Role of Nuclear Claudin-4 in Renal Cell Carcinoma

 , and
, and
Abstract
:1. Introduction
2. Results
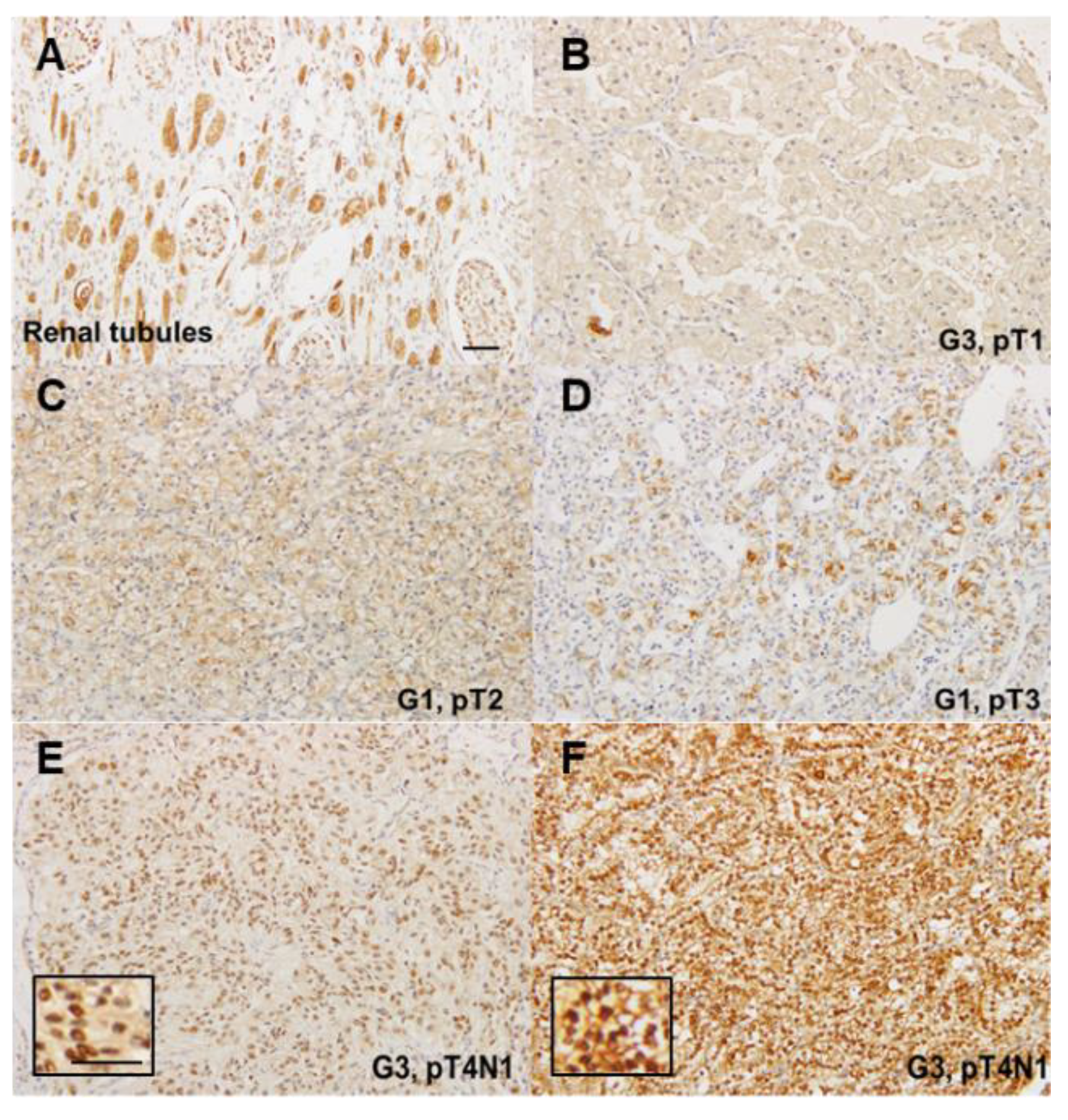
2.1. Expression of CLDN4 in Renal Cell Carcinomas
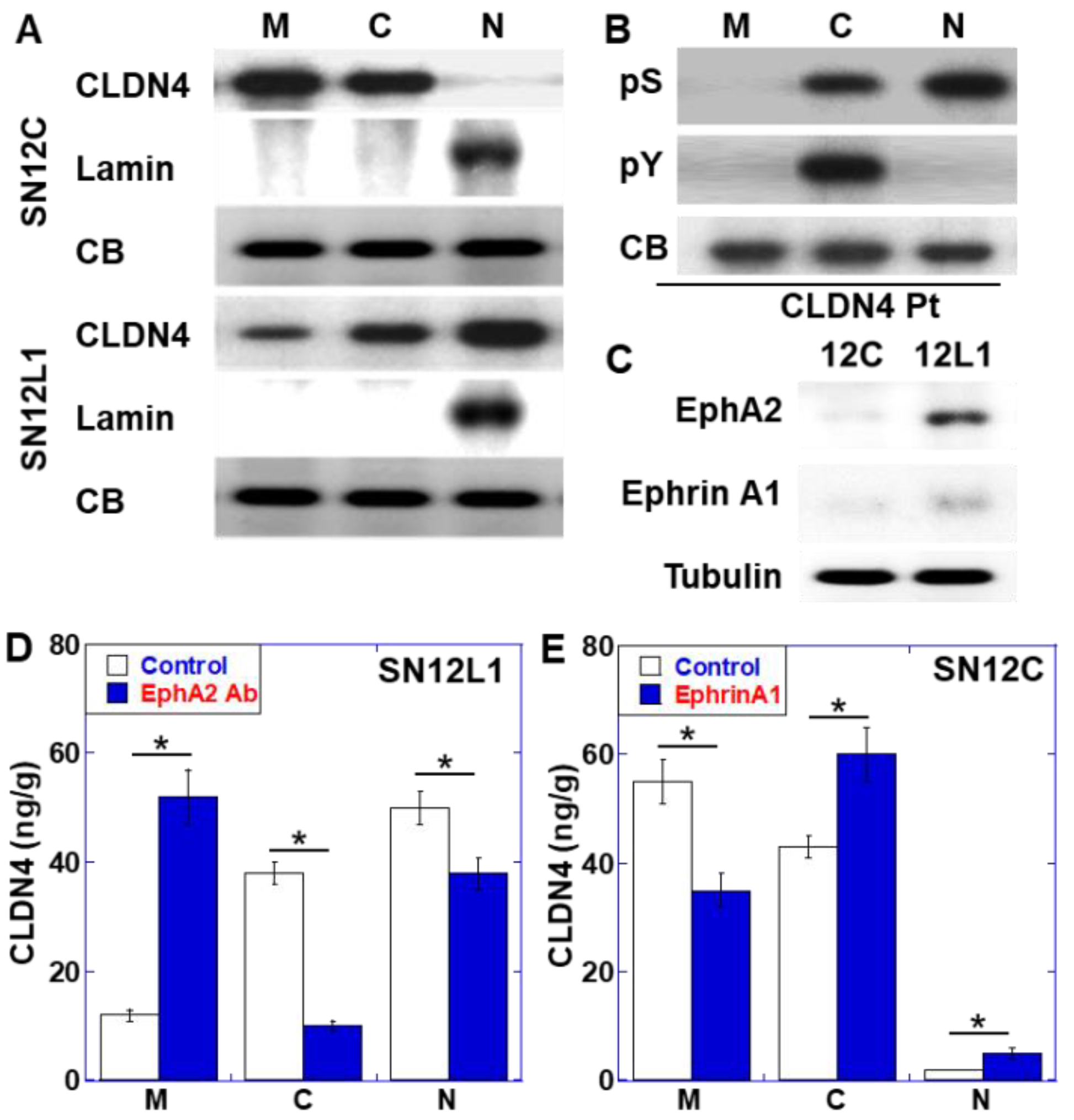
2.2. Role of EphA2 in CLDN4 Cytoplasmic Translocation
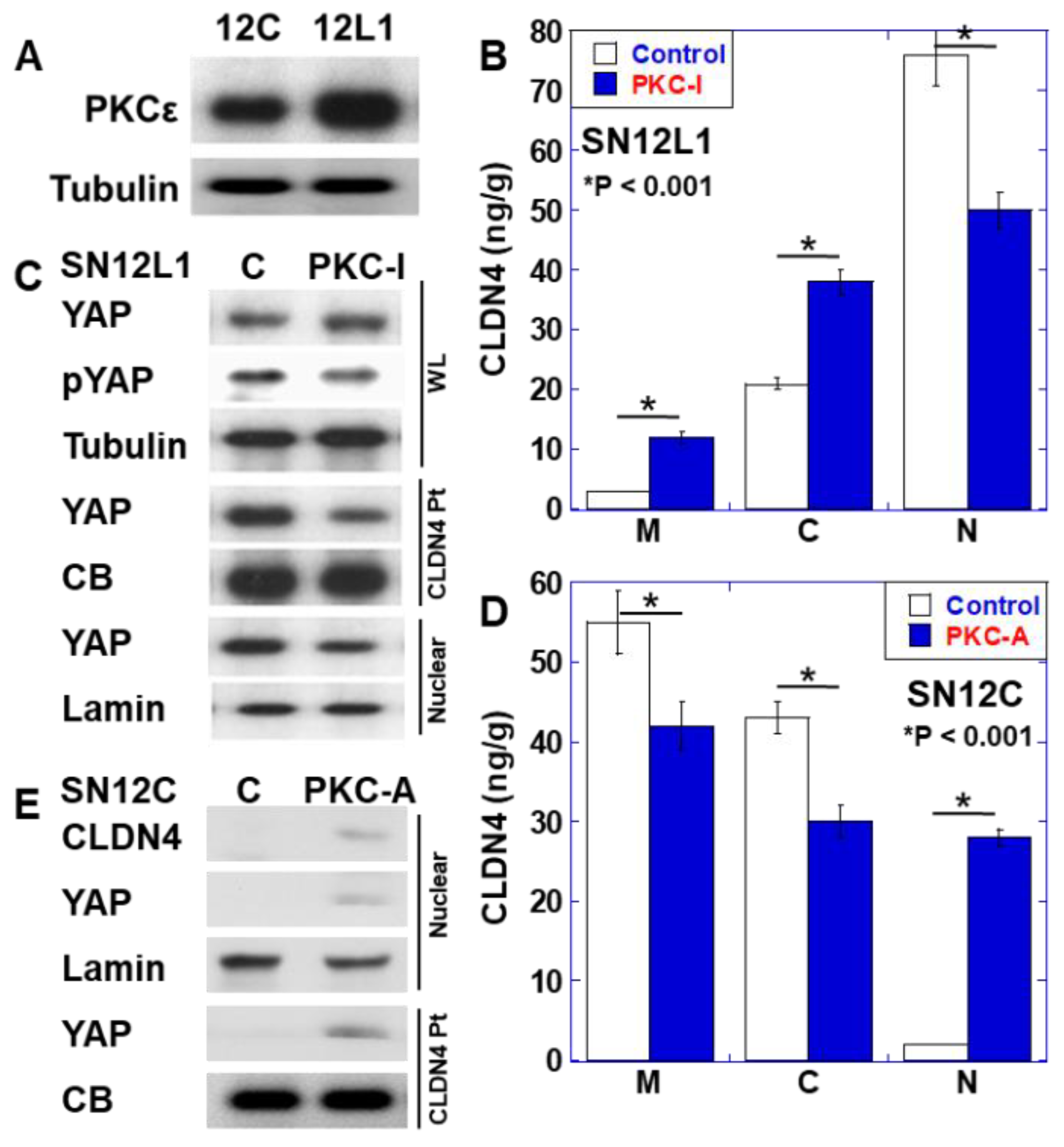
2.3. Role of PKCε in CLDN4 Nuclear Translocation and YAP Activation
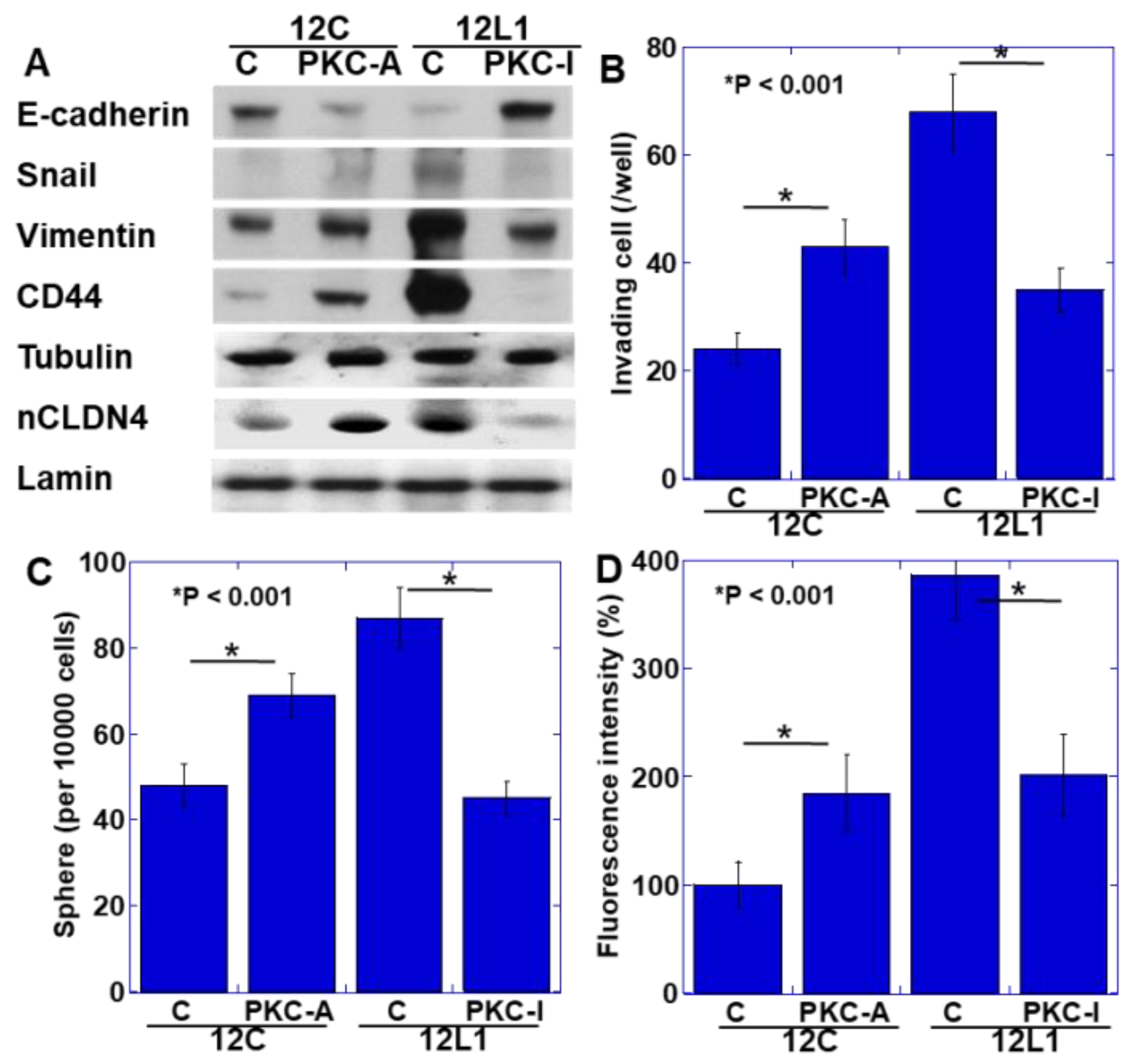
2.4. YAP Activation and Induction of the EMT Phenotype
3. Discussion
4. Materials and Methods
4.1. Surgical Specimens
4.2. Cell Lines
4.3. Sphere Assay
4.4. Antibody and Reagents
4.5. Immunohistochemistry
4.6. Protein Extraction
4.7. Immunoblot Analysis
4.8. Immunoprecipitation
4.9. Enzyme-Linked Immunosorbent Assay (ELISA) and Colorimetric Assay
4.10. Animals
4.11. Lung Metastasis Model
4.12. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviation
| RCC | renal cell carcinoma; |
| CCC | clear cell renal cell carcinoma |
| HIF | hypoxia-inducible factor |
| VHL | von Hippel-Lindau |
| EMT | epithelial-mesenchymal transition |
| CLDN | claudin |
| CPE | Clostridium perfringens enterotoxin |
| YAP | Yes-associated protein |
| PKC | protein kinase C |
References
- Tamaki, T.; Dong, Y.; Ohno, Y.; Sobue, T.; Nishimoto, H.; Shibata, A. The burden of rare cancer in Japan: Application of the RARECARE definition. Cancer Epidemiol. 2014, 38, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Motzer, R.J.; Bander, N.H.; Nanus, D.M. Renal-cell carcinoma. N. Engl. J. Med. 1996, 335, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Buchner, A.; Riesenberg, R.; Kotter, I.; Hofstetter, A.; Stief, C.; Oberneder, R. Frequency and prognostic relevance of disseminated tumor cells in bone marrow of patients with metastatic renal cell carcinoma. Cancer 2006, 106, 1514–1520. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.J.; Le, V.H.; Oyama, T.; Ricketts, C.J.; Ho, T.H.; Cheng, E.H. Chromosome 3p Loss-Orchestrated VHL, HIF, and Epigenetic Deregulation in Clear Cell Renal Cell Carcinoma. J. Clin. Oncol. 2018, 36, Jco2018792549. [Google Scholar] [CrossRef]
