A Novel Selective Sphingosine Kinase 2 Inhibitor, HWG-35D, Ameliorates the Severity of Imiquimod-Induced Psoriasis Model by Blocking Th17 Differentiation of Naïve CD4 T Lymphocytes

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Neither SK1 nor DES1 Protein Levels Were Altered by a Novel Selective SK2 Inhibitor, HWG-35D
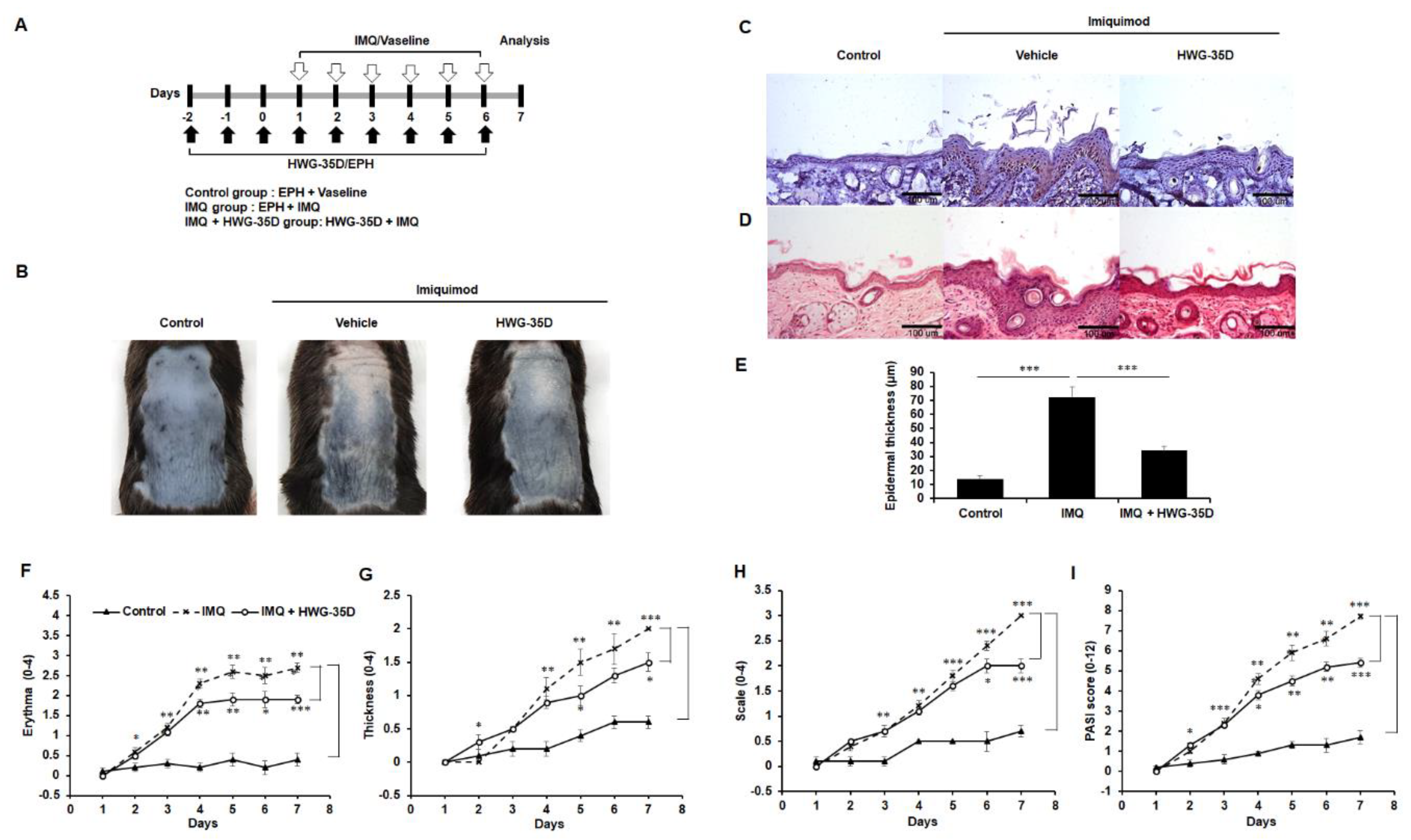
2.2. Selective SK2 Inhibition Using HWG-35D Improves Severity of Psoriasis-Like Skin Lesions in a Murine Model of Psoriasis With the Dosage of 25 µG/40 µL Surfactant
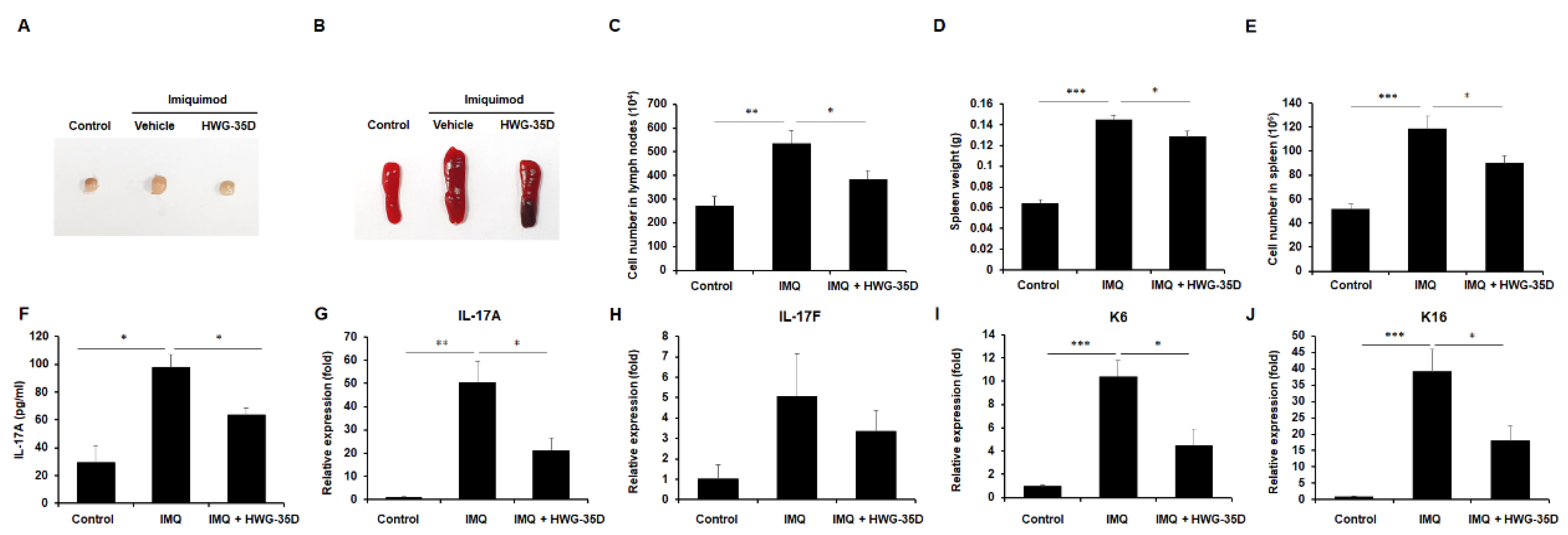
2.3. Systemic Immune Reaction Caused by IMQ Was Diminished With Topical Application of HWG-35D
2.4. Topical HWG-35D Application Normalises mRNA Levels of Genes Associated With Th17 Response and Keratinization
2.5. Inhibition of SK2 With HWG-35D Blocks Th17 Polarization In Vitro
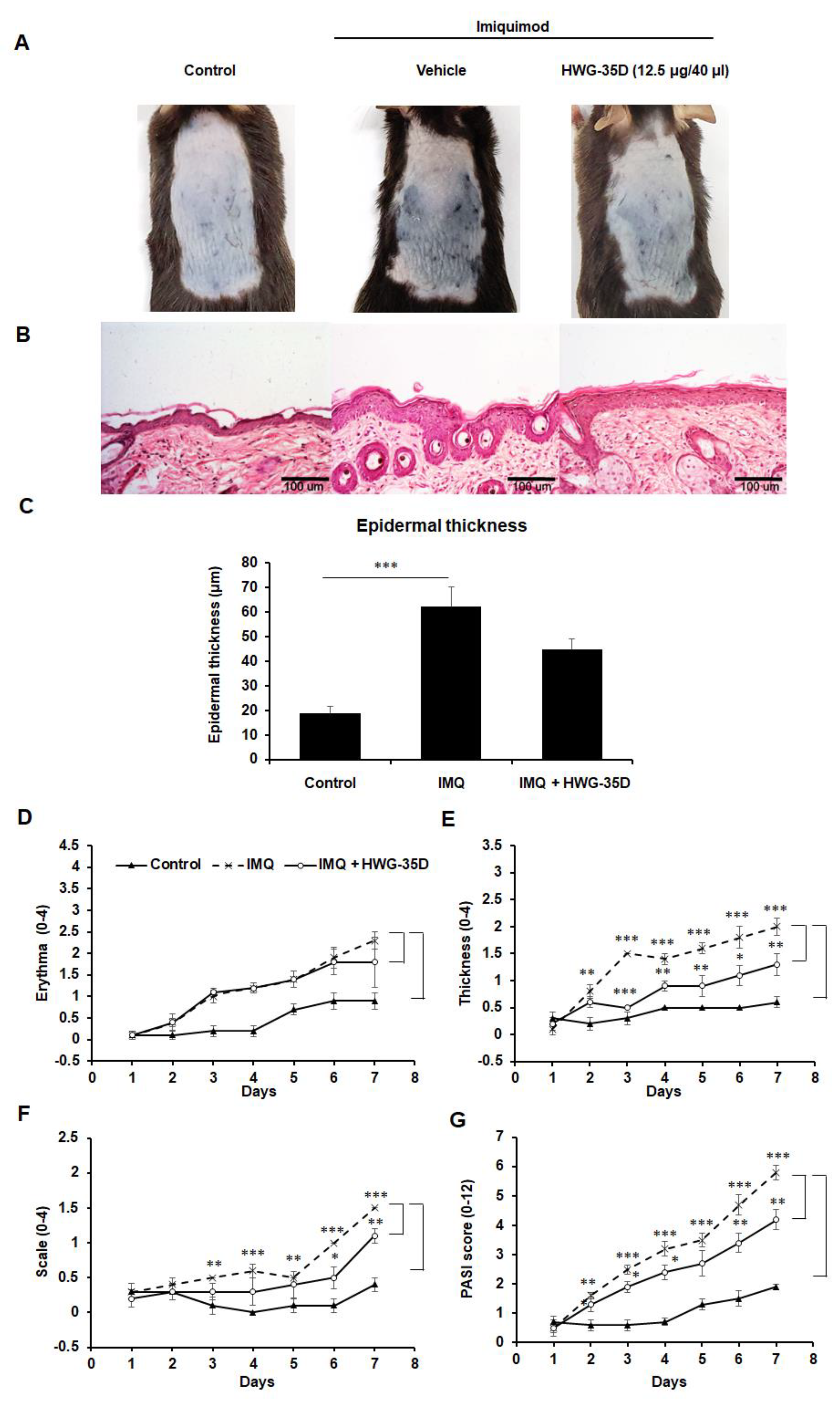
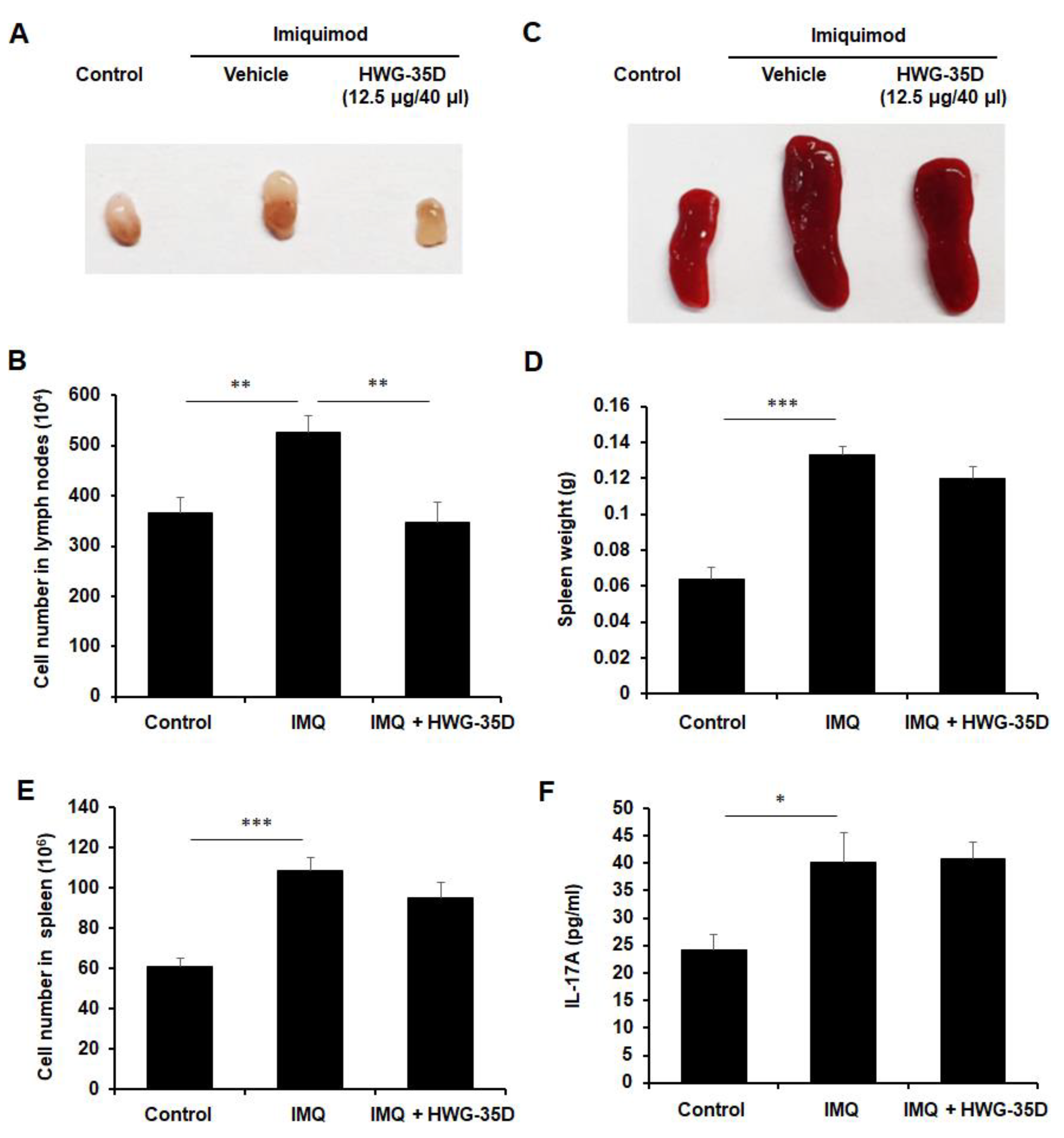
2.6. The Protective Effects of Topical HWG-35D Application in Psoriasis-Like Skin Disease Were Investigated With the Dosage of 12.5 µG/40 µL Surfactant
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Animal Experiments
4.4. Scoring the Severity of Psoriasis-Like Skin Lesion
4.5. Histological Evaluation and Immunohistochemistry
4.6. Enzyme-Linked Immunosorbent Assay
4.7. Real-Time Polymerase Chain Reaction
4.8. Western Blotting
4.9. In Vitro Induction of T Cell Differentiation
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DES1 | Dihydroceramide desaturase 1 |
| ELISA | Enzyme-linked immunosorbent assay |
| EPH | Ethanol, propylene glycol and H2O |
| H & E | Hematoxylin and eosin |
| IFN | Interferon |
| IL | Interleukin |
| IMQ | Imiquimod |
| PASI | Psoriasis Area and Severity Index |
| SK | Sphingosine kinase |
| S1P | Sphingosine-1-phosphate |
| SOCS | Suppressor of cytokine signaling |
| STAT3 | Signal transducer and activator of transcription |
| Th | T-helper |
| TNF-α | Tumor necrosis factor-α |
References
