Tip60 Phosphorylation at Ser 99 Is Essential for Autophagy Induction in Bombyx mori


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
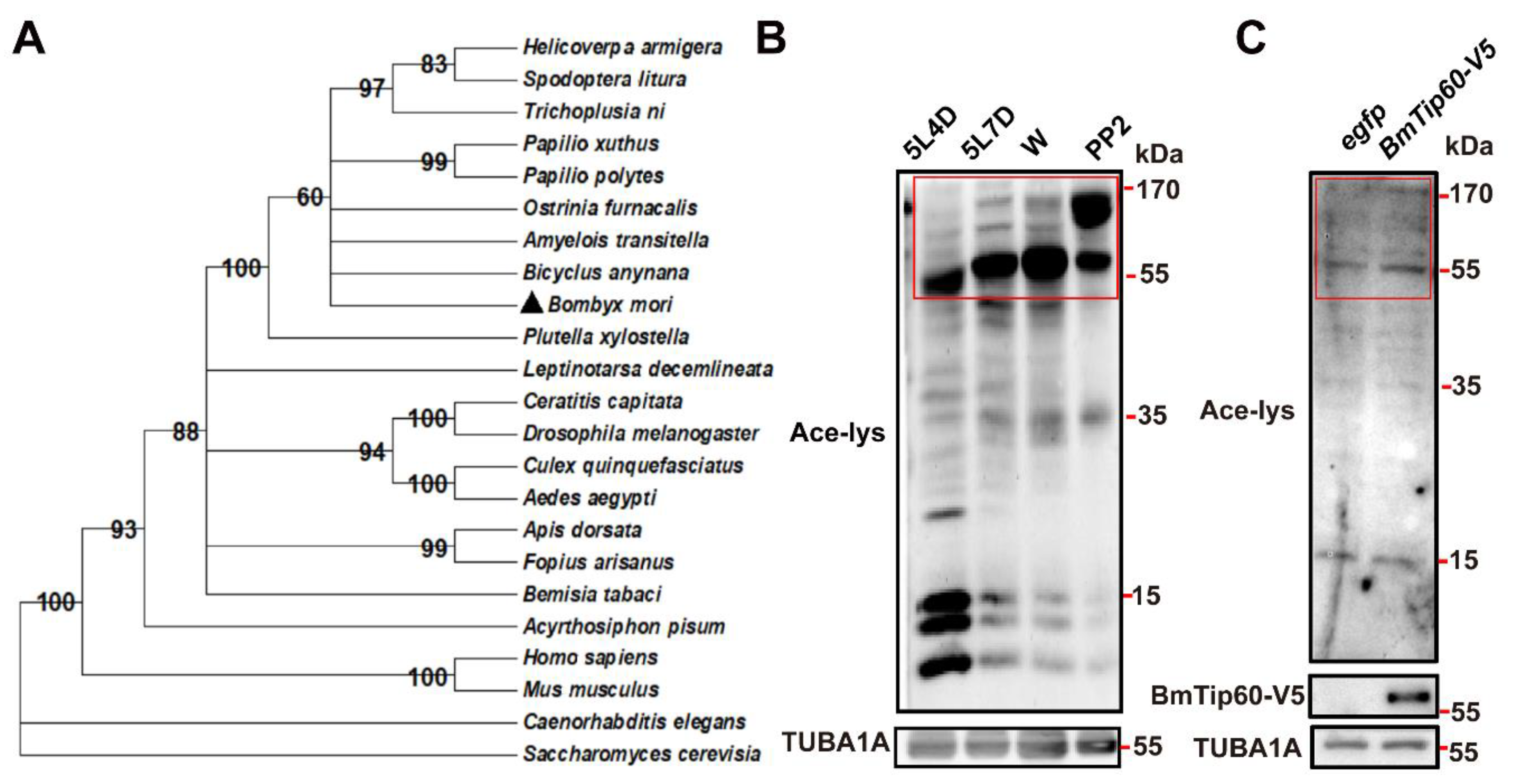
2.1. BmTip60 Is Highly Conserved from Insects to Mammals and Is Able to Catalyze Protein Acetylation
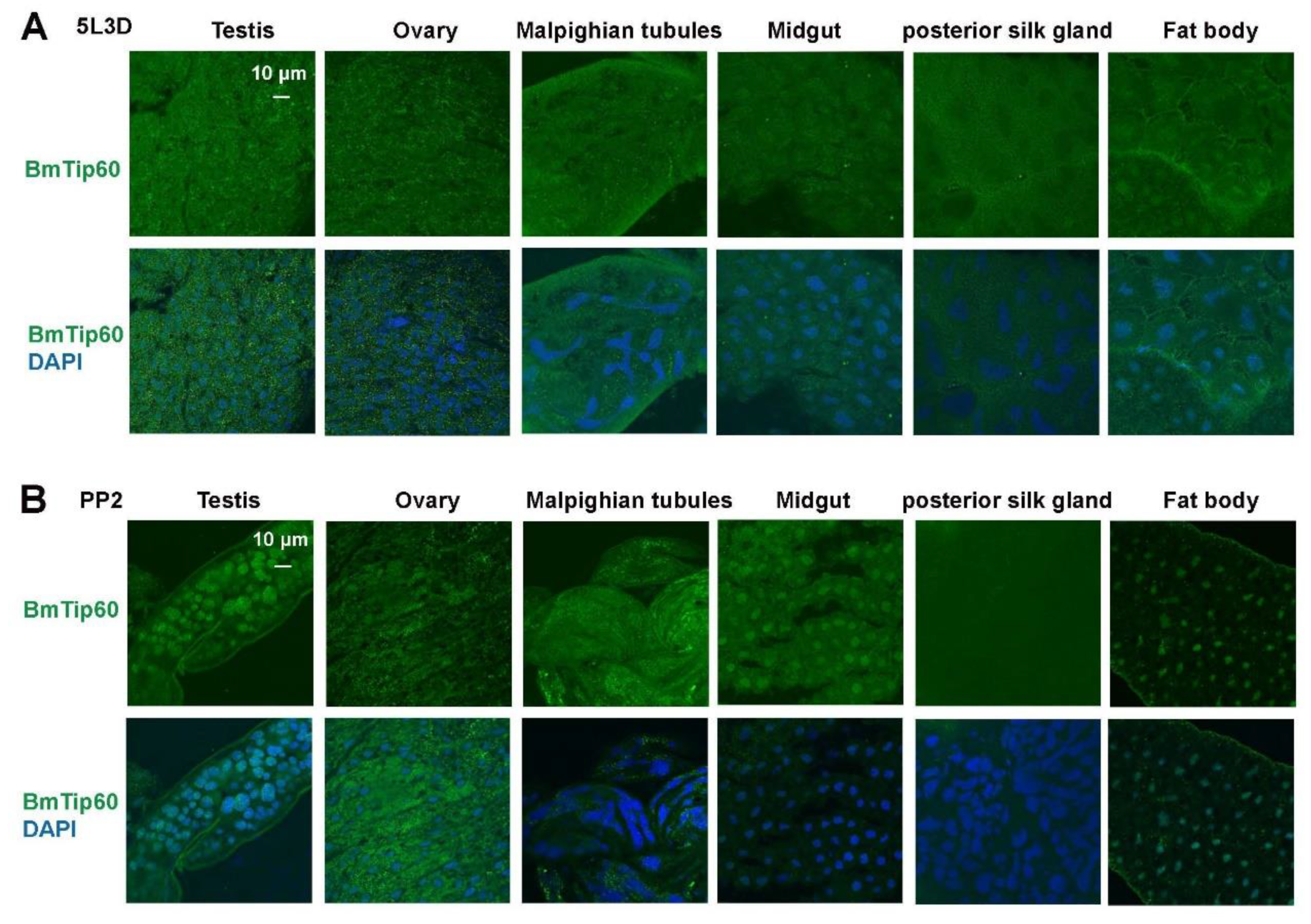
2.2. Tissue Distribution of BmTip60 in B. mori
2.3. Migration of BmTip60 Protein Band Is Consistent with Autophagy Occurrence in Fat Body
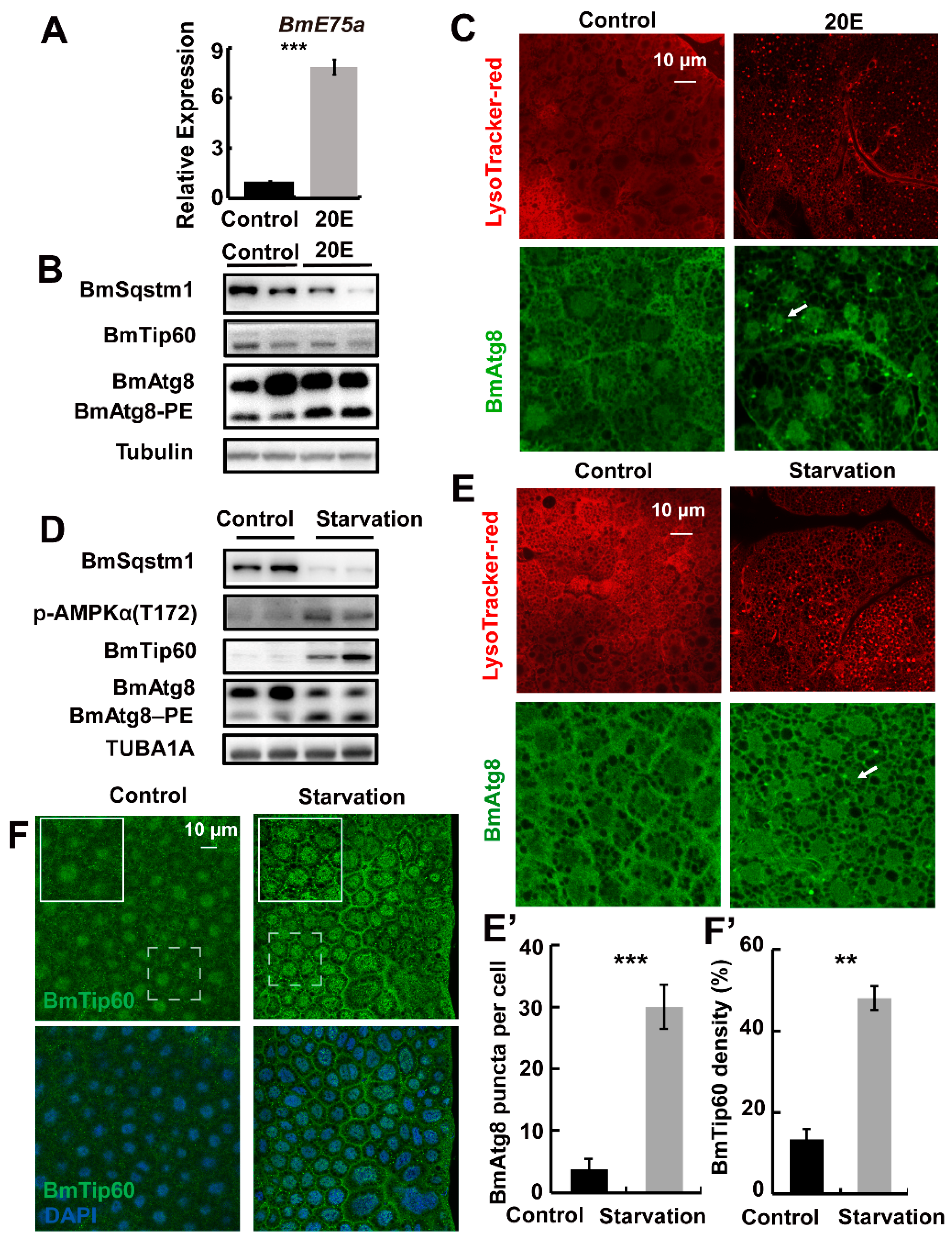
2.4. Starvation but Not 20E Promotes BmTip60 in Addition to Autophagy Induction in B. mori
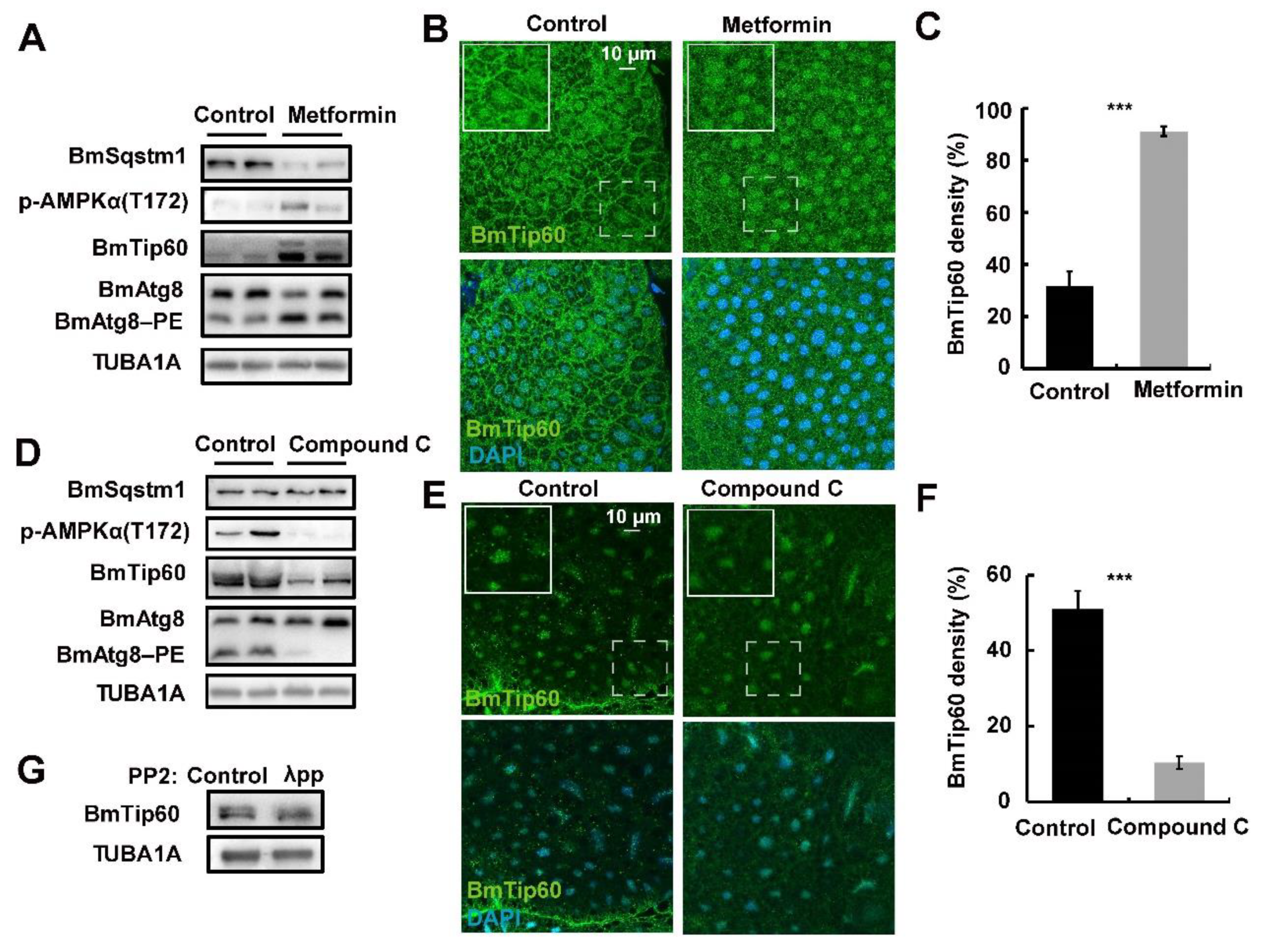
2.5. AMPK Activity Mediates the Migration of BmTip60 Protein
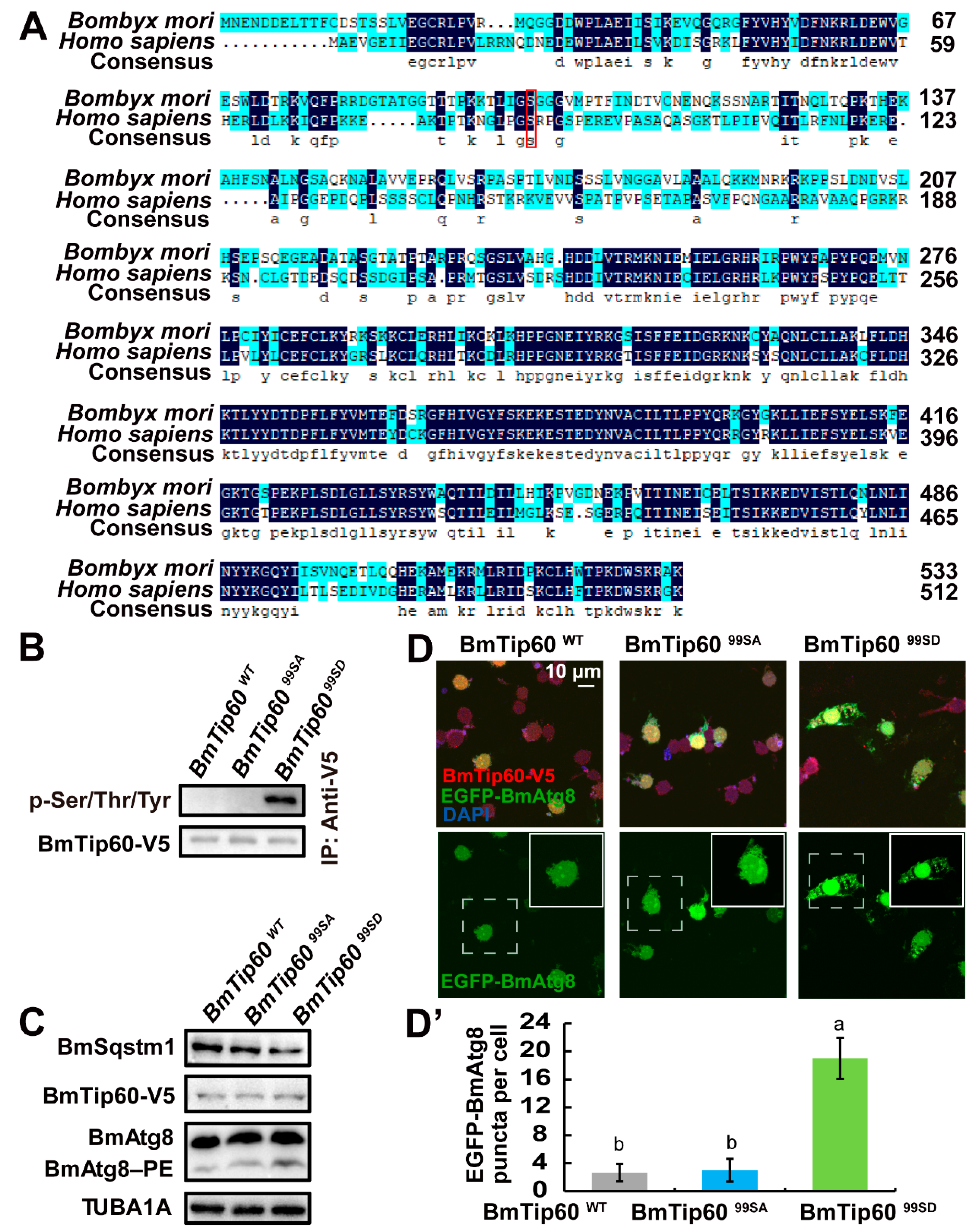
2.6. Ser99, an Evolutionarily Conserved Phosphorylation Site, Is Indispensable for BmTip60-Mediated Autophagy
3. Discussion
3.1. Phosphorylation of BmTip60 Is Regulated by AMPK Activity and Requisite for Autophagy Induction
3.2. 20E Functions Differently with Starvation in Regulating BmTip60
4. Materials and Methods
4.1. Silkworms and Cells
4.2. Plasmid Construction and Transfection
4.3. Starvation and 20E Treatment
4.4. Chemical Treatments
4.5. Immunoprecipitation and Western Blots
4.6. Immunofluorescent Staining
4.7. LysoTracker Staining
4.8. λ-Phosphatase Treatment of B. mori Fat Body Cells
4.9. Mutation of Ser99 in BmTip60
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 20E | 20-hydroxyecdysone |
| Tip60 | Tat-interactive protein, 60 KDa |
| Atg | Autophagy-related |
| AMPK | AMP-activated protein kinase |
| B. mori | Bombyx mori |
| BmSqstm1 | Bombyx sequestosome 1 |
| lys | Lysine |
