Compounds with Antiviral, Anti-Inflammatory and Anticancer Activity Identified in Wine from Hungary’s Tokaj Region via High Resolution Mass Spectrometry and Bioinformatics Analyses

 ,
,  , , and
, , and
Abstract
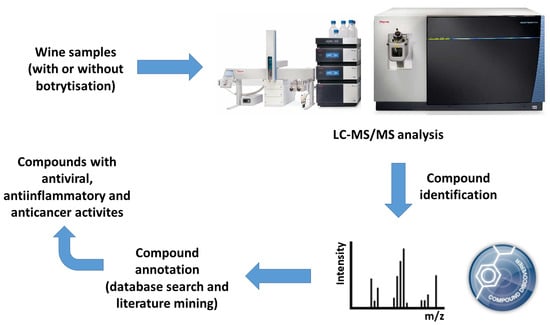
:
1. Introduction
2. Results
2.1. Biological Roles of the Identified Wine Compounds
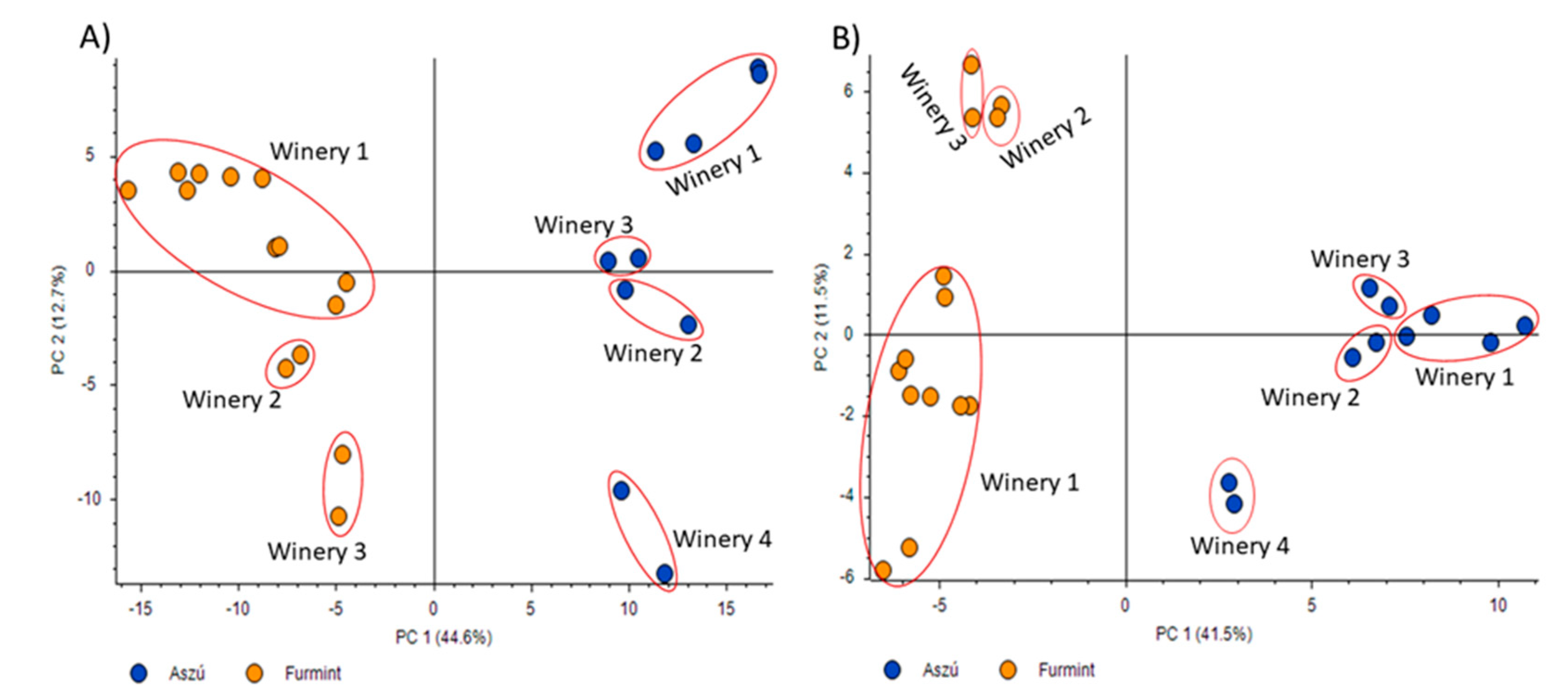
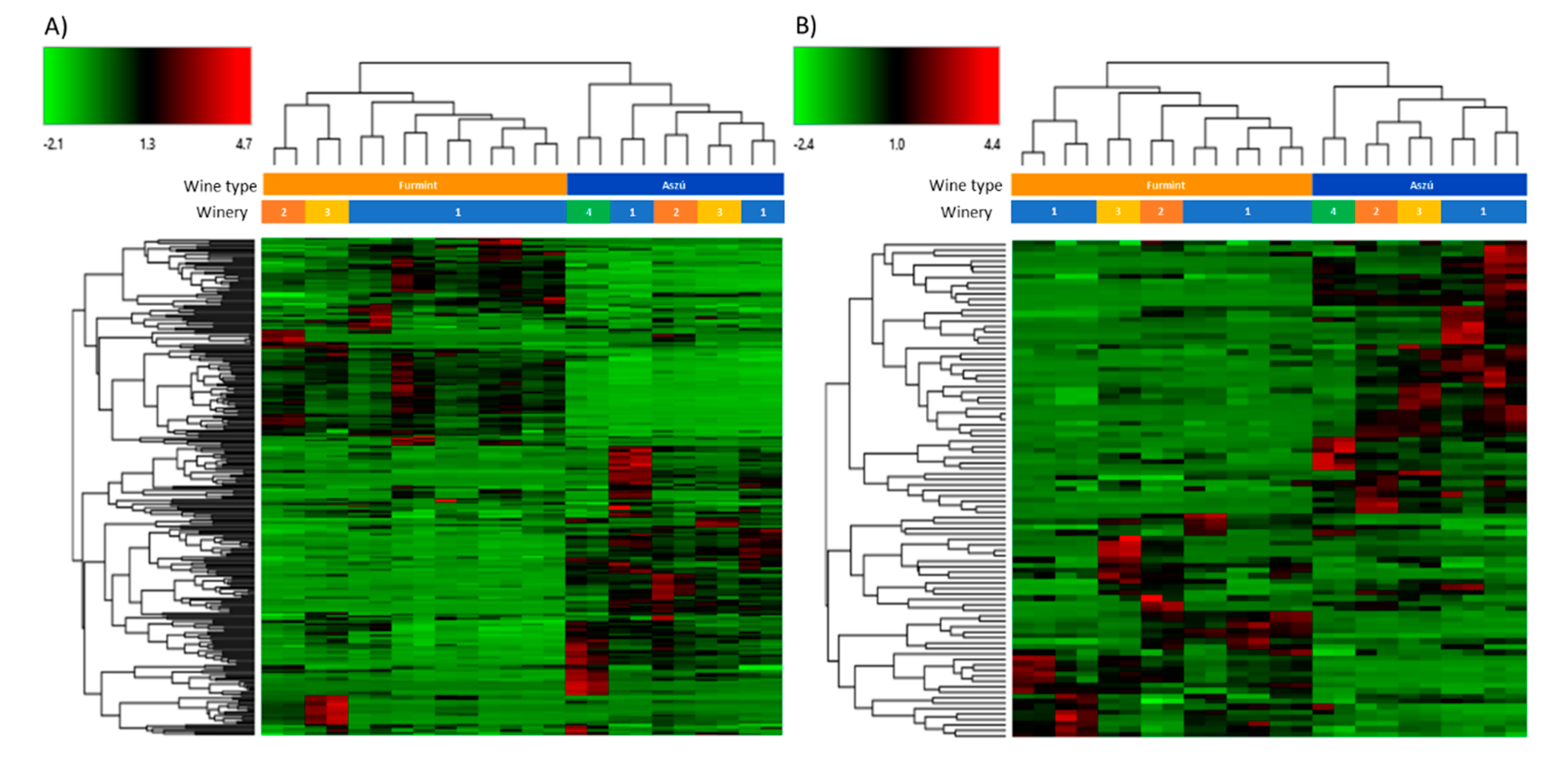
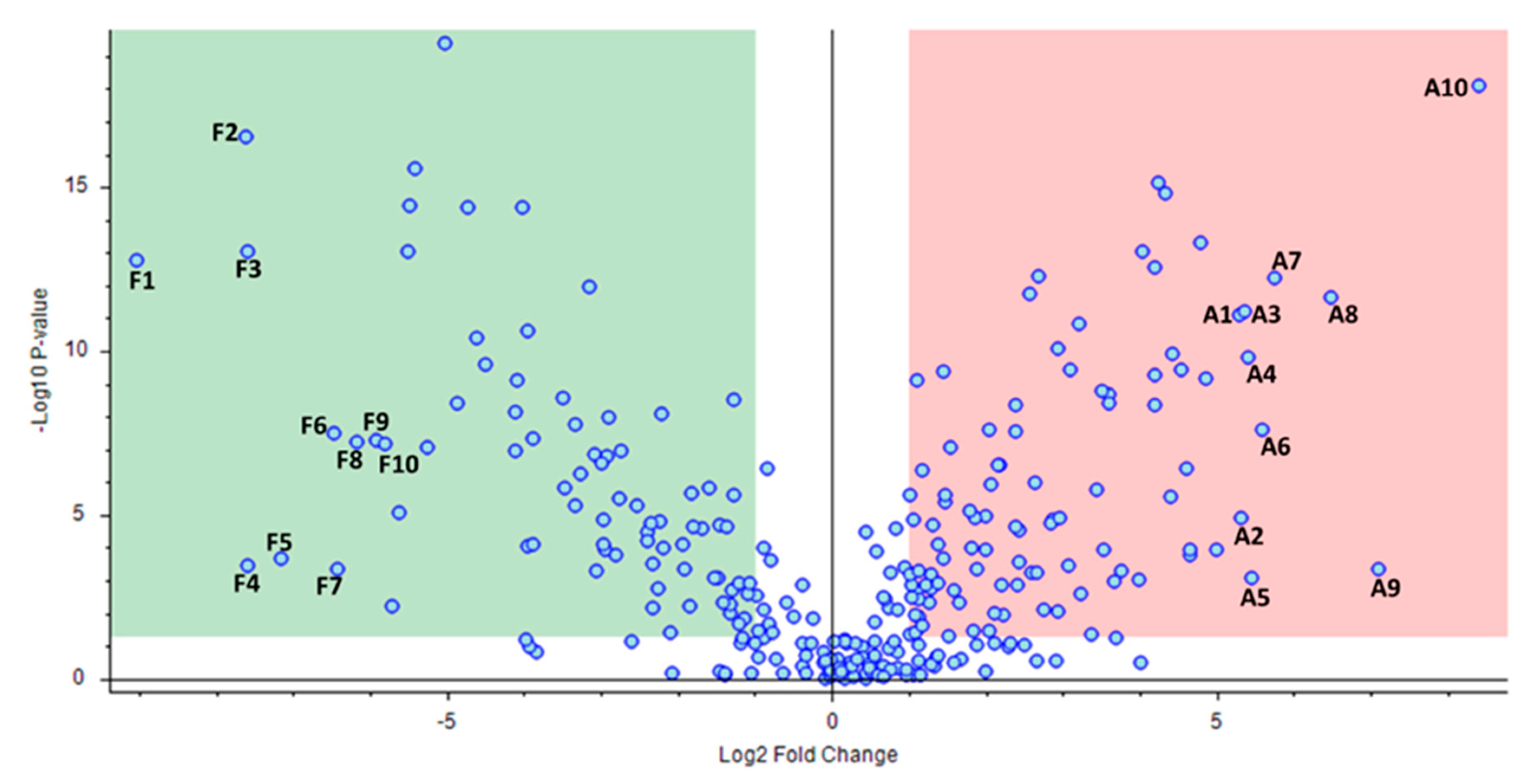
2.2. Comparative Analysis of “aszú” and “furmint” Wines
3. Discussion
4. Materials and Methods
4.1. Wine Samples
4.2. LC-MS Analysis
4.3. Data Analysis and Compound Identification
4.4. Annotation of the Identified Compounds
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| UNESCO | United Nations Educational, Scientific and Cultural Organization |
| NMR | Nuclear magnetic resonance |
| GC-MS | Gas chromatography-mass spectrometry |
| LC-MS | Liquid chromatography-mass spectrometry |
| MS/MS | Tandem mass spectrometry |
| FDA | Food and Drug Administration |
| LC-MS/MS | Liquid chromatography-tandem mass spectrometry |
| SARS-CoV | Severe acute respiratory syndrome coronavirus 2 |
| COX-2 | Cyclooxygenase-2 |
| NF-κB | Nuclear factor-κB |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin-6 |
| IL-8 | Interleunik-8 |
| TNF-α | Tumor necrosis factor α |
| INF-γ | Interferon γ |
| NO | Nitrogen monoxide |
| Nrf2 | Nuclear factor erythroid 2–related factor 2 |
| CID | PubChem compound identifier |
References
- Wu, C.D. Grape Products and Oral Health. J. Nutr. 2009, 139, 1818S–1823S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weseler, A.R.; Bast, A. Masquelier’s grape seed extract: From basic flavonoid research to a well-characterized food supplement with health benefits. Nutr. J. 2017, 16, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirmala, J.G.; Narendhirakannan, R.T. Vitis vinifera peel and seed gold nanoparticles exhibit chemopreventive potential, antioxidant activity and induce apoptosis through mutant p53, Bcl-2 and pan cytokeratin down-regulation in experimental animals. Biomed. Pharmacother. 2017, 89, 902–917. [Google Scholar] [CrossRef] [PubMed]
- Zunino, S.J. Type 2 Diabetes and Glycemic Response to Grapes or Grape Products. J. Nutr. 2009, 139, 1794S–1800S. [Google Scholar] [CrossRef] [Green Version]
- Rasines-Perea, Z.; Teissedre, P.L. Grape Polyphenols’ effects in human cardiovascular diseases and diabetes. Molecules 2017, 22, 68. [Google Scholar] [CrossRef]
- Baiano, A.; Scrocco, C.; Sepielli, G.; Del Nobile, M.A. Wine Processing: A Critical Review of Physical, Chemical, and Sensory Implications of Innovative Vinification Procedures. Crit. Rev. Food Sci. Nutr. 2016, 56, 2391–2407. [Google Scholar] [CrossRef]
- Diamantidou, D.; Zotou, A.; Theodoridis, G. Wine and grape marc spirits metabolomics. Metabolomics 2018, 14, 159. [Google Scholar] [CrossRef]