- Braga, E.A.; Fridman, M.V.; Loginov, V.I.; Dmitriev, A.A.; Morozov, S.G. Molecular Mechanisms in Clear Cell Renal Cell Carcinoma: Role of miRNAs and Hypermethylated miRNA Genes in Crucial Oncogenic Pathways and Processes. Front. Genet. 2019, 10, 320. [Google Scholar] [CrossRef] [PubMed]
- Rankin, E.B.; Fuh, K.C.; Castellini, L.; Viswanathan, K.; Finger, E.C.; Diep, A.N.; LaGory, E.L.; Kariolis, M.S.; Chan, A.; Lindgren, D.; et al. Direct regulation of GAS6/AXL signaling by HIF promotes renal metastasis through SRC and MET. Proc. Natl. Acad. Sci. USA 2014, 111, 13373–13378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukita, S.; Furuse, M.; Itoh, M. Multifunctional strands in tight junctions. Nat. Rev. Mol. Cell Biol. 2001, 2, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Turksen, K.; Troy, T.C. Junctions gone bad: Claudins and loss of the barrier in cancer. Biochim. Biophys. Acta 2011, 1816, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Shang, X.; Manorek, G.; Howell, S.B. Regulation of the Epithelial-Mesenchymal Transition by Claudin-3 and Claudin-4. PLoS ONE 2013, 8, e67496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredriksson, K.; Van Itallie, C.M.; Aponte, A.; Gucek, M.; Tietgens, A.J.; Anderson, J.M. Proteomic analysis of proteins surrounding occludin and claudin-4 reveals their proximity to signaling and trafficking networks. PLoS ONE 2015, 10, e0117074. [Google Scholar] [CrossRef]
- Li, W.; Xu, C.; Wang, K.; Ding, Y.; Ding, L. Non-tight junction-related function of claudin-7 in interacting with integrinβ1 to suppress colorectal cancer cell proliferation and migration. Cancer Manag. Res. 2019, 11, 1443–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Kim, D.H.; Fan, J.; Lu, Q.; Verbanac, K.; Ding, L.; Renegar, R.; Chen, Y.H. A non-tight junction function of claudin-7-Interaction with integrin signaling in suppressing lung cancer cell proliferation and detachment. Mol. Cancer 2015, 14, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiguchi, Y.; Fujiwara-Tani, R.; Sasaki, T.; Luo, Y.; Ohmori, H.; Kishi, S.; Mori, S.; Goto, K.; Yasui, W.; Sho, M.; et al. Targeting claudin-4 enhances CDDP-chemosensitivity in gastric cancer. Oncotarget 2019, 10, 2189–2202. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, C.; Yamamoto, K.; Kishi, S.; Sasaki, T.; Ohmori, H.; Fujiwara-Tani, R.; Mori, S.; Kawahara, I.; Nishiguchi, Y.; Mori, T.; et al. Clostridium perfringens enterotoxin induces claudin-4 to activate YAP in oral squamous cell carcinomas. Oncotarget 2020, 11, 309–321. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara-Tani, R.; Fujii, K.; Mori, S.; Kishi, S.; Sasaki, T.; Ohmori, H.; Nakashima, C.; Kawahara, I.; Nishiguchi, Y.; Mori, T.; et al. Role of Clostridium perfringens Enterotoxin on YAP Activation in Colonic Sessile Serrated Adenoma/ Polyps with Dysplasia. Int. J. Mol. Sci. 2020, 21, 3840. [Google Scholar] [CrossRef]
- Sasaki, T.; Mori, S.; Kishi, S.; Fujiwara-Tani, R.; Ohmori, H.; Nishiguchi, Y.; Hojo, Y.; Kawahara, I.; Nakashima, C.; Fujii, K.; et al. Effect of Proton Pump Inhibitors on Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 3877. [Google Scholar] [CrossRef] [PubMed]
- Sobin, L.H.; Wittekind, C. UICC TNM Classification of Malignant Tumours, 6th ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2003. [Google Scholar]
- Tanaka, M.; Kamata, R.; Sakai, R. EphA2 phosphorylates the cytoplasmic tail of Claudin-4 and mediates paracellular permeability. J. Biol. Chem. 2005, 280, 42375–42382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, T.; Indig, F.E.; Morin, P.J. Phosphorylation of claudin-4 by PKCepsilon regulates tight junction barrier function in ovarian cancer cells. Exp. Cell Res. 2007, 313, 3364–3375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwada, M.; Chihara, Y.; Luo, Y.; Li, X.; Nishiguchi, Y.; Fujiwara, R.; Sasaki, T.; Fujii, K.; Ohmori, H.; Fujimoto, K.; et al. Pro-chemotherapeutic effects of antibody against