Downregulation of the Polycomb-Associated Methyltransferase Ezh2 during Maturation of Hippocampal Neurons Is Mediated by MicroRNAs Let-7 and miR-124

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. MicroRNA Expression Profile in Mature Hippocampal Neurons
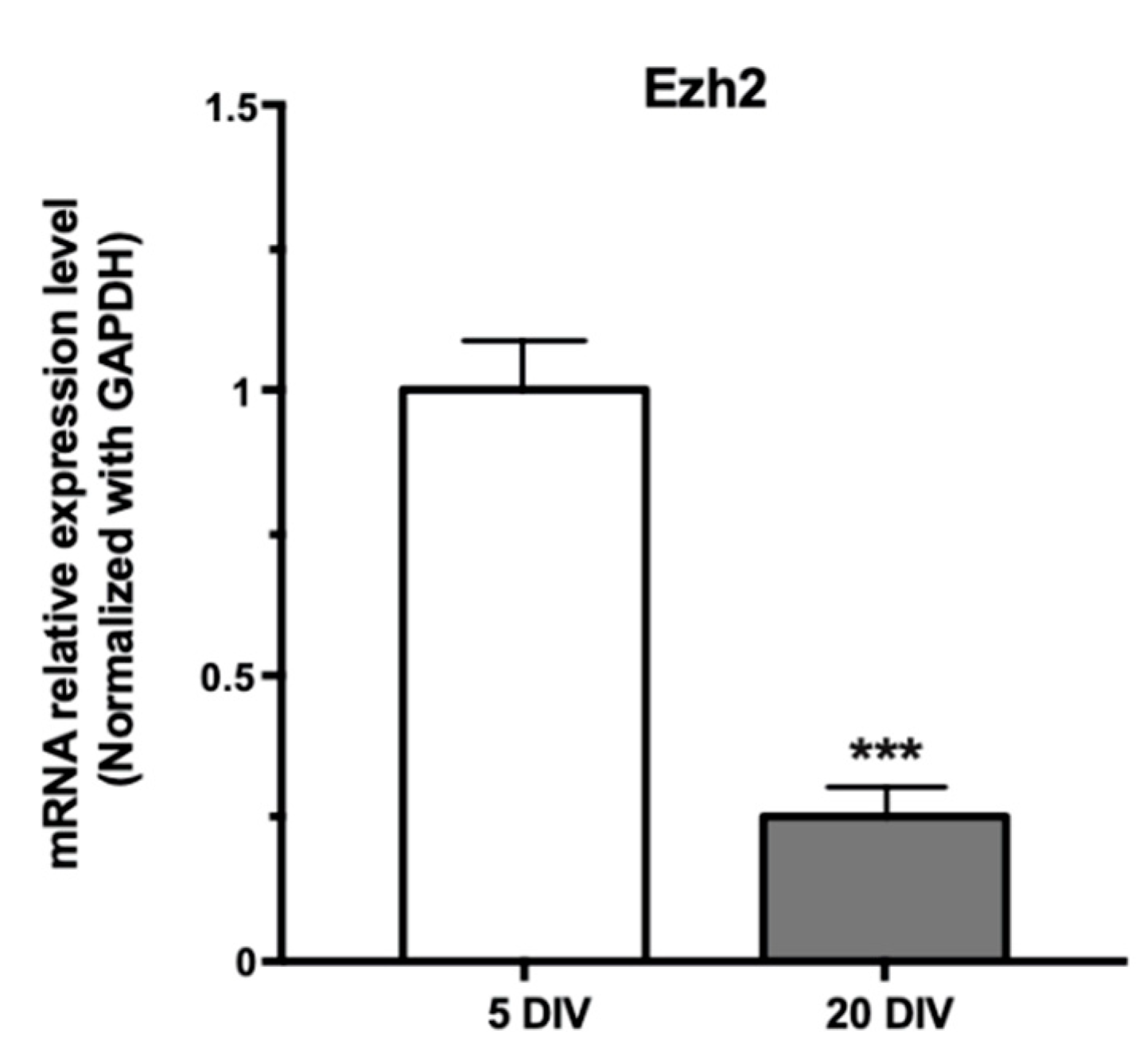
2.2. Downregulation of Ezh2 Expression during Hippocampal Maturation Requires Dicer
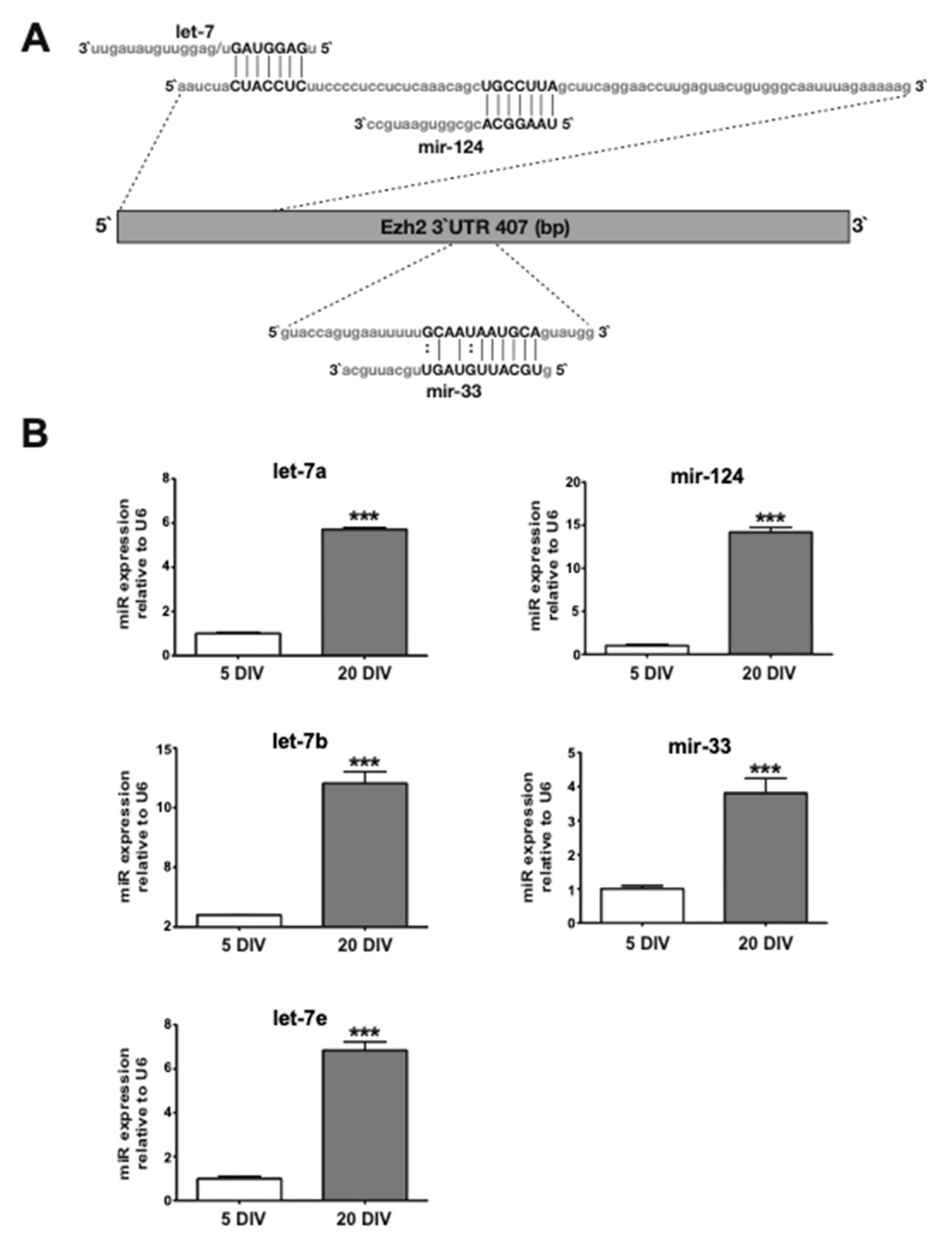
2.3. Candidate miRs for Mediating Ezh2 Downregulation Exhibit Increased Expression during Hippocampal Neuronal Maturation
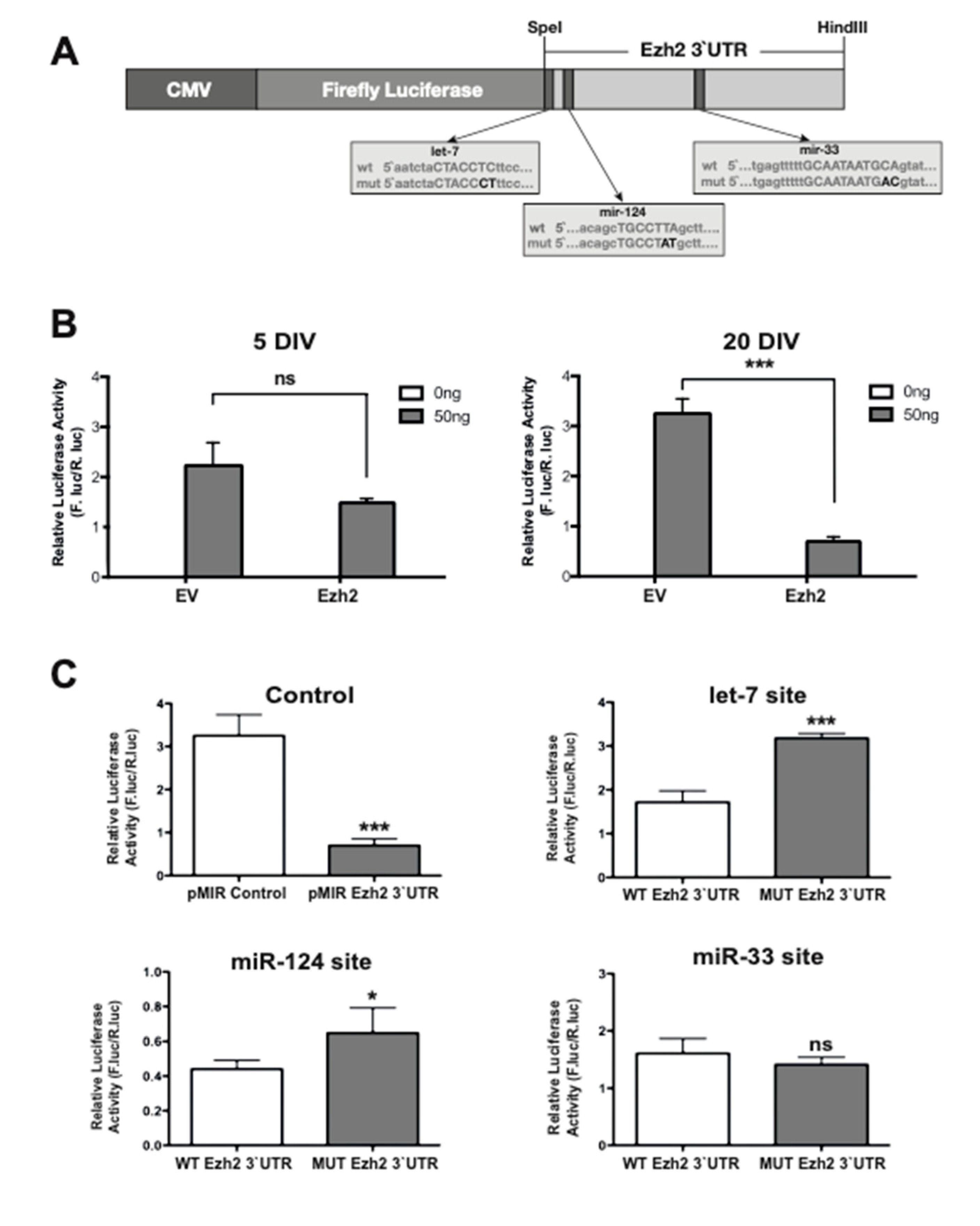
2.4. The 3′-UTR of the Ezh2 Gene Contains Sequence Elements that Mediate Decreased Expression of Immediately Upstream Protein-Coding Genes
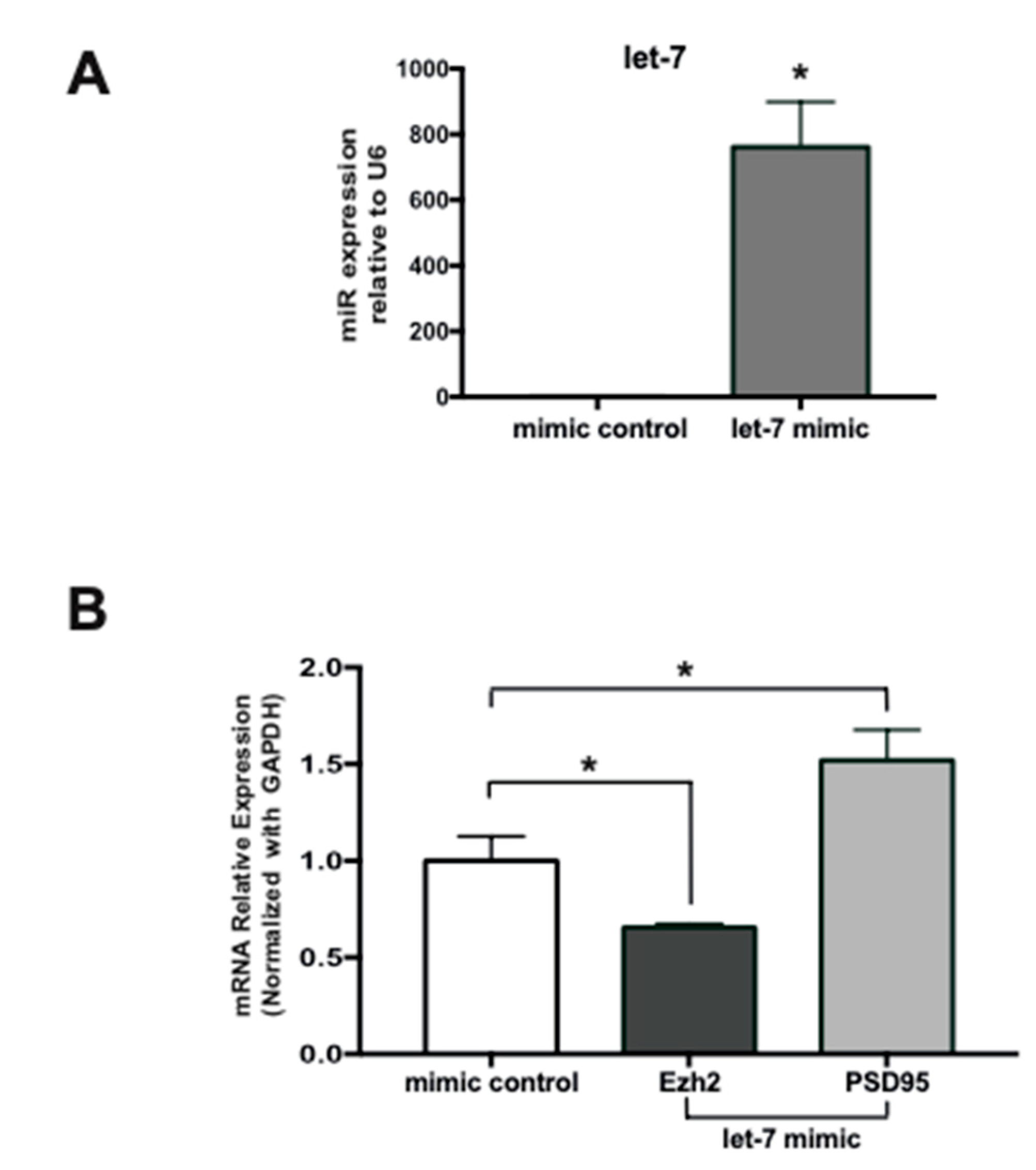
2.5. Expression of a Let-7 Mimic in Immature Hippocampal Neurons Inhibits Ezh2 Expression
3. Discussion
4. Materials and Methods
4.1. Primary Hippocampal Cultures
4.2. MicroRNA Expression Analyses Using Microarrays
4.3. MicroRNA Quantitation
4.4. Plasmid Construction
4.5. Transient Transfections
4.6. Lentivirus Production and Infection of Hippocampal Neurons
4.7. Reverse Transcriptase and Real-Time Quantitative PCR (RT-qPCR)
4.8. Luciferase Reporter Assay
4.9. MicroRNA Target Prediction
4.10. Expression of Let-7 Mimics
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| miRs | MicroRNAs |
| CNS | Central nervous system |
| IFU | Infectious units of virus |
| FMRP | Fragile X mental retardation protein |
| BDNF | Brain-derived neurotrophic factor |
References
- Gräff, J.; Kim, D.; Dobbin, M.M.; Tsai, L.-H. Epigenetic regulation of gene expression in physiological and pathological brain processes. Physiol. Rev. 2011, 91, 603–649. [Google Scholar] [CrossRef]
- Sweatt, J.D. The emerging field of neuroepigenetics. Neuron 2013, 80, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Allis, C.D.; Jenuwein, T. The molecular hallmarks of epigenetic control. Nat. Rev. Genet. 2016, 17, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Margueron, R.; Reinberg, D. The Polycomb complex PRC2 and its mark in life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.-R.; Lee, C.-H.; Oksuz, O.; Stafford, J.M.; Reinberg, D. PRC2 is high maintenance. Genes Dev. 2019, 33, 903–935. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, R.; Bustos, F.J.; Saez, M.; Rojas, A.; Allende, M.L.; van Wijnen, A.J.; van Zundert, B.; Montecino, M. Polycomb PRC2 complex mediates epigenetic silencing of a critical osteogenic master regulator in the hippocampus. Biochim. Biophys. Acta 2016, 1859, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriquez, B.; Bustos, F.J.; Aguilar, R.; Becerra, A.; Simon, F.; Montecino, M.; van Zundert, B. Ezh1 and Ezh2 differentially regulate PSD-95 gene transcription in developing hippocampal neurons. Mol. Cell. Neurosci. 2013, 57, 130–143. [Google Scholar] [CrossRef]
- Pereira, J.D.; Sansom, S.N.; Smith, J.; Dobenecker, M.-W.; Tarakhovsky, A.; Livesey, F.J. Ezh2, the histone methyltransferase of PRC2, regulates the balance between self-renewal and differentiation in the cerebral cortex. Proc. Natl. Acad. Sci. USA 2010, 107, 15957–15962. [Google Scholar] [CrossRef] [Green Version]
- Sher, F.; Boddeke, E.; Copray, S. Ezh2 expression in astrocytes induces their dedifferentiation toward neural stem cells. Cell. Reprogram. 2011, 13, 1–6. [Google Scholar] [CrossRef]
- Shivram, H.; Le, S.V.; Iyer, V.R. MicroRNAs reinforce repression of PRC2 transcriptional targets independently and through a feed-forward regulatory network. Genome Res. 2019, 29, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.-L.; Chou, R.-H.; Shyu, W.C.; Hsieh, S.C.; Wu, C.S.; Chiang, S.Y.; Chang, W.J.; Chen, J.N.; Tseng, Y.J.; Lin, Y.H.; et al. Smurf2-mediated degradation of EZH2 enhances neuron differentiation and improves functional recovery after ischaemic stroke. EMBO Mol. Med. 2013, 5, 531–547. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Sun, G.; Ye, P.; Li, S.; Shi, Y. MicroRNA let-7d regulates the TLX/microRNA-9 cascade to control neural cell fate and neurogenesis. Sci. Rep. 2013, 3, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, F.; Lv, X.; Jiao, J. The role of microRNAs in neural stem cells and neurogenesis. J. Genet Genom. 2013, 40, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Krichevsky, A.M.; King, K.S.; Donahue, C.P.; Khrapko, K.; Kosik, K.S. A microRNA array reveals extensive regulation of microRNAs during brain development. RNA 2003, 9, 1274–1281. [Google Scholar] [CrossRef] [Green Version]
