Influence of Natural Killer Cells and Natural Killer T Cells on Periodontal Disease: A Systematic Review of the Current Literature

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. NK Cell-Mediated Mechanisms in Periodontitis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mechanism | Evaluation of the Influence | Study Design | Reference |
|---|---|---|---|
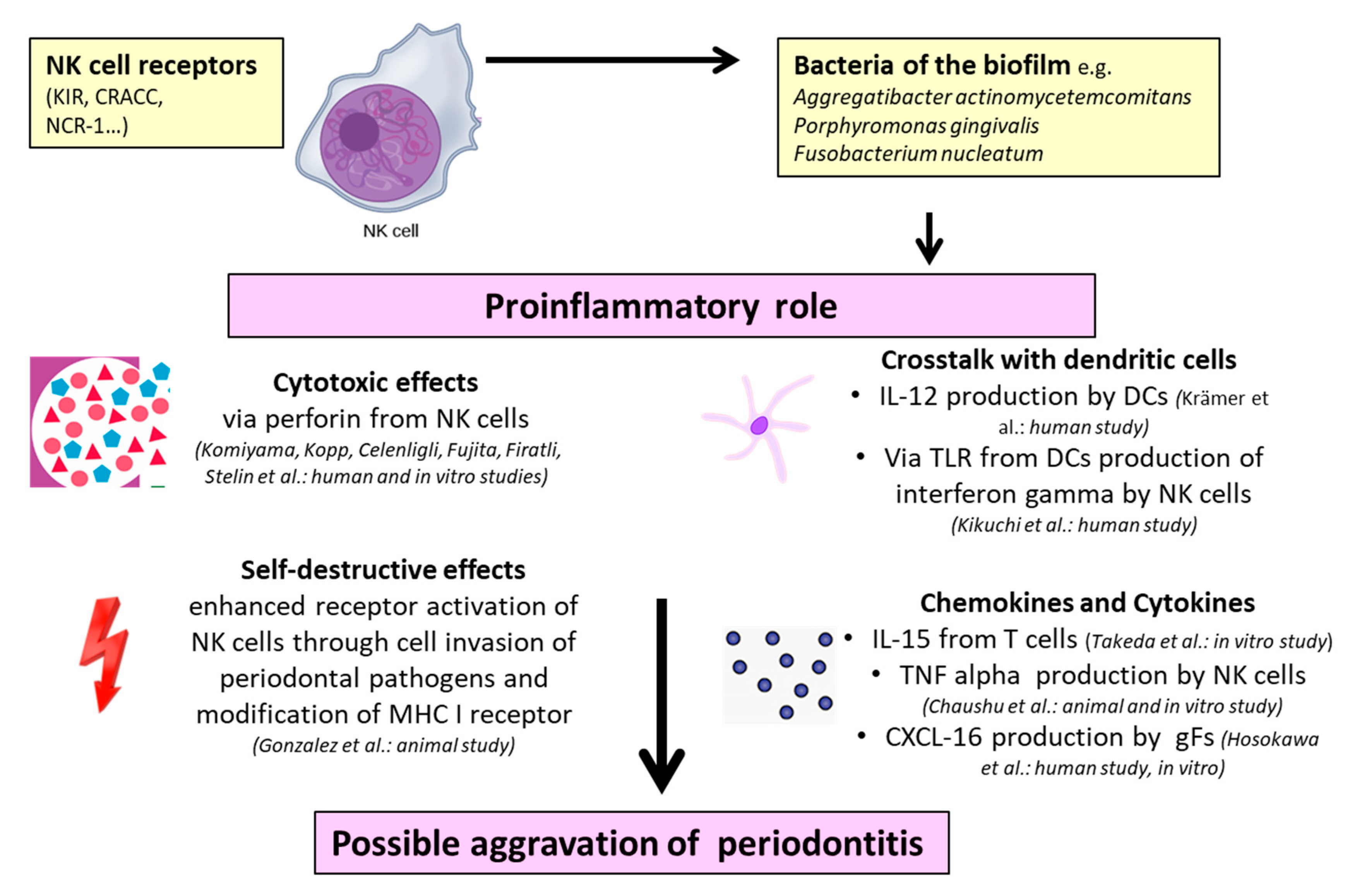
| NK cells may participate in local responses through cytotoxic and/or immunoregulatory mechanisms. | Proinflammatory through tissue damage | Human biopsies Cell cultivation Immunostaining | Komiyama et al. (1988) [54] |
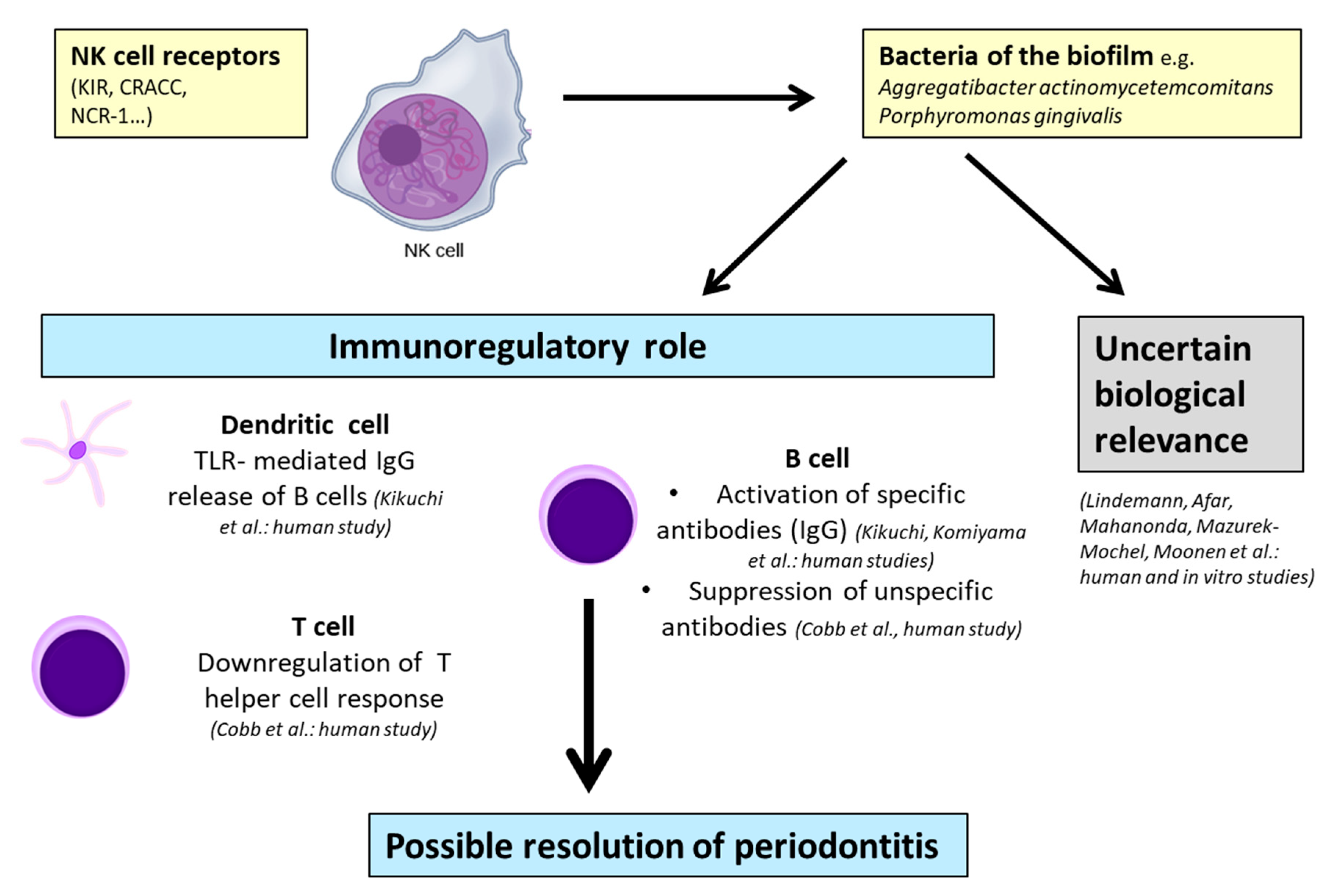
| Immunoregulatory through modulation of B-cell activity | |||
| Increased concentrations of NK cells before and decreased concentrations after periodontal therapy demonstrate that periodontitis represents an altering pathogenetic environment influencing NK cells. | Proinflammatory through upregulation and cytotoxic immunoreactive effects | Human biopsies Cell cultivation Immunostaining | Kopp et al. (1988) [55] |
| NK cells may have immunoregulatory functions in periodontal disease | Immunoregulatory through regulation of T-cell proliferation and suppression of B-cell immunoglobulin production | Human biopsies Cell cultivation Immunostaining | Cobb at al. (1989) [56] |
| Cytotoxicity was exclusively found in NK-enriched low-density large granular lymphocyte fractions after activation by LPS from A.a. without stimulating high levels of proliferation | Uncertain biological relevance | Lymphocytes from PB Human cell cultivation | Lindemann et al. (1989) [57] |
| NK cells seemed to be significantly elevated in the peripheral blood of patients with juvenile and rapidly progressive periodontitis | Proinflammatory through immuno-pathogenetic effects | Lymphocyte analysis from human PB Immunostaining | Celenligli et al. (1990) [58] |
| Exact nature of the role of NK cells needs further investigation | Uncertain biological relevance | Lymphocyte analysis from human PB Immunostaining | Afar at el. (1992) [59] |
| Leu-11b-positive cells (CD 16+ NK cells) appeared more frequently in severe forms of periodontitis | Proinflammatory through destructive reactions | Human biopsies Immunostaining | Fujita et al. (1992) [60] |