- Osier, E.; Wang, A.S.; Tollefson, M.M.; Cordoro, K.M.; Daniels, S.R.; Eichenfield, A.; Gelfand, J.M.; Gottlieb, A.B.; Kimball, A.B.; Lebwohl, M.; et al. Pediatric Psoriasis Comorbidity Screening Guidelines. JAMA Dermatol 2017, 153, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Eberle, F.C.; Bruck, J.; Holstein, J.; Hirahara, K.; Ghoreschi, K. Recent advances in understanding psoriasis. F1000Res 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, M.; Ghoreschi, K. The role of IL-4 in psoriasis. Expert. Rev. Clin. Immunol. 2017, 13, 171–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, S.; Nakashima, A.; Shima, T.; Ito, M. Th1/Th2/Th17 and regulatory T-cell paradigm in pregnancy. Am. J. Reprod. Immunol. 2010, 63, 601–610. [Google Scholar] [CrossRef]
- Ouyang, W.; Kolls, J.K.; Zheng, Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity 2008, 28, 454–467. [Google Scholar] [CrossRef] [Green Version]
- Marinoni, B.; Ceribelli, A.; Massarotti, M.S.; Selmi, C. The Th17 axis in psoriatic disease: Pathogenetic and therapeutic implications. Auto Immun Highlights 2014, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.P.; Ruether, A.; Stuart, P.E.; Jenisch, S.; Tejasvi, T.; Hiremagalore, R.; Schreiber, S.; Kabelitz, D.; Lim, H.W.; Voorhees, J.J.; et al. Polymorphisms of the IL12B and IL23R genes are associated with psoriasis. J. Invest. Dermatol. 2008, 128, 1653–1661. [Google Scholar] [CrossRef] [Green Version]
- Tsoi, L.C.; Spain, S.L.; Knight, J.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; Ding, J.; Li, Y.; Tejasvi, T.; Gudjonsson, J.E.; et al. Identification of 15 new psoriasis susceptibility loci highlights the role of innate immunity. Nat. Genet. 2012, 44, 1341–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lande, R.; Botti, E.; Jandus, C.; Dojcinovic, D.; Fanelli, G.; Conrad, C.; Chamilos, G.; Feldmeyer, L.; Marinari, B.; Chon, S.; et al. The antimicrobial peptide LL37 is a T-cell autoantigen in psoriasis. Nat. Commun. 2014, 5, 5621. [Google Scholar] [CrossRef]
- Carreira, A.C.; Santos, T.C.; Lone, M.A.; Zupancic, E.; Lloyd-Evans, E.; de Almeida, R.F.M.; Hornemann, T.; Silva, L.C. Mammalian sphingoid bases: Biophysical, physiological and pathological properties. Prog. Lipid. Res. 2019, 75, 100988. [Google Scholar] [CrossRef]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Pyne, N.J.; Adams, D.R.; Pyne, S. Sphingosine Kinase 2 in Autoimmune/Inflammatory