- Somel, M.; Liu, X.; Tang, L.; Yan, Z.; Hu, H.; Guo, S.; Jiang, X.; Zhang, X.; Xu, G.; Xie, G.; et al. MicroRNA-driven developmental remodeling in the brain distinguishes humans from other primates. PLoS Biol. 2011, 9, e1001214. [Google Scholar] [CrossRef]
- Khudayberdiev, S.; Fiore, R.; Schratt, G. MicroRNA as modulators of neuronal responses. Commun. Integr. Biol. 2009, 2, 411–413. [Google Scholar] [CrossRef] [Green Version]
- Schratt, G. microRNAs at the synapse. Nat. Rev. Neurosci. 2009, 10, 842–849. [Google Scholar] [CrossRef]
- De Pietri Tonelli, D.; Pulvers, J.N.; Haffner, C.; Murchison, E.P.; Hannon, G.J.; Huttner, W.B. miRNAs are essential for survival and differentiation of newborn neurons but not for expansion of neural progenitors during early neurogenesis in the mouse embryonic neocortex. Development 2008, 135, 3911–3921. [Google Scholar] [CrossRef] [Green Version]
- Kawase-Koga, Y.; Otaegi, G.; Sun, T. Different timings of Dicer deletion affect neurogenesis and gliogenesis in the developing mouse central nervous system. Dev. Dyn. 2009, 238, 2800–2812. [Google Scholar] [CrossRef] [Green Version]
- Makeyev, E.V.; Zhang, J.; Carrasco, M.A.; Maniatis, T. The MicroRNA miR-124 promotes neuronal differentiation by triggering brain-specific alternative pre-mRNA splicing. Mol. Cell. 2007, 27, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Deo, M.; Yu, J.-Y.; Chung, K.-H.; Tippens, M.; Turner, D.L. Detection of mammalian microRNA expression by in situ hybridization with RNA oligonucleotides. Dev. Dyn. 2006, 235, 2538–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visvanathan, J.; Lee, S.; Lee, B.; Lee, J.W.; Lee, S.-K. The microRNA miR-124 antagonizes the anti-neural REST/SCP1 pathway during embryonic CNS development. Genes Dev. 2007, 21, 744–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conaco, C.; Otto, S.; Han, J.-J.; Mandel, G. Reciprocal actions of REST and a microRNA promote neuronal identity. Proc. Natl. Acad. Sci. USA 2006, 103, 2422–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, A.S.; Sun, A.X.; Li, L.; Shcheglovitov, A.; Portmann, T.; Li, Y.; Lee-Messer, C.; Dolmetsch, R.E.; Tsien, R.W.; Crabtree, G.R. MicroRNA-mediated conversion of human fibroblasts to neurons. Nature 2011, 476, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Ouyang, K.; Huang, J.; Zhou, Y.; Ouyang, H.; Li, H.; Wang, G.; Wu, Q.; Wei, C.; Bi, Y.; et al. Direct conversion of fibroblasts to neurons by reprogramming PTB-regulated microRNA circuits. Cell 2013, 152, 82–96. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.-C.; Pastrana, E.; Tavazoie, M.; Doetsch, F. miR-124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nat. Neurosci. 2009, 12, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Sanuki, R.; Onishi, A.; Koike, C.; Muramatsu, R.; Watanabe, S.; Muranishi, Y.; Irie, S.; Uneo, S.; Koyasu, T.; Matsui, R.; et al. miR-124a is required for hippocampal axogenesis and retinal cone survival through Lhx2 suppression. Nat. Neurosci. 2011, 14, 1125–1134. [Google Scholar] [CrossRef]
- Neo, W.H.; Yap, K.; Lee, S.H.; Looi, L.S.; Khandelia, P.; Neo, S.X.; Makeyev, E.V.; Su, I.-H. MicroRNA miR-124 controls the choice between neuronal and astrocyte differentiation by fine-tuning Ezh2 expression. J. Biol. Chem. 2014, 289, 20788–20801. [Google Scholar] [CrossRef] [Green Version]
- Doeppner, T.R.; Doehring, M.; Bretschneider, E.; Zechariah, A.; Kaltwasser, B.; Müller, B.; Koch, J.C.; Bähr, M.; Hermann, D.M.; Michel, U. MicroRNA-124 protects against focal cerebral ischemia via mechanisms involving Usp14-dependent REST degradation. Acta Neuropathol. 2013, 126, 251–265. [Google Scholar] [CrossRef]
- Lee, S.W.; Oh, Y.M.; Lu, Y.-L.; Kim, W.K.; Yoo, A.S. MicroRNAs Overcome Cell Fate Barrier by Reducing EZH2-Controlled REST Stability during Neuronal Conversion of Human Adult Fibroblasts. Dev. Cell 2018, 46, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Thomson, J.M.; Parker, J.; Perou, C.M.; Hammond, S.M. A custom microarray platform for analysis of microRNA gene expression. Nat. Methods 2004, 1, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulman, B.R.M.; Esquela-Kerscher, A.; Slack, F.J. Reciprocal expression of lin-41 and the microRNAs let-7 and mir-125 during mouse embryogenesis. Dev. Dyn. 2005, 234, 1046–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, J.M.; Newman, M.; Parker, J.S.; Morin-Kensicki, E.M.; Wright, T.; Hammond, S.M. Extensive post-transcriptional regulation of microRNAs and its implications for cancer. Genes Dev. 2006, 20, 2202–2207. [Google Scholar] [CrossRef] [Green Version]
