GLP-2 Prevents Neuronal and Glial Changes in the Distal Colon of Mice Chronically Treated with Cisplatin

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
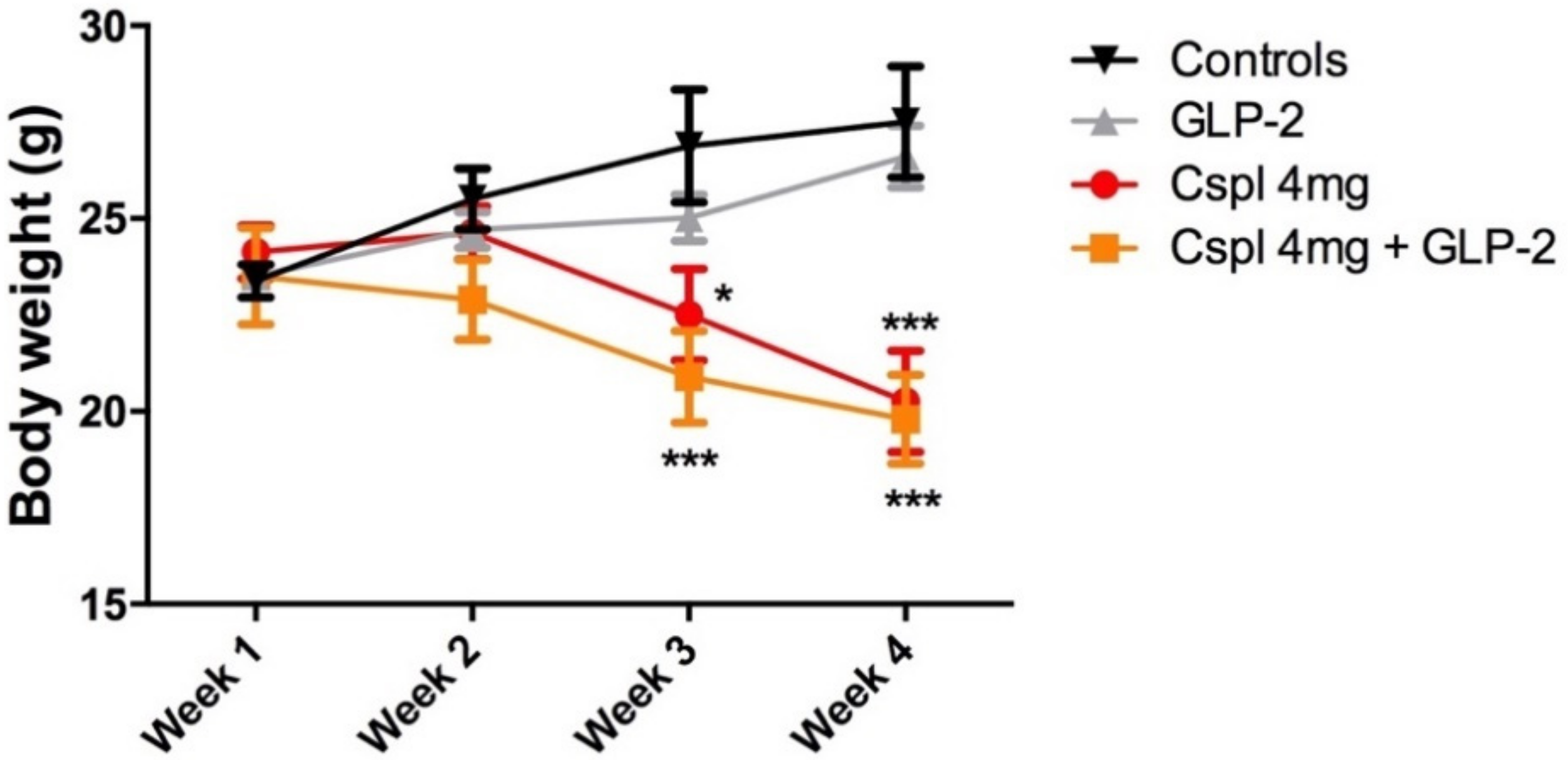
2.1. GLP-2 Does Not Affect the Weight Loss Induced by Chronic Cisplatin Treatment
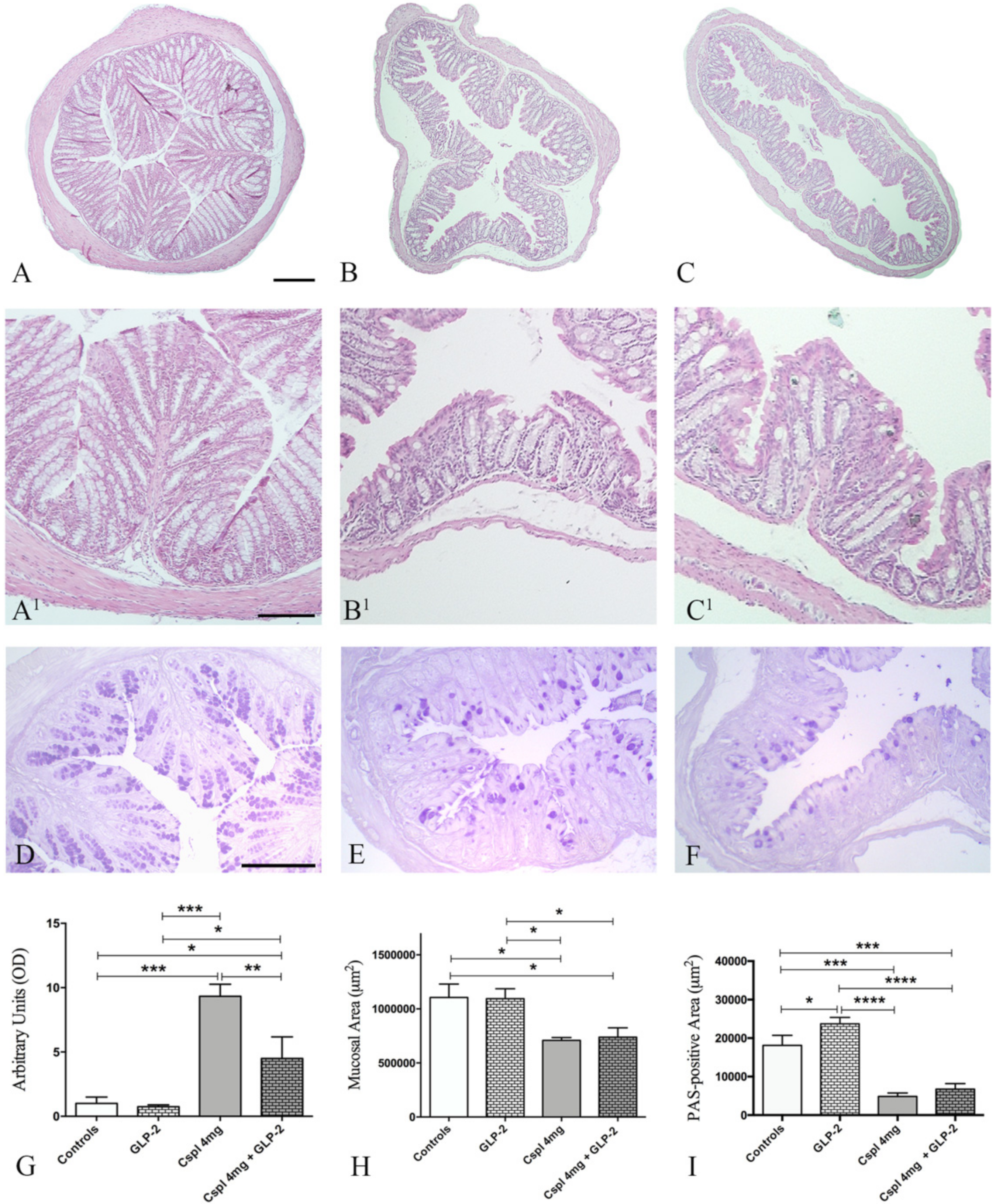
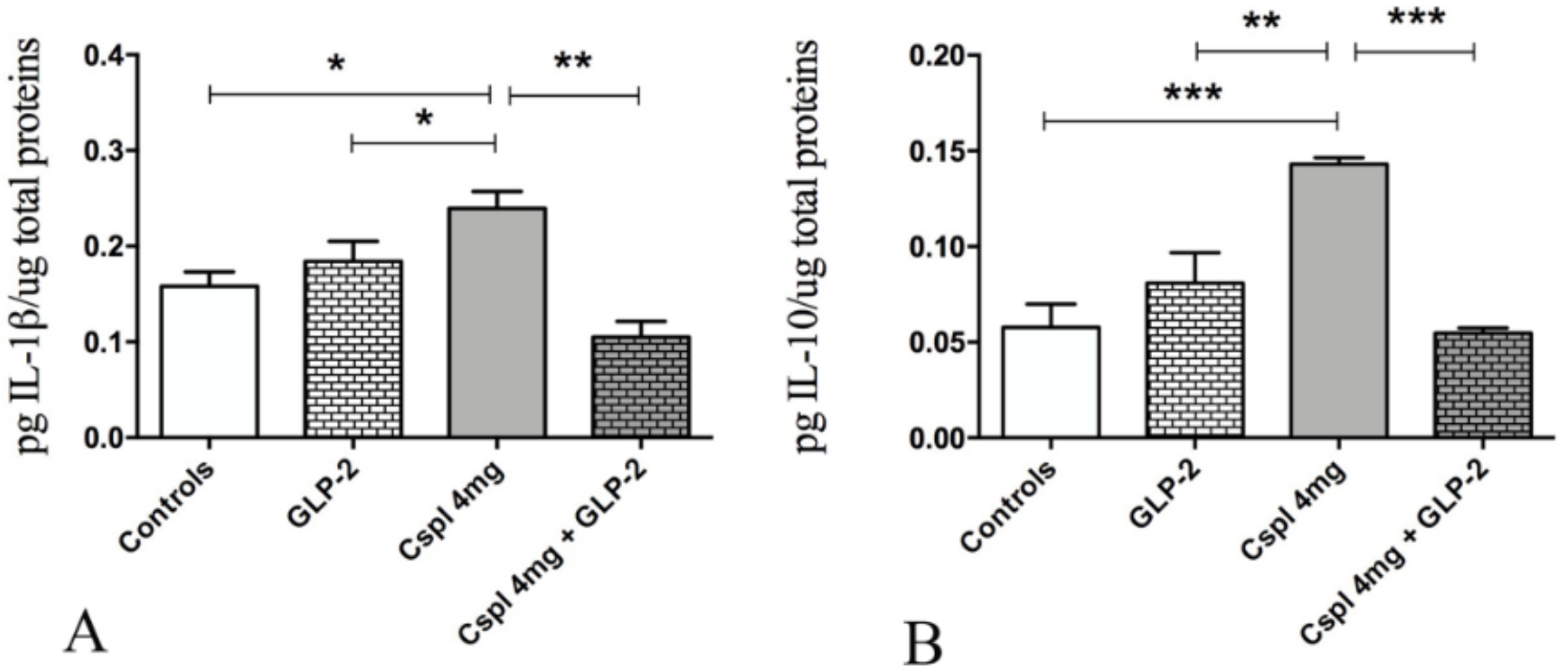
2.2. GLP-2 Prevents Cytokines Increase and Partially Ameliorate Mucosal Damage Induced by Cisplatin in the Distal Colon
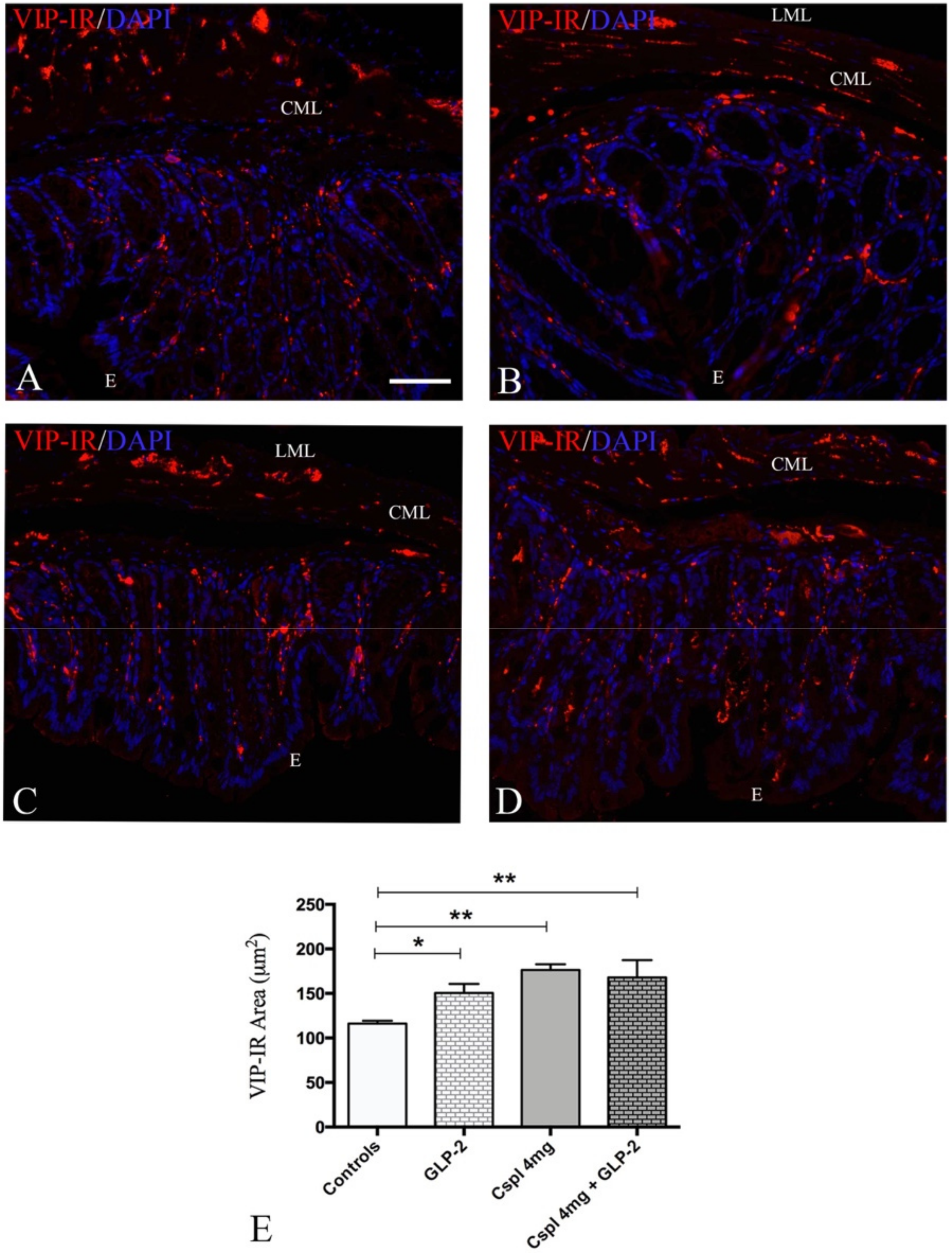
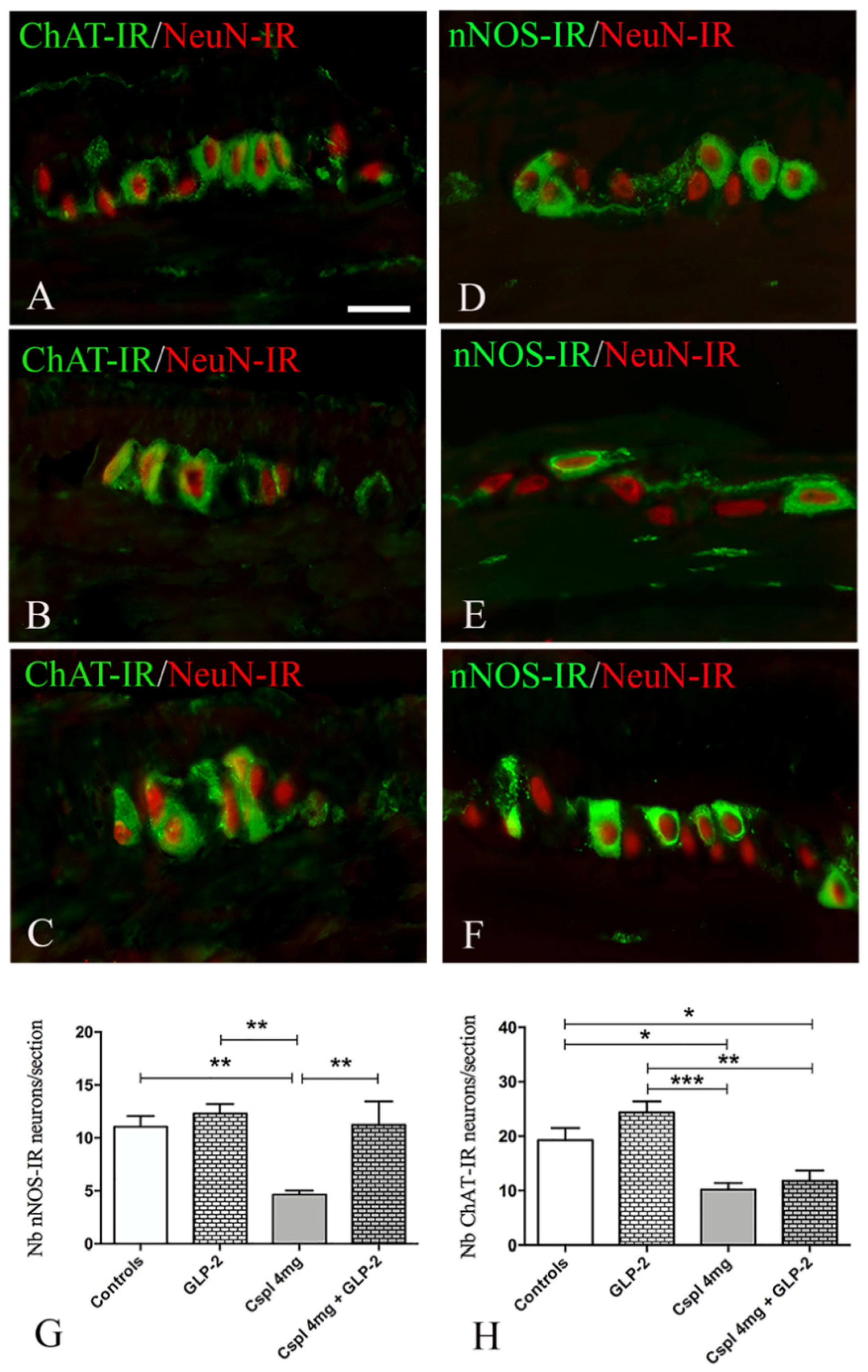
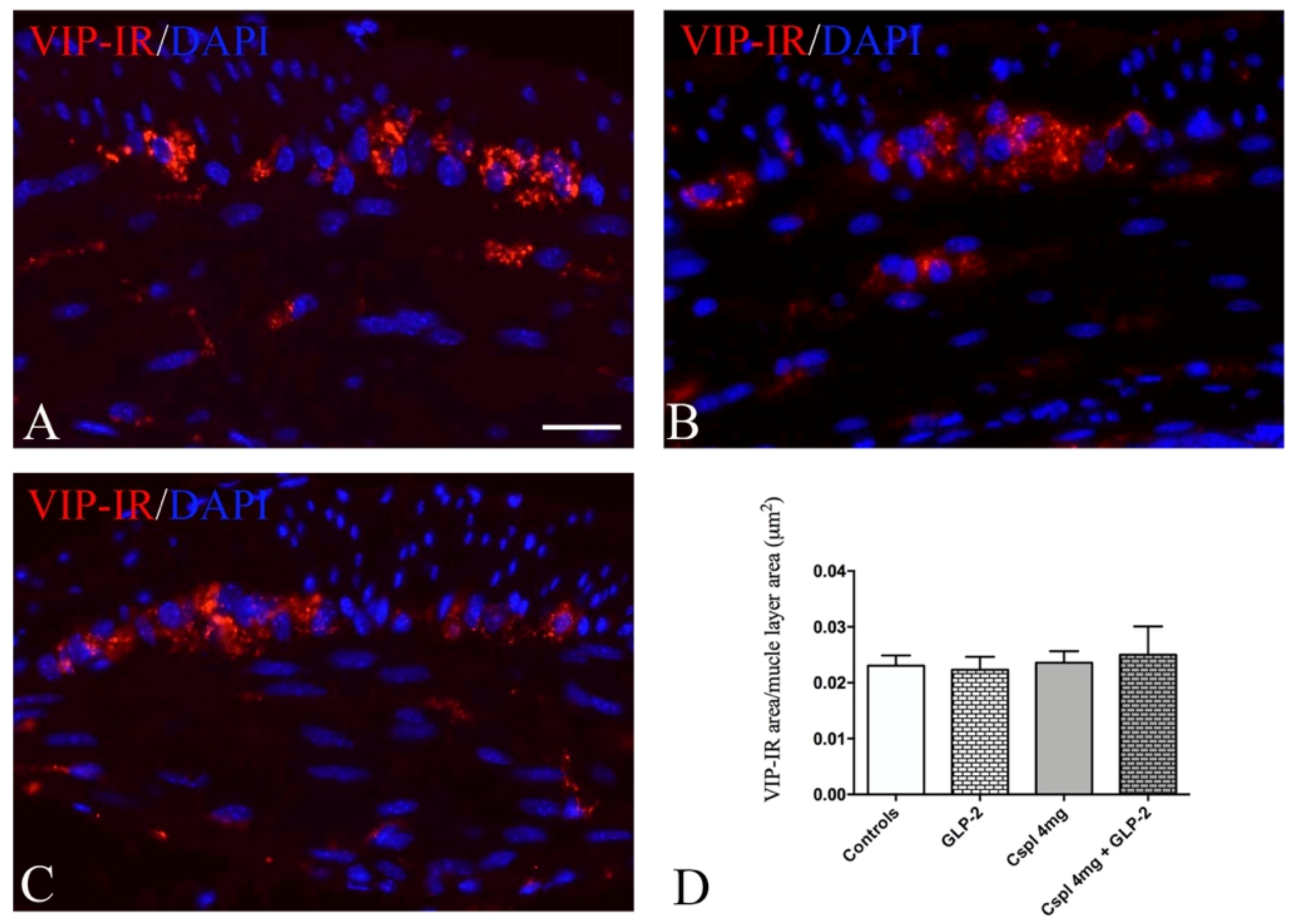
2.3. GLP-2 Significantly Prevents Neuron and Glia Loss in the Myenteric Plexus Induced by Cisplatin in the Distal Colon
3. Discussion
4. Materials and Methods
4.1. Animals Experimental Design
4.2. Tissue Sampling
4.3. Morphological Studies
4.3.1. Histology and Histochemistry
4.3.2. Immunohistochemistry
4.4. Biochemical Study
4.4.1. Cytokine Enzyme-Linked Immune-Absorbent Assay
4.4.2. Data Analysis and Statistical Test
Author Contributions
Funding
Conflicts of Interest
References
- Dilruba, S.; Kalayda, G.V. Platinum-based drugs: Past, present and future. Cancer Chemother. Pharmacol. 2016, 77, 1103–1124. [Google Scholar] [CrossRef] [PubMed]
- Di Fiore, F.; Van Cutsem, E. Acute and long-term gastrointestinal consequences of chemotherapy. Best Pract. Res. Clin. Gastroenterol. 2009, 23, 113–124. [Google Scholar] [CrossRef] [PubMed]
- McQuade, R.M.; Stojanovska, V.; Abalo, R.; Bornstein, J.C.; Nurgali, K. Chemotherapy-Induced Constipation and Diarrhea: Pathophysiology, Current and Emerging Treatments. Front. Pharmacol. 2016, 7, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse effects of cancer chemotherapy: Anything new to improve tolerance and reduce sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef]
- Stojanovska, V.; Sakkal, S.; Nurgali, K. Platinum-based chemotherapy: Gastrointestinal immunomodulation and enteric nervous system toxicity. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G223–G232. [Google Scholar] [CrossRef] [Green Version]