| In all of the investigated patients, cytotoxic effects of NK cells were enhanced | Proinflammatory through enhanced cytotoxic effects | Lymphocyte analysis from human PB Immunostaining | Firatli et al. (1996) [61] |
| Influence of NK cells and T cells in the diseased group is uncertain | Uncertain biological relevance | Lymphocytes from PB Cell cultivation Immunostaining | Mahanonda et al. (2004) [62] |
| A.a.–LPS–TLR4 interactions on dendritic cells initiate the pathway leading to the production of IFN-ɣ by NK cells in periodontitis; this results in high levels of IgG2 | Proinflammatory through destructive effects via IFN-ɣ production induced by IL-12 | Lymphocyte analysis from human PB Cell cultivation Immunostaining | Kikuchi et al. (2004) [63] |
| Immunoregulatory through protective effects via IgG2 production | |||
| P.g.–DC–NK interactions can result in reciprocal activation and increase of cytokine production by both DCs and NK cells. NK cells may provide IFN-ɣ needed to induce the P.g.-specific IgG2 in periodontitis | Immunoregulatory through induction of IgG2 response | Lymphocyte analysis from human PB Cell cultivation PCR | Kikuchi et al. (2005) [64] |
| Higher levels of CD57+ NK cells in tissue with periodontal disease indicate pathological progress | Proinflammatory through cytotoxic effects | Human biopsies Immunostaining | Stelin et al. (2009) [65] |
| Bone loss in the presence of NCR1 after infection with F.n. F.n. triggers the secretion of TNF-α which is dependent on NCR1 and binds directly on NCR1. | Proinflammatory through secretion of TNF-α | Induction of experimental periodontitis in mice Murine cell cultivation Immunostaining | Chaushu et al. (2012) [66] |
| Increased CRACC induction in aggressive periodontitis and in response to infections with A.a. CRACC-mediated NK cell activation could lead to an accelerated course of tissue destruction in aggressive periodontitis | Proinflammatory through CRACC induction activated by dendritic cells and subsequent IL-12 signaling | Human biopsies Transcriptomic analysis Cell cultivation | Krämer et al. (2013) [67] |
