Colon Transcriptomics Reveals Sex-Dependent Metabolic Signatures in Response to 2-Amino-1-methyl-6-phenylimidazo[4,5-b]pyridine Treatment in C57BL/6N Mice

 , , and
, , and
Abstract
:1. Introduction
2. Results
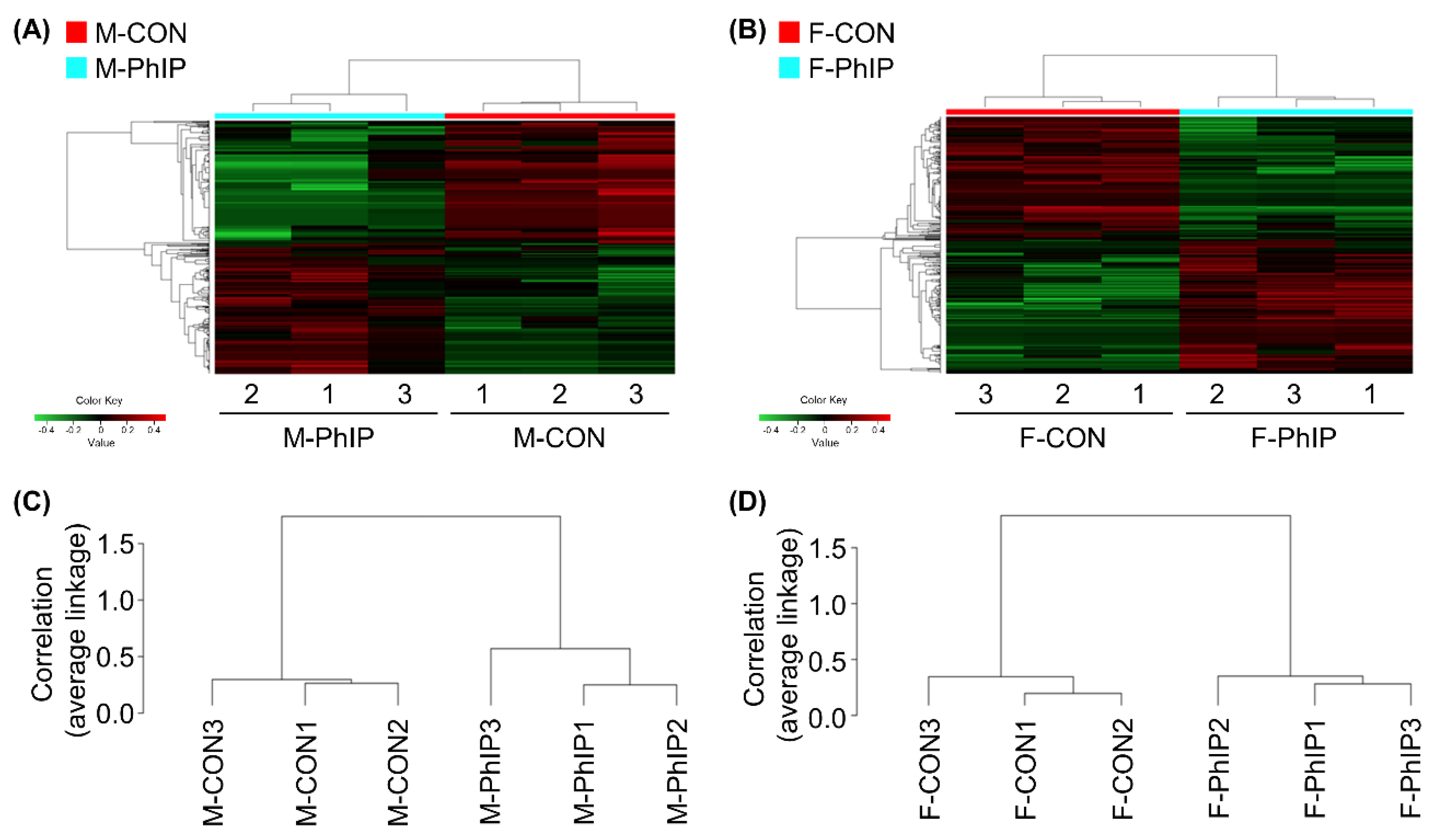
2.1. Identification of DEGs in Response to PhIP Injection
2.2. Comparison of Sex-Dependent Gene Ontology of DEGs Using the PANTHER
2.3. Comparison of Sex-Dependent Canonical Pathways of DEGs Using the IPA
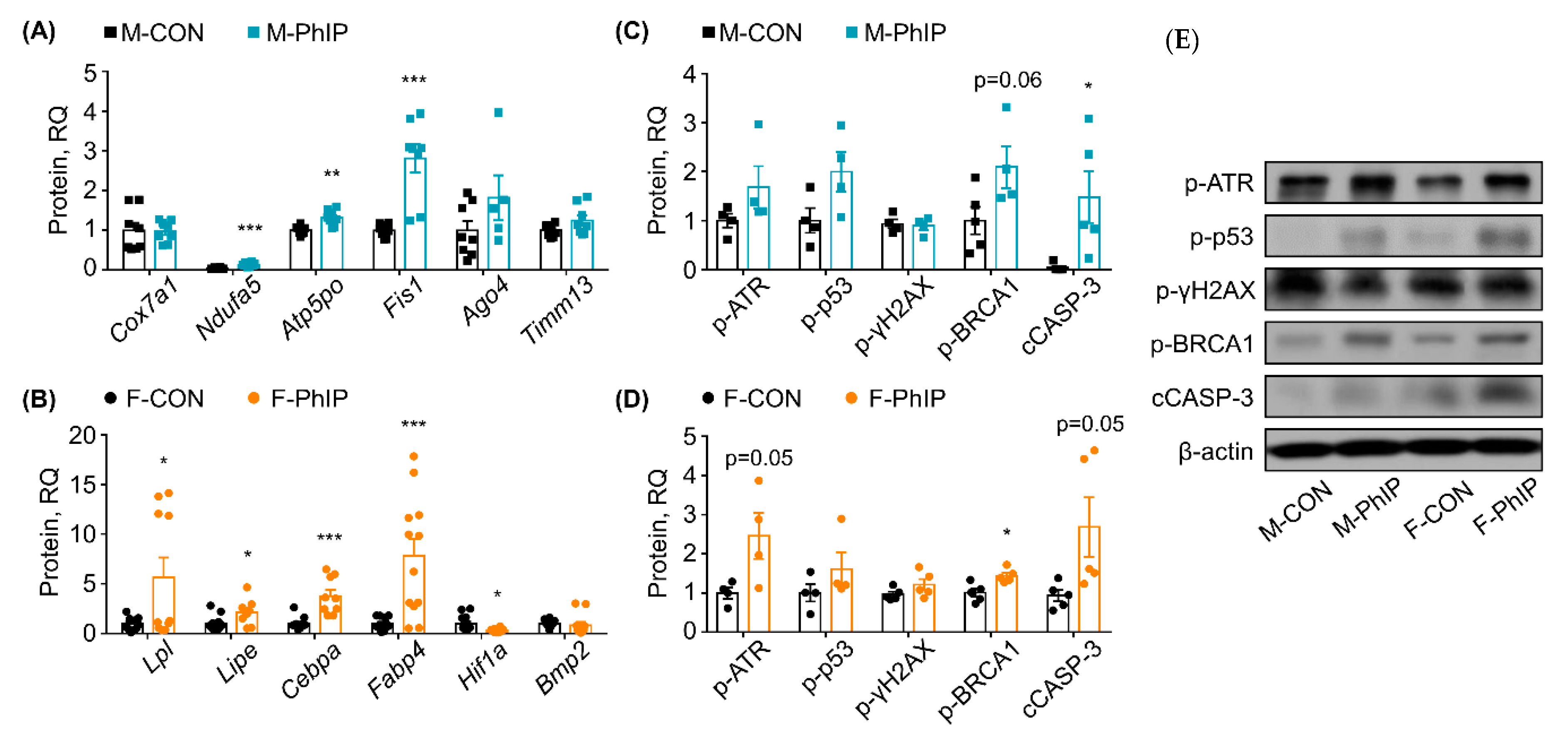
2.4. Quantitative RT-PCR Validation for Predicted Canonical Pathways
2.5. Comparison of Sex-Dependent Effects of PhIP on DNA Damage Response and Apoptosis
3. Discussion
4. Materials and Methods
4.1. Animal Housing and Care
4.2. Study Design and PhIP Treatment
4.3. Colon Transcriptomic Analysis
4.4. Bioinformatics Analyses
4.5. Quantitative RT-PCR Analysis
4.6. Western Blot Analysis
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PhIP | 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine |
| Ago4 | Argonaute 4 |
| Anxa1 | Annexin A1 |
| Atp5po | Mitochondrial ATP synthase subunit O |
| Bmp2 | Bone morphogenetic protein 2 |
| cCASP-3 | Cleaved caspase-3 |
| Cebpa | CCAAT/enhancer-binding protein alpha |
| Ces | Carboxylic ester hydrolase |
| Cox7a1 | Mitochondrial cytochrome c oxidase polypeptide 7a1 |
| CYP | Cytochrome P450 |
| Ddhd2 | DDHD domain-containing protein 2 |
| DDR | DNA damage response |
| DEGs | Differentially expressed genes |
| Fabp4 | Fatty acid binding protein 4 |
| F-CON | Female control group |
| Fis1 | Mitochondrial fission 1 |
| F-PhIP | Female PhIP group |
| GO | Gene ontology |
| HAAs | Heterocyclic aromatic amines |
| Hif1a | Hypoxia-inducible factor 1-alpha |
| IPA | Ingenuity pathway analysis |
| Lipe | Hormone-sensitive lipase |
| Lpl | Lipoprotein lipase |
| M-CON | Male control group |
| M-PhIP | Male PhIP group |
| mtDNA | Mitochondrial DNA |
| Ndufa5 | NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 5 |
| OXPHOS | Oxidative phosphorylation |
| PANTHER | Protein annotation through evolutionary relationships |
| p-ATR | Phosphorylated ataxia telangiectasia and Rad3 related |
| p-BRCA1 | Phosphorylated breast cancer type 1 |
| p-γH2AX | Phosphorylated H2A histone family member X |
| p-p53 | Phosphorylated tumor protein p53 |
| qPCR | Quantitative real-time PCR |
| RIN | RNA integrity number |