- Zaukuu, J.L.Z.; Soós, J.; Bodor, Z.; Felföldi, J.; Magyar, I.; Kovacs, Z. Authentication of Tokaj Wine (Hungaricum) with the Electronic Tongue and Near Infrared Spectroscopy. J. Food Sci. 2019, 84, 3437–3444. [Google Scholar] [CrossRef]
- Bodor, P.; Szoke, A.; Toth-Lencses, K.; Veres, A.; Deak, T.; Kozma, P.; Bisztray, G.D.; Kiss, E. Differentiation of grapevine (Vitis vinifera L.) conculta members based on molecular tools. Biotechnol. Biotechnol. Equip. 2014, 28, 14–20. [Google Scholar] [CrossRef]
- Magyar, I.; Soós, J. Botrytized wines—Current perspectives. Int. J. Wine Res. 2016, 8, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Magyar, I. Botrytized wines. In Advances in Food and Nutrition Research; Academic Press Inc.: Cambridge, MA, USA, 2011; Volume 63, pp. 147–206. [Google Scholar]
- Yoo, Y.J.; Saliba, A.J.; Prenzler, P.D. Should Red Wine Be Considered a Functional Food? Compr. Rev. Food Sci. Food Saf. 2010, 9, 530–551. [Google Scholar] [CrossRef]
- Haseeb, S.; Alexander, B.; Baranchuk, A. Wine and Cardiovascular Health. Circulation 2017, 136, 1434–1448. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, L.; Narváez, A.; Izzo, L.; Graziani, G.; Gaspari, A.; Di Minno, G.; Ritieni, A. Red wine consumption and cardiovascular health. Molecules 2019, 24, 3626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigo, R.; Miranda, A.; Vergara, L. Modulation of endogenous antioxidant system by wine polyphenols in human disease. Clin. Chim. Acta 2011, 412, 410–424. [Google Scholar] [CrossRef]
- Cueva, C.; Gil-Sánchez, I.; Ayuda-Durán, B.; González-Manzano, S.; González-Paramás, A.M.; Santos-Buelga, C.; Bartolomé, B.; Moreno-Arribas, M.V. An Integrated View of the Effects of Wine Polyphenols and Their Relevant Metabolites on Gut and Host Health. Molecules 2017, 22, 99. [Google Scholar] [CrossRef]
- Fernandes, I.; Pérez-Gregorio, R.; Soares, S.; Mateus, N.; De Freitas, V.; Santos-Buelga, C.; Feliciano, A.S. Wine flavonoids in health and disease prevention. Molecules 2017, 22, 292. [Google Scholar] [CrossRef]
- Hong, Y.S. NMR-based metabolomics in wine science. Magn. Reson. Chem. 2011, 49, S13–S21. [Google Scholar] [CrossRef]
- Amargianitaki, M.; Spyros, A. NMR-based metabolomics in wine quality control and authentication. Chem. Biol. Technol. Agric. 2017, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Fournié-Zaluski, M.C.; Soroca-Lucas, E.; Roques, B.P.; Chaillet, P.; Margais-Collado, H.; Costentin, J. New Carboxyalkyl Inhibitors of Brain Enkephalinase: Synthesis, Biological Activity, and Analgesic Properties. J. Med. Chem. 1983, 26, 60–65. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Database. Available online: https://pubchem.ncbi.nlm.nih.gov/bioassay/1565#sid=7978036 (accessed on 5 May 2020).