| MTOR | mechanistic target of rapamycin kinase |
| PBS | Phosphate buffered saline |
| PTMs | Post-translational modifications |
| HATs | Histone acetyltransferases |
References
- Klionsky, D.J.; Abdelmohsen, K.; Abe, A.; Abedin, M.J.; Abeliovich, H.; Acevedo Arozena, A.; Adachi, H.; Adams, C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12, 1–222. [Google Scholar] [CrossRef] [Green Version]
- Levine, B.; Kroemer, G. Biological functions of autophagy genes: A disease perspective. Cell 2019, 176, 11–42. [Google Scholar] [CrossRef] [Green Version]
- Mariño, G.; Niso-Santano, M.; Baehrecke, E.H.; Kroemer, G. Self-consumption: The interplay of autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2014, 15, 81–94. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Klionsky, D.J. Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet. 2009, 43, 67–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.; Ma, L.; Guo, E.; Deng, X.; Ma, S.; Xia, Q.; Cao, Y.; Li, S. 20-Hydroxyecdysone upregulates Atg genes to induce autophagy in the Bombyx fat body. Autophagy 2013, 9, 1172–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Tian, L.; Guo, Z.; Guo, S.; Zhang, J.; Gu, S.H.; Palli, S.R.; Cao, Y.; Li, S. 20-Hydroxyecdysone (20E) Primary response gene E75 isoforms mediate steroidogenesis autoregulation and regulate developmental timing in Bombyx. J. Biol. Chem. 2016, 291, 18163–18175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, K.; Tian, L.; Guo, X.; Li, K.; Li, J.; Deng, X.; Li, Q.; Xia, Q.; Zhong, Y.; Huang, Z.; et al. BmATG5 and BmATG6 mediate apoptosis following autophagy induced by 20-hydroxyecdysone or starvation. Autophagy 2016, 12, 381–396. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Luo, M.; Li, K.; Dai, Y.; Yi, H.; Zhong, Y.; Cao, Y.; Tettamanti, G.; Tian, L. Cholesterol derivatives induce dephosphorylation of the histone deacetylases Rpd3/HDAC1 to upregulate autophagy. Autophagy 2020, 12, 1–17. [Google Scholar] [CrossRef]
- Popelka, H.; Klionsky, D.J. Post-translationally-modified structures in the autophagy machinery: An integrative perspective. FEBS J. 2015, 282, 3474–3488. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.H.; Finkel, T. Regulation of autophagy by the P300 acetyltransferase. J. Biol. Chem. 2009, 284, 6322–6328. [Google Scholar] [CrossRef] [Green Version]
- Bánréti, A.; Sass, M.; Graba, Y. The emerging role of acetylation in the regulation of autophagy. Autophagy 2013, 9, 819–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.H.; Cao, L.; Mostoslavsky, R.; Lombard, D.B.; Liu, J.; Bruns, N.E.; Tsokos, M.; Alt, F.W.; Finkel, T. A role for the NAD-dependent deacetylase Sirt1 in the regulation of autophagy. Proc. Natl. Acad. Sci. USA 2008, 105, 3374–3379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.P.; Zhou, D.; Ouyang, D.Y.; Xu, L.H.; Wang, Y.; Wang, L.X.; Pan, H.; He, X.H. LC3B-II deacetylation by histone deacetylase 6 is involved in serum-starvation-induced autophagic degradation. Biochem. Biophys. Res. Commun. 2013, 441, 970–975. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, E.; Shimaji, K.; Umegawachi, T.; Tomida, S.; Yoshida, H.; Yoshimoto, N.; Izawa, S.; Kimura, H.; Yamaguchi, M. The Histone deacetylase gene Rpd3 is required for starvation stress resistance. PLoS ONE 2016, 11, e0167554. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.A.; Bautista-Jimenez, R.; Denlinger, D.L. Changes in histone acetylation as potential mediators of pupal diapause in the flesh fly, Sarcophaga bullata. Insect Biochem. Mol. Biol. 2016, 76, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Jiang, F.; Yang, P.; Liu, Q.; Wang, X.; Kang, L. Characteristics and expression patterns of histone-modifying enzyme systems in the migratory locust. Insect Biochem. Mol. Biol. 2016, 76, 18–28. [Google Scholar] [CrossRef]
- Özbek, R.; Mukherjee, K.; Uçkan, F.; Vilcinskas, A. Reprograming of epigenetic mechanisms controlling host insect immunity and development in response to egg-laying by a parasitoid wasp. Proc. R. Soc. B 2020, 287, 20200704. [Google Scholar] [CrossRef]
- Narita, T.; Weinert, B.T.; Choudhary, C. Functions and mechanisms of non-histone protein acetylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 156–174. [Google Scholar] [CrossRef]
- Ikura, T.; Ogryzko, V.V.; Grigoriev, M.; Groisman, R.; Wang, J.; Horikoshi, M.; Scully, R.; Qin, J.; Nakatani, Y. Involvement of the TIP60 histone acetylase complex in DNA repair and apoptosis. Cell 2000, 102, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Utley, R.T.; Côté, J. The MYST family of histone acetyltransferases. Curr. Top Microbiol. Immunol. 2003, 274, 203–236. [Google Scholar]