Disease and the Development of Sphingosine Kinase 2 Inhibitors. Trends. Pharmacol. Sci. 2017, 38, 581–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattenberg, B.W. Role of sphingosine kinase localization in sphingolipid signaling. World. J. Biol. Chem. 2010, 1, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, H.A.; Pitson, S.M. Roles, regulation and inhibitors of sphingosine kinase 2. FEBS J 2013, 280, 5317–5336. [Google Scholar] [CrossRef] [PubMed]
- Antoon, J.W.; White, M.D.; Meacham, W.D.; Slaughter, E.M.; Muir, S.E.; Elliott, S.; Rhodes, L.V.; Ashe, H.B.; Wiese, T.E.; Smith, C.D.; et al. Antiestrogenic effects of the novel sphingosine kinase-2 inhibitor ABC294640. Endocrinology 2010, 151, 5124–5135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsanafi, M.; Kelly, S.L.; Jubair, K.; McNaughton, M.; Tate, R.J.; Merrill, A.H., Jr.; Pyne, S.; Pyne, N.J. Native and Polyubiquitinated Forms of Dihydroceramide Desaturase Are Differentially Linked to Human Embryonic Kidney Cell Survival. Mol. Cell Biol. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.R.; Tawati, S.; Berretta, G.; Rivas, P.L.; Baiget, J.; Jiang, Z.; Alsfouk, A.; Mackay, S.P.; Pyne, N.J.; Pyne, S. Topographical Mapping of Isoform-Selectivity Determinants for J-Channel-Binding Inhibitors of Sphingosine Kinases 1 and 2. J Med. Chem. 2019, 62, 3658–3676. [Google Scholar] [CrossRef] [Green Version]
- Maceyka, M.; Harikumar, K.B.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate signaling and its role in disease. Trends Cell Biol. 2012, 22, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Mysliwiec, H.; Baran, A.; Harasim-Symbor, E.; Choromanska, B.; Mysliwiec, P.; Milewska, A.J.; Chabowski, A.; Flisiak, I. Increase in circulating sphingosine-1-phosphate and decrease in ceramide levels in psoriatic patients. Arch. Dermatol. Res. 2017, 309, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Checa, A.; Xu, N.; Sar, D.G.; Haeggström, J.Z.; Ståhle, M.; Wheelock, C.E. Circulating levels of sphingosine-1-phosphate are elevated in severe, but not mild psoriasis and are unresponsive to anti-TNF-α treatment. Scientific Reports 2015, 5, 12017. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.H.; Cho, K.A.; Hahn, S.; Lee, Y.; Kim, Y.H.; Woo, S.Y.; Ryu, K.H.; Park, W.J.; Park, J.W. Inhibiting Sphingosine Kinase 2 Derived-sphingosine-1-phosphate Ameliorates Psoriasis-like Skin Disease via Blocking Th17 Differentiation of Naive CD4 T Lymphocytes in Mice. Acta. Derm. Venereol. 2019, 99, 594–601. [Google Scholar] [CrossRef] [Green Version]
- Ji, M.; Xue, N.; Lai, F.; Zhang, X.; Zhang, S.; Wang, Y.; Jin, J.; Chen, X. Validating a Selective S1P1 Receptor Modulator Syl930 for Psoriasis Treatment. Biol. Pharm. Bull. 2018, 41, 592–596. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Xue, N.; Liu, Y.; Fu, R.; Wang, M.; Ji, M.; Lai, F.; Hu, J.; Wang, X.; Xiao, Q.; et al. A novel S1P1 modulator IMMH002 ameliorates psoriasis in multiple animal models. Acta. Pharm. Sin. B. 2020, 10, 276–288. [Google Scholar] [CrossRef]
- McNaughton, M.; Pitman, M.; Pitson, S.M.; Pyne, N.J.; Pyne, S. Proteasomal degradation of sphingosine kinase 1 and inhibition of dihydroceramide desaturase by the sphingosine kinase inhibitors, SKi or ABC294640, induces growth arrest in androgen-independent LNCaP-AI prostate cancer cells. Oncotarget 2016, 7, 16663–16675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, K.G.; Sun, C.; Bittman, R.; Pyne, N.J.; Pyne, S. (R)-FTY720 methyl ether is a specific sphingosine kinase 2 inhibitor: Effect on sphingosine kinase 2 expression in HEK 293 cells and actin rearrangement and survival of MCF-7 breast cancer cells. Cell Signal. 2011, 23, 1590–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Fits, L.; Mourits, S.; Voerman, J.S.; Kant, M.; Boon, L.; Laman, J.D.; Cornelissen, F.; Mus, A.M.; Florencia, E.; Prens, E.P.; et al. Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J. Immunol. 2009, 182, 5836–5845. [Google Scholar] [CrossRef] [PubMed]
- Lowes, M.A.; Suarez-Farinas, M.; Krueger, J.G. Immunology of psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef] [Green Version]