| Substantive increase in expression of genes related to NK cell interactions with antigen-presenting cells in periodontal tissues | Proinflammatory through cell invasion of periodontal pathogens and autoimmunity | Tissue samples of rhesus monkeys Gene expression microarray analysis | Gonzalez et al. (2014) [68] |
| IL-15 (bone destructive factor) activated NK cells are responsible for the induction of osteoblast apoptosis | Proinflammatory through IL-15 activation | Murine cell cultivation Immunostaining Gene expression | Takeda et al. (2014) [69] |
| Tissue damage is not associated with the presence or absence of different KIR genes | Uncertain biological relevance | Lymphocytes from human PB Gene analysis | Mazurek-Mochel et al. (2014) [70] |
| KIR presence/absence polymorphism is not a significant factor involved in the pathogenesis of periodontitis in contrast to tobacco smoking | Uncertain biological relevance | Lymphocytes from human PB Gene analysis | Mazurek-Mochol et al. (2017) [71] |
| Numbers of NK cells were increased in the presence of gFs. GFs support the retention and survival of NK cells by LFA-1 expression | Uncertain biological relevance | Cell cultivation Immunostaining Gene analysis | Moonen et al. (2018) [72] |
2.1.1. Proinflammatory Properties of NK Cells in Periodontitis
2.1.2. Immunoregulatory Properties of NK Cells in Periodontitis
2.1.3. Uncertain Biological Relevance of NK Cells in Periodontitis
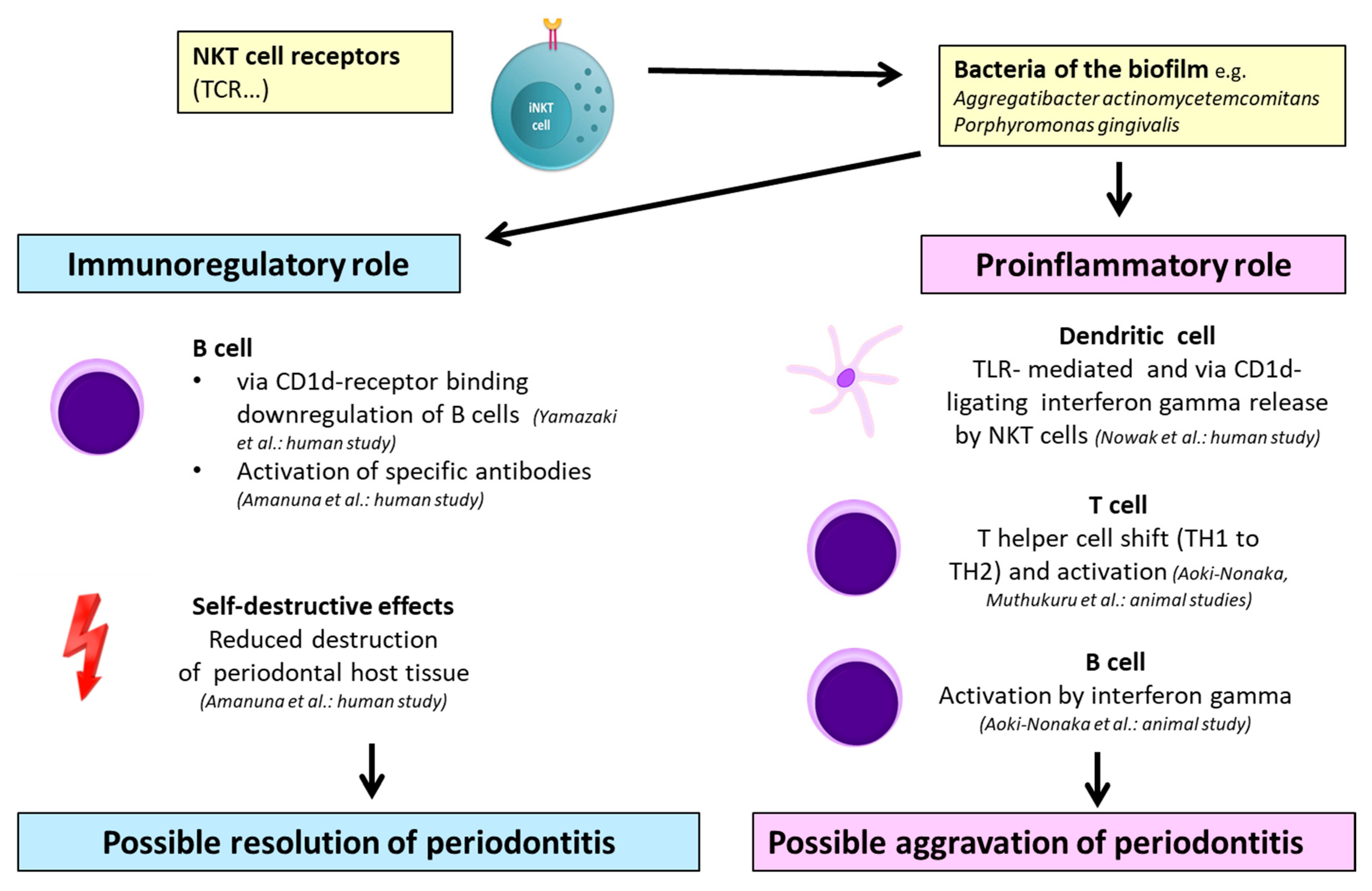
2.2. NKT Cell-Mediated Mechanisms in Periodontitis
| Mechanism | Evaluation of the Influence | Study Design | Reference |
|---|---|---|---|
| Invariant NKT cells infiltrating periodontal lesions seem to have a downregulating role of autoimmune responses | Immunoregulatory through association with CD1d(+) cells | Human biopsies Lymphocytes from human peripheral blood Gene analysis Immunostaining | Yamazaki et al. (2001) [73] |
| NKT cells can provide direct help for B-cell proliferation and antibody production against autoimmune reactions | Immunoregulatory through activation by CD1d-expressing B cells | Human biopsies Immunostaining | Amanuma et al. (2006) [74] |
| Type I NKT cell activation and IFN-ɣ secretion possibly aggravate tissue destruction in aggressive periodontitis | Proinflammatory through activation of type I NKT cells with subsequent production of IFN-ɣ triggered by A.a. | Human biopsies Transcriptomic analysis Murine cell cultivation | Nowak et al. (2013) [75] |