- Squatrito, M.; Gorrini, C.; Amati, B. Tip60 in DNA damage response and growth control: Many tricks in one HAT. Trends Cell Biol. 2006, 16, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, J.; Wang, J.; Wang, R.; Liu, Z.; Yu, Y.; Lu, H. ING5 is a Tip60 cofactor that acetylates p53 in response to DNA damage. Cancer Res. 2013, 73, 3749–3760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.S.; Seo, J.; Choi, D.Y.; Lee, E.W.; Ko, A.; Ha, N.C.; Yoon, J.B.; Lee, H.W.; Kim, K.P.; Song, J. Stabilization of p21 (Cip1/WAF1) following Tip60-dependent acetylation is required for p21-mediated DNA damage response. Cell Death Differ. 2013, 20, 620–629. [Google Scholar] [CrossRef]
- Niida, H.; Katsuno, Y.; Sengoku, M.; Shimada, M.; Yukawa, M.; Ikura, M.; Ikura, T.; Kohno, K.; Shima, H.; Suzuki, H.; et al. Essential role of Tip60-dependent recruitment of ribonucleotide reductase at DNA damage sites in DNA repair during G1 phase. Genes Dev. 2010, 24, 333–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charvet, C.; Wissler, M.; Brauns-Schubert, P.; Wang, S.J.; Tang, Y.; Sigloch, F.C.; Mellert, H.; Brandenburg, M.; Lindner, S.E.; Breit, B.; et al. Phosphorylation of Tip60 by GSK-3 determines the induction of PUMA and apoptosis by p53. Mol. Cell 2011, 42, 584–596. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.Y.; Li, T.Y.; Liu, Q.; Zhang, C.; Li, X.; Chen, Y.; Zhang, S.M.; Lian, G.; Liu, Q.; Ruan, K.; et al. GSK3-TIP60-ULK1 signaling pathway links growth factor deprivation to autophagy. Science 2012, 336, 477–481. [Google Scholar] [CrossRef]
- Cheng, X.; Ma, X.; Zhu, Q.; Song, D.; Ding, X.; Li, L.; Jiang, X.; Wang, X.; Tian, R.; Su, H.; et al. Pacer is a mediator of mTORC1 and GSK3-TIP60 signaling in regulation of autophagosome maturation and lipid metabolism. Mol. Cell 2019, 73, 788–802. [Google Scholar] [CrossRef] [Green Version]