- McQuade, R.M.; Bornstein, J.C.; Nurgali, K. Anti-Colorectal Cancer Chemotherapy-Induced Diarrhoea: Current Treatments and Side-Effects. Int. J. Clin. Med. 2014, 5, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Ishihara, K.; Takeda, Y.; Koizumi, W.; Ichikawa, T. Changes in the mucus barrier during cisplatin-induced intestinal mucositis in rats. BioMed Res. Int. 2013, 2013, 276186. [Google Scholar] [CrossRef]
- Cabezos, P.A.; Vera, G.; Martín-Fontelles, M.I.; Fernández-Pujol, R.; Abalo, R. Cisplatin-induced gastrointestinal dysmotility is aggravated after chronic administration in the rat. Comparison with pica. Neurogastroenterol. Motil. 2010, 22, 797–805. [Google Scholar] [CrossRef]
- Vera, G.; Castillo, M.; Cabezos, P.A.; Chiarlone, A.; Martín, M.I.; Gori, A.; Pasquinelli, G.; Barbara, G.; Stanghellini, V.; Corinaldesi, R.; et al. Enteric neuropathy evoked by repeated cisplatin in the rat. Neurogastroenterol. Motil. 2011, 23, 370–378. [Google Scholar] [CrossRef]
- Wafai, L.; Taher, M.; Jovanovska, V.; Bornstein, J.C.; Dass, C.R.; Nurgali, K. Effects of oxaliplatin on mouse myenteric neurons and colonic motility. Front. Neurosci. 2013, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Robinson, A.M.; Stojanovska, V.; Rahman, A.; McQuade, R.M.; Senior, P.V.; Nurgali, K. Effects of Oxaliplatin Treatment on the Enteric Glial Cells and Neurons in the Mouse Ileum. J. Histochem. Cytochem. 2016, 64, 530–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pini, A.; Garella, R.; Idrizaj, E.; Calosi, L.; Baccari, M.C.; Vannucchi, M.G. Glucagon-like peptide 2 counteracts the mucosal damage and the neuropathy induced by chronic treatment with cisplatin in the mouse gastric fundus. Neurogastroenterol. Motil. 2016, 28, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Uranga, J.A.; García-Martínez, J.M.; García-Jiménez, C.; Vera, G.; Martín-Fontelles, M.I.; Abalo, R. Alterations in the small intestinal wall and motor function after repeated cisplatin in rat. Neurogastroenterol. Motil. 2017, 29, e13047. [Google Scholar] [CrossRef] [PubMed]