extracellular domain of claudin-4 in bladder cancer. Cancer Lett. 2015, 369, 212–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara-Tani, R.; Sasaki, T.; Luo, Y.; Goto, K.; Kawahara, I.; Nishiguchi, Y.; Kishi, S.; Mori, S.; Ohmori, H.; Kondoh, M.; et al. Anti-claudin-4 extracellular domain antibody enhances the antitumoral effects of chemotherapeutic and antibody drugs in colorectal cancer. Oncotarget 2018, 9, 37367–37378. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Fujiwara-Tani, R.; Kishi, S.; Mori, S.; Luo, Y.; Ohmori, H.; Goto, K.; Nishiguchi, Y.; Mori, T.; Sho, M.; et al. Targeting claudin-4 enhances chemosensitivity of pancreatic ductal carcinomas. Cancer Med. 2019, 8, 6700–6708. [Google Scholar] [CrossRef]
- Luo, Y.; Kishi, S.; Sasaki, T.; Ohmori, H.; Fujiwara-Tani, R.; Mori, S.; Goto, K.; Nishiguchi, Y.; Mori, T.; Kawahara, I.; et al. Targeting claudin-4 enhances chemosensitivity in breast cancer. Cancer Sci. 2020, 111, 1840–1850. [Google Scholar] [CrossRef] [Green Version]
- Gossage, L.; Eisen, T.; Maher, E.R. VHL, the story of a tumour suppressor gene. Nat. Rev. Cancer 2015, 15, 55–64. [Google Scholar] [CrossRef]
- Harten, S.K.; Shukla, D.; Barod, R.; Hergovich, A.; Balda, M.S.; Matter, K.; Esteban, M.A.; Maxwell, P.H. Regulation of renal epithelial tight junctions by the von Hippel-Lindau tumor suppressor gene involves occludin and claudin 1 and is independent of E-cadherin. Mol. Biol. Cell 2009, 20, 1089–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajduković, J. HIF-1--a big chapter in the cancer tale. Exp. Oncol. 2016, 38, 9–12. [Google Scholar] [CrossRef]
- Virman, J.; Soini, Y.; Kujala, P.; Luukkaala, T.; Salminen, T.; Sunela, K.; Kellokumpu-Lehtinen, P.L. Claudins as prognostic factors for renal cell cancer. Anticancer Res. 2014, 34, 4181–4187. [Google Scholar]
- Men, W.; Martin, T.A.; Ruge, F.; Zhang, N.; Du, P.; Yang, Y.; Jiang, W.G. Expression of claudins in human clear cell renal cell carcinoma. Cancer Genom. Proteom. 2015, 12, 1–8. [Google Scholar]