- Libbrecht, R.; Nadrau, D.; Foitzik, S. A role of histone acetylation in the regulation of circadian rhythm in ants. iScience 2020, 23, 100846. [Google Scholar] [CrossRef] [Green Version]
- Ri, H.; Lee, J.; Sonn, J.Y.; Yoo, E.; Lim, C.; Choe, J. Drosophila CrebB is a substrate of the nonsense-mediated mRNA decay pathway that sustains circadian behaviors. Cells 2019, 42, 301–312. [Google Scholar]
- Mizushima, N. The role of the Atg1/ULK1 complex in autophagy regulation. Curr. Opin. Cell Biol. 2010, 22, 132–139. [Google Scholar] [CrossRef]
- Sengupta, S.; Peterson, T.R.; Sabatini, D.M. Regulation of the mTOR complex 1 pathway by nutrients, growth factors, and stress. Mol. Cell 2010, 40, 310–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neufeld, T.P. TOR-dependent control of autophagy: Biting the hand that feeds. Curr. Opin. Cell Biol. 2010, 22, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, D.; Zhou, S.; Liu, S.; Li, K.; Zhao, H.; Long, S.; Liu, H.; Xie, Y.; Su, Y.; Yu, F.; et al. The AMPK-PP2A axis in insect fat body is activated by 20-hydroxyecdysone to antagonize insulin/IGF signaling and restrict growth rate. Proc. Natl. Acad. Sci. USA 2020, 117, 9292–9301. [Google Scholar] [CrossRef]
- Li, S.; Xu, J.; Xu, X.; Ma, W.; Tian, L.; Li, K. Functional identification of Bombyx mori Atg13 in autophagy. Arch. Insect Biochem. Physiol. 2020, 9, e21718. [Google Scholar]
- Chang, Y.Y.; Neufeld, T.P. Autophagy takes flight in Drosophila. FEBS Lett. 2010, 584, 1342–1349. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Li, K.; Wu, W.; Wu, K.; Yi, H.; Li, W.; Xiao, Y.; Zhong, Y.; Cao, Y.; Tian, L. Steroid hormone 20-hydroxyecdysone induces the transcription and complex assembly of V-ATPases to facilitate autophagy in Bombyx mori. Insect Biochem. Mol. Biol. 2020, 116, 103255. [Google Scholar] [CrossRef]
