2.1. Reconstruction and Analysis of the Networks Associated with Hyperglycemia and Hypoglycemia
At the first step, molecular networks related to hyperglycemia (
Figure S1, Table S1) and hypoglycemia (
Figure S2, Table S2) were reconstructed using the ANDSystem [
33,
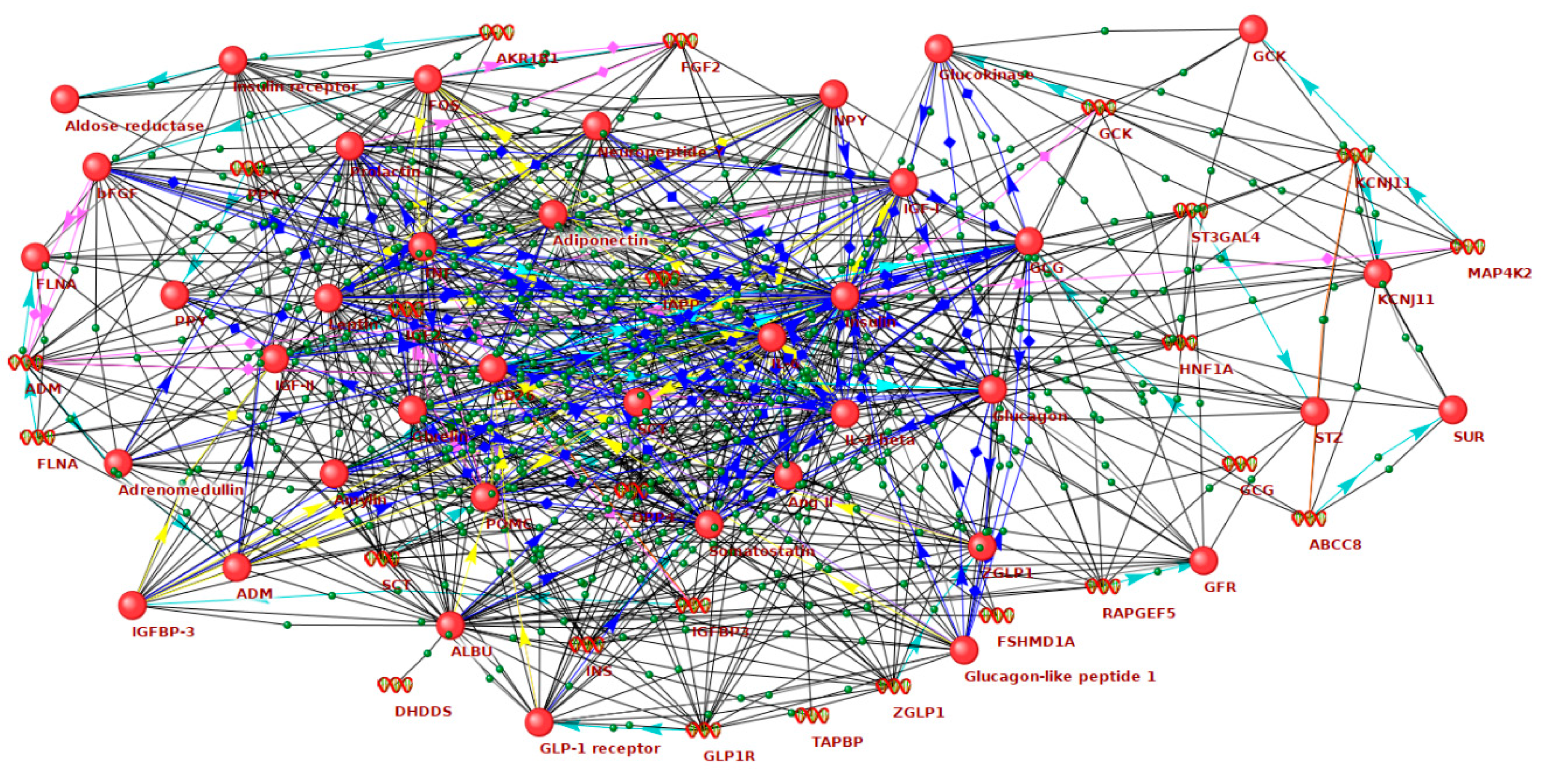
34]. The gene network of hyperglycemia included 209 genes/proteins and 5818 interactions between them. The gene network of hypoglycemia included 128 genes/proteins and 2467 interactions. At the intersection of these gene networks, we identified 37 genes/proteins. These molecules were associated with both hyperglycemia and hypoglycemia, being highly connected with each other in the gene network (
Table S3,
Figure 1). It can be assumed that these genes and proteins are most closely related to enhanced GV, which is characterized by the transition from hyperglycemia to hypoglycemia and vice versa. In what follows, we will call such genes as GV-related ones.