- Fritzsche, F.R.; Oelrich, B.; Johannsen, M.; Kristiansen, I.; Moch, H.; Jung, K.; Kristiansen, G. Claudin-1 protein expression is a prognostic marker of patient survival in renal cell carcinomas. Clin. Cancer Res. 2008, 14, 7035–7042. [Google Scholar] [CrossRef] [Green Version]
- Lechpammer, M.; Resnick, M.B.; Sabo, E.; Yakirevich, E.; Greaves, W.O.; Sciandra, K.T.; Tavares, R.; Noble, L.C.; DeLellis, R.A.; Wang, L.J. The diagnostic and prognostic utility of claudin expression in renal cell neoplasms. Mod. Pathol. 2008, 21, 1320–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, P.; Saha, T.; Sheikh, I.A.; Chakraborty, S.; Aoun, J.; Chakrabarti, M.K.; Rajendran, V.M.; Ameen, N.A.; Dutta, S.; Hoque, K.M. Zinc ameliorates intestinal barrier dysfunctions in shigellosis by reinstating claudin-2 and -4 on the membranes. Am. J. Physiol. Gastrointest Liver Physiol. 2019, 316, G229–G246. [Google Scholar] [CrossRef]
- Dhawan, P.; Singh, A.B.; Deane, N.G.; No, Y.; Shiou, S.R.; Schmidt, C.; Neff, J.; Washington, M.K.; Beauchamp, R.D. Claudin-1 regulates cellular transformation and metastatic behavior in colon cancer. J. Clin. Investig. 2005, 115, 1765–1776. [Google Scholar] [CrossRef] [Green Version]
- McCole, D.F. Phosphatase regulation of intercellular junctions. Tissue Barriers 2013, 1, e26713. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.C.; Cho, S.Y.; Yoon, C.Y.; Lee, S.B.; Kwak, C.; Kim, H.H.; Jeong, H. EphA2 Is a Potential Player of Malignant Cellular Behavior in Non-Metastatic Renal Cell Carcinoma Cells but Not in Metastatic Renal Cell Carcinoma Cells. PLoS ONE 2015, 10, e0130975. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, J.; Cui, L.; Zhang, H.; Zhang, S.; Bai, Y. High EphA2 protein expression in renal cell carcinoma is associated with a poor disease outcome. Oncol. Lett. 2014, 8, 687–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrem, C.J.; Tatsumi, T.; Olson, K.S.; Shirai, K.; Finke, J.H.; Bukowski, R.M.; Zhou, M.; Richmond, A.L.; Derweesh, I.; Kinch, M.S.; et al. Expression of EphA2 is prognostic of disease-free interval and overall survival in surgically treated patients with renal cell carcinoma. Clin. Cancer Res. 2005, 11, 226–231. [Google Scholar]
