 13, 427. [Google Scholar] [CrossRef]
- Herbert, J.; Thiermann, H.; Worek, F.; Wille, T. Precision cut lung slices as test system for candidate therapeutics in organophosphate poisoning. Toxicology 2017, 389, 94–100. [Google Scholar] [CrossRef]
- Alencar Filho, E.B.; Santos, A.A.; Oliveira, B.G. A quantum chemical study of molecular properties and QSPR modeling of oximes, amidoximes and hydroxamic acids with nucleophilic activity against toxic organophosphorus agents. J. Mol. Struct. 2017, 1133, 338–347. [Google Scholar] [CrossRef]
- Malfatti, M.A.; Enright, H.A.; Be, N.A.; Kuhn, E.A.; Hok, S.; McNerney, M.W.; Lao, V.; Nguyen, T.H.; Lightstone, F.C.; Carpenter, T.S.; et al. The biodistribution and pharmacokinetics of the oxime acetylcholinesterase reactivator RS194B in guinea pigs. Chem. Biol. Interact. 2017, 277, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Kuča, K.; Kassa, J. A comparison of the ability of a new bispyridinium oxime—1-(4-hydroxyiminomethylpyridinium)-4-(4-carbamoylpyridinium)butane dibromide and currently used oximes to reactivate nerve agent-inhibited rat brain acetylcholinesterase by in vitro methods. J. Enzym. Inhib. Med. Chem. 2003, 18, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Delfino, R.T.; Ribeiro, T.S.; Figueroa-Villar, J.D. Organophosphorus compounds as chemical warfare agents: A review. Sect. Title Toxicol. 2009, 20, 407–428. [Google Scholar] [CrossRef]
- Bajgar, J.; Fusek, J.; Kuca, K.; Bartosova, L.; Jun, D. Treatment of Organophosphate Intoxication Using Cholinesterase Reactivators:Facts and Fiction. Mini-Rev. Med. Chem. 2007, 7, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Polisel, D.A.; de Castro, A.A.; Mancini, D.T.; da Cunha, E.F.F.; França, T.C.C.; Ramalho, T.C.; Kuca, K. Slight difference in the isomeric oximes K206 and K203 makes huge difference for the reactivation of organophosphorus-inhibited AChE: Theoretical and experimental aspects. Chem. Biol. Interact. 2019, 309, 108671. [Google Scholar] [CrossRef] [PubMed]
- Matos, K.S.; Mancini, D.T.; da Cunha, E.F.F.; Kuča, K.; França, T.C.C.; Ramalho, T.C. Molecular aspects of the reactivation process of acetylcholinesterase inhibited by cyclosarin. J. Braz. Chem. Soc. 2011, 6, 286–289. [Google Scholar] [CrossRef] [Green Version]
- Giacoppo, J.O.S.; França, T.C.C.; Kuča, K.; Da Cunha, E.F.F.; Abagyan, R.; Mancini, D.T.; Ramalho, T.C. Molecular modeling and in vitro reactivation study between the oxime BI-6 and acetylcholinesterase inhibited by different nerve agents. J. Biomol. Struct. Dyn. 2015, 33, 2048–2058. [Google Scholar] [CrossRef]
- Kuca, K.; Musilek, K.; Jun, D.; Zdarova-Karasova, J.; Nepovimova, E.; Soukup, O.; Hrabinova, M.; Mikler, J.; Franca, T.C.C.; Da Cunha, E.F.F.; et al. A newly developed oxime K203 is the most effective reactivator of tabun-inhibited acetylcholinesterase. BMC Pharmacol. Toxicol. 2018, 19, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Musilek, K.; Holas, O.; Kuca, K.; Jun, D.; Dohnal, V.; Opletalova, V.; Dolezal, M. Synthesis of monooxime-monocarbamoyl bispyridinium compounds bearing (E)-but-2-ene linker and evaluation of their reactivation activity against tabun- and paraoxon-inhibited acetylcholinesterase. J. Enzym. Inhib. Med. Chem. 2008, 23, 70–76. [Google Scholar] [CrossRef]
- Jun, D.; Stodulka, P.; Kuca, K.; Dolezal, B. High-performance liquid chromatography analysis of by-products and intermediates arising during the synthesis of the acetylcholinesterase reactivator HI-6. J. Chromatogr. Sci. 2010, 48, 694–696. [Google Scholar] [CrossRef] [Green Version]
- Kuča, K.; Cabal, J. Evaluation of Newly Synthesized Reactivators of the Brain Cholinesterase Inhibited by Sarin Nerve Agent. Toxicol. Mech. Methods 2005, 15, 247–252. [Google Scholar] [CrossRef]
- Gaussian 09, Revision A.02; Gaussian, Inc.: Wallingford, CT, USA, 2009.