Ten GV-related genes showed the largest number of connections with other molecules in the network (
Table 1). Among these molecules, as expected, there are insulin (
INS) and glucagon (
GCG), two principal regulators of blood glucose level in humans, as well as other hormones that affect glucose metabolism directly or indirectly, including leptin (
LEP), insulin-like growth factor 1 (
IGF1), somatostatin (
SST), adiponectin (
ADIPOQ), prolactin (
PRL), ghrelin (
GHRL), glucagon like peptide 1 receptor (
GLP1R), proopiomelanocortin (
POMC), amylin (
IAPP), adrenomedullin (
ADM), angiotensin II (
AGT), secretin (
SCT), and pancreatic polypeptide (
PPY). The network includes genes encoding proinflammatory cytokines (
IL6, IL1B, TNF), growth factors (
FGF2, IGF2), molecules involved in the orchestration of insulin secretion and signaling (
GCK, KCNJ11, ABCC8, INSR, FLNA), transcriptional factors (
HNF1A, FOS, ZGLP1), neurotransmitter (
NPY), enzymes (
DPP4, AKR1B1, ST3GAL4, MAP4K2, RAPGEF5), and binding proteins (
ALB, IGFBP3, TAPBP).Thus, the network associated with GV includes a variety of molecular components. Hormones constitute a significant part of this network. This is consistent with the well-known fact that maintenance of glycemic fluctuations within the physiological limits is controlled by fine and well-balanced hormonal regulation. Circadian rhythms in secretion of growth hormone, melatonin, prolactin, cortisol, leptin, and ghrelin are highly correlated with sleep and can contribute to the intra-day glycemic fluctuations [
35,
36,
37].
Among non-hormonal factors, interleukin-6 (
IL6) showed the highest number of connections in the GV-related network. Circadian rhythm in IL-6 secretion with an elevation in day-time hours was described [
38]. As IL-6 modulates glucose disposal and insulin sensitivity [
39], one could speculate it can affect intraday GV. On the other hand, excessive glucose fluctuations can increase IL-6 production. In cultured human endothelial cells, an intermittent increase in glucose levels caused a more pronounced increase in
IL6 expression than a consistently high glucose [
40]. The association between GV and plasma level of IL-6 has been shown in individuals with metabolic syndrome [
41]. In patients with type 1 diabetes, an increase in the IL-6 levels was observed after a two-hour episode of hypoglycemia [
42]. Hyperglycemia, following hypoglycemia, caused a further increase in IL-6 concentration [
18].
The genes associated with insulin secretion and signaling turned out to be important nodes of the GV network. In humans, mutations in these genes (
GCK, HNF1A, ABCC8, and
KCNJ11) are causes of monogenic forms of diabetes known as maturity-onset diabetes of the young [
43]. An increased GV was reported in patients with diabetes caused by mutations in the
HNF1A [
44] and
ABCC8 [
45]. Mutations in the insulin receptor gene (
INSR) are associated with insulin resistance and hyperglycaemia and can lead to hyperinsulinemic hypoglycaemia in adults and children [
46,
47]. The
GCK, KCNJ11, ABCC8, and
INSR were identified as genetic variants associated with type 2 diabetes and/or glycemic traits in genome-wide association studies [
48]. However, the contribution of the polymorphisms of these genes to GV in healthy individuals and patients with diabetes has not been studied yet. The role of a number of molecules, identified as the network components, in GV phenomenon remains to be clarified.