- Stojanovska, V.; McQuade, R.M.; Miller, S.; Nurgali, K. Effects of Oxaliplatin Treatment on the Myenteric Plexus Innervation and Glia in the Murine Distal Colon. J. Histochem. Cytochem. 2018, 66, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQuade, R.M.; Stojanovska, V.; Bornstein, J.C.; Nurgali, K. PARP inhibition in platinum-based chemotherapy: Chemopotentiation and neuroprotection. Pharmacol. Res. 2018, 137, 104–113. [Google Scholar] [CrossRef]
- Drucker, D.J.; Yusta, B. Physiology and pharmacology of the enteroendocrine hormone glucagon-like peptide-2. Annu. Rev. Physiol. 2014, 76, 561–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, X.; Karpen, H.E.; Stephens, J.; Bukowski, J.T.; Niu, S.; Zhang, G.; Stoll, B.; Finegold, M.J.; Holst, J.J.; Hadsell, D.L.; et al. GLP-2 receptor localizes to enteric neurons and endocrine cells expressing vasoactive peptides and mediates increased blood flow. Gastroenterology 2006, 130, 150–164. [Google Scholar] [CrossRef]
- Amato, A.; Baldassano, S.; Serio, R.; Mulè, F. Glucagon-like peptide-2 relaxes mouse stomach through vasoactive intestinal peptide release. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G678–G684. [Google Scholar] [CrossRef]
- Amato, A.; Rotondo, A.; Cinci, L.; Baldassano, S.; Vannucchi, M.G.; Mulè, F. Role of cholinergic neurons in the motor effects of glucagon-like peptide-2 in mouse colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G1038–G1044. [Google Scholar] [CrossRef]
- Cinci, L.; Faussone-Pellegrini, M.S.; Rotondo, A.; Mulè, F.; Vannucchi, M.G. GLP-2 receptor expression in excitatory and inhibitory enteric neurons and its role in mouse duodenum contractility. Neurogastroenterol. Motil. 2011, 23, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, J.; Pedersen, N.B.; Brix, S.W.; Grunddal, K.V.; Rosenkilde, M.M.; Hartmann, B.; Orskov, C.; Poulsen, S.S.; Holst, J.J. The glucagon-like peptide 2 receptor is expressed in enteric neurons and not in the epithelium of the intestine. Peptides 2015, 67, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Rowland, K.J.; Brubaker, P.L. The “cryptic” mechanism of action of glucagon-like peptide-2. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G1–G8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldassano, S.; Amato, A. GLP-2: What do we know? What are we going to discover? Regul. Pept. 2014, 194–195, 6–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldassano, S.; Amato, A.; Mulè, F. Influence of glucagon-like peptide 2 on energy homeostasis. Peptides 2016, 86, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Austin, K.; Markovic, M.A.; Brubaker, P.L. Current and potential therapeutic targets of glucagon-like peptide-2. Curr. Opin. Pharmacol. 2016, 31, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; Yusta, B.; Boushey, R.P.; Forest, L.D.E.; Brubaker, P.L.; Daniel, J.; Yusta, B.; Boushey, R.P.; Deforest, L.; Human, P.L.B. Human [Gly 2] GLP-2 reduces the severity of colonic injury in a murine model of experimental colitis. Am. Physiol. Soc. 1999, 276, 79–91. [Google Scholar]
- Tavakkolizadeh, A.; Shen, R.; Abraham, P.; Kormi, N.; Seifert, P.; Edelman, E.R.; Jacobs, D.O.; Zinner, M.J.; Ashley, S.W.; Whang, E.E. Glucagon-like Peptide 2: A New Treatment for Chemotherapy-Induced Enteritis. J. Surg. Res. 2000, 91, 77–82. [Google Scholar] [CrossRef]
- Prasad, R.; Alavi, K.; Schwartz, M.Z. GLP-2 Accelerates Recovery of Mucosal Absorptive Function After Intestinal Ischemia/Reperfusion. J. Pediatr. Surg. 2001, 36, 570–572. [Google Scholar] [CrossRef]
- Sigalet, D.L.; Wallace, L.; De Heuval, E.; Sharkey, K.A. The effects of glucagon-like peptide 2 on enteric neurons in intestinal inflammation. Neurogastroenterol. Motil. 2010, 22, 1318–1325. [Google Scholar] [CrossRef]
- Sigalet, D.L.; Wallace, L.E.; Holst, J.J.; Martin, G.R.; Kaji, T.; Tanaka, H.; Sharkey, K.A. Enteric neural pathways mediate the anti-inflammatory actions of glucagon-like peptide 2. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G211–G221. [Google Scholar] [CrossRef]