- Glennon, R.A.; Liebowitz, S.M. Serotonin Receptor Affinity of Cathinone and Related Analogues. J. Med. Chem. 1982, 25, 393–397. [Google Scholar] [CrossRef]
- Glennon, R.A.; Liebowitz, S.M.; Anderson, G.M. Serotonin Receptor Affinities of Psychoactive Phenalkylamine Analogues. J. Med. Chem. 1980, 23, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Rahnasto, M.; Raunio, H.; Poso, A.; Wittekindt, C.; Juvonen, R.O. Quantitative structure-activity relationship analysis of inhibitors of the nicotine metabolizing CYP2A6 enzyme. J. Med. Chem. 2005, 48, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Lewin, A.H.; Navarro, H.A.; Wayne Mascarella, S. Structure-activity correlations for β-phenethylamines at human trace amine receptor 1. Bioorg. Med. Chem. 2008, 16, 7415–7423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Center for Biotechnology Information. PubChem Database. Source=The Scripps Research Institute Molecular Screening Center, AID=793. Available online: https://pubchem.ncbi.nlm.nih.gov/bioassay/793 (accessed on 5 May 2020).
- National Center for Biotechnology Information. PubChem Database. Source=The Scripps Research Institute Molecular Screening Center, AID=1040. Available online: https://pubchem.ncbi.nlm.nih.gov/bioassay/1040 (accessed on 5 May 2020).
- National Center for Biotechnology Information. PubChem Database. Source=1043, AID=1159608. Available online: https://pubchem.ncbi.nlm.nih.gov/bioassay/1159608 (accessed on 5 May 2020).
- National Center for Biotechnology Information. PubChem Database. Source=NCGC, AID=938. Available online: https://pubchem.ncbi.nlm.nih.gov/bioassay/938 (accessed on 5 May 2020).
- National Center for Biotechnology Information. PubChem Database. Source=NCGC, AID=893. Available online: https://pubchem.ncbi.nlm.nih.gov/bioassay/893 (accessed on 5 May 2020).
- Aung, H.T.; Furukawa, T.; Nikai, T.; Niwa, M.; Takaya, Y. Contribution of cinnamic acid analogues in rosmarinic acid to inhibition of snake venom induced hemorrhage. Bioorg. Med. Chem. 2011, 19, 2392–2396. [Google Scholar] [CrossRef]
- Bagnardi, V.; Rota, M.; Botteri, E.; Tramacere, I.; Islami, F.; Fedirko, V.; Scotti, L.; Jenab, M.; Turati, F.; Pasquali, E.; et al. Alcohol consumption and site-specific cancer risk: A comprehensive dose-response meta-analysis. Br. J. Cancer 2015, 112, 580–593. [Google Scholar] [CrossRef]
- Piano, M.R. Alcohol’s Effects on the Cardiovascular System. Alcohol Res. 2017, 38, 219–241. [Google Scholar]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. Effect of Maturity and Vine Water Status on Grape Skin and Wine Flavonoids. Am. J. Enol. Vitic. 2002, 53, 268–274. [Google Scholar]
- Bavinger, J.C.; Shantha, J.G.; Yeh, S. Ebola, COVID-19, and emerging infectious disease: Lessons learned and future preparedness. Curr. Opin. Ophthalmol. 2020, 31, 416–422. [Google Scholar] [CrossRef]
- Lin, L.T.; Hsu, W.C.; Lin, C.C. Antiviral natural products and herbal medicines. J. Tradit. Complement. Med. 2014, 4, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Konowalchuk, J.; Speirs, J.I. Antiviral activity of fruit extracts. J. Food Sci. 1976, 41, 1013–1017. [Google Scholar] [CrossRef]
- Nikolaeva-Glomb, L.; Mukova, L.; Nikolova, N.; Badjakov, I.; Dincheva, I.; Kondakova, V.; Doumanova, L.; Galabov, A.S. In vitro antiviral activity of a series of wild berry fruit extracts against representatives of Picorna-, Orthomyxo- and Paramyxoviridae. Nat. Prod. Commun. 2014, 9, 51–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danaher, R.J.; Wang, C.; Dai, J.; Mumper, R.J.; Miller, C.S. Antiviral effects of blackberry extract against herpes simplex virus type 1. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2011, 112, e31–e35. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.R.; Gu, Q.; Wang, Y.H.; Zhang, X.M.; Yang, L.M.; Zhou, J.; Chen, J.J.; Zheng, Y.T. Anti-HIV-1 activities of compounds isolated from the medicinal plant Rhus chinensis. J. Ethnopharmacol. 2008, 117, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Byung, S.M.; Hyeong, K.L.; Sang, M.L.; Young, H.K.; Ki, H.B.; Otake, T.; Nakamura, N.; Hattori, M. Anti-human immunodeficiency virus-type 1 activity of constituents from Juglans mandshurica. Arch. Pharm. Res. 2002, 25, 441–445. [Google Scholar] [CrossRef]
- Pu, J.X.; Yang, L.M.; Xiao, W.L.; Li, R.T.; Lei, C.; Gao, X.M.; Huang, S.X.; Li, S.H.; Zheng, Y.T.; Huang, H.; et al. Compounds from Kadsura heteroclita and related anti-HIV activity. Phytochemistry 2008, 69, 1266–1272. [Google Scholar] [CrossRef]
- Yang, X.; Zhu, X.; Ji, H.; Deng, J.; Lu, P.; Jiang, Z.; Li, X.; Wang, Y.; Wang, C.; Zhao, J.; et al. Quercetin synergistically reactivates human immunodeficiency virus type 1 latency by activating nuclear factor-κB. Mol. Med. Rep. 2018, 17, 2501–2508. [Google Scholar] [CrossRef]
- Mahmood, N.; Piacente, S.; Pizza, C.; Burke, A.; Khan, A.I.; Hayt, A.J. The anti-HIV activity and mechanisms of action of pure compounds isolated from Rosa damascena. Biochem. Biophys. Res. Commun. 1996, 229, 73–79. [Google Scholar] [CrossRef]
- Esposito, F.; Sanna, C.; Del Vecchio, C.; Cannas, V.; Venditti, A.; Corona, A.; Bianco, A.; Serrilli, A.M.; Guarcini, L.; Parolin, C.; et al. Hypericum hircinum L. components as new single-molecule inhibitors of both HIV-1 reverse transcriptase-associated DNA polymerase and ribonuclease H activities. Pathog. Dis. 2013, 68, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Tamura, H.; Akioka, T.; Ueno, K.; Chujyo, T.; Okazaki, K.; King, P.J.; Robinson, W.E. Anti-human immunodeficiency virus activity of 3,4,5-tricaffeoylquinic acid in cultured cells of lettuce leaves. Mol. Nutr. Food Res. 2006, 50, 396–400. [Google Scholar] [CrossRef]
- Xia, C.-N.; Li, H.-B.; Liu, F.; Hu, W.-X. Synthesis of trans-caffeate analogues and their bioactivities against HIV-1 integrase and cancer cell lines. Bioorg. Med. Chem. Lett. 2008, 18, 6553–6557. [Google Scholar] [CrossRef] [PubMed]
- Biziagos, E.; Crance, J.-M.; Passagot, J.; Deloince, R. Effect of antiviral substances on hepatitis A virus replication in vitro. J. Med. Virol. 1987, 22, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Khachatoorian, R.; Arumugaswami, V.; Raychaudhuri, S.; Yeh, G.K.; Maloney, E.M.; Wang, J.; Dasgupta, A.; French, S.W. Divergent antiviral effects of bioflavonoids on the hepatitis C virus life cycle. Virology 2012, 433, 346–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Sun, G.; Guo, W.; Huang, Y.; Sun, W.; Zhao, F.; Hu, K. Inhibition of hepatitis B virus replication by quercetin in human hepatoma cell lines. Virol. Sin. 2015, 30, 261–268. [Google Scholar] [CrossRef]
- Wang, G.F.; Shi, L.P.; Ren, Y.D.; Liu, Q.F.; Liu, H.F.; Zhang, R.J.; Li, Z.; Zhu, F.H.; He, P.L.; Tang, W.; et al. Anti-hepatitis B virus activity of chlorogenic acid, quinic acid and caffeic acid in vivo and in vitro. Antivir. Res. 2009, 83, 186–190. [Google Scholar] [CrossRef]
- Shen, H.; Yamashita, A.; Nakakoshi, M.; Yokoe, H.; Sudo, M.; Kasai, H.; Tanaka, T.; Fujimoto, Y.; Ikeda, M.; Kato, N.; et al. Inhibitory Effects of Caffeic Acid Phenethyl Ester Derivatives on Replication of Hepatitis C Virus. PLoS ONE 2013, 8, e82299. [Google Scholar] [CrossRef]