- Wulczyn, F.G.; Smirnova, L.; Rybak, A.; Brandt, C.; Kwidzinski, E.; Ninnemann, O.; Strehle, M.; Seiler, A.; Schumacher, S.; Nitsch, R. Post-transcriptional regulation of the let-7 microRNA during neural cell specification. FASEB J. 2007, 21, 415–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwamborn, J.C.; Berezikov, E.; Knoblich, J.A. The TRIM-NHL protein TRIM32 activates microRNAs and prevents self-renewal in mouse neural progenitors. Cell 2009, 136, 913–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimadamore, F.; Amador-Arjona, A.; Chen, C.; Huang, C.-T.; Terskikh, A.V. SOX2-LIN28/let-7 pathway regulates proliferation and neurogenesis in neural precursors. Proc. Natl. Acad. Sci. USA 2013, 110, E3017–E3026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Sun, G.; Li, S.; Lang, M.-F.; Yang, S.; Li, W.; Shi, Y. MicroRNA let-7b regulates neural stem cell proliferation and differentiation by targeting nuclear receptor TLX signaling. Proc. Natl. Acad. Sci. USA 2010, 107, 1876–1881. [Google Scholar] [CrossRef] [Green Version]
- Thomas, K.T.; Gross, C.; Bassell, G.J. microRNAs Sculpt Neuronal Communication in a Tight Balance That Is Lost in Neurological Disease. Front. Mol. Neurosci. 2018, 11, 455. [Google Scholar] [CrossRef]
- Kong, D.; Heath, E.; Chen, W.; Cher, M.L.; Powell, I.; Heilbrun, L.; Li, Y.; Ali, S.; Sethi, S.; Hassan, O.; et al. Loss of let-7 up-regulates EZH2 in prostate cancer consistent with the acquisition of cancer stem cell signatures that are attenuated by BR-DIM. PLoS ONE 2012, 7, e33729. [Google Scholar] [CrossRef] [Green Version]
- Bustos, F.J.; Varela-Nallar, L.; Campos, M.; Henriquez, B.; Phillips, M.; Opazo, C.; Aguayo, L.G.; Montecino, M.; Constantine-Paton, M.; Inestrosa, N.C.; et al. PSD95 suppresses dendritic arbor development in mature hippocampal neurons by occluding the clustering of NR2B-NMDA receptors. PLoS ONE 2014, 9, e94037. [Google Scholar] [CrossRef] [Green Version]
- Hutvágner, G.; McLachlan, J.; Pasquinelli, A.E.; Balint, E.; Tuschl, T.; Zamore, P.D. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science 2001, 293, 834–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.; Patkaniowska, A.; Urlaub, H.; Lührmann, R.; Tuschl, T. Single-stranded antisense siRNAs guide target RNA cleavage in RNAi. Cell 2002, 110, 563–574. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Price, N.L.; Fernández-Hernando, C. Non-coding RNAs in lipid metabolism. Vascul. Pharmacol. 