| Activation of NKT cells promoted a systematic inflammatory response. P.g. induced alveolar bone resorption via NKT cell activation | Proinflammatory through T-cell and B-cell activation mediated by NKT cells via IFN-ɣ | Murine periodontal infection Murine biopsy, blood, and gene analysis | Aoki-Nonaka et al. (2014) [76] |
2.2.1. Immunoregulatory Properties of NKT Cells in Periodontitis
2.2.2. Proinflammatory Properties of NKT Cells in Periodontitis
2.3. Combined NK Cell-/NKT Cell-Mediated Mechanisms in Periodontitis
| Mechanism | Evaluation of the Influence | Study Design | Reference |
|---|---|---|---|
| CXCL 16 produced by human gFs in diseased periodontal tissues controls the migration of NK and NKT cells, leading to bone resorption | Proinflammatory through IFN-ɣ and further attraction by chemokines | Human biopsies Cell cultivation Gene analysis | Hosokawa et al. (2007) [77] |
| Increased number of NK and NKT cells in tissues from patients with chronic periodontitis | Proinflammatory through downregulation of T-helper cells | Human biopsies Cell cultivation Immunostaining | Muthukuru et al. (2012) [78] |
3. Discussion
3.1. Role of NK Cells in Periodontitis
3.2. Role of NKT Cells in Periodontitis
3.3. Combined Role of NK/NKT Cells in Periodontitis
3.4. Role of Different Bacteria
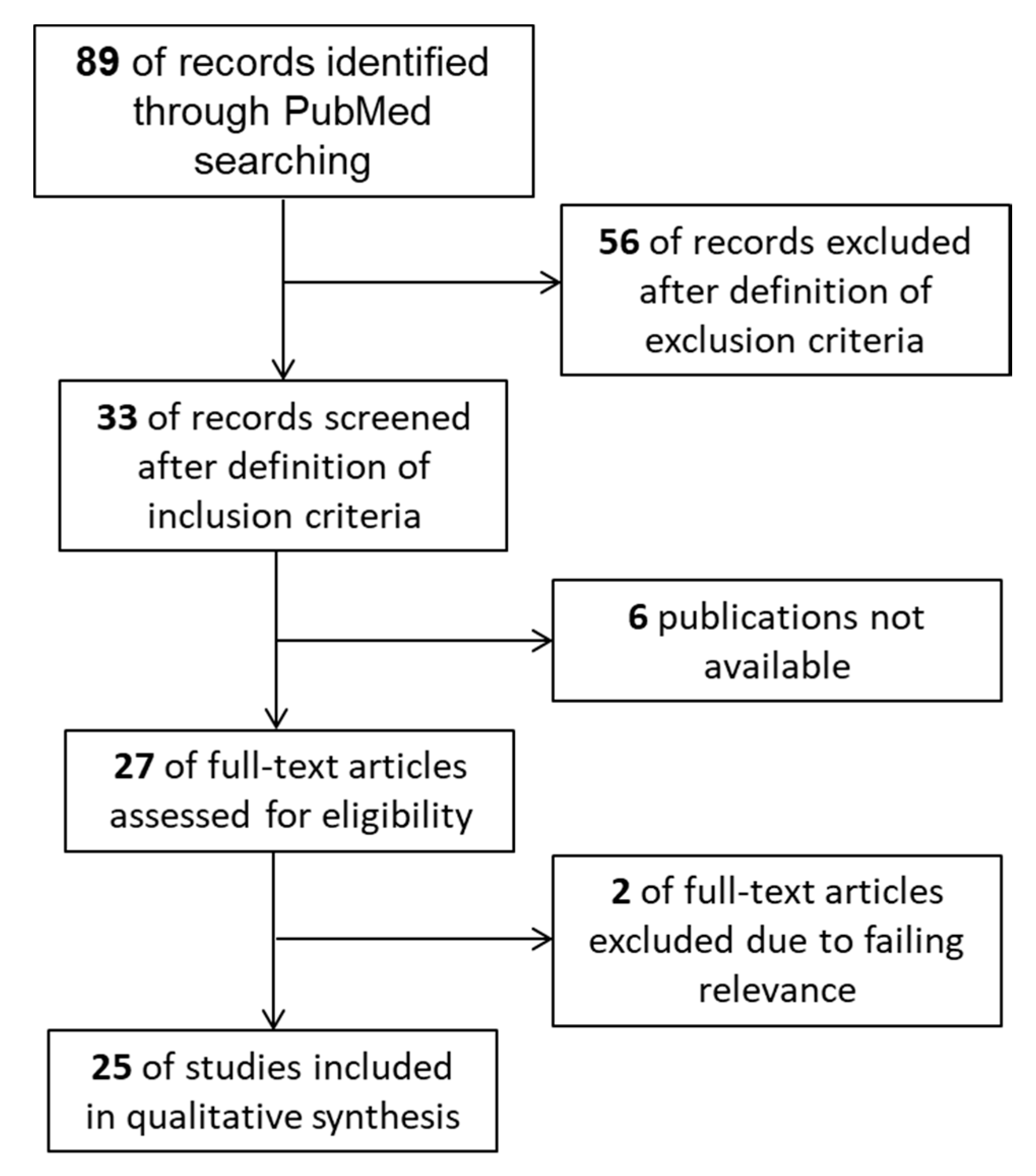
4. Materials and Methods
4.1. Study Design
4.2. Search Strategy
4.3. Study Selection
- Original publications regarding animals, humans (peripheral lymphocyte analysis and human biopsies), and in vitro studies (cell cultivation) were relevant;
- The results had to include the term ‘marginal periodontitis’;
- Reviews were excluded and were only used for discussing the outcome;
- Publications containing titles of other diseases, such as peri-apical periodontitis, saliva-associated diseases, hypophosphatasia, hypercholesterolemia, cancer, psychological stress disorders, and cranio-facial syndromes, were excluded;
- Abstracts fulfilling the inclusion criteria and dealing with different immune cells, but not with NK cells and NKT cells, were extracted.