- Zhao, Y.; Geng, C.A.; Sun, C.L.; Ma, Y.B.; Huang, X.Y.; Cao, T.W.; He, K.; Wang, H.; Zhang, X.M.; Chen, J.J. Polyacetylenes and anti-hepatitis B virus active constituents from Artemisia capillaris. Fitoterapia 2014, 95, 187–193. [Google Scholar] [CrossRef]
- Chen, C.; Jiang, Z.Y.; Yu, B.; Wu, X.L.; Dai, C.Q.; Zhao, C.L.; Ju, D.H.; Chen, X.Y. Study on the anti-H1N1 virus effects of quercetin and oseltamivir and their mechanism related to TLR7 pathway. J. Asian Nat. Prod. Res. 2012, 14, 877–885. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, J.; Li, W.; Wang, X.; Xu, J.; Xie, J.; Tao, K.; Shen, L.; Zhang, R. Molecular docking of potential inhibitors for influenza H7N9. Comput. Math. Methods Med. 2015, 2015, 480764. [Google Scholar] [CrossRef]
- Ibrahim, A.K.; Youssef, A.I.; Arafa, A.S.; Ahmed, S.A. Anti-H5N1 virus flavonoids from Capparis sinaica Veill. Nat. Prod. Res. 2013, 27, 2149–2153. [Google Scholar] [CrossRef]
- Davis, J.M.; Murphy, E.A.; McClellan, J.L.; Carmichael, M.D.; Gangemi, J.D. Quercetin reduces susceptibility to influenza infection following stressful exercise. Am. J. Physiol. Integr. Comp. Physiol. 2008, 295, R505–R509. [Google Scholar] [CrossRef]
- Boon, A.C.M.; Vos, A.P.; Graus, Y.M.F.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E. In vitro Effect of Bioactive Compounds on Influenza Virus Specific B- and T-Cell Responses. Scand. J. Immunol. 2002, 55, 24–32. [Google Scholar] [CrossRef]
- Gamaleldin Elsadig Karar, M.; Matei, M.F.; Jaiswal, R.; Illenberger, S.; Kuhnert, N. Neuraminidase inhibition of Dietary chlorogenic acids and derivatives-potential antivirals from dietary sources. Food Funct. 2016, 7, 2052–2059. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Cao, Z.; Cao, L.; Ding, G.; Wang, Z.; Xiao, W. Antiviral activity of chlorogenic acid against influenza A (H1N1/H3N2) virus and its inhibition of neuraminidase. Sci. Rep. 2017, 7, 45723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, L.C.; Chiang, W.; Liu, M.C.; Lin, C.C. In vitro antiviral activities of Caesalpinia pulcherrima and its related flavonoids. J. Antimicrob. Chemother. 2003, 52, 194–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, L.C.; Chiang, W.; Chang, M.Y.; Ng, L.T.; Lin, C.C. Antiviral activity of Plantago major extracts and related compounds in vitro. Antivir. Res. 2002, 55, 53–62. [Google Scholar] [CrossRef]
- Guo, Y.J.; Luo, T.; Wu, F.; Mei, Y.W.; Peng, J.; Liu, H.; Li, H.R.; Zhang, S.L.; Dong, J.H.; Fang, Y.; et al. Involvement of TLR2 and TLR9 in the anti-inflammatory effects of chlorogenic acid in HSV-1-infected microglia. Life Sci. 2015, 127, 12–18. [Google Scholar] [CrossRef]
- Kratz, J.M.; Andrighetti-Fröhner, C.R.; Kolling, D.J.; Leal, P.C.; Cirne-Santos, C.C.; Yunes, R.A.; Nunes, R.J.; Trybala, E.; Bergström, T.; Frugulhetti, I.C.P.P.; et al. Anti-HSV-1 and anti-HIV-1 activity of gallic acid and pentyl gallate. Mem. Inst. Oswaldo Cruz 2008, 103, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Xi, C.; Hu, K.; Gao, W.; Cai, X.; Qin, J.; Lv, S.; Du, C.; Wei, Y. Inhibition of enterovirus 71 replication and viral 3C protease by quercetin. Virol. J. 2018, 15, 116. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, Y.; Hou, X.; Peng, H.; Zhang, L.; Jiang, Q.; Shi, M.; Ji, Y.; Wang, Y.; Shi, W. Chlorogenic Acid Inhibits the Replication and Viability of Enterovirus 71 In Vitro. PLoS ONE 2013, 8, e76007. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Li, J.; Luo, C.; Liu, H.; Xu, W.; Chen, G.; Liew, O.W.; Zhu, W.; Puah, C.M.; Shen, X.; et al. Binding interaction of quercetin-3-β-galactoside and its synthetic derivatives with SARS-CoV 3CLpro: Structure-activity relationship studies reveal salient pharmacophore features. Bioorg. Med. Chem. 2006, 14, 8295–8306. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Faris, A.N.; Comstock, A.T.; Wang, Q.; Nanua, S.; Hershenson, M.B.; Sajjan, U.S. Quercetin inhibits rhinovirus replication in vitro and in vivo. Antivir. Res. 2012, 94, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Son, M.; Ryu, E.; Shin, Y.S.; Kim, J.G.; Kang, B.W.; Cho, H.; Kang, H. Quercetin-induced apoptosis prevents EBV infection. Oncotarget 2015, 6, 12603–12624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galochkina, A.V.; Anikin, V.B.; Babkin, V.A.; Ostrouhova, L.A.; Zarubaev, V.V. Virus-inhibiting activity of dihydroquercetin, a flavonoid from Larix sibirica, against coxsackievirus B4 in a model of viral pancreatitis. Arch. Virol. 2016, 161, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Parang, K.; Wiebe, L.I.; Knaus, E.E.; Huang, J.S.; Tyrrell, D.L.; Csizmadia, F. In vitro antiviral activities of myristic acid analogs against human immunodeficiency and hepatitis B viruses. Antivir. Res. 1997, 34, 75–90. [Google Scholar] [CrossRef]
- Bourinbaiar, A.S.; Tan, X.; Nagorny, R. Inhibitory effect of coumarins on HIV-1 replication and cell-mediated or cell-free viral transmission. Acta Virol. 1993, 37, 241–250. [Google Scholar]
- Chen, H.C.; Chou, C.K.; Lee, S.D.; Wang, J.C.; Yeh, S.F. Active compounds from Saussurea lappa Clarks that suppress hepatitis B virus surface antigen gene expression in human hepatoma cells. Antivir. Res. 1995, 27, 99–109. [Google Scholar] [CrossRef]
- Wang, Y.H.; Tang, J.G.; Wang, R.R.; Yang, L.M.; Dong, Z.J.; Du, L.; Shen, X.; Liu, J.K.; Zheng, Y.T. Flazinamide, a novel β-carboline compound with anti-HIV actions. Biochem. Biophys. Res. Commun. 2007, 355, 1091–1095. [Google Scholar] [CrossRef]
- Anderson, G.; Maes, M.; Markus, R.P.; Rodriguez, M. Ebola virus: Melatonin as a readily available treatment option. J. Med. Virol. 2015, 87, 537–543. [Google Scholar] [CrossRef]
- Cicchese, J.M.; Evans, S.; Hult, C.; Joslyn, L.R.; Wessler, T.; Millar, J.A.; Marino, S.; Cilfone, N.A.; Mattila, J.T.; Linderman, J.J.; et al. Dynamic balance of pro- and anti-inflammatory signals controls disease and limits pathology. Immunol. Rev. 2018, 285, 147–167. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Sah, S.K.; Yang, H.S.; Lee, J.H.; Shin, J.; Kim, T.Y. Rhododendrin inhibits toll-like receptor-7-mediated psoriasis-like skin inflammation in mice. Exp. Mol. Med. 2017, 49, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Dong, L.; Jiang, J.; Zhao, J.; Zhao, G.; Dang, X.; Lu, X.; Jia, M. Chlorogenic acid reduces liver inflammation and fibrosis through inhibition of toll-like receptor 4 signaling pathway. Toxicology 2013, 303, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Nam, Y.J.; Lee, C.S. Taxifolin reduces the cholesterol oxidation product-induced neuronal apoptosis by suppressing the Akt and NF-κB activation-mediated cell death. Brain Res. Bull. 2017, 134, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.P.; Mahajan, S.; Reynolds, J.L.; Aalinkeel, R.; Nair, H.; Schwartz, S.A.; Kandaswami, C. The flavonoid quercetin inhibits proinflammatory cytokine (tumor necrosis factor alpha) gene expression in normal peripheral blood mononuclear cells via modulation of the NF-κβ system. Clin. Vaccine Immunol. 2006, 13, 319–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicentini, F.T.M.C.; He, T.; Shao, Y.; Fonseca, M.J.V.; Verri, W.A.; Fisher, G.J.; Xu, Y. Quercetin inhibits UV irradiation-induced inflammatory cytokine production in primary human keratinocytes by suppressing NF-κB pathway. J. Dermatol. Sci. 2011, 61, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Murase, T.; Kume, N.; Hase, T.; Shibuya, Y.; Nishizawa, Y.; Tokimitsu, I.; Kita, T. Gallates Inhibit Cytokine-Induced Nuclear Translocation of NF-κB and Expression of Leukocyte Adhesion Molecules in Vascular Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1412–1420. [Google Scholar] [CrossRef] [Green Version]