- Artursson, E.; Andersson, P.O.; Akfur, C.; Linusson, A.; Börjegren, S.; Ekström, F. Catalytic-site conformational equilibrium in nerve-agent adducts of acetylcholinesterase: Possible implications for the HI-6 antidote substrate specificity. Biochem. Pharmacol. 2013, 85, 1389–1397. [Google Scholar] [CrossRef]
- Thomsen, R.; Christensen, M.H. MolDock: A New Technique for High-Accuracy Molecular Docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.C.; dos Santos Pires, M.; de Castro, A.A.; da Cunha, E.F.F.; Caetano, M.S.; Ramalho, T.C. Molecular insight into the inhibition mechanism of plant and rat 4-hydroxyphenylpyruvate dioxygenase by molecular docking and DFT calculations. Med. Chem. Res. 2015, 24, 3958–3971. [Google Scholar] [CrossRef]
- Guimaraes, A.P.; Oliveira, A.A.; da Cunha, E.F.F.; Ramalho, T.C.; Franco, T.C.C. Analysis of Bacillus anthracis nucleoside hydrolase via in silico docking with inhibitors and molecular dynamics simulation. J. Mol. Model. 2011, 17, 2939–2951. [Google Scholar] [CrossRef]
- da Cunha, E.F.F.; Barbosa, E.F.; Oliveira, A.A.; Ramalho, T.C. Molecular Modeling of Mycobacterium Tuberculosis DNA Gyrase and its Molecular Docking Study with Gatifloxacin Inhibitors. J. Biomol. Struct. Dyn. 2010, 27, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Souza, T.C.S.; Josa, D.; Ramalho, T.C.; Caetano, M.S.; da Cunha, E.F.F. Molecular modelling of Mycobacterium tuberculosis acetolactate synthase catalytic subunit and its molecular docking study with inhibitors. Mol. Simul. 2008, 34, 707–713. [Google Scholar] [CrossRef]
- Nemukhin, A.V.; Grigorenko, B.L.; Morozov, D.I.; Kochetov, M.S.; Lushchekina, S.V.; Varfolomeev, S.D. On quantum mechanical-molecular mechanical (QM/MM) approaches to model hydrolysis of acetylcholine by acetylcholinesterase. Chem. Biol. Interact. 2013, 203, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Gonçalves, A.; França, T.C.C.; Caetano, M.S.; Ramalho, T.C. Reactivation steps by 2-PAM of tabun-inhibited human acetylcholinesterase: Reducing the computational cost in hybrid QM/MM methods. J. Biomol. Struct. Dyn. 2014, 32, 301–307. [Google Scholar] [CrossRef]
- Matos, K.; Cunha, E.; Abagyan, R.; Ramalho, T. Computational Evidence for the Reactivation Process of Human Acetylcholinesterase Inhibited by Carbamates. Comb. Chem. High Throughput Screen. 2013, 17, 554–564. [Google Scholar] [CrossRef]
- Heyden, A.; Lin, H.; Truhlar, D.G. Adaptive partitioning in combined quantum mechanical and molecular mechanical calculations of potential energy functions for multiscale simulations. J. Phys. Chem. B 2007, 111, 2231–2241. [Google Scholar] [CrossRef]
- Ramalho, T.C.; Caetano, M.S.; da Cunha, E.F.F.; Souza, T.C.S.; Rocha, M.V.J. Construction and Assessment of Reaction Models of Class I EPSP Synthase: Molecular Docking and Density Functional Theoretical Calculations. J. Biomol. Struct. Dyn. 2009, 27, 195–207. [Google Scholar] [CrossRef]
- Kuca, K.; Musilek, K.; Jun, D.; Nepovimova, E.; Soukup, O.; Korabecny, J.; França, T.C.C.; de Castro, A.A.; Krejcar, O.; da Cunha, E.F.F.; et al. Oxime K074—In vitro and in silico reactivation of acetylcholinesterase inhibited by nerve agents and pesticides. Toxin Rev. 2020, 39, 157–166. [Google Scholar] [CrossRef]