- Jeppesen, P.B. Teduglutide, a novel glucagon-like peptide 2 analog, in the treatment of patients with short bowel syndrome. Therap. Adv. Gastroenterol. 2012, 5, 159–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boushey, R.P.; Yusta, B.; Drucker, D.J. Glucagon-like peptide (GLP)-2 reduces chemotherapy-associated mortality and enhances cell survival in cells expressing a transfected GLP-2 receptor. Cancer Res. 2001, 61, 687–693. [Google Scholar] [PubMed]
- Voss, U.; Sand, E.; Hellström, P.M.; Ekblad, E. Glucagon-like peptides 1 and 2 and vasoactive intestinal peptide are neuroprotective on cultured and mast cell co-cultured rat myenteric neurons. BMC Gastroenterol. 2012, 12, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang-Christensen, M.; Vrang, N.; Larsen, P.J. Glucagon-like peptide containing pathways in the regulation of feeding behaviour. Int. J. Obes. 2001, 25, S42–S47. [Google Scholar] [CrossRef]
- Keefe, D.M.K. Mucositis management in patients with cancer. Support. Cancer Ther. 2006, 3, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Kissow, H.; Viby, N.E.; Hartmann, B.; Holst, J.J.; Timm, M.; Thim, L.; Poulsen, S.S. Exogenous glucagon-like peptide-2 (GLP-2) prevents chemotherapy-induced mucositis in rat small intestine. Cancer Chemother. Pharmacol. 2012, 70, 39–48. [Google Scholar] [CrossRef]
- Kaji, T.; Tanaka, H.; Holst, J.J.; Redstone, H.; Wallace, L.; de Heuval, E.; Sigalet, D.L. The effects of variations in dose and method of administration on glucagon like peptide-2 activity in the rat. Eur. J. Pharmacol. 2008, 596, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, A.R.; Viby, N.E.; Hare, K.J.; Hartmann, B.; Thim, L.; Holst, J.J.; Poulsen, S.S. The intestinotrophic peptide, GLP-2, counteracts the gastrointestinal atrophy in mice induced by the epidermal growth factor receptor inhibitor, Erlotinib, and cisplatin. Dig. Dis. Sci. 2010, 55, 2785–2796. [Google Scholar] [CrossRef]
- El-jamal, N.; Erdual, E.; Neunlist, M.; Koriche, D.; Dubuquoy, C.; Maggiotto, F.; Chevalier, J.; Berrebi, D.; Dubuquoy, L.; Boulanger, E.; et al. Glugacon-like peptide-2: Broad receptor expression, limited therapeutic effect on intestinal inflammation and novel role in liver regeneration. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G274–G285. [Google Scholar] [CrossRef] [Green Version]