At the next step, we performed the comparisons of expression patterns of GV-related genes in different tissues with the use of Bgee [
49]. The greatest expression of GV-related genes was observed in the cardiovascular system, followed by pancreas, adipose and muscle tissues, gastrointestinal tract, and kidney; retina and nerves demonstrated a less maximal expression score (
Table S4). This fact gives further support to notion that GV could be the unifying link between impairment of insulin synthesis and signaling, glucose metabolism, and diabetic vascular complications.
Table 2 demonstrates the results of the Gene Ontology (GO) enrichment analysis by DAVID service [
50], aimed at revealing the overrepresented biological processes for GV-related genes. In addition to the regulation of insulin secretion and different aspects of glucose metabolism, this analysis revealed some signaling pathways considered as the cornerstones in regulation of cellular metabolism (mitogen-activated protein kinase (MAPK) cascade, protein kinase B signaling, JAK-STAT cascade, regulation of peptidyl-tyrosine phosphorylation), cell cycle (regulation of mitotic nuclear division and cell proliferation), and cell-cell signaling. Positive regulation of nitric oxide biosynthetic process was identified among overrepresented processes.
It turned out that the biological processes identified as overrepresented are closely related to oxidative stress, glycation, endothelial dysfunction, and other processes that are considered as important players in the development of diabetes complications. In particular, oxidative stress can directly alter glucose homeostasis [
51] and it itself is triggered by hyperglycemia [
52]. The intensity of glycation is proportional to glucose concentration and depends on glucose metabolism [
53,
54]. MAPK cascade and nitric oxide biosynthesis are known as principal pathways in the biology of endothelial cells [
55,
56].
These data support the notion that GV-related genes could play an important role in pathogenesis of diabetic complications. We tested this assumption at the next stage of our work.
2.2. The Role of GV-Related Genes in the Networks of Diabetes Complications
The ANDSystem was used to construct gene networks associated with diabetic complications: cardiovascular disease (
Figure S3, Table S5), diabetic nephropathy (
Figure S4, Table S6), diabetic retinopathy (
Figure S5, Table S7), and diabetic neuropathy (
Figure S6, Table S8). For each of these gene networks, the role of GV-related genes was assessed by analyzing the betweenness centrality coefficients of the network participants (
Table 3). Betweenness centrality reflects the number of shortest pathways between all pairs of genes in the analyzed gene network that go through a given gene. It describes the functional importance of a gene in a gene network.
Analysis shows that GV-related genes are significantly overrepresented in all networks of diabetes complications. In addition, the centrality of the GV-related genes was, on average, significantly higher than the average centrality of all participants of the considered gene networks (
Table 3). These data support the key role of GV-related genes in the molecular orchestra of diabetes complications.
In order to find out what biological processes involved in pathogenesis of diabetic complications are triggered by glycemic fluctuations, the following analysis was carried out. Among the participants of the gene networks of diabetes complications, we selected those that were directly regulated by GV-related genes, according to ANDSystem. Thus, we identified 141 genes in the network of cardiovascular diseases, 219 genes in the network of diabetic nephropathy, 132 genes in the network of diabetic retinopathy, and 44 genes in the network of diabetic neuropathy. Further, using the DAVID service [
50], the GO biological processes overrepresented for these genes were identified (
Table S9).
It was found that the response to hypoxia was overrepresented in all four sets of genes, and in all cases, it was among the top ten most significantly overrepresented biological processes. This suggests an important role of hypoxia as a mediator between GV and diabetes complications. Diabetes is characterized by altered oxygen deprivation signaling in the targeted organs, including the vasculature and kidney. Cellular response to hypoxia is controlled by hypoxia-inducible factor (HIF), a transcriptional regulator. The HIF family is considered as a master regulator of endothelial biology with rising interest in the field of diabetes-driven atherosclerosis [
57]. It is postulated that a shift in the balance of HIF-1α and HIF-2α promotes proinflammatory and profibrotic pathways in glomerular and renal tubular cells [
58]. A growing body of research indicates HIF-1α and its target genes as the contributors to the retinal neovascularization in diabetic retinopathy [
59]. In obesity and type 2 diabetes, inflammation and hypoxia of adipose tissue may play an important role in the development of insulin resistance [
60]. In its turn, intermittent hypoxia, which is used to improve the adaptation to hypoxia, reduced blood glucose in patients with type 2 diabetes [
61]. These data are in agreement with our findings indicating the response to hypoxia as an important mechanism linking GV and chronic diabetic complications.