- Bang, B.W.; Park, D.; Kwon, K.S.; Lee, D.H.; Jang, M.-J.; Park, S.K.; Kim, J.-Y. BST-104, a Water Extract of Lonicera japonica, Has a Gastroprotective Effect via Antioxidant and Anti-Inflammatory Activities. J. Med. Food 2019, 22, 140–151. [Google Scholar] [CrossRef]
- Khan, A.Q.; Khan, R.; Qamar, W.; Lateef, A.; Ali, F.; Tahir, M.; Tahir, M.; Muneeb-U-Rehman, S.S. Caffeic acid attenuates 12-O-tetradecanoyl-phorbol-13-acetate (TPA)-induced NF-κB and COX-2 expression in mouse skin: Abrogation of oxidative stress, inflammatory responses and proinflammatory cytokine production. Food Chem. Toxicol. 2012, 50, 175–183. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Lo, C.P.; Chen, Y.P.; Wang, S.Y.; Yang, N.S.; Kuo, Y.H.; Shyur, L.F. Ethyl caffeate suppresses NF-κB activation and its downstream inflammatory mediators, iNOS, COX-2, and PGE 2 in vitro or in mouse skin. Br. J. Pharmacol. 2005, 146, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Amirova, K.M.; Dimitrova, P.; Marchev, A.S.; Aneva, I.Y.; Georgiev, M.I. Clinopodium vulgare L. (wild basil) extract and its active constituents modulate cyclooxygenase-2 expression in neutrophils. Food Chem. Toxicol. 2019, 124, 1–9. [Google Scholar] [CrossRef]
- Meng, Z.Q.; Tang, Z.H.; Yan, Y.X.; Guo, C.R.; Cao, L.; Ding, G.; Huang, W.Z.; Wang, Z.Z.; Wang, K.D.G.; Xiao, W.; et al. Study on the anti-gout activity of chlorogenic acid: Improvement on hyperuricemia and gouty inflammation. Am. J. Chin. Med. 2014, 42, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhou, J.; Wang, L.; Li, B.; Guo, J.; Guan, X.; Han, Q.; Zhang, H. Caffeic Acid Reduces Cutaneous Tumor Necrosis Factor Alpha (TNF-α), IL-6 and IL-1β Levels and Ameliorates Skin Edema in Acute and Chronic Model of Cutaneous Inflammation in Mice. Biol. Pharm. Bull. 2014, 37, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerriero, E.; Sorice, A.; Capone, F.; Costantini, S.; Palladino, P.; D’ischia, M.; Castello, G. Effects of Lipoic Acid, Caffeic Acid and a Synthesized Lipoyl-Caffeic Conjugate on Human Hepatoma Cell Lines. Molecules 2011, 16, 6365–6377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.J.; Kim, Y.W.; Park, Y.; Lee, H.J.; Kim, K.W. Anti-inflammatory effects of chlorogenic acid in lipopolysaccharide- stimulated RAW 264.7 cells. Inflamm. Res. 2014, 63, 81–90. [Google Scholar] [CrossRef]
- Kurokawa, M.; Watanabe, W.; Shimizu, T.; Sawamura, R.; Shiraki, K. Modulation of cytokine production by 7-hydroxycoumarin in vitro and its efficacy against influenza infection in mice. Antivir. Res. 2010, 85, 373–380. [Google Scholar] [CrossRef]
- Kabel, A.M.; Al-Shehri, A.H.; Al-Talhi, R.A.; Abd Elmaaboud, M.A. The promising effect of linagliptin and/or indole-3-carbinol on experimentally-induced polycystic ovarian syndrome. Chem. Biol. Interact. 2017, 273, 190–199. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Liu, C.; Reiter, R.J. COVID-19: Melatonin as a potential adjuvant treatment. Life Sci. 2020, 250, 117583. [Google Scholar] [CrossRef]
- Alamili, M.; Bendtzen, K.; Lykkesfeldt, J.; Rosenberg, J.; Gögenur, I. Melatonin suppresses markers of inflammation and oxidative damage in a human daytime endotoxemia model. J. Crit. Care 2014, 29, 184.e9–184.e13. [Google Scholar] [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Y.H.; Lu, Q. Pro-oncogenic and anti-oncogenic pathways: Opportunities and challenges of cancer therapy. Future Oncol. 2010, 6, 587–603. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Sato, S.; Nagase, S.; Shimosato, K.; Saito, T. Polyamine levels in various tissues of rats treated with 3-hydroxy-4-methoxycinnamic acid and 3,4-dimethoxycinnamic acid. Anticancer Drugs 1996, 7, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.; Husheem, M.; Härkönen, P.; Pihlaja, K. Inhibition of cancer cell growth by crude extract and the phenolics of Terminalia chebula retz. fruit. J. Ethnopharmacol. 2002, 81, 327–336. [Google Scholar] [CrossRef]
- Kim, W.H.; Song, H.O.; Choi, H.J.; Bang, H.I.; Choi, D.Y.; Park, H. Ethyl gallate induces apoptosis of HL-60 cells by promoting the expression of caspases-8, -9, -3, apoptosis-inducing factor and endonuclease G. Int. J. Mol. Sci. 2012, 13, 11912–11922. [Google Scholar] [CrossRef]
- Manigandan, K.; Manimaran, D.; Jayaraj, R.L.; Elangovan, N.; Dhivya, V.; Kaphle, A. Taxifolin curbs NF-κB-mediated Wnt/β-catenin signaling via up-regulating Nrf2 pathway in experimental colon carcinogenesis. Biochimie 2015, 119, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Razak, S.; Afsar, T.; Ullah, A.; Almajwal, A.; Alkholief, M.; Alshamsan, A.; Jahan, S. Taxifolin, a natural flavonoid interacts with cell cycle regulators causes cell cycle arrest and causes tumor regression by activating Wnt/ β -catenin signaling pathway 06 Biological Sciences 0601 Biochemistry and Cell Biology 11 Medical and Health Sciences 1112 Oncology and Carcinogenesis. BMC Cancer 2018, 18. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Liu, Z.; Wang, M.; Chen, D.; Zhou, L.; Guo, L. Taxifolin inhibits the development of scar cell carcinoma by inducing apoptosis, cell cycle arrest, and suppression of PI3K/ AKT/mTOR pathway. J. BUON 2019, 24, 853–858. [Google Scholar]
- Zhang, Z.R.; Al Zaharna, M.; Wong, M.M.K.; Chiu, S.K.; Cheung, H.Y. Taxifolin Enhances Andrographolide-Induced Mitotic Arrest and Apoptosis in Human Prostate Cancer Cells via Spindle Assembly Checkpoint Activation. PLoS ONE 2013, 8, e54577. [Google Scholar] [CrossRef] [Green Version]
- Kabala-Dzik, A.; Rzepecka-Stojko, A.; Kubina, R.; Jastrzȩbska-Stojko, Ż.; Stojko, R.; Wojtyczka, R.D.; Stojko, J. Migration rate inhibition of breast cancer cells treated by caffeic acid and caffeic acid phenethyl ester: An in vitro comparison study. Nutrients 2017, 9, 1144. [Google Scholar] [CrossRef]
- Viveros-Valdez, E.; Rivas-Morales, C.; Oranday-Cárdenas, A.; Castro-Garza, J.; Carranza-Rosales, P. Antiproliferative effect from the Mexican Poleo (Hedeoma drummondii). J. Med. Food 2010, 13, 740–742. [Google Scholar] [CrossRef]
- Tyszka-Czochara, M.; Bukowska-Strakova, K.; Majka, M. Metformin and caffeic acid regulate metabolic reprogramming in human cervical carcinoma SiHa/HTB-35 cells and augment anticancer activity of Cisplatin via cell cycle regulation. Food Chem. Toxicol. 2017, 106, 260–272. [Google Scholar] [CrossRef]