| RNAseq | RNA sequencing |
| Timm13 | Mitochondrial import inner membrane translocase subunit Tim13 |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Turner, N.D.; Lloyd, S.K. Association between red meat consumption and colon cancer: A systematic review of experimental results. Exp. Biol. Med. (Maywood) 2017, 242, 813–839. [Google Scholar] [CrossRef] [Green Version]
- Cross, A.J.; Sinha, R. Meat-related mutagens/carcinogens in the etiology of colorectal cancer. Environ. Mol. Mutagen. 2004, 44, 44–55. [Google Scholar] [CrossRef]
- Ochiai, M.; Watanabe, M.; Kushida, H.; Wakabayashi, K.; Sugimura, T.; Nagao, M. DNA adduct formation, cell proliferation and aberrant crypt focus formation induced by PhIP in male and female rat colon with relevance to carcinogenesis. Carcinogenesis 1996, 17, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Buonarati, M.H.; Turteltaub, K.W.; Shen, N.H.; Felton, J.S. Role of sulfation and acetylation in the activation of 2-hydroxyamino-1-methyl-6-phenylimidazo[4,5-b]pyridine to intermediates which bind DNA. Mutat. Res. Lett. 1990, 245, 185–190. [Google Scholar] [CrossRef]
- Hein, D.W.; Rustan, T.D.; Ferguson, R.J.; Doll, M.A.; Gray, K. Metabolic activation of aromatic and heterocyclic N-hydroxyarylamines by wild-type and mutant recombinant human NAT1 and NAT2 acetyltransferases. Arch. Toxicol. 1994, 68, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Muckel, E.; Frandsen, H.; Glatt, H.R. Heterologous expression of human N-acetyltransferases 1 and 2 and sulfotransferase 1A1 in Salmonella typhimurium for mutagenicity testing of heterocyclic amines. Food Chem. Toxicol. 2002, 40, 1063–1068. [Google Scholar] [CrossRef]
- Chou, H.C.; Lang, N.P.; Kadlubar, F.F. Metabolic activation of N-hydroxy arylamines and N-hydroxy heterocyclic amines by human sulfotransferase(s). Cancer Res. 1995, 55, 525–529. [Google Scholar]
- Malfatti, M.A.; Felton, J.S. N-glucuronidation of 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) and N_hydroxy-PhIP by specific human UDP-glucuronosyltransferases. Carcinogenesis 2001, 22, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Coles, B.; Nowell, S.A.; MacLeod, S.L.; Sweeney, C.; Lang, N.P.; Kadlubar, F.F. The role of human glutathione S-transferases (hGSTs) in the detoxification of the food-derived carcinogen metabolite N-acetoxy-PhIP and the effect of a polymorphism in hGSTA1 on colorectal cancer risk. Mutat. Res 2001, 482, 3–10. [Google Scholar] [CrossRef]