- Fattet, L.; Jung, H.Y.; Matsumoto, M.W.; Aubol, B.E.; Kumar, A.; Adams, J.A.; Chen, A.C.; Sah, R.L.; Engler, A.J.; Pasquale, E.B.; et al. Matrix Rigidity Controls Epithelial-Mesenchymal Plasticity and Tumor Metastasis via a Mechanoresponsive EPHA2/LYN Complex. Dev. Cell 2020, 54, 302–316. [Google Scholar] [CrossRef]
- French, A.D.; Fiori, J.L.; Camilli, T.C.; Leotlela, P.D.; O’Connell, M.P.; Frank, B.P.; Subaran, S.; Indig, F.E.; Taub, D.D.; Weeraratna, A.T. PKC and PKA phosphorylation affect the subcellular localization of claudin-1 in melanoma cells. Int. J. Med. Sci. 2009, 6, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Cao, K.; Li, X.; Guo, S.; Mao, X.; Wang, Z.; Zhuang, J.; Pan, J.; Mo, C.; Chen, J.; et al. The expression and role of protein kinase C (PKC) epsilon in clear cell renal cell carcinoma. J. Exp. Clin. Cancer Res. 2011, 30, 88. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Fu, S.J.; Fan, W.Z.; Wang, Z.H.; Chen, Z.B.; Guo, S.J.; Chen, J.X.; Qiu, S.P. PKCε inhibits isolation and stemness of side population cells via the suppression of ABCB1 transporter and PI3K/Akt, MAPK/ERK signaling in renal cell carcinoma cell line 769P. Cancer Lett. 2016, 376, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Brenner, W.; Benzing, F.; Gudejko-Thiel, J.; Fischer, R.; Färber, G.; Hengstler, J.G.; Seliger, B.; Thüroff, J.W. Regulation of beta1 integrin expression by PKCepsilon in renal cancer cells. Int. J. Oncol. 2004, 25, 1157–1163. [Google Scholar] [PubMed]
- Aziz, M.H.; Manoharan, H.T.; Sand, J.M.; Verma, A.K. Protein kinase Cepsilon interacts with Stat3 and regulates its activation that is essential for the development of skin cancer. Mol. Carcinog. 2007, 46, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Giaccia, A.J.; Strulovici, B.; Brown, J.M. Differential expression of protein kinase C epsilon protein in lung cancer cell lines by ionising radiation. Br. J. Cancer 1992, 66, 844–849. [Google Scholar] [CrossRef]