4.4. Data Collection Process
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| A.a. | Aggregatibacter actinomycetemcomitans |
| CD | Cluster of differentiation |
| CRACC | CD2-like receptor-activating cytotoxic cell |
| CXCL | CXC-motif ligand chemokine |
| DC | Dendritic cell |
| F.n. | Fusobacterium nucleatum |
| gF | Gingival fibroblast |
| IgG | Immunoglobulin G |
| IL | Interleukin |
| IFN-ɣ | Interferon gamma |
| KIR | Killer-cell immunoglobulin-like receptor |
| LFA | Lymphocyte function-associated antigen |
| LPS | Lipopolysaccharide |
| NCR-1 | Natural cytotoxicity triggering receptor 1 |
| NK cell | Natural killer cell |
| NKT cell | Natural killer T cell |
| iNKT cell | Invariant natural killer T cell |
| MHC | Major histocompatibility complex |
| PB | Peripheral blood |
| P.g. | Porphyromonas gingivalis |
| TCR | T cell receptor |
| TLR | Toll-like receptor |
| TH | T helper cell |
| TNF | Tumor necrosis factor |
References
- Polak, D.; Shapira, L. An update on the evidence for pathogenic mechanisms that may link periodontitis and diabetes. J. Clin. Periodontol. 2018, 45, 150–166. [Google Scholar] [CrossRef]
- Mathews, M.J.; Mathews, E.H.; Mathews, G.E. Oral health and coronary heart disease. BMC Oral Health 2016, 16, 122. [Google Scholar] [CrossRef] [Green Version]
- Van Dyke, T.E.; Kornman, K.S. Inflammation and factors that may regulate inflammatory response. J. Periodontol. 2008, 79, 1503–1507. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Lamont, R.J. Beyond the red complex and into more complexity: The polymicrobial synergy and dysbiosis (PSD) model of periodontal disease etiology. Mol. Oral Microbiol. 2012, 27, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Hajishengallis, G.; Liang, S.; Payne, M.A.; Hashim, A.; Jotwani, R.; Eskan, M.A.; McIntosh, M.L.; Alsam, A.; Kirkwood, K.L.; Lambris, J.D.; et al. Low-abundance biofilm species orchestrates inflammatory periodontal disease through the commensal microbiota and complement. Cell Host Microbe. 2011, 10, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Darveau, R.P.; Hajishengallis, G.; Curtis, M.A. Porphyromonas gingivalis as a potential community activist for disease. J. Dent. Res. 2012, 91, 816–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajishengallis, G. Immunomicrobial pathogenesis of periodontitis: Keystones, pathobionts, and host response. Trends Immunol. 2014, 35, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dyke, T.E.; Lester, M.A.; Shapira, L. The role of the host response in periodontal disease progression: Implications for future treatment strategies. J. Periodontol. 1993, 64 (Suppl. S8), 792–806. [Google Scholar] [CrossRef]
- Grossi, S.G.; Zambon, J.J.; Ho, A.W.; Koch, G.; Dunford, R.G.; Machtei, E.E.; Norderyd, O.M.; Genco, R.J. Assessment of risk for periodontal disease. I. Risk indicators for attachment loss. J. Periodontol. 1994, 65, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Deo, V.; Bhongade, M.L. Pathogenesis of periodontitis: Role of cytokines in host response. Dent. Today 2010, 29, 60–62, 64–66; quiz 68–69. [Google Scholar] [PubMed]
- Diaz, P.I.; Hoare, A.; Hong, B.Y. Subgingival microbiome shifts and community dynamics in periodontal diseases. J. Calif. Dent. Assoc. 2016, 44, 421–435. [Google Scholar] [PubMed]
- Lamont, R.J.; Hajishengallis, G. Polymicrobial synergy and dysbiosis in inflammatory disease. Trends Mol. Med. 2015, 21, 172–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiessling, R.; Klein, E.; Wigzell, H. “Natural” killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype. Eur. J. Immunol. 1975, 5, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Wilensky, A.; Chaushu, S.; Shapira, L. The role of natural killer cells in periodontitis. Periodontol. 2000 2015, 69, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Loza, M.J.; Perussia, B. Final steps of natural killer cell maturation: A model for type 1-type 2 differentiation? Nat. Immunol. 2001, 2, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Sivori, S.; Cantoni, C.; Parolini, S.; Marcenaro, E.; Conte, R.; Moretta, L.; Moretta, A. IL-21 induces both rapid maturation of human CD34+ cell precursors towards NK cells and acquisition of surface killer Ig-like receptors. Eur. J. Immunol. 2003, 33, 3439–3447. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, W.M.; Kim, S.; French, A.R. The dynamic life of natural killer cells. Annu. Rev. Immunol. 2004, 22, 405–429. [Google Scholar] [CrossRef]
- Parisi, L.; Bassani, B.; Tremolati, M.; Gini, E.; Farronato, G.; Bruno, A. Natural killer cells in the orchestration of chronic inflammatory diseases. J. Immunol. Res. 2017, 2017, 4218254. [Google Scholar] [CrossRef]
- Clayton, A.; Mitchell, J.P.; Court, J.; Linnane, S.; Mason, M.D.; Tabi, Z. Human tumor-derived exosomes down-modulate NKG2D expression. J. Immunol. 2008, 180, 7249–7258. [Google Scholar] [CrossRef] [Green Version]
- Clayton, A.; Tabi, Z. Exosomes and the MICA-NKG2D system in cancer. Blood Cells Mol. Dis. 2005, 34, 206–213. [Google Scholar] [CrossRef]
- Muller, L.; Mitsuhashi, M.; Simms, P.; Gooding, W.E.; Whiteside, T.L. Tumor-derived exosomes regulate expression of immune function-related genes in human T cell subsets. Sci. Rep. 2016, 6, 20254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greening, D.W.; Gopal, S.K.; Xu, R.; Simpson, R.J.; Chen, W. Exosomes and their roles in immune regulation and cancer. Semin. Cell Dev. Biol. 2015, 40, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Webber, J.; Yeung, V.; Clayton, A. Extracellular vesicles as modulators of the cancer microenvironment. Semin. Cell Dev. Biol. 2015, 40, 27–34. [Google Scholar] [CrossRef]