Other 15 biological processes were revealed to be overrepresented simultaneously for three sets of genes important for cardiovascular disease, diabetic nephropathy, and diabetic retinopathy (
Table 4). They are involved in the inflammatory response, platelet function, angiogenesis, regulation of blood pressure, and nitric oxide biosynthesis. These processes, along with endothelial dysfunction, are considered as the cornerstones in the pathogenesis of diabetic vascular complications [
62].
Molecular pathways, implicated in the regulation of cell cycle, proliferation, and cell-cell signaling, have been recognized among significantly overrepresented biological processes related to GV and complications. Among them, extracellular signal-regulated kinase (ERK) 1 and 2 cascade should be mentioned. ERK cascade plays a crucial role in multiple cellular processes, such as cell proliferation, differentiation, adhesion, migration, and survival [
63]. Glucose can activate ERK1/2 in beta-cells, contributing to an increase in mitogenesis [
64]. Moreover, activation of ERK is a component of insulin signaling. Accordingly, the targeting of the ERK cascade is considered as a potential treatment for insulin resistance and type 2 diabetes [
65]. The activation of ERK1/2 is involved in pathogenesis of cardiac hypertrophy and dysfunction; recent studies showed the role of ERK in development of diabetic cardiomyopathy [
66]. High glucose increases ERK phosphorylation in mesangial cells, promoting the synthesis of extracellular matrix, a key event in diabetic kidney disease [
67].
Thus, it was revealed that GV-related genes occupy central positions in the gene networks of diabetes complications. The response to hypoxia, low-grade inflammation, abnormalities of angiogenesis and platelets function, disturbances in the regulation of blood pressure and vasodilatation, as well as alterations of cellular signaling pathways, could link GV with the development of diabetic complications.
2.3. Identification of New Candidate Genes in the GV Network
At the first step of this study, we identified by the ANDSystem the set of genes that are directly associated with both hyperglycemia and hypoglycemia, attributed them to GV. The Pathway Wizard tool of the ANDSystem allows one to identify indirect interactions between objects in the gene networks that pass through genes-intermediaries. Using this opportunity, it was possible to find genes/proteins for which there is no evidence on a direct connection with hypoglycemia and hyperglycemia, but there are indications of indirect relationships with glucose abnormalities through genes-intermediaries. As genes-intermediaries, we considered the GV-related genes described in the first section of this article. The genes/proteins that were associated with genes-intermediaries, we considered as candidate genes potentially involved in GV network. The analysis identified 334 such candidate genes. Further, in order to rank candidates and determine the most promising ones, gene prioritization was carried out using the ToppGene system [
68]. The GV-related genes were used as a training set.
The results of prioritization could be found in
Table S10. The top ten most-priority genes are shown in
Table 5. These genes are important for cell-cell interactions (
THBS1, FN1), intracellular signal transmission (
MAPK1, STAT3, TP53), stress response (
HSP90AA1, TP53, GSK3B, PTEN), cell proliferation and differentiation (
MAPK1, TP53, EGFR, EGF), apoptosis (
STAT3, TP53, GSK3B), and glucose metabolism (
GSK3B).
The
THBS1 gene received the highest priority. This gene encodes an adhesive glycoprotein thrombospondin 1 (THSP1), which regulates cell-cell and cell-matrix interactions. THSP1 is an important player in the regulation of adhesion, mobility, and growth of endothelial cells; it has important regulatory function in angiogenesis, inflammation, and tissue remodeling also [
69]. Patients with diabetes and coronary artery disease have increased THSP1 levels in plasma [
70]. The enhanced expression of
THBS1 was observed in mice with streptozotocin-induced diabetes. In cultured fibroblasts, high glucose increased expression of
THBS1 [
71]. Being involved in atherosclerosis, inflammation, ischemia/reperfusion injury, cardiac hypertrophy, and heart failure,
THBS1 plays a significant role in the development of cardiovascular diseases [
72]. In diabetic complications, this molecule is considered to be a key inductor of fibrosis [
73].