- Nasr Bouzaiene, N.; Kilani Jaziri, S.; Kovacic, H.; Chekir-Ghedira, L.; Ghedira, K.; Luis, J. The effects of caffeic, coumaric and ferulic acids on proliferation, superoxide production, adhesion and migration of human tumor cells in vitro. Eur. J. Pharmacol. 2015, 766, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, S.K. Growth inhibition by caffeic acid, one of the phenolic constituents of honey, in HCT 15 colon cancer cells. Sci. World J. 2012, 2012, 372345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelinson, L.P.; Assmann, C.E.; Palma, T.V.; da Cruz, I.B.M.; Pillat, M.M.; Mânica, A.; Stefanello, N.; Weis, G.C.C.; de Oliveira Alves, A.; de Andrade, C.M.; et al. Antiproliferative and apoptotic effects of caffeic acid on SK-Mel-28 human melanoma cancer cells. Mol. Biol. Rep. 2019, 46, 2085–2092. [Google Scholar] [CrossRef] [PubMed]
- Dziedzic, A.; Kubina, R.; Kabała-Dzik, A.; Wojtyczka, R.D.; Morawiec, T.; Bułdak, R.J. Caffeic acid reduces the viability and migration rate of oral carcinoma cells (SCC-25) exposed to low concentrations of ethanol. Int. J. Mol. Sci. 2014, 15, 18725–18741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.T.; Chen, I.L.; Chou, C.T.; Liang, W.Z.; Kuo, D.H.; Shieh, P.; Jan, C.R. Effect of caffeic acid on Ca2+ homeostasis and apoptosis in SCM1 human gastric cancer cells. Arch. Toxicol. 2013, 87, 2141–2150. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.J.; Lee, K.W.; Shin, B.J.; Jung, S.K.; Hwang, M.K.; Bode, A.M.; Heo, Y.S.; Lee, H.J.; Dong, Z. Caffeic acid, a phenolic phytochemical in coffee, directly inhibits Fyn kinase activity and UVB-induced COX-2 expression. Carcinogenesis 2009, 30, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.K.; Hwang, J.T.; Kwon, D.Y.; Surh, Y.J.; Park, O.J. Induction of apoptosis by quercetin is mediated through AMPKα1/ASK1/p38 pathway. Cancer Lett. 2010, 292, 228–236. [Google Scholar] [CrossRef]
- Ranelletti, F.O.; Maggiano, N.; Serra, F.G.; Ricci, R.; Larocca, L.M.; Lanza, P.; Scambia, G.; Fattorossi, A.; Capelli, A.; Piantelli, M. Quercetin inhibits p21-ras expression in human colon cancer cell lines and in primary colorectal tumors. Int. J. Cancer 2000, 85, 438–445. [Google Scholar] [CrossRef]
- Yang, Y.M.; Son, Y.O.; Lee, S.A.; Jeon, Y.M.; Lee, J.C. Quercetin inhibits α-MSH-stimulated melanogenesis in B16F10 melanoma cells. Phyther. Res. 2011, 25, 1166–1173. [Google Scholar] [CrossRef]
- Siegelin, M.D.; Reuss, D.E.; Habel, A.; Rami, A.; Von Deimling, A. Quercetin promotes degradation of survivin and thereby enhances death-receptor- mediated apoptosis in glioma cells. Neuro Oncol. 2009, 11, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Papp, L.V.; Fang, J.; Rodriguez-Nieto, S.; Zhivotovsky, B.; Holmgren, A. Inhibition of mammalian thioredoxin reductase by some flavonoids: Implications for myricetin and quercetin anticancer activity. Cancer Res. 2006, 66, 4410–4418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Zhao, X.H.; Wang, Z.J. Flavones and flavonols exert cytotoxic effects on a human oesophageal adenocarcinoma cell line (OE33) by causing G2/M arrest and inducing apoptosis. Food Chem. Toxicol. 2008, 46, 2042–2053. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Lee, J.O.; Kim, J.H.; Lee, S.K.; You, G.Y.; Park, S.H.; Park, J.M.; Kim, E.K.; Suh, P.G.; An, J.K.; et al. Quercetin suppresses HeLa cell viability via AMPK-induced HSP70 and EGFR down-regulation. J. Cell. Physiol. 2010, 223, 408–414. [Google Scholar] [CrossRef]
- Haghiac, M.; Walle, T. Quercetin induces necrosis and apoptosis in SCC-9 oral cancer cells. Nutr. Cancer 2005, 53, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Mu, C.; Jia, P.; Yan, Z.; Liu, X.; Li, X.; Liu, H. Quercetin induces cell cycle G1 arrest through elevating Cdk inhibitors p21 and p27 in human hepatoma cell line (HepG2). Methods Find. Exp. Clin. Pharmacol. 2007, 29, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Monasterio, A.; Urdaci, M.C.; Pinchuk, I.V.; López-Moratalla, N.; Martínez-Irujo, J.J. Flavonoids induce apoptosis in human leukemia U937 cells through caspase- and caspase-calpain-dependent pathways. Nutr. Cancer 2004, 50, 90–100. [Google Scholar] [CrossRef]
- Liang, W.; Li, X.; Li, C.; Liao, L.; Gao, B.; Gan, H.; Yang, Z.; Liao, L.; Chen, X. Quercetin-mediated apoptosis via activation of the mitochondrial-dependent pathway in MG-63 osteosarcoma cells. Mol. Med. Rep. 2011, 4, 1017–1023. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.T.; Huang, Y.T.; Hwang, J.J.; Lee, P.P.H.; Ke, F.C.; Nair, M.P.; Kanadaswami, C.; Lee, M.T. Blockade of the epidermal growth factor receptor tyrosine kinase activity by quercetin and luteolin leads to growth inhibition and apoptosis of pancreatic tumor cells. Anticancer Res. 2002, 22, 1615–1627. [Google Scholar]
- Deka, S.; Gorai, S.; Manna, D.; Trivedi, V. Evidence of PKC Binding and Translocation to Explain the Anticancer Mechanism of Chlorogenic Acid in Breast Cancer Cells. Curr. Mol. Med. 2017, 17, 79–89. [Google Scholar] [CrossRef]
- Barahuie, F.; Saifullah, B.; Dorniani, D.; Fakurazi, S.; Karthivashan, G.; Hussein, M.Z.; Elfghi, F.M. Graphene oxide as a nanocarrier for controlled release and targeted delivery of an anticancer active agent, chlorogenic acid. Mater. Sci. Eng. C 2017, 74, 177–185. [Google Scholar] [CrossRef]
- Hou, N.; Liu, N.; Han, J.; Yan, Y.; Li, J. Chlorogenic acid induces reactive oxygen species generation and inhibits the viability of human colon cancer cells. Anticancer Drugs 2017, 28, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Satoh, K.; Watanabe, S.; Kusama, K.; Sakagami, H. Inhibition of chlorogenic acid-induced cytotoxicity by COCl2. Anticancer Res. 2001, 21, 3349–3353. [Google Scholar] [PubMed]
- m/z cloud. Available online: https://www.mzcloud.org/ (accessed on 5 May 2020).
- Pence, H.E.; Williams, A. Chemspider: An online chemical information resource. J. Chem. Educ. 2010, 87, 1123–1124. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids Database: Structures, chemical fingerprints and distribution among organisms. Database 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FooDB. Available online: https://foodb.ca/ (accessed on 5 May 2020).
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Lee, S.O.; Kim, K.R.; Lee, H.J. Sphingosine kinase-1 involves the inhibitory action of HIF-1α by chlorogenic acid in hypoxic DU145 cells. Int. J. Mol. Sci. 2017, 18, 325. [Google Scholar] [CrossRef] [Green Version]
- The LIPID MAPS® Lipidomics Gateway. Available online: https://www.lipidmaps.org (accessed on 5 May 2020).
- Rothwell, J.A.; Perez-Jimenez, J.; Neveu, V.; Medina-Remón, A.; M’Hiri, N.; García-Lobato, P.; Manach, C.; Knox, C.; Eisner, R.; Wishart, D.S.; et al. Phenol-Explorer 3.0: A major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content. Database 2013, 2013. [Google Scholar] [CrossRef]
- Plant Metabolics Network (PMN). Available online: www.plantcyc.org (accessed on 5 May 2020).
- Ramirez-Gaona, M.; Marcu, A.; Pon, A.; Guo, A.C.; Sajed, T.; Wishart, N.A.; Karu, N.; Djoumbou Feunang, Y.; Arndt, D.; Wishart, D.S. YMDB 2.0: A significantly expanded version of the yeast metabolome database. Nucleic Acids Res. 2016, 45, D440–D445. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem substance and compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [Green Version]
- PubMed. Available online: www.pubmed.ncbi.nlm.nih.gov (accessed on 5 May 2020).