- Kim, J.K.; Gallaher, D.D.; Chen, C.; Yao, D.; Trudo, S.P. Apiaceous Vegetable Consumption Decreases PhIP-Induced DNA Adducts and Increases Methylated PhIP Metabolites in the Urine Metabolome in Rats. J. Nutr. 2015, 145, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Glickman, B.W.; de Boer, J.G. Sex-specific induction of mutations by PhIP in the kidney of male and female rats and its modulation by conjugated linoleic acid. Environ. Mol. Mutagen. 2002, 40, 116–121. [Google Scholar] [CrossRef]
- Esumi, H.; Ohgaki, H.; Kohzen, E.; Takayama, S.; Sugimura, T. Induction of lymphoma in CDF1 mice by the food mutagen, 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine. Jpn. J Cancer Res. 1989, 80, 1176–1178. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Hasegawa, R.; Sano, M.; Tamano, S.; Esumi, H.; Takayama, S.; Sugimura, T. A new colon and mammary carcinogen in cooked food, 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP). Carcinogenesis 1991, 12, 1503–1506. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Wang, R.; Dashwood, W.M.; Lohr, C.V.; Williams, D.E.; Ho, E.; Mertens-Talcott, S.; Dashwood, R.H. A miRNA signature for an environmental heterocyclic amine defined by a multi-organ carcinogenicity bioassay in the rat. Arch. Toxicol. 2017, 91, 3415–3425. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.M.; Lee, Y.K.; Park, I.; Kwon, S.M.; Min, S.; Yoon, G. Lactic acidosis caused by repressed lactate dehydrogenase subunit B expression down-regulates mitochondrial oxidative phosphorylation via the pyruvate dehydrogenase (PDH)-PDH kinase axis. J. Biol. Chem. 2019, 294, 7810–7820. [Google Scholar] [CrossRef]
- Yu, M. Generation, function and diagnostic value of mitochondrial DNA copy number alterations in human cancers. Life Sci. 2011, 89, 65–71. [Google Scholar] [CrossRef]
- Amunts, A.; Brown, A.; Toots, J.; Scheres, S.H.W.; Ramakrishnan, V.; Ribosome. The structure of the human mitochondrial ribosome. Science 2015, 348, 95–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.K.; Gallaher, D.D.; Trudo, S.P. Apiaceous vegetable intake modulates expression of DNA damage response genes and microRNA in the rat colon. J. Funct. Foods 2018, 45, 138–145. [Google Scholar] [CrossRef]