2019, 114, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.E.; Martin, M.M.; Feldman, D.S.; Terry, A.V.; Nuovo, G.J.; Elton, T.S. Experimental validation of miRNA targets. Methods 2008, 44, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebustini, I.T. A Functional MicroRNA Screening Method for Organ Morphogenesis. Curr. Protoc. Cell Biol. 2017, 74, 19.19.1–19.19.17. [Google Scholar]
- Muddashetty, R.S.; Nalavadi, V.C.; Gross, C.; Yao, X.; Xing, L.; Laur, O.; Warren, S.T.; Bassell, G.J. Reversible inhibition of PSD-95 mRNA translation by miR-125a, FMRP phosphorylation, and mGluR signaling. Mol. Cell. 2011, 42, 673–688. [Google Scholar] [CrossRef] [Green Version]
- Ifrim, M.F.; Williams, K.R.; Bassell, G.J. Single-Molecule Imaging of PSD-95 mRNA Translation in Dendrites and Its Dysregulation in a Mouse Model of Fragile X Syndrome. J. Neurosci. 2015, 35, 7116–7130. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Xie, X. Comparative sequence analysis reveals an intricate network among REST, CREB and miRNA in mediating neuronal gene expression. Genome Biol. 2006, 7, R85. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Han, S.; Kwon, C.S.; Lee, D. Biogenesis and regulation of the let-7 miRNAs and their functional implications. Protein Cell 2016, 7, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-W.A.; Ruiz, C.R.; Eyler, E.C.H.; Lin, K.; Meffert, M.K. Dual regulation of miRNA biogenesis generates target specificity in neurotrophin-induced protein synthesis. Cell 2012, 148, 933–946. [Google Scholar] [CrossRef] [Green Version]
- Sepulveda, F.J.; Bustos, F.J.; Inostroza, E.; Zúñiga, F.A.; Neve, R.L.; Montecino, M.; van Zundert, B. Differential roles of NMDA Receptor Subtypes NR2A and NR2B in dendritic branch development and requirement of RasGRF1. J. Neurophysiol. 2010, 103, 1758–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela-Nallar, L.; Grabowski, C.P.; Alfaro, I.E.; Alvarez, A.R.; Inestrosa, N.C. Role of the Wnt receptor Frizzled-1 in presynaptic differentiation and function. Neural Dev. 2009, 4, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Rehmsmeier, M.; Steffen, P.; Hochsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimson, A.; Farh, K.K.-H.; Johnston, W.K.; Garrett-Engele, P.; Lim, L.P.; Bartel, D.P. MicroRNA targeting specificity in mammals: Determinants beyond seed pairing. Mol. Cell. 2007, 27, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, N.; Wang, X. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015, 43, D146–D152. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ProbeSetID | 20 DIV n1 | 20 DIV n2 | 20 DIV n3 | Mean 20 DIV |