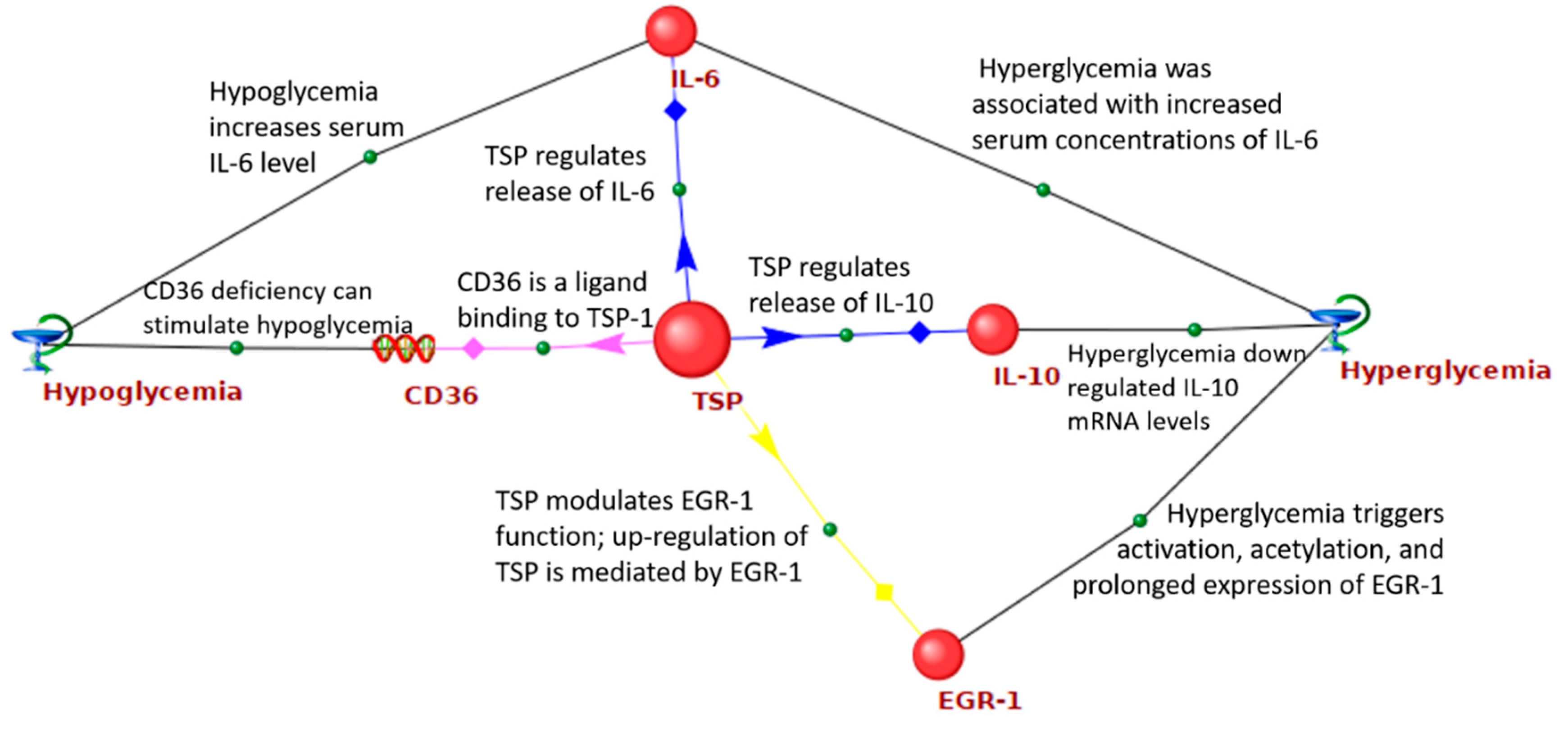
Figure 2 shows the network of interactions between the
THBS1 and GV-related genes. It was demonstrated that
THBS1 is able to bind to CD36, a transporter and sensor of free fatty acids. Binding of CD36 to
THBS1 initiates anti-angiogenic signals, induction of endothelial cell apoptosis, and the inhibition of angiogenesis [
74].