- Fahrer, J.; Kaina, B. Impact of DNA repair on the dose-response of colorectal cancer formation induced by dietary carcinogens. Food Chem. Toxicol. 2017, 106, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, I.J. Lipoprotein lipase and lipolysis: Central roles in lipoprotein metabolism and atherogenesis. J. Lipid Res. 1996, 37, 693–707. [Google Scholar]
- Blaner, W.S.; Obunike, J.C.; Kurlandsky, S.B.; al-Haideri, M.; Piantedosi, R.; Deckelbaum, R.J.; Goldberg, I.J. Lipoprotein lipase hydrolysis of retinyl ester. Possible implications for retinoid uptake by cells. J. Biol. Chem. 1994, 269, 16559–16565. [Google Scholar]
- Schupp, M.; Lefterova, M.I.; Janke, J.; Leitner, K.; Cristancho, A.G.; Mullican, S.E.; Qatanani, M.; Szwergold, N.; Steger, D.J.; Curtin, J.C.; et al. Retinol saturase promotes adipogenesis and is downregulated in obesity. Proc. Natl. Acad. Sci. USA 2009, 106, 1105–1110. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.R.; Schulze, A. Lipid metabolism in cancer. FEBS J. 2012, 279, 2610–2623. [Google Scholar] [CrossRef]
- Yi, M.; Li, J.; Chen, S.; Cai, J.; Ban, Y.; Peng, Q.; Zhou, Y.; Zeng, Z.; Peng, S.; Li, X.; et al. Emerging role of lipid metabolism alterations in Cancer stem cells. J. Exp. Clin. Cancer Res. 2018, 37, 118. [Google Scholar] [CrossRef]
- Zhao, J.; Zhi, Z.; Wang, C.; Xing, H.; Song, G.; Yu, X.; Zhu, Y.; Wang, X.; Zhang, X.; Di, Y. Exogenous lipids promote the growth of breast cancer cells via CD36. Oncol. Rep. 2017, 38, 2105–2115. [Google Scholar] [CrossRef] [Green Version]
- Van der Paal, J.; Neyts, E.C.; Verlackt, C.C.W.; Bogaerts, A. Effect of lipid peroxidation on membrane permeability of cancer and normal cells subjected to oxidative stress. Chem. Sci. 2016, 7, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Rohrig, F.; Schulze, A. The multifaceted roles of fatty acid synthesis in cancer. Nat. Rev. Cancer 2016, 16, 732–749. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.A.; Xing, X.; Harris, J.W.; Zaytseva, Y.Y.; Mitov, M.I.; Napier, D.L.; Weiss, H.L.; Mark Evers, B.; Gao, T. Adipocytes activate mitochondrial fatty acid oxidation and autophagy to promote tumor growth in colon cancer. Cell Death Dis. 2017, 8, e2593. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Gao, Z.; Kolonin, M.G. Transient inflammatory signaling promotes beige adipogenesis. Sci. Signal 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.C.; Hsieh, L.L.; Tang, R.; Chang-Chieh, C.R.; Sung, F.C. MS-920: DNA repair gene polymorphisms, diet and colorectal cancer risk in Taiwan. Cancer Lett. 2005, 224, 279–288. [Google Scholar] [CrossRef]