- de Heuvel, E.; Wallace, L.; Sharkey, K.A.; Sigalet, D.L. Glucagon-like peptide 2 induces vasoactive intestinal polypeptide expression in enteric neurons via phophatidylinositol 3-kinase- signaling. AJP Endocrinol. Metab. 2012, 303, E994–E1005. [Google Scholar] [CrossRef] [Green Version]
- Ekblad, E.; Bauer, A.J. Role of vasoactive intestinal peptide and inflammatory mediators in enteric neuronal plasticity. Neurogastroenterol. Motil. 2004, 16, 123–128. [Google Scholar] [CrossRef] [PubMed]
- McQuade, R.M.; Carbone, S.E.; Stojanovska, V.; Rahman, A.; Gwynne, R.M.; Robinson, A.M.; Goodman, C.A.; Bornstein, J.C.; Nurgali, K. Role of oxidative stress in oxaliplatin-induced enteric neuropathy and colonic dysmotility in mice. Br. J. Pharmacol. 2016, 173, 3502–3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, S.E.; Jovanovska, V.; Brookes, S.J.H.; Nurgali, K. Electrophysiological and morphological changes in colonic myenteric neurons from chemotherapy-treated patients: A pilot study. Neurogastroenterol. Motil. 2016, 28, 975–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boussios, S.; Pentheroudakis, G.; Katsanos, K.; Pavlidis, N. Systemic treatment-induced gastrointestinal toxicity: Incidence, clinical presentation and management. Ann. Gastroenterol. 2012, 25, 106–118. [Google Scholar] [PubMed]
- McQuade, R.M.; Stojanovska, V.; Donald, E.L.; Rahman, A.A.; Campelj, D.G.; Abalo, R.; Rybalka, E.; Bornstein, J.C.; Nurgali, K. Irinotecan-induced gastrointestinal dysfunction is associated with enteric neuropathy, but increased numbers of cholinergic myenteric neurons. Front. Physiol. 2017, 8, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQuade, R.M.; Stojanovska, V.; Donald, E.; Abalo, R.; Bornstein, J.C.; Nurgali, K. Gastrointestinal dysfunction and enteric neurotoxicity following treatment with anticancer chemotherapeutic agent 5-fluorouracil. Neurogastroenterol. Motil. 2016, 28, 1861–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojanovska, V.; McQuade, R.; Rybalka, E.; Nurgali, K. Neurotoxicity Associated with Platinum-Based Anti-Cancer Agents: What are the Implications of Copper Transporters? Curr. Med. Chem. 2017, 24, 1520–1536. [Google Scholar] [CrossRef]