THBS1 differentially regulates the release of IL-6 and IL-10 by human monocytes [
75]. Both hypoglycemia and hyperglycemia increased serum concentrations of IL-6 [
76,
77], while hyperglycemia down-regulated
IL-10 mRNA levels [
78].
THBS1 modulates epidermal growth factor 1 (EGR1) function [
79]. At the same time, up-regulation of
THBS1 is mediated by EGR1 [
80]. Hyperglycemia is responsible for EGR1 activation, acetylation, and its prolonged expression [
81].
Fibronectin is a component of the extracellular matrix, accumulating in the retinal and glomerular basement membranes and mesangial matrix in diabetic retinopathy and diabetic kidney disease [
82,
83]. High ambient glucose increases fibronectin production in human mesangial cells [
84] and retinal pigment epithelial cells [
85]. In mesangial cells, the effect was mediated via THSP and TGF-beta [
84].
Heat shock protein 90α (Hsp90α), encoded by the
HSP90AA1 gene, is the stress-inducible isoform of the molecular chaperone Hsp90. The expression of
HSP90AA1 increases in some forms of cancer [
86]. Previously,
HSP90AA1 was recognized by bioinformatic analysis as a node in a protein-protein interaction network in diabetic nephropathy [
87]. However, the exact role of Hsp90α in diabetes and its complications remains to be clarified.
Both
EGF and
EGFR, encoding epidermal growth factor (EGF) and its receptor respectively, were present in the list of gene-intermediates. In diabetes, EGF/EGFR system was discussed primarily as a promoter of wound healing [
88]. Recently it was demonstrated that EGF signal is involved in diabetes-induced vascular dysfunction, remodeling, and transcriptome dysregulation associated with renal involvement [
89]. In patients with type 2 diabetes, soluble EGFR correlates with fasting blood glucose, insulin resistance, and glycated hemoglobin HbA1c [
90]. Interestingly, EGFR can regulate the transcription of proopiomelanocortin [
91], a hypothalamic appetite regulator and ACTH precursor modulated by glucose level [
92]. Both EGFR and TP53 regulate expression of
IGFBP3 [
93,
94], which level is reduced in response to hyperglycemia [
95].
Mitogen-activated protein kinase 1 (MAPK1), also known as ERK2, is a molecular target of metformin, a principal antihyperglycemic agent [
96]. MAPK1 is one of the essential molecules implicated in canonical insulin signaling, an important modulator of protein synthesis [
96]. In the liver, MAPK1 is involved in regulation of glucose and lipid metabolism required for physiological metabolic adaptation [
97]. In mice on a high-fat/high-sucrose diet, MAPK1 knockout promoted insulin resistance and impairment of glucose tolerance [
98].
Signal transducer and activator of transcription 3 (STAT3) is a principal modulator of β-cell survival and function [
99]. It is involved in the regulation of hepatic insulin sensitivity and gluconeogenesis [
100]. The excessive STAT3 signaling contributes to skeletal muscle insulin resistance [
101]. Recent data indicated the role of STAT3 signaling in high-glucose-induced podocyte injury [
102], tubular cell dysfunction [
103], and diabetic retinopathy [
104].
Tumor protein p53 (TP53), a tumor suppressor, was recently identified as a metabolic regulator. In the liver, TP53 improves insulin sensitivity via inhibition of MAPKs and NF-κB pathways [
105]. The protein regulates the function of glucose transporters affecting their transcription and translocation. It negatively regulates glycolysis and positively regulates gluconeogenesis [
106]. In the beta cells, TP53 mediates gluco- and lipotoxicity and induces apoptosis [
106]. In patients with type 2 diabetes, the level of TP53 increased with the age, duration of diabetes, and waist-to-hip ratio [
107].