- Acs, P.; Szallasi, Z.; Kazanietz, M.G.; Blumberg, P.M. Differential activation of PKC isozymes by 14–3-3 zeta protein. Biochem. Biophys. Res. Commun. 1995, 216, 103–109. [Google Scholar] [CrossRef]
- Masui, O.; White, N.M.; DeSouza, L.V.; Krakovska, O.; Matta, A.; Metias, S.; Khalil, B.; Romaschin, A.D.; Honey, R.J.; Stewart, R.; et al. Quantitative proteomic analysis in metastatic renal cell carcinoma reveals a unique set of proteins with potential prognostic significance. Mol. Cell Proteom. 2013, 12, 132–144. [Google Scholar] [CrossRef] [Green Version]
- Anzai, H.; Kitadai, Y.; Bucana, C.D.; Sanchez, R.; Omoto, R.; Fidler, I.J. Intratumoral heterogeneity and inverse correlation between expression of E-cadherin and collagenase type IV in human gastric carcinomas. Differentiation 1996, 60, 119–127. [Google Scholar] [CrossRef]
- Naito, S.; von Eschenbach, A.C.; Fidler, I.J. Different growth pattern and biologic behavior of human renal cell carcinoma implanted into different organs of nude mice. J. Natl. Cancer Inst. 1987, 78, 377–385. [Google Scholar]
- Kuniyasu, H.; Oue, N.; Wakikawa, A.; Shigeishi, H.; Matsutani, N.; Kuraoka, K.; Ito, R.; Yokozaki, H.; Yasui, W. Expression of receptors for advanced glycation end-products (RAGE) is closely associated with the invasive and metastatic activity of gastric cancer. J. Pathol. 2002, 196, 163–170. [Google Scholar] [CrossRef]
- Kuniyasu, H.; Yano, S.; Sasaki, T.; Sasahira, T.; Sone, S.; Ohmori, H. Colon cancer cell-derived high mobility group 1/amphoterin induces growth inhibition and apoptosis in macrophages. Am. J. Pathol. 2005, 166, 751–760. [Google Scholar] [CrossRef] [Green Version]
- Matsushima-Otsuka, S.; Fujiwara-Tani, R.; Sasaki, T.; Ohmori, H.; Nakashima, C.; Kishi, S.; Nishiguchi, Y.; Fujii, K.; Luo, Y.; Kuniyasu, H. Significance of intranuclear angiotensin-II type 2 receptor in oral squamous cell carcinoma. Oncotarget 2018, 9, 36561–36574. [Google Scholar] [CrossRef] [Green Version]