- Cimprich, K.A.; Cortez, D. ATR: An essential regulator of genome integrity. Nat. Rev. Mol. Cell. Biol. 2008, 9, 616–627. [Google Scholar] [CrossRef] [Green Version]
- Tibbetts, R.S.; Brumbaugh, K.M.; Williams, J.M.; Sarkaria, J.N.; Cliby, W.A.; Shieh, S.Y.; Taya, Y.; Prives, C.; Abraham, R.T. A role for ATR in the DNA damage-induced phosphorylation of p53. Genes Dev. 1999, 13, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Santoro, R.; Marani, M.; Blandino, G.; Muti, P.; Strano, S. Melatonin triggers p53Ser phosphorylation and prevents DNA damage accumulation. Oncogene 2012, 31, 2931–2942. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.P.; Liu, F.; Wang, W. Two-phase dynamics of p53 in the DNA damage response. Proc. Natl. Acad. Sci. USA 2011, 108, 8990–8995. [Google Scholar] [CrossRef] [Green Version]
- Hartman, A.R.; Ford, J.M. BRCA1 induces DNA damage recognition factors and enhances nucleotide excision repair. Nat. Genet. 2002, 32, 180–184. [Google Scholar] [CrossRef]
- Glass-Holmes, M.; Aguilar, B.J.; Gragg, R.D., 3rd; Darling-Reed, S.; Goodman, C.B. Characterization of 2-amino-1-methyl-6-phenylimidazo[4,5b]pyridine at androgen receptor: Mechanistic support for its role in prostate cancer. Am. J. Cancer Res. 2015, 5, 191–200. [Google Scholar]
- Wang, R.; Dashwood, W.M.; Nian, H.; Lohr, C.V.; Fischer, K.A.; Tsuchiya, N.; Nakagama, H.; Ashktorab, H.; Dashwood, R.H. NADPH oxidase overexpression in human colon cancers and rat colon tumors induced by 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP). Int. J. Cancer 2011, 128, 2581–2590. [Google Scholar] [CrossRef] [PubMed]
- Ohgaki, H.; Takayama, S.; Sugimura, T. Carcinogenicities of heterocyclic amines in cooked food. Mutat. Res. 1991, 259, 399–410. [Google Scholar] [CrossRef]
- Schut, H.A.; Yao, R. Tea as a potential chemopreventive agent in PhIP carcinogenesis: Effects of green tea and black tea on PhIP-DNA adduct formation in female F-344 rats. Nutr. Cancer 2000, 36, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Glickman, B.W.; de Boer, J.G. Effect of conjugated linoleic acid on the formation of spontaneous and PhIP-induced mutation in the colon and cecum of rats. Mutat. Res. 2002, 500, 157–168. [Google Scholar] [CrossRef]