- Garella, R.; Idrizaj, E.; Traini, C.; Squecco, R.; Vannucchi, M.G.; Baccari, M.C. Glucagon-like peptide-2 modulates the nitrergic neurotransmission in strips from the mouse gastric fundus. World J. Gastroenterol. 2017, 23, 7211–7220. [Google Scholar] [CrossRef]
- Sandgren, K.; Lin, Z.; Svenningsen, Å.F.; Ekblad, E. Vasoactive Intestinal Peptide and Nitric Oxide Promote Survival of Adult Rat Myenteric Neurons in Culture. Evaluation 2003, 602, 595–602. [Google Scholar] [CrossRef]
- Lipton, S.A. Neuronal protection and destruction by NO. Cell Death Differ. 1999, 6, 943–951. [Google Scholar] [CrossRef]
- Boesmans, W.; Lasrado, R.; Vanden Berghe, P.; Pachnis, V. Heterogeneity and phenotypic plasticity of glial cells in the mammalian enteric nervous system. Glia 2015, 63, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Giorgio, R.; Giancola, F.; Boschetti, E.; Abdo, H.; Lardeux, B.; Neunlist, M. Enteric glia and neuroprotection: Basic and clinical aspects. AJP Gastrointest. Liver Physiol. 2012, 303, G887–G893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharkey, K.A. Emerging roles for enteric glia in gastrointestinal disorders. J. Clin. Investig. 2015, 125, 918–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neunlist, M.; Rolli-Derkinderen, M.; Latorre, R.; Van Landeghem, L.; Coron, E.; Derkinderen, P.; De Giorgio, R. Enteric glial cells: Recent developments and future directions. Gastroenterology 2014, 147, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Bassotti, G.; Villanacci, V.; Fisogni, S.; Rossi, E.; Baronio, P.; Clerici, C.; Maurer, C.A.; Cathomas, G.; Antonelli, E. Enteric glial cells and their role in gastrointestinal motor abnormalities: Introducing the neuro-gliopathies. World J. Gastroenterol. 2007, 13, 4035–4041. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, P.B.; Gilroy, R.; Pertkiewicz, M.; Allard, J.P.; Messing, B.; O’Keefe, S.J. Randomised placebo-controlled trial of teduglutide in reducing parenteral nutrition and/or intravenous fluid requirements in patients with short bowel syndrome. Gut 2011, 60, 902–914. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Species | Source | Concentration |