- Baginska, J.; Viry, E.; Paggetti, J.; Medves, S.; Berchem, G.; Moussay, E.; Janji, B. The critical role of the tumor microenvironment in shaping natural killer cell-mediated anti-tumor immunity. Front. Immunol. 2013, 4, 490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenbarth, G.S. Type I diabetes mellitus. A chronic autoimmune disease. N. Engl. J. Med. 1986, 314, 1360–1368. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.A.; Rayner, M.L.; Mijovic, C.H.; Barnett, A.H. Molecular aspects of type 1 diabetes. Mol. Pathol. 2003, 56, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.G.; Smyth, M.J. The role of NK cells in autoimmune disease. Autoimmunity 2002, 35, 1–14. [Google Scholar] [CrossRef]
- Rodacki, M.; Milech, A.; de Oliveira, J.E. NK cells and type 1 diabetes. Clin. Dev. Immunol. 2006, 13, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Rodacki, M.; Svoren, B.; Butty, V.; Besse, W.; Laffel, L.; Benoist, C.; Mathis, D. Altered natural killer cells in type 1 diabetic patients. Diabetes 2007, 56, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Bobryshev, Y.V.; Lord, R.S. Identification of natural killer cells in human atherosclerotic plaque. Atherosclerosis 2005, 180, 423–427. [Google Scholar] [CrossRef]
- Whitman, S.C.; Rateri, D.L.; Szilvassy, S.J.; Yokoyama, W.; Daugherty, A. Depletion of natural killer cell function decreases atherosclerosis in low-density lipoprotein receptor null mice. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1049–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allavena, P.; Bianchi, G.; Zhou, D.; van Damme, J.; Jílek, P.; Sozzani, S.; Mantovani, A. Induction of natural killer cell migration by monocyte chemotactic protein-1, -2 and -3. Eur. J. Immunol. 1994, 24, 3233–3236. [Google Scholar] [CrossRef] [PubMed]
- Umehara, H.; Bloom, E.T.; Okazaki, T.; Nagano, Y.; Yoshie, O.; Imai, T. Fractalkine in vascular biology: From basic research to clinical disease. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Sobenin, I.A.; Orekhov, A.N.; Bobryshev, Y.V. Dendritic cells in atherosclerotic inflammation: The complexity of functions and the peculiarities of pathophysiological effects. Front. Physiol. 2014, 5, 196. [Google Scholar] [CrossRef] [PubMed]
- Mallat, Z.; Corbaz, A.; Scoazec, A.; Graber, P.; Alouani, S.; Esposito, B.; Humbert, Y.; Chvatchko, Y.; Tedgui, A. Interleukin-18/interleukin-18 binding protein signaling modulates atherosclerotic lesion development and stability. Circ. Res. 2001, 89, E41–E45. [Google Scholar] [CrossRef] [Green Version]
- Uyemura, K.; Demer, L.L.; Castle, S.C.; Jullien, D.; Berliner, J.A.; Gately, M.K.; Warrier, R.R.; Pham, N.; Fogelman, A.M.; Modlin, R.L. Cross-regulatory roles of interleukin (IL)-12 and IL-10 in atherosclerosis. J. Clin. Investig. 1996, 97, 2130–2138. [Google Scholar] [CrossRef]
- Bonaccorsi, I.; De Pasquale, C.; Campana, S.; Barberi, C.; Cavaliere, R.; Benedetto, F.; Ferlazzo, G. Natural killer cells in the innate immunity network of atherosclerosis. Immunol. Lett. 2015, 168, 51–57. [Google Scholar] [CrossRef]
- Van Kaer, L.; Parekh, V.V.; Wu, L. Invariant natural killer T cells: Bridging innate and adaptive immunity. Cell Tissue Res. 2011, 343, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Bendelac, A.; Savage, P.B.; Teyton, L. The biology of NKT cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef] [Green Version]
- Brigl, M.; Brenner, M.B. CD1: Antigen presentation and T cell function. Annu. Rev. Immunol. 2004, 22, 817–890. [Google Scholar] [CrossRef]
- Godfrey, D.I.; MacDonald, H.R.; Kronenberg, M.; Smyth, M.J.; Van Kaer, L. NKT cells: What’s in a name? Nat. Rev. Immunol. 2004, 4, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Arrenberg, P.; Halder, R.; Kumar, V. Cross-regulation between distinct natural killer T cell subsets influences immune response to self and foreign antigens. J. Cell Physiol. 2009, 218, 246–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, J.L.; Mallevaey, T.; Scott-Browne, J.; Gapin, L. CD1d-restricted iNKT cells, the ‘Swiss-Army knife’ of the immune system. Curr. Opin. Immunol. 2008, 20, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parekh, V.V.; Wilson, M.T.; Olivares-Villagómez, D.; Singh, A.K.; Wu, L.; Wang, C.R.; Joyce, S.; van Kaer, L. Glycolipid antigen induces long-term natural killer T cell anergy in mice. J. Clin. Investig. 2005, 115, 2572–2583. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, D.I.; Kronenberg, M. Going both ways: Immune regulation via CD1d-dependent NKT cells. J. Clin. Investig. 2004, 114, 1379–1388. [Google Scholar] [CrossRef] [Green Version]
- Nowak, M.; Stein-Streilein, J. Invariant NKT cells and tolerance. Int. Rev. Immunol. 2007, 26, 95–119. [Google Scholar] [CrossRef]
- Pillai, A.B.; George, T.I.; Dutt, S.; Teo, P.; Strober, S. Host NKT cells can prevent graft-versus-host disease and permit graft antitumor activity after bone marrow transplantation. J. Immunol. 2007, 178, 6242–6251. [Google Scholar] [CrossRef] [Green Version]
- Boeck, C.L.; Amberger, D.C.; Doraneh-Gard, F.; Sutanto, W.; Guenther, T.; Schmohl, J.; Schuster, F.; Salih, H.; Babor, F.; Borkhardt, A.; et al. Significance of frequencies, compositions, and/or antileukemic activity of (DC-stimulated) invariant NKT, NK and CIK cells on the outcome of patients with AML, ALL and CLL. J. Immunother. 2017, 40, 224–248. [Google Scholar] [CrossRef]
- Terabe, M.; Berzofsky, J.A. The role of NKT cells in tumor immunity. Adv. Cancer Res. 2008, 101, 277–348. [Google Scholar] [CrossRef] [Green Version]
- Van Kaer, L. NKT cells: T lymphocytes with innate effector functions. Curr. Opin. Immunol. 2007, 19, 354–364. [Google Scholar] [CrossRef]