- Kim, S.; Thiessen, P.A.; Cheng, T.; Yu, B.; Bolton, E.E. An update on PUG-REST: RESTful interface for programmatic access to PubChem. Nucleic Acids Res. 2018, 46, W563–W570. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiao, J.; Suzek, T.O.; Zhang, J.; Wang, J.; Zhou, Z.; Han, L.; Karapetyan, K.; Dracheva, S.; Shoemaker, B.A.; et al. PubChem’s BioAssay Database. Nucleic Acids Res. 2012, 40, D400–D412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Thiessen, P.A.; Cheng, T.; Zhang, J.; Gindulyte, A.; Bolton, E.E. PUG-View: Programmatic access to chemical annotations integrated in PubChem. J. Cheminform. 2019, 11, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | PubChem ID | Role, Biological Activity |
|---|---|---|
| Achalensolide | 21634938 | NF-κB inhibitory activity in Jurkat T cells |
| DL-alanyl-DL-phenylalanine | 2080 | Cytotoxic effect against aggressive human metastatic breast adenocarcinoma MDA-MB-231 and MCF-7 cells |
| Asperlin | 35319 | Induction of apoptosis via ROS production in human cervical carcinoma HeLa cells |
| Inhibition of iNOS, suppression of COX-2 expression, reduction in COX-derived PGE2 | ||
| Bestatin | 439299 | Antiviral activity against coronaviruses |
| Caffeic acid | 689043 | Anticancer activity against human cell lines originating from breast cancer (MCF-7, MDA-MB-231) cervical cancer (HeLa), metastatic cervical cancer (SiHa), hepatocellular carcinoma (HepG2, Huh7), lung cancer (A-549), colon carcinoma (HT29-D4, HCT 15), melanoma (SK-Mel-28), oral squamous cell carcinoma (SCC-25), gastric cancer (SCM1), suppression of UVB-induced skin carcinogenesis |
| Reduction in lipid peroxidation and of TNF-α, IL-6, IL-1β, IFN-γ, NF-κB/p65 and TGF-β levels | ||
| Dimethylcaffeic acid | 717531 | Antiviral activity against HIV-1 |
| Beneficial effect in prostate-, thymus- and stomach-, lung- and brain cancer | ||
| (E)-Ethyl caffeate | 5317238 | Antiviral activity against HBV, HIV-1 |
| Anticancer activity against cancer cell lines: human hepatocellular carcinoma BEL-7404 and HepG2, SK-OV-3 human ovarian cancer, human breast MCF-7 adenocarcinoma, human lung A549 adenocarcinoma and human gastric cancer BCG823 | ||
| Suppression of NF-κB activation and its downstream inflammatory mediators, iNOS, COX-2, and PGE2 | ||
| Chlorogenic acid | 1794427 | Antiviral activity against hepatitis C, hepatitis B, Ebola virus, HIV-1, adenoviruses, H1N1/H3N2 influenza strains |
| Anticancer activity against human cell lines originating from breast cancer (MCF-7, MDA-MB-231), cervical cancer (HeLa), hepatocellular carcinoma (HepG2), lung cancer (A-549), colon carcinoma (CT-26, Caco-2, HT29-D4, HCT 116), melanoma (SK-Mel-28), oral squamous cell carcinoma (HSC-2), salivary gland cancer (HSG), pancreatic cancer (PANC-1), leukemia (U937, HL-60, K562), prostate cancer (DU145) | ||
| Anti-inflammatory activity, inhibition of NO and proinflammatory cytokine production | ||
| Coumarin | 323 | Coumarin derivatives exert anti-coagulant, anti-tumor, anti-viral, anti-inflammatory and antioxidant effects, as well as anti-microbial and enzyme inhibition properties |
| Anticancer activity against human cell lines originating from gastric carcinoma, colon-carcinoma cell line (Caco-2), hepatoma-derived cell line (HepG2), lymphoblastic cell line (CCRF CEM), lung adenocarcinoma (A427, Calu-1, SK-MES-1, SK-LU-1), renal carcinoma (786-O, A-498), malignant prostatic cancer (DU145, LNCaP), beneficial effects in renal cell carcinoma and malignant melanoma | ||
| (E)-p-coumaric acid | 637542 | Antiviral activity against oseltamivir- and peramivir-sensitive and oseltamivir- and peramivir-resistant influenza viruses and hepatitis C |
| Anticancer activity in mammalian, colon and hepatic cancer and neuroblastoma cell lines | ||
| Inhibition of the STAT1 activation, decrease in TNF-α expression, ROS scavenger function | ||
| Cyclo(phenylalanyl-prolyl) | 99895 | Growth inhibition and apoptosis induction in HT-29 colon cancer cells |
| Dehydrocostus lactone | 73174 | Inhibition of Norovirus infection |
| Anticancer activity against human cell lines originating from ovarian cancer (SK-OV-3), breast cancer (MCF-7, MDA-MB-231), cervical cancer (HeLa), hepatocellular carcinoma (HepG2), lung adenocarcinoma (A-549, NCI-H520, NCI-H460), prostate cancer (DU145), sarcoma (liposarcoma–SW-872, synovial sarcoma – SW-982, TE-671 – rhabdomyosarcoma), neuroblastoma (IMR-32, NB-39, SK-N-SH, LA-N-1) | ||
| Reduced production of chemokines induced by TNF-α and IFN-γ | ||
| Dodecanedioic acid | 12736 | Cytotoxic effect on B16 melanoma cells |
| Eicosapentaenoic acid | 446284 | Growth inhibition effect on colon cancer cell lines (HT-29, Caco-2, DLD-1), antiproliferative effects on hepatoma (HepG2), leukemia (HL-60) cell lines, inhibition of macrophage-induced gastric cancer cell migration |
| Attenuation of pro-inflammatory properties of VLDL via decrease in lipoprotein-lipase activity to hydrolyze VLDL | ||
| Ethyl gallate | 13250 | Anti-herpes simplex virus type 1 activity |
| Anticancer activity against human cell lines originating from leukemia (HL-60), prostate cancer (PC-3), human (MCF-7) and mouse (S115) breast cancer, osteosarcoma (HOS-1), ovarian cancer (OVCAR-3), renal cancer (A-498), lung cancer (NCI-H460), colon cancer (KM20L2) and melanoma (SK-MEL-5) | ||
| Inhibition of LPS induced cell adhesion molecules expression, attenuation of acute lung injury | ||
| Fertaric acid | 22298372 | Hepatoprotective effects |
| 16-Heptadecyne-1,2,4-triol | 3015189 | Cytotoxic activity in human lung carcinoma (A-549), mammary adenocarcinoma (MCF-7), colon adenocarcinoma (HT-29), kidney carcinoma (A-498), pancreatic carcinoma (PaCa-2), prostate adenocarcinoma (PC-3) cell lines |
| L-Histidinol | 165271 | Pro-apoptotic activity in CCRF-CEM human leukemia cell line, inhibition of B16 melanoma cell proliferation |
| Indole-3-carbinol | 3712 | Promotion of apoptosis of Epstein–Barr virus (EBV)-positive but not of EBV-negative Burkitt’s lymphoma cell lines |
| Anticancer activity against human cell lines originating from various cancer types such as prostate cancer (LnCaP, PC-3), breast cancer (MCF7, MDA-MB-468, MDA-MB-231, HBL100), colon cancer (HT-29, HCT-116), lung cancer (A-549), cervical cancer (CaSki, SiHa, C33-A), melanoma (SK-MEL-2, SK-MEL-5), ovarian cancer (SK-OV-3), oral squamous cell carcinoma (SCC2095, SCC9, SCC15), hepatocellular carcinoma (HepG2, Huh-7, SNU449), pancreatic cancer (BxPC-3, Mia Paca-2, PL-45, AsPC-1, PANC-1), leukemia (U937, HL-60, K562, BCP-ALL NALM-6), osteosarcoma (U2OS) etc. | ||
| Reversal of liver fibrosis, reduction in hepatocyte degeneration, necrosis, promotion of hepatic stellate cell apoptosis, anti-inflammatory effects by inhibiting the productions of NO, TNF-alpha, and IL-10 | ||
| Kynurenic acid | 3845 | Modulation of IL-23 and IL-17 expression in dendritic cells and Th17 cells |
| Linamarin | 11128 | Cytotoxic effects on MCF-7, HT-29 and HL60 cell lines |
| Melatonin | 896 | Indirect support against Ebola virus infection, Potential adjuvant treatment in COVID-19, and other viral infections |