- Guo, S.Y.; Wu, W.M.; Li, S.Y.; Liu, Y.; Ruan, Z.F.; Ye, M.Q.; Xiao, Y.; Zhong, Y.J.; Cao, Y.; Li, K.; et al. 20-Hydroxyecdysone-upregulated proteases involved in Bombyx larval fat body destruction. Insect Mol. Biol. 2018, 27, 724–738. [Google Scholar] [CrossRef]
- Lu, F.; Wei, Z.; Luo, Y.; Guo, H.; Zhang, G.; Xia, Q.; Wang, Y. SilkDB 3.0: Visualizing and exploring multiple levels of data for silkworm. Nucleic Acids Res. 2020, 48, 749–755. [Google Scholar] [CrossRef] [Green Version]
- Brian, R.K.; Suleyman, V.; Sanjit, P.; Alex, B.; Chittibabu, G. ngLOC: Software and web server for predicting protein subcellular localization in prokaryotes and eukaryotes. BMC Res. Notes 2012, 5, 351. [Google Scholar]
- Zhou, S.; Zhou, Q.; Liu, Y.; Wang, S.; Wen, D.; He, Q.; Wang, W.; Bendena, W.G.; Li, S. Two Tor genes in the silkworm Bombyx mori. Insect Mol. Biol. 2010, 19, 727–735. [Google Scholar] [CrossRef]
- Yi, C.; Ma, M.; Ran, L.; Zheng, J.; Tong, J.; Zhu, J.; Ma, C.; Sun, Y.; Zhang, S.; Feng, W.; et al. Function and molecular mechanism of acetylation in autophagy regulation. Science 2012, 336, 474–477. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, A.; Deiulio, A.; Martin, E.T.; Upadhyay, M.; Rangana, P. Tip60 complex promotes expression of a differentiation factor to regulate germline differentiation in female Drosophila. Mol. Biol. Cell 2018, 29, 2933–2945. [Google Scholar] [CrossRef] [PubMed]