- Brembilla, N.C.; Senra, L.; Boehncke, W.H. The IL-17 Family of Cytokines in Psoriasis: IL-17A and Beyond. Front. Immunol. 2018, 9, 1682. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yin, M.; Zhang, L.J. Keratin 6, 16 and 17-Critical Barrier Alarmin Molecules in Skin Wounds and Psoriasis. Cells 2019, 8, 807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, R.A.; Eichner, R.; Sun, T.T. Monoclonal antibody analysis of keratin expression in epidermal diseases: A 48- and 56-kdalton keratin as molecular markers for hyperproliferative keratinocytes. J. Cell Biol. 1984, 98, 1397–1406. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, A.; Suzuki, M.; Sakaguchi, R.; Hanada, T.; Yasukawa, H. SOCS, Inflammation, and Autoimmunity. Front. Immunol. 2012, 3, 20. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Ichiyama, K.; Hashimoto, M.; Yoshida, H.; Takimoto, T.; Takaesu, G.; Torisu, T.; Hanada, T.; Yasukawa, H.; Fukuyama, S.; et al. Loss of suppressor of cytokine signaling 1 in helper T cells leads to defective Th17 differentiation by enhancing antagonistic effects of IFN-gamma on STAT3 and Smads. J. Immunol. 2008, 180, 3746–3756. [Google Scholar] [CrossRef] [Green Version]
- Zaba, L.C.; Krueger, J.G.; Lowes, M.A. Resident and “inflammatory” dendritic cells in human skin. J. Invest. Dermatol. 2009, 129, 302–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barra, G.; Lepore, A.; Gagliardi, M.; Somma, D.; Matarazzo, M.R.; Costabile, F.; Pasquale, G.; Mazzoni, A.; Gallo, C.; Nuzzo, G.; et al. Sphingosine Kinases promote IL-17 expression in human T lymphocytes. Sci. Rep. 2018, 8, 13233. [Google Scholar] [CrossRef]
- Cingolani, F.; Casasampere, M.; Sanllehi, P.; Casas, J.; Bujons, J.; Fabrias, G. Inhibition of dihydroceramide desaturase activity by the sphingosine kinase inhibitor SKI II. J. Lipid. Res. 2014, 55, 1711–1720. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.; Xin, C.; Pfeilschifter, J.; Huwiler, A. A novel mode of action of the putative sphingosine kinase inhibitor 2-(p-hydroxyanilino)-4-(p-chlorophenyl) thiazole (SKI II): Induction of lysosomal sphingosine kinase 1 degradation. Cell Physiol. Biochem. 2010, 26, 97–104. [Google Scholar] [CrossRef]
- Powell, J.A.; Pitman, M.R.; Zebol, J.R.; Moretti, P.A.B.; Neubauer, H.A.; Davies, L.T.; Lewis, A.C.; Dagley, L.F.; Webb, A.I.; Costabile, M.; et al. Kelch-like protein 5-mediated ubiquitination of lysine 183 promotes proteasomal degradation of sphingosine kinase 1. Biochem. J. 2019, 476, 3211–3226. [Google Scholar] [CrossRef]