- da Silva, J.A.; Pereira, A.F.; LaPlante, S.R.; Kuca, K.; Ramalho, T.C.; França, T.C. Reactivation of VX-Inhibited Human Acetylcholinesterase by Deprotonated Pralidoxime. A Complementary Quantum Mechanical Study. Biomolecules 2020, 10, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besler, B.H.; Merz, K.M.; Kollman, P.A. Atomic charges derived from semiempirical methods. J. Comput. Chem. 1990, 11, 431–439. [Google Scholar] [CrossRef]
- Singh, U.C.; Kollman, P.A. An approach to computing electrostatic charges for molecules. J. Comput. Chem. 1984, 5, 129–145. [Google Scholar] [CrossRef]
- Li, R.; Liu, Y.; Zhang, J.; Chen, K.; Li, S.; Jiang, J. An isofenphos-methyl hydrolase (Imh) capable of hydrolyzing the P-O-Z moiety of organophosphorus pesticides containing an aryl or heterocyclic group. Appl. Microbiol. Biotechnol. 2012, 94, 1553–1564. [Google Scholar] [CrossRef] [PubMed]
- Gorecki, L.; Korabecny, J.; Musilek, K.; Malinak, D.; Nepovimova, E.; Dolezal, R.; Jun, D.; Soukup, O.; Kuca, K. SAR study to find optimal cholinesterase reactivator against organophosphorous nerve agents and pesticides. Arch. Toxicol. 2016, 90, 2831–2859. [Google Scholar] [CrossRef] [PubMed]
- da Cunha, E.F.F.; Ramalho, T.C.; Reynolds, R.C. Binding mode analysis of 2,4-diamino-5-methyl-5-deaza-6-substituted pteridines with mycobacterium tuberculosis and human dihydrofolate reductases. J. Biomol. Struct. Dyn. 2008, 25, 377–385. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Trimedoxime | |
|---|---|---|
| React. (%) Conc. 10−5 M | React. (%) Conc. 10−3 M | |
| AChE-GA | 0 | 30 |
| AChE-GB | 7 | 54 |
| AChE-GF | 0 | 0 |
| AChE-GD | 0 | 0 |
| AChE-VX | 9.8 | 85.3 |
| AChE-POX | 50 | 46 |
| AChE-DDVP | 17.3 | 31.5 |
| System | Trimedoxime | |
|---|---|---|
| ∆E * (kcal mol−1) | Residues | |
| AChE-GA | −140.9 | Ser298 |
| AChE-GB | −154.7 | Tyr124, Ser298, Arg296 |
| AChE-GF | −161.3 | Tyr124, Glu285 |
| AChE-GD | −157.7 | Tyr124, Glu285 |
| AChE-VX | −115.0 | Tyr124, Phe295, Arg296 |
| AChE-POX | −144.1 | Tyr124, Glu285 |
| AChE-DDVP | −164.8 | Arg296, Ser298, Trp286 |
| System | Trimedoxime | |
|---|---|---|
| ∆∆E# * (kcal mol−1) | React. (%) Conc. 10−3 M | |
| AChE-GA | 46.83 | 30 |
| AChE-GB | 33.43 | 54 |
| AChE-GF | - | 0 |
| AChE-GD | - | 0 |
| AChE-VX | 0 | 85.3 |
| AChE-POX | 41.59 | 46 |
| AChE-DDVP | 47.75 | 31.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Castro, A.A.; Polisel, D.A.; Pereira, B.T.L.; da Cunha, E.F.F.; Kuca, K.; Nepovimova, E.; Ramalho, T.C. Understanding the Interaction Modes and Reactivity of Trimedoxime toward MmAChE Inhibited by Nerve Agents: Theoretical and Experimental Aspects. Int. J. Mol. Sci. 2020, 21, 6510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186510
de Castro AA, Polisel DA, Pereira BTL, da Cunha EFF, Kuca K, Nepovimova E, Ramalho TC. Understanding the Interaction Modes and Reactivity of Trimedoxime toward MmAChE Inhibited by Nerve Agents: Theoretical and Experimental Aspects. International Journal of Molecular Sciences. 2020; 21(18):6510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186510
Chicago/Turabian Stylede Castro, Alexandre A., Daniel A. Polisel, Bruna T. L. Pereira, Elaine F. F. da Cunha, Kamil Kuca, Eugenie Nepovimova, and Teodorico C. Ramalho. 2020. "Understanding the Interaction Modes and Reactivity of Trimedoxime toward MmAChE Inhibited by Nerve Agents: Theoretical and Experimental Aspects" International Journal of Molecular Sciences 21, no. 18: 6510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186510