|---|---|---|---|
| Primary Antisera | |||
| nNOS | Rabbit | Millipore (Bedford, MA, USA) | 1:2000 |
| NeuN | Mouse | Millipore | 1:200 |
| ChAT | Goat | Millipore | 1:200 |
| VIP | Mouse | Santa Cruz Biotech (Santa Cruz, CA, USA) | 1:200 |
| SOX-10 | Goat | SantaCruz | 1:500 |
| HuCD | Rabbit | SantaCruz | 1:200 |
| S100β | Rabbit | Dako (Dako, Santa Clara, CA, USA) | Ready to use |
| GFAP | Mouse | Sigma (St Louis, MO, USA) | 1:300 |
| Secondary Antisera | |||
| Alexa Fluor 488 | Goat | Invitrogen (Carlsbad, CA, USA) | 1:333 |
| Alexa Fluor 594 | Mouse | Jackson ImmunoResearch (Ely, Cambridgeshire, UK) | 1:333 |
| Alexa Fluor 488 | Rabbit | Jackson ImmunoResearch | 1:333 |
| Anti-mouse C3 | Donkey | Jackson ImmunoResearch | 1:500 |
| Anti-rabbit-FP488 | Donkey | Interchim (Montluçon, France) | 1:200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nardini, P.; Pini, A.; Bessard, A.; Duchalais, E.; Niccolai, E.; Neunlist, M.; Vannucchi, M.G. GLP-2 Prevents Neuronal and Glial Changes in the Distal Colon of Mice Chronically Treated with Cisplatin. Int. J. Mol. Sci. 2020, 21, 8875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228875
Nardini P, Pini A, Bessard A, Duchalais E, Niccolai E, Neunlist M, Vannucchi MG. GLP-2 Prevents Neuronal and Glial Changes in the Distal Colon of Mice Chronically Treated with Cisplatin. International Journal of Molecular Sciences. 2020; 21(22):8875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228875
Chicago/Turabian StyleNardini, Patrizia, Alessandro Pini, Anne Bessard, Emilie Duchalais, Elena Niccolai, Michel Neunlist, and Maria Giuliana Vannucchi. 2020. "GLP-2 Prevents Neuronal and Glial Changes in the Distal Colon of Mice Chronically Treated with Cisplatin" International Journal of Molecular Sciences 21, no. 22: 8875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228875