- Cai, Y.; Fleming, C.; Yan, J. New insights of T cells in the pathogenesis of psoriasis. Cell Mol. Immunol. 2012, 9, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Diani, M.; Altomare, G.; Reali, E. T Helper Cell Subsets in Clinical Manifestations of Psoriasis. J. Immunol. Res. 2016, 2016, 7692024. [Google Scholar] [CrossRef] [Green Version]
- Schlaak, J.F.; Buslau, M.; Jochum, W.; Hermann, E.; Girndt, M.; Gallati, H.; Meyer zum Buschenfelde, K.H.; Fleischer, B. T cells involved in psoriasis vulgaris belong to the Th1 subset. J. Invest. Dermatol. 1994, 102, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, E.; Sato, Y.; Minagawa, A.; Okuyama, R. Pathogenesis of psoriasis and development of treatment. J. Dermatol. 2018, 45, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Detmar, M.; Orfanos, C.E. Tumor necrosis factor-alpha inhibits cell proliferation and induces class II antigens and cell adhesion molecules in cultured normal human keratinocytes in vitro. Arch. Dermatol. Res. 1990, 282, 238–245. [Google Scholar] [CrossRef]
- de Oliveira, P.S.; Cardoso, P.R.; Lima, E.V.; Pereira, M.C.; Duarte, A.L.; Pitta Ida, R.; Rego, M.J.; Pitta, M.G. IL-17A, IL-22, IL-6, and IL-21 Serum Levels in Plaque-Type Psoriasis in Brazilian Patients. Mediators Inflamm 2015, 2015, 819149. [Google Scholar] [CrossRef] [Green Version]
- Silfvast-Kaiser, A.; Paek, S.Y.; Menter, A. Anti-IL17 therapies for psoriasis. Expert Opin Biol Ther 2019, 19, 45–54. [Google Scholar] [CrossRef]
- Newcomb, D.C.; Cephus, J.Y.; Boswell, M.G.; Fahrenholz, J.M.; Langley, E.W.; Feldman, A.S.; Zhou, W.; Dulek, D.E.; Goleniewska, K.; Woodward, K.B.; et al. Estrogen and progesterone decrease let-7f microRNA expression and increase IL-23/IL-23 receptor signaling and IL-17A production in patients with severe asthma. J. Allergy. Clin. Immunol. 2015, 136, 1025–1034.e11. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.Y.; Fan, Y.M.; Zhang, Q.; Liu, S.; Li, Q.; Ke, G.L.; Li, C.; You, Z. Estradiol inhibits Th17 cell differentiation through inhibition of RORgammaT transcription by recruiting the ERalpha/REA complex to estrogen response elements of the RORgammaT promoter. J. Immunol. 2015, 194, 4019–4028. [Google Scholar] [CrossRef] [Green Version]