|---|---|---|---|---|
| rno-miR-125b-5p_st | 14,19586735 | 14,11524634 | 14,02201945 | 14,1110444 |
| rno-let-7c_st | 14,05654371 | 13,7811261 | 14,33908404 | 14,058918 |
| rno-miR-124_st | 13,14528831 | 14,67637917 | 13,74687013 | 13,8561792 |
| rno-let-7b_st | 13,77215925 | 13,48533913 | 14,02162067 | 13,7597064 |
| hp_rno-mir-124-3_s_st | 12,66468198 | 14,17408833 | 13,4111353 | 13,4166352 |
| hp_rno-mir-124-1_s_st | 12,55263545 | 14,12533502 | 13,40486244 | 13,3609443 |
| hp_rno-mir-124-2_s_st | 12,50969099 | 13,9984208 | 13,40293019 | 13,3036807 |
| rno-miR-103_st | 12,86020989 | 13,58644686 | 13,02428652 | 13,1569811 |
| rno-miR-99a_st | 13,4265623 | 13,03790836 | 12,6824616 | 13,0489774 |
| rno-miR-107_st | 12,54827077 | 13,33459961 | 12,92074488 | 12,9345384 |
| rno-miR-16_st | 13,06704109 | 13,08511435 | 12,45829264 | 12,8701494 |
| rno-miR-138_st | 11,97407534 | 13,46384005 | 13,02131292 | 12,8197428 |
| rno-miR-24_st | 13,22507587 | 12,5403456 | 12,45652393 | 12,7406485 |
| rno-miR-22_st | 12,80712275 | 12,92439752 | 12,4873622 | 12,7396275 |
| rno-let-7a_st | 12,7448344 | 12,65429313 | 12,66395779 | 12,6876951 |
| rno-miR-181a_st | 12,18081162 | 13,05708698 | 12,71542614 | 12,6511082 |
| rno-miR-191_st | 12,97155739 | 12,54972548 | 12,42775979 | 12,6496809 |
| rno-miR-23a_st | 13,4611495 | 12,41864159 | 12,06288726 | 12,6475594 |
| rno-let-7d_st | 12,33069725 | 12,736612 | 12,86714228 | 12,6448172 |
| rno-miR-26a_st | 12,50207374 | 12,94821494 | 12,21461208 | 12,5549669 |
| rno-let-7e_st | 12,13864311 | 12,47117626 | 12,48884926 | 12,3662229 |
| rno-miR-125a-5p_st | 12,67967 | 12,30327676 | 11,86511822 | 12,2826883 |
| rno-miR-130a_st | 12,38279452 | 12,62256525 | 11,75596603 | 12,2537753 |
| rno-miR-127_st | 11,38137317 | 12,79650642 | 12,28065679 | 12,1528455 |
| rno-miR-99b_st | 12,31702088 | 11,70084872 | 11,9723612 | 11,9967436 |
| Target | microRNA | Chromosome | Coding Details |
|---|---|---|---|
| let-7a-1 | 17 | Clustered miR-3596b, miR-3596d, let-7d; let-7f-1 | |