Glycogen synthase kinase 3 beta (GSK3B) is a serine/threonine protein kinase that is involved in glycogen metabolism, regulation of gene transcription, cytoskeleton organization, cell cycle, and apoptosis. The enzyme inhibits glycogen synthase and decreases glycogen storage in skeletal muscles and, to a lesser extent, in the liver; it also deteriorates glucose tolerance and insulin sensitivity. Insulin inactivates GSK3B via protein kinase B (Akt) [
108]. It was demonstrated that GSK3B modulates inflammatory response in the islets [
109]. Women with gestational diabetes had increased GSK3B activity in skeletal muscle and adipose tissue, where the enzyme promoted the inflammatory response [
110]. Blood glucose fluctuations suppressed the expression of phosphorylated GSK3B and contributed to cardiac fibrosis in a model of diabetic cardiomyopathy [
111].
Phosphatase and tensin homolog (PTEN) is essential for both cellular growth and metabolism. PTEN counteracts insulin signaling via PI3K/Akt pathway and reduces insulin sensitivity in mouse adipose tissue, liver, and β-cells [
112]. In humans, mutations in
PTEN gene increase the risks of obesity and cancer, but decrease the risk of type 2 diabetes [
113]. Recent experimental studies indicate the role of PTEN in β-cell function and apoptosis [
99], hyperglycemia-induced angiogenesis suppression [
114], and development of diabetic nephropathy [
115].
As an indirect confirmation of the relevance of performed gene prioritization, we checked the co-occurrence of the candidate gene together with the terms denoting GV in full-text PubMed Central articles. We used information from full-text articles to test our prioritization because ANDSystem stores the information gathered from the abstracts, but not full-text articles. For each candidate gene, using the hypergeometric distribution, the likelihood of meeting the name of this gene, together with the terms denoting GV in full-text of PubMed Central articles, was estimated (
Table S11). At the next step, the Spearman’s rank correlation coefficient was calculated between the prioritization rank of a gene and the probability to discover the gene in articles together with the terms describing GV by chance. The correlation coefficient was found to be 0.3 and the statistical significance of the correlation was 1.6 × 10
−8. This indirectly confirms the adequacy of the gene prioritization. It means that the top candidate genes are promising for experimental verification of their relationship with GV.
For further verification of obtained results, we used transcriptomic data stored in GEO (
https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/geo/info/), the largest open repository of experiments in the field of gene expression. We were able to find only one study (GEO ID: GSE40779) related to glucose fluctuations in diabetes. In this study, carried out in
Rattus norvegicus, non-coding RNA profile was assessed in the heart under glucose fluctuations compared to sustained hyperglycemia [
116]. It was found that the expression of 20 of 350 scanned miRNAs significantly differed between diabetic hearts and diabetic hearts with glucose fluctuations [
116]. We have conducted the search for the target genes of these 20 differentially expressed miRNAs in the miRTarBase database [
117]. For 16 differentially expressed miRNAs, 69 target genes in
Rattus norvegicus have been found. Four miRNAs (rno-miR-136, rno-miR-296*, rno-miR-326, and rno-miR-30c-2*) had no target genes in miRTarBase. At the next step, human orthologous genes were found for these 69 target genes using the NCBI Orthologs service (
Table S12). Next, we matched these genes with those we identified as the candidate genes most likely associated with GV.
It turned out that five genes, namely PTEN, TNFRSF1A, CCND1, TLR2, and RELA, which we proposed as the candidate genes for experimental verification, are targets of microRNAs differentially expressed in the GSE40779 experiment. According to the hypergeometric distribution, the probability of observing this for random reasons is low (p-value = 0.006). In addition, all of these five genes were among the top 100 candidate genes with highest priority predicted by us, which, according to the hypergeometric distribution, is unlikely by chance (p-value = 0.002). Moreover, three of five discussed genes were among the top 50 candidate genes with highest priority, which is also unlikely to be observed for random reasons (p-value = 0.03), and the PTEN was among the top ten most-priority candidates. This indirectly confirms the results of our bioinformatics analysis.






{kind=link}
{kind=link}