- Cheng, Z.N.; Shu, Y.; Liu, Z.Q.; Wang, L.S.; Ou-Yang, D.S.; Zhou, H.H. Role of cytochrome P450 in estradiol metabolism in vitro. Acta Pharmacol. Sin. 2001, 22, 148–154. [Google Scholar]
- Reed, L.; Arlt, V.M.; Phillips, D.H. The role of cytochrome P450 enzymes in carcinogen activation and detoxication: An in vivo-in vitro paradox. Carcinogenesis 2018, 39, 851–859. [Google Scholar] [CrossRef]
- Fotsis, T.; Zhang, Y.; Pepper, M.S.; Adlercreutz, H.; Montesano, R.; Nawroth, P.P.; Schweigerer, L. The endogenous oestrogen metabolite 2-methoxyoestradiol inhibits angiogenesis and suppresses tumour growth. Nature 1994, 368, 237–239. [Google Scholar] [CrossRef]
- Seegers, J.C.; Aveling, M.L.; Van Aswegen, C.H.; Cross, M.; Koch, F.; Joubert, W.S. The cytotoxic effects of estradiol-17 beta, catecholestradiols and methoxyestradiols on dividing MCF-7 and HeLa cells. J. Steroid. Biochem. 1989, 32, 797–809. [Google Scholar] [CrossRef]
- Blanck, A.; Hansson, T.; Eriksson, L.C.; Gustafsson, J.A. On mechanisms of sex differences in chemical carcinogenesis: Effects of implantation of ectopic pituitary grafts on the early stages of liver carcinogenesis in the rat. Carcinogenesis 1984, 5, 1257–1262. [Google Scholar] [CrossRef]
- Krais, A.M.; Speksnijder, E.N.; Melis, J.P.; Singh, R.; Caldwell, A.; Gamboa da Costa, G.; Luijten, M.; Phillips, D.H.; Arlt, V.M. Metabolic activation of 2-amino-1-methyl-6-phenylimidazo [4,5-b]pyridine and DNA adduct formation depends on p53: Studies in Trp53(+/+),Trp53(+/−) and Trp53(−/−) mice. Int. J. Cancer 2016, 138, 976–982. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Suzuki, R.; Kohno, H.; Sugie, S.; Takahashi, M.; Wakabayashi, K. Colonic adenocarcinomas rapidly induced by the combined treatment with 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine and dextran sodium sulfate in male ICR mice possess beta-catenin gene mutations and increases immunoreactivity for beta-catenin, cyclooxygenase-2 and inducible nitric oxide synthase. Carcinogenesis 2005, 26, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.W.; Hudson, N.; Seo, D.; Lee, S.; Khatri, B.; Lassiter, K.; Cook, D.; Piekarski, A.; Dridi, S.; Anthony, N.; et al. RNA sequencing for global gene expression associated with muscle growth in a single male modern broiler line compared to a foundational Barred Plymouth Rock chicken line. BMC Genom. 2017, 18, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Canonical Pathways 1 | Genes Up-Regulated | Genes Down-Regulated |