| let-7a-2 | 8 | Clustered miR-100, miR-3695a, in lnc215 gene | |
| let-7b | 7 | Clustered let-7c-2 | |
| let-7c-1 | 11 | Clustered miR-99a | |
| let-7c-2 | 7 | Clustered let-7b | |
| let-7d | 17 | Clustered miR-3596b, miR-3596d, let-7a-1, let-7f-1, in Spaca6 gene | |
| let-7e | 1 | Clustered miR-99b, miR-3596c, miR-125a | |
| Ezh2 | let-7f-1 | 17 | Clustered 3596b, miR-3596d, let-7a-1, let-7d |
| let-7f-2 | X | Clustered miR-98, in Huwe1 gene | |
| let-7g | 8 | IR | |
| let-7i | 7 | IR | |
| miR-124-1 | 15 | IR | |
| miR-124-2 | 2 | IR | |
| miR-124-3 | 3 | IR | |
| miR-33 | 7 | In Srebf2 gene |
| miR Analyzed | Ezh2 3′UTR | MRE—miR Alignments Predicted | ΔG MRE WT | ΔG MRE MUT |
|---|---|---|---|---|
| WT | 5′AAUCUACUACCUCU 3′ | |||
| let-7 | 7–13 | 3′UGAUAUGUUGGAGGAUGGAGU 5′ | −20.1 | −15.4 |
| MUT | 5′AAUCUACUACCCUU 3′ | |||
| WT | 5′CUCCUCUCAAACAGCUGCCUUAG 3′ | |||
| miR-124 | 33–39 | 3′CCGUAAGUGGCGC-ACGGAAU 5′ | −19.9 | −15.8 |
| MUT | 5′CUCCUCUCAAACAGCUGCCUAUG 3′ | |||
| WT | 5′UGAAUUUUUGCAAUAAUGCAG 3′ | |||
| miR-33 | 195–216 | 3′ACGUUACGUUGAUGUUACGUG 5′ | −18.2 | −14.2 |
| MUT | 5′UGAAUUUUUGCAAUAAUGACG 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guajardo, L.; Aguilar, R.; Bustos, F.J.; Nardocci, G.; Gutiérrez, R.A.; van Zundert, B.; Montecino, M. Downregulation of the Polycomb-Associated Methyltransferase Ezh2 during Maturation of Hippocampal Neurons Is Mediated by MicroRNAs Let-7 and miR-124. Int. J. Mol. Sci. 2020, 21, 8472. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228472
Guajardo L, Aguilar R, Bustos FJ, Nardocci G, Gutiérrez RA, van Zundert B, Montecino M. Downregulation of the Polycomb-Associated Methyltransferase Ezh2 during Maturation of Hippocampal Neurons Is Mediated by MicroRNAs Let-7 and miR-124. International Journal of Molecular Sciences. 2020; 21(22):8472. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228472
Chicago/Turabian StyleGuajardo, Laura, Rodrigo Aguilar, Fernando J. Bustos, Gino Nardocci, Rodrigo A. Gutiérrez, Brigitte van Zundert, and Martin Montecino. 2020. "Downregulation of the Polycomb-Associated Methyltransferase Ezh2 during Maturation of Hippocampal Neurons Is Mediated by MicroRNAs Let-7 and miR-124" International Journal of Molecular Sciences 21, no. 22: 8472. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228472