Spatio-Temporal Bone Remodeling after Hematopoietic Stem Cell Transplantation

 , , ,
, , , 