- Fuseini, H.; Cephus, J.Y.; Wu, P.; Davis, J.B.; Contreras, D.C.; Gandhi, V.D.; Rathmell, J.C.; Newcomb, D.C. ERalpha Signaling Increased IL-17A Production in Th17 Cells by Upregulating IL-23R Expression, Mitochondrial Respiration, and Proliferation. Front. Immunol. 2019, 10, 2740. [Google Scholar] [CrossRef] [Green Version]
- Sarac, G.; Koca, T.T.; Baglan, T. A brief summary of clinical types of psoriasis. North. Clin. Istanb. 2016, 3, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Lee, K.M.; Ucmak, D.; Brodsky, M.; Atanelov, Z.; Farahnik, B.; Abrouk, M.; Nakamura, M.; Zhu, T.H.; Liao, W. Erythrodermic psoriasis: Pathophysiology and current treatment perspectives. Psoriasis (Auckl) 2016, 6, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.; Park, M.; Kim, Y.H.; Ryu, K.H.; Lee, K.H.; Cho, K.A.; Woo, S.Y. Tonsil-derived mesenchymal stem cells (T-MSCs) prevent Th17-mediated autoimmune response via regulation of the programmed death-1/programmed death ligand-1 (PD-1/PD-L1) pathway. J. Tissue. Eng. Regen. Med. 2018, 12, e1022–e1033. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic. Acids. Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Mailleux, J.; Timmermans, S.; Nelissen, K.; Vanmol, J.; Vanmierlo, T.; van Horssen, J.; Bogie, J.F.J.; Hendriks, J.J.A. Low-Density Lipoprotein Receptor Deficiency Attenuates Neuroinflammation through the Induction of Apolipoprotein, E. Front. Immunol. 2017, 8, 1701. [Google Scholar] [CrossRef]
- Zheng, T.; Zhao, W.; Li, H.; Xiao, S.; Hu, R.; Han, M.; Liu, H.; Liu, Y.; Otsu, K.; Liu, X.; et al. p38alpha signaling in Langerhans cells promotes the development of IL-17-producing T cells and psoriasiform skin inflammation. Sci. Signal. 2018, 11, eaao1685. [Google Scholar] [CrossRef] [Green Version]
- Park, W.J.; Kim, S.Y.; Kim, Y.R.; Park, J.W. Bortezomib alleviates drug-induced liver injury by regulating CYP2E1 gene transcription. Int. J. Mol. Med. 2016, 37, 613–622. [Google Scholar] [CrossRef] [Green Version]