- Coppieters, K.; Van Beneden, K.; Jacques, P.; Dewint, P.; Vervloet, A.; Vander Cruyssen, B.; van Calenbergh, S.; Chen, G.; Franck, R.W.; Verbruggen, G.; et al. A single early activation of invariant NK T cells confers long-term protection against collagen-induced arthritis in a ligand-specific manner. J. Immunol. 2007, 179, 2300–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, S.; Van Kaer, L. Invariant natural killer T cells trigger adaptive lymphocytes to churn up bile. Cell Host Microbe 2008, 3, 275–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komiyama, K.; Hirsch, H.Z.; Moro, I.; Umemura, S.; Mestecky, J. HNK-1+ (Leu-7) cells and natural killer cell activity in inflamed human gingival tissue. J. Oral. Pathol. 1988, 17, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Kopp, W. Density and localization of lymphocytes with natural-killer (NK) cell activity in periodontal biopsy specimens from patients with severe periodontitis. J. Clin. Periodontol. 1988, 15, 595–600. [Google Scholar] [CrossRef]
- Cobb, C.M.; Singla, O.; Feil, P.H.; Theisen, F.C.; Schultz, R.E. Comparison of NK-cell (Leu-7+ and Leu-11b+) populations in clinically healthy gingiva, chronic gingivitis and chronic adult periodontitis. J. Periodontal. Res. 1989, 24, 1–7. [Google Scholar] [CrossRef]
- Lindemann, R.A.; Eilber, F. Activation of human natural killer cells by lipopolysaccharide from Actinobacillus actinomycetemcomitans. Arch. Oral Biol. 1989, 34, 459–463. [Google Scholar] [CrossRef]
- Celenligil, H.; Kansu, E.; Eratalay, K. Juvenile and rapidly progressive periodontitis. Peripheral blood lymphocyte subpopulations. J. Clin. Periodontol. 1990, 17, 207–210. [Google Scholar] [CrossRef]
- Afar, B.; Engel, D.; Clark, E.A. Activated lymphocyte subsets in adult periodontitis. J. Periodontal. Res. 1992, 27, 126–133. [Google Scholar] [CrossRef]
- Fujita, S.; Takahashi, H.; Okabe, H.; Ozaki, Y.; Hara, Y.; Kato, I. Distribution of natural killer cells in periodontal diseases: An immunohistochemical study. J. Periodontol. 1992, 63, 686–689. [Google Scholar] [CrossRef]
- Firatli, E.; Gürel, N.; Efeoğlu, A.; Cebeci, I. Generalized prepubertal periodontitis. A report of 4 cases with the immunological findings. J. Clin. Periodontol. 1996, 23, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Mahanonda, R.; Sa-Ard-Iam, N.; Charatkulangkun, O.; Promsudthi, A.; Schifferle, R.E.; Yongvanichit, K.; Pichyangkul, S. Monocyte activation by Porphyromonas gingivalis LPS in aggressive periodontitis with the use of whole-blood cultures. J. Dent. Res. 2004, 83, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Hahn, C.L.; Tanaka, S.; Barbour, S.E.; Schenkein, H.A.; Tew, J.G. Dendritic cells stimulated with Actinobacillus actinomycetemcomitans elicit rapid gamma interferon responses by natural killer cells. Infect. Immun. 2004, 72, 5089–5096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, T.; Willis, D.L.; Liu, M.; Purkall, D.B.; Sukumar, S.; Barbour, S.E.; Schenkein, H.A.; Tew, J.G. Dendritic-NK cell interactions in P. gingivalis-specific responses. J. Dent. Res. 2005, 84, 858–862. [Google Scholar] [CrossRef]
- Stelin, S.; Ramakrishan, H.; Talwar, A.; Arun, K.V.; Kumar, T.S. Immunohistological analysis of CD1a langerhans cells and CD57 natural killer cells in healthy and diseased human gingival tissue: A comparative study. J. Indian Soc. Periodontol. 2009, 13, 150–154. [Google Scholar] [CrossRef]
- Chaushu, S.; Wilensky, A.; Gur, C.; Shapira, L.; Elboim, M.; Halftek, G.; Polak, D.; Achdout, H.; Bachrach, G.; Mandelboim, O. Direct recognition of Fusobacterium nucleatum by the NK cell natural cytotoxicity receptor NKp46 aggravates periodontal disease. PLoS Pathog. 2012, 8, e1002601. [Google Scholar] [CrossRef] [Green Version]
- Krämer, B.; Kebschull, M.; Nowak, M.; Demmer, R.T.; Haupt, M.; Körner, C.; Perner, S.; Jepsen, S.; Nattermann, J.; Papapanou, P.N. Role of the NK cell-activating receptor CRACC in periodontitis. Infect. Immun. 2013, 81, 690–696. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, O.A.; Novak, M.J.; Kirakodu, S.; Orraca, L.; Chen, K.C.; Stromberg, A.; Gonzalez-Martinez, J.; Ebersole, J.L. Comparative analysis of gingival tissue antigen presentation pathways in ageing and periodontitis. J. Clin. Periodontol. 2014, 41, 327–339. [Google Scholar] [CrossRef]
- Takeda, H.; Kikuchi, T.; Soboku, K.; Okabe, I.; Mizutani, H.; Mitani, A.; Ishihara, Y.; Noguchi, T. Effect of IL-15 and natural killer cells on osteoclasts and osteoblasts in a mouse coculture. Inflammation 2014, 37, 657–669. [Google Scholar] [CrossRef]
- Mazurek-Mochol, M.; Majorczyk, E.; Banach, J.; Dembowska, E.; Pawlik, A.; Kuśnierczyk, P. Are KIR genes associated with clinical parameters in the course of periodontitis? Postepy Hig. Med. Dosw. (Online) 2014, 68, 1145–1151. [Google Scholar] [CrossRef]
- Mazurek-Mochol, M.; Majorczyk, E.; Banach, J.; Dembowska, E.; Kuśnierczyk, P.; Safranow, K.; Pawlik, A. The influence of KIR gene presence/absence polymorphisms on the development of periodontal disease in smokers and non-smokers. Cent. Eur. J. Immunol. 2017, 42, 347–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moonen, C.G.J.; Alders, S.T.; Bontkes, H.J.; Schoenmaker, T.; Nicu, E.A.; Loos, B.G.; de Vries, T.J. Survival, retention, and selective proliferation of lymphocytes is mediated by gingival fibroblasts. Front. Immunol. 2018, 9, 1725. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Ohsawa, Y.; Yoshie, H. Elevated proportion of natural killer T cells in periodontitis lesions: A common feature of chronic inflammatory diseases. Am. J. Pathol. 2001, 158, 1391–1398. [Google Scholar] [CrossRef]