- Kuniyasu, H.; Yasui, W.; Pettaway, C.A.; Yano, S.; Oue, N.; Tahara, E.; Fidler, I.J. Interferon-alpha prevents selection of doxorubicin-resistant undifferentiated- androgen-insensitive metastatic human prostate cancer cells. Prostate 2001, 49, 19–29. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter 1 | n | Expression of CLDN4 2 | p3 | |
|---|---|---|---|---|
| Age | −60 y | 52 | 53 ± 8 | NS |
| 61 y - | 150 | 56 ± 6 | ||
| Sex | Male | 111 | 56 ± 9 | NS |
| Female | 91 | 54 ± 7 | ||
| Histology | Clear cell | 200 | 55 ± 9 | NS |
| Papillary | 2 | 52 ± 10 | ||
| Grade | 1 | 161 | 52 ± 7 | NS |
| 2 | 34 | 59 ± 6 | ||
| 3 | 7 | 41 ± 7 | ||
| pT | 1 | 145 | 50 ± 6 | NS |
| 2 | 46 | 64 ± 7 | ||
| 3 | 11 | 49 ± 7 | ||
| pN | 0 | 192 | 57 ± 8 | NS |
| 1 | 10 | 55 ± 8 | ||
| Stage | I | 146 | 49 ± 6 | NS |
| II | 44 | 68 ± 8 | ||
| III-IV | 12 | 54 ± 7 |
| CLDN4 | n | pT3 1) | p2) | |
|---|---|---|---|---|
| Nuclear | 5 | 5 | (100%) | |
| Non-nuclear | 197 | 6 | (3%) | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Owari, T.; Sasaki, T.; Fujii, K.; Fujiwara-Tani, R.; Kishi, S.; Mori, S.; Mori, T.; Goto, K.; Kawahara, I.; Nakai, Y.; et al. Role of Nuclear Claudin-4 in Renal Cell Carcinoma. Int. J. Mol. Sci. 2020, 21, 8340. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218340
Owari T, Sasaki T, Fujii K, Fujiwara-Tani R, Kishi S, Mori S, Mori T, Goto K, Kawahara I, Nakai Y, et al. Role of Nuclear Claudin-4 in Renal Cell Carcinoma. International Journal of Molecular Sciences. 2020; 21(21):8340. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218340
Chicago/Turabian StyleOwari, Takuya, Takamitsu Sasaki, Kiyomu Fujii, Rina Fujiwara-Tani, Shingo Kishi, Shiori Mori, Takuya Mori, Kei Goto, Isao Kawahara, Yasushi Nakai, and et al. 2020. "Role of Nuclear Claudin-4 in Renal Cell Carcinoma" International Journal of Molecular Sciences 21, no. 21: 8340. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218340