- Tettamanti, G.; Casartelli, M. Cell death during complete metamorphosis. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2019, 374, 20190065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Dai, F.; Guo, E.; Li, K.; Ma, L.; Tian, L.; Cao, Y.; Zhang, G.; Palli, S.R.; Li, S. 20-Hydroxyecdysone (20E) Primary response gene E93 modulates 20E signaling to promote Bombyx larval-pupal metamorphosis. J. Biol. Chem. 2015, 290, 27370–27383. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, S.; Liu, H.; Wang, J.; Zhou, S.; Jiang, R.J.; Bendena, W.G.; Li, S. 20-hydroxyecdysone reduces insect food consumption resulting in fat body lipolysis during molting and pupation. J. Mol. Cell Biol. 2010, 2, 128–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to image j: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Li, K.; Zhao, H.; Xu, X.; Xu, J.; Luo, M.; Xiao, Y.; Tian, L. Tip60 Phosphorylation at Ser 99 Is Essential for Autophagy Induction in Bombyx mori. Int. J. Mol. Sci. 2020, 21, 6893. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186893
Wu W, Li K, Zhao H, Xu X, Xu J, Luo M, Xiao Y, Tian L. Tip60 Phosphorylation at Ser 99 Is Essential for Autophagy Induction in Bombyx mori. International Journal of Molecular Sciences. 2020; 21(18):6893. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186893
Chicago/Turabian StyleWu, Wenmei, Kang Li, Haigang Zhao, Xianying Xu, Jing Xu, Man Luo, Yang Xiao, and Ling Tian. 2020. "Tip60 Phosphorylation at Ser 99 Is Essential for Autophagy Induction in Bombyx mori" International Journal of Molecular Sciences 21, no. 18: 6893. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186893