- Amanuma, R.; Nakajima, T.; Yoshie, H.; Yamazaki, K. Increased infiltration of CD1d and natural killer T cells in periodontal disease tissues. J. Periodontal. Res. 2006, 41, 73–79. [Google Scholar] [CrossRef]
- Nowak, M.; Krämer, B.; Haupt, M.; Papapanou, P.N.; Kebschull, J.; Hoffmann, P.; Schmidt-Wolf, I.G.; Jepsen, S.; Brossart, P.; Perner, S.; et al. Activation of invariant NK T cells in periodontitis lesions. J. Immunol. 2013, 190, 2282–2291. [Google Scholar] [CrossRef] [Green Version]
- Aoki-Nonaka, Y.; Nakajima, T.; Miyauchi, S.; Miyazawa, H.; Yamada, H.; Domon, H.; Tabeta, K.; Yamazaki, K. Natural killer T cells mediate alveolar bone resorption and a systemic inflammatory response in response to oral infection of mice with Porphyromonas gingivalis. J. Periodontal. Res. 2014, 49, 69–76. [Google Scholar] [CrossRef]
- Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Nakae, H.; Matsuo, T. CXC chemokine ligand 16 in periodontal diseases: Expression in diseased tissues and production by cytokine-stimulated human gingival fibroblasts. Clin. Exp. Immunol. 2007, 149, 146–154. [Google Scholar] [CrossRef]
- Muthukuru, M. Technical advance: Decreased helper T cells and increased natural killer cells in chronic periodontitis analyzed by a novel method for isolating resident lymphocytes. J. Leukoc. Biol. 2012, 92, 683–692. [Google Scholar] [CrossRef]
- Pessino, A.; Sivori, S.; Bottino, C.; Malaspina, A.; Morelli, L.; Moretta, L.; Biassoni, R.; Moretta, A. Molecular cloning of NKp46: A novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicity. J. Exp. Med. 1998, 188, 953–960. [Google Scholar] [CrossRef] [Green Version]
- Vitale, M.; Bottino, C.; Sivori, S.; Sanseverino, L.; Castriconi, R.; Marcenaro, E.; Augugliaro, R.; Moretta, L.; Moretta, A. NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non-major histocompatibility complex-restricted tumor cell lysis. J. Exp. Med. 1998, 187, 2065–2072. [Google Scholar] [CrossRef]
- Pende, D.; Parolini, S.; Pessino, A.; Sivori, S.; Augugliaro, R.; Morelli, L.; Marcenaro, E.; Accame, L.; Malaspina, A.; Biassoni, R.; et al. Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J. Exp. Med. 1999, 190, 1505–1516. [Google Scholar] [CrossRef] [PubMed]
- Biassoni, R.; Cantoni, C.; Pende, D.; Sivori, S.; Parolini, S.; Vitale, M.; Bottino, C.; Moretta, A. Human natural killer cell receptors and co-receptors. Immunol. Rev. 2001, 181, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Meyle, J.; Chapple, I. Molecular aspects of the pathogenesis of periodontitis. Periodontol. 2000 2015, 69, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, J.R. T- and B-cell subsets in periodontitis. Periodontol. 2000 2015, 69, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Górska, R.; Gregorek, H.; Kowalski, J.; Laskus-Perendyk, A.; Syczewska, M.; Madaliński, K. Relationship between clinical parameters and cytokine profiles in inflamed gingival tissue and serum samples from patients with chronic periodontitis. J. Clin. Periodontol. 2003, 30, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Gemmell, E.; Yamazaki, K.; Seymour, G.J. The role of T cells in periodontal disease: Homeostasis and autoimmunity. Periodontol. 2000 2007, 43, 14–40. [Google Scholar] [CrossRef]
- Kajiya, M.; Giro, G.; Taubman, M.A.; Han, X.; Mayer, M.P.; Kawai, T. Role of periodontal pathogenic bacteria in RANKL-mediated bone destruction in periodontal disease. J. Oral Microbiol. 2010, 2. [Google Scholar] [CrossRef]
- De Araújo, M.F.; Etchebehere, R.M.; de Melo, M.L.R.; Beghini, M.; Severino, V.O.; de Castro Côbo, E.; Rocha Rodrigues, D.B.; de Lima Pereira, S.A. Analysis of CD15, CD57 and HIF-1α in biopsies of patients with peri-implantitis. Pathol. Res. Pract. 2017, 213, 1097–1101. [Google Scholar] [CrossRef]
- Heitz-Mayfield, L.J.; Lang, N.P. Comparative biology of chronic and aggressive periodontitis vs. peri-implantitis. Periodontol. 2000 2010, 53, 167–181. [Google Scholar] [CrossRef]
- Brutkiewicz, R.R. CD1d ligands: The good, the bad, and the ugly. J. Immunol. 2006, 177, 769–775. [Google Scholar] [CrossRef] [Green Version]
- Venkataswamy, M.M.; Porcelli, S.A. Lipid and glycolipid antigens of CD1d-restricted natural killer T cells. Semin. Immunol. 2010, 22, 68–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berglundh, T.; Donati, M.; Zitzmann, N. B cells in periodontitis: Friends or enemies? Periodontol. 2000 2007, 45, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Hasturk, H.; Kantarci, A.; Van Dyke, T.E. Paradigm shift in the pharmacological management of periodontal diseases. Front. Oral Biol. 2012, 15, 160–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seidel, A.; Seidel, C.L.; Weider, M.; Junker, R.; Gölz, L.; Schmetzer, H. Influence of Natural Killer Cells and Natural Killer T Cells on Periodontal Disease: A Systematic Review of the Current Literature. Int. J. Mol. Sci. 2020, 21, 9766. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249766
Seidel A, Seidel CL, Weider M, Junker R, Gölz L, Schmetzer H. Influence of Natural Killer Cells and Natural Killer T Cells on Periodontal Disease: A Systematic Review of the Current Literature. International Journal of Molecular Sciences. 2020; 21(24):9766. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249766
Chicago/Turabian StyleSeidel, Andreas, Corinna L. Seidel, Matthias Weider, Rüdiger Junker, Lina Gölz, and Helga Schmetzer. 2020. "Influence of Natural Killer Cells and Natural Killer T Cells on Periodontal Disease: A Systematic Review of the Current Literature" International Journal of Molecular Sciences 21, no. 24: 9766. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249766