|---|---|---|
| M-PhIP vs. M-CON 2 | ||
| Oxidative Phosphorylation | Atp5g1, Atp5po, Cox17, Cox6b1, Cox7a1, Cyb5a, Atp6, Ndufa2, Ndufa3, Ndufa5, Ndufa8, Ndufa11, Ndufb7, Ndufs6, Ndufs7, Ndufv3 | - |
| Mitochondrial Dysfunction | Atp5g1, Atp5po, Cox17, Cox6b1, Cox7a1, Cyb5a, Fis1, Glrx2, Atp6, Ndufa2, Ndufa3, Ndufa5, Ndufa8, Ndufa11, Ndufb7, Ndufs6, Ndufs7, Ndufv3 | - |
| eIF2 Signaling | Eif3k, Map2k2, Rpl18, Rpl24, Rpl28, Rpl35, Rpl38, Rpl27a, Rpl37a, Rplp2, Rps11, Rps19, Rps20, Rps26 | Ago4, Eif2ak3, Pik3cb, Vegfa |
| Sirtuin Signaling Pathway | Atp5g1, Bax, Map1lc3a, Atp6, Ndufa2, Ndufa3, Ndufa5, Ndufa8, Ndufa11, Ndufb7, Ndufs6, Ndufs7, Ndufv3, Pam16, Sod1, Timm13, Tomm6 | Acss2, Ppargc1a |
| F-PhIP vs. F-CON | ||
| Triacylglycerol Degradation | Abhd6, Ddhd2, Lipe, Lpl | Aadac, Prdx6 |
| Retinol Biosynthesis | Ces1g, Ddhd2, Lipe, Lpl | Aadac |
| Adipogenesis Pathway | Cebpa, Fabp4, Fgfrl1, Lpl, Sap30 | Bmp2, Hif1a, Noct |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, J.H.; Cicalo, C.; Le, B.; Jeon, S.; Kim, S.; Hwang, K.A.; Kong, B.; Lee, J.H.; Kim, J.K. Colon Transcriptomics Reveals Sex-Dependent Metabolic Signatures in Response to 2-Amino-1-methyl-6-phenylimidazo[4,5-b]pyridine Treatment in C57BL/6N Mice. Int. J. Mol. Sci. 2020, 21, 6620. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186620
Pan JH, Cicalo C, Le B, Jeon S, Kim S, Hwang KA, Kong B, Lee JH, Kim JK. Colon Transcriptomics Reveals Sex-Dependent Metabolic Signatures in Response to 2-Amino-1-methyl-6-phenylimidazo[4,5-b]pyridine Treatment in C57BL/6N Mice. International Journal of Molecular Sciences. 2020; 21(18):6620. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186620
Chicago/Turabian StylePan, Jeong Hoon, Cara Cicalo, Brandy Le, Suwon Jeon, Sangyub Kim, Kyung A. Hwang, Byungwhi Kong, Jin Hyup Lee, and Jae Kyeom Kim. 2020. "Colon Transcriptomics Reveals Sex-Dependent Metabolic Signatures in Response to 2-Amino-1-methyl-6-phenylimidazo[4,5-b]pyridine Treatment in C57BL/6N Mice" International Journal of Molecular Sciences 21, no. 18: 6620. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186620