- Heymans, S.; Corsten, M.F.; Verhesen, W.; Carai, P.; van Leeuwen, R.E.; Custers, K.; Peters, T.; Hazebroek, M.; Stoger, L.; Wijnands, E.; et al. Macrophage microRNA-155 promotes cardiac hypertrophy and failure. Circulation 2013, 128, 1420–1432. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Ward, P.A.; Wang, X.; Gao, H. Myeloid depletion of SOCS3 enhances LPS-induced acute lung injury through CCAAT/enhancer binding protein delta pathway. FASEB J. 2013, 27, 2967–2976. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Reference |
|---|---|---|
| IL-17A | F: ATCAGGACGCGCAAACATGA | [52] |
| R: TTGGACACGCTGAGCTTTGA | ||
| IL-17F | F: GTCGCCATTCAGCAAGAAAT | [21] |
| R: GGTGCAGCCAACTTTTAGGA | ||
| K6 | F: CTGGTAGTGGCTTTGGCTTC | [21] |
| R: AGGCTCTGGTTGATGGTGAC | ||
| K16 | F: GGTGGCCTCTAACAGTGATCT | [53] |
| R: TGCATACAGTATCTGCCTTTGG | ||
| Gapdh | F: CACTCTTCCACCTTCGATGC | [54] |
| R: CCCTGTTGCTGTAGCCGTAT | ||
| SOCS1 | F: CCTCCTCGTCCTCGTCTTC | [55] |
| R: AAGGTGCGGAAGTGAGTGTC | ||
| SOCS3 | F: AGCTCCAAAAGCGAGTACCA | [56] |
| R: AGCTGTCGCGGATAAGAAAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.-H.; Kim, H.-Y.; Yoon, H.-S.; Park, W.-J.; Adams, D.R.; Pyne, N.J.; Pyne, S.; Park, J.-W. A Novel Selective Sphingosine Kinase 2 Inhibitor, HWG-35D, Ameliorates the Severity of Imiquimod-Induced Psoriasis Model by Blocking Th17 Differentiation of Naïve CD4 T Lymphocytes. Int. J. Mol. Sci. 2020, 21, 8371. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218371
Shin S-H, Kim H-Y, Yoon H-S, Park W-J, Adams DR, Pyne NJ, Pyne S, Park J-W. A Novel Selective Sphingosine Kinase 2 Inhibitor, HWG-35D, Ameliorates the Severity of Imiquimod-Induced Psoriasis Model by Blocking Th17 Differentiation of Naïve CD4 T Lymphocytes. International Journal of Molecular Sciences. 2020; 21(21):8371. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218371
Chicago/Turabian StyleShin, Sun-Hye, Hee-Yeon Kim, Hee-Soo Yoon, Woo-Jae Park, David R. Adams, Nigel J. Pyne, Susan Pyne, and Joo-Won Park. 2020. "A Novel Selective Sphingosine Kinase 2 Inhibitor, HWG-35D, Ameliorates the Severity of Imiquimod-Induced Psoriasis Model by Blocking Th17 Differentiation of Naïve CD4 T Lymphocytes" International Journal of Molecular Sciences 21, no. 21: 8371. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218371