| Upregulation of Fas/Fas ligand in Ewing’s sarcoma cells, cell cycle arrest and apoptosis in hepatocarcinoma HepG2 cell line, induction of pro-apoptotic signaling pathway in human pancreatic carcinoma cells, anticancer activity in breast cancer (MCF-7) cells, inhibition of the proliferation and invasion of glioma cells, lung adenocarcinoma (A-549) cells, inhibition of estrogen receptor transactivation in breast cancer cells, negative mitogenic hormonal regulator of human prostate epithelial cells | ||
| Decreases serum and tissue inflammatory cytokines levels, tissue lipid peroxidation and neutrophil infiltration | ||
| 9S,13R-12-Oxophytodienoic acid | 14037063 | Induces growth arrest in MDA-MB-231 and T47D breast cancer cells followed by progressive reduction in cyclin D1 expression |
| Suppression of NF-κB, inhibition of p38, and activation of SOCS-1 signaling | ||
| Perlolyrine | 160179 | Antiproliferative activity against human stomach cancer cell lines |
| Phytosphingosine | 122121 | Induction of apoptotic cell death via caspase 8 activation and Bax translocation in human cancer cells |
| Quercetin | 5280343 | Antiviral activity against HIV-1, hepatitis B and C viruses, adenoviruses, herpes simplex viruses, noroviruses, H1N1, H5N1 etc. |
| Anticancer activity against human cell lines originating from various cancer types such as breast cancer (MCF7), colon cancer (HT-29, HCT-116, Caco-2, DLD-1), lung cancer (A-549), esophageal squamous cell carcinoma cell line (KYSE-510, OE33), cervical cancer (HeLa), oral squamous cell carcinoma (SCC9), hepatocellular carcinoma (HepG2), leukemia (U937, HL-60), osteosarcoma (MG-63), pancreatic cancer (PC3, EPP85-181P, EPP85-181RDB), melanoma (B16F10), glioma (U87, U139MG), inhibition of angiogenesis in tamoxifen-resistant breast cancer cells | ||
| Inhibition of inflammatory cytokine production, Inhibition of histamine release, Reduction in neutrophil recruitment, Ameliorating endothelial insulin resistance through inhibition of reactive oxygen species-associated inflammation. | ||
| (-)-Rhododendrin | 442538 | Inhibition of toll-like receptor-7-mediated inflammation |
| Sphinganine | 91486 | Anticancer activity against human cell lines originating from leukemia (HL-60), prostate cancer (PC-3, LnCaP), breast cancer (MDA-MB-231), colon cancer (HT-29, HCT-116) and melanoma (939, 294, C8161, A2058), oral squamous cell carcinoma (SAS, Ca9-22, HSC-3) |
| Taxifolin | 439533 | Antiviral activity against HIV-1, coxsackieviruses B4, hepatitis A |
| Anticancer effect due to Nrf2, inflammatory and Wnt/β-catenin cascade modulation, inhibition of breast cancer MDA-MB-231 and 4T1 cell proliferation, cell cycle arrest in human colorectal cancer HCT116 and HT29 cells, activation of apoptosis in prostate carcinoma DU145 cells | ||
| Antioxidant and anti-inflammatory effects by inhibition of NO and PGE2 production, ICAM-1, COX-2 and PLA2 expression | ||
| Tetrahydroharman-3-carboxylic acid | 73530 | Inhibition of nitric oxide and prostaglandin E2 production |
| Umbelliferone | 5281426 | Antiviral activity against HIV-1 |
| Cytostatic activity in human malignant cell lines A549, ACHN, Caki-2, Dakiki, HS-Sultan, H727, HCT-15, HL-60, K562, LNCaP, PC-3, Du 145 COLO-232, MCF-7 and RP-1788; stimulation of apoptosis in HL-60 cells, Growth inhibition of human bladder carcinoma E-J cell line, lung adenocarcinoma A-427 cells, proliferation inhibition of gastric carcinoma, colon-carcinoma (Caco-2), a hepatoma-derived (HepG2), and a lymphoblastic cell line (CCRF CEM), chemoprotective effect in early-stage (Ln- Cap) and late-stage (PC3) prostate cancer cells | ||
| Anti-inflammatory and antipyretic effects, reduction of IL-4, IL-5 and IL-13, suppression of Th1 cytokine production during influenza virus infection | ||
| Zedoarondiol | 14632997 | Inhibition of iNOS, COX-2 activity and of the production of NO, PGE2, TNF-alpha, IL-6, and IL-1beta |
| Name | Log2 Fold Change | Adjusted p-Value | Biological Roles |
|---|---|---|---|
| DL-isoleucyl-DL-isoleucyl-DL-histidine | −9.04 | 2.43 × 10−11 | No information |
| DL-alanyl-DL-isoleucyl-DL-isoleucyl-DL-threonine | −7.62 | 2.18 × 10−14 | No information |
| DL-leucyl-DL-leucyl-DL-leucine | −7.60 | 1.43 × 10−11 | No information |
| DL-valyl-DL-valyl-DL-valine | −7.58 | 1.35 × 10−3 | No information |
| L-phenylalanyl-L-leucine | −7.16 | 8.81 × 10−4 | Membrane metalloendopeptidase inhibitor in mouse model [20] |
| Plant metabolite | |||
| DL-tyrosyl-DL-prolyl-DL-isoleucine | −6.46 | 5.77 × 10−7 | No information |
| (2S)-2-[(2R)-7-(2-Methoxyethoxy)-5,8-dimethyl-1,2,3,4-tetrahydro-2-naphthalenyl]-1-(1-piperidinyl)-1-propanone | −6.42 | 1.76 × 10−3 | No information |
| DL-isoleucyl-DL-alpha-glutamyl-DL-lysine | −6.18 | 9.00 × 10−7 | No information |
| 1-(4-Methoxyphenyl)-N-[3-(4-morpholinyl) propyl]-5-oxo-3-pyrrolidine carboxamide | −5.92 | 7.99 × 10−7 | Phosphoethanolamine/phosphocholine phosphatase 1 inhibitor [21] |
| N-Isobutyrylglycylglycine | −5.79 | 1.04 × 10−6 | No information |
| Phenethylamine | 5.29 | 5.97 × 10−10 | 5-hydroxytryptamine receptor agonist in rat [22,23] |
| CYP450 inhibitor [24] | |||
| Trace amine associated receptor agonist [25] | |||
| Perillartine | 5.32 | 7.90 × 10−5 | No information |
| DL-glutaminyl-DL-threonyl-DL-lysine | 5.37 | 4.92 × 10−10 | No information |
| N-Boc-Tyramine | 5.40 | 6.91 × 10−9 | No information |
| 4-(2,3-Dihydro-1,4-benzodioxin-6-yl)-4-oxobutanoic acid | 5.45 | 2.92 × 10−3 | Neuropeptide Y receptor agonist [26,27,28] |
| Thyroid stimulating hormone receptor agonist [29] | |||
| 1-Hydroxyhexane-1,2,6-tricarboxylate | 5.58 | 4.60 × 10−7 | No information |
| 2-Hydroxy-1-(6-hydroxy-2-isopropenyl-2,3-dihydro-1-benzofuran-5-yl)ethanone | 5.75 | 7.35 × 10−11 | No information |
| Rosin | 6.49 | 2.23 × 10−10 | No information |
| (2E)-3-(3,4-dimethoxypheny)prop-2-enoic acid | 7.11 | 1.77 × 10−3 | Inhibitors of HSD17B4|hydroxysteroid (17-beta) dehydrogenase 4 [30] |
| Antihemorrhagic activity in ddY mouse [31] | |||
| N-[3-(4,11-Dimethyl-2-oxo-6,7,8,9-tetrahydro-2H-[1]benzofuro[3,2-g]chromen-3-yl)propanoyl]glycine | 8.42 | 1.57 × 10−15 | No information |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalló, G.; Kunkli, B.; Győri, Z.; Szilvássy, Z.; Csősz, É.; Tőzsér, J. Compounds with Antiviral, Anti-Inflammatory and Anticancer Activity Identified in Wine from Hungary’s Tokaj Region via High Resolution Mass Spectrometry and Bioinformatics Analyses. Int. J. Mol. Sci. 2020, 21, 9547. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249547
Kalló G, Kunkli B, Győri Z, Szilvássy Z, Csősz É, Tőzsér J. Compounds with Antiviral, Anti-Inflammatory and Anticancer Activity Identified in Wine from Hungary’s Tokaj Region via High Resolution Mass Spectrometry and Bioinformatics Analyses. International Journal of Molecular Sciences. 2020; 21(24):9547. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249547
Chicago/Turabian StyleKalló, Gergő, Balázs Kunkli, Zoltán Győri, Zoltán Szilvássy, Éva Csősz, and József Tőzsér. 2020. "Compounds with Antiviral, Anti-Inflammatory and Anticancer Activity Identified in Wine from Hungary’s Tokaj Region via High Resolution Mass Spectrometry and Bioinformatics Analyses" International Journal of Molecular Sciences 21, no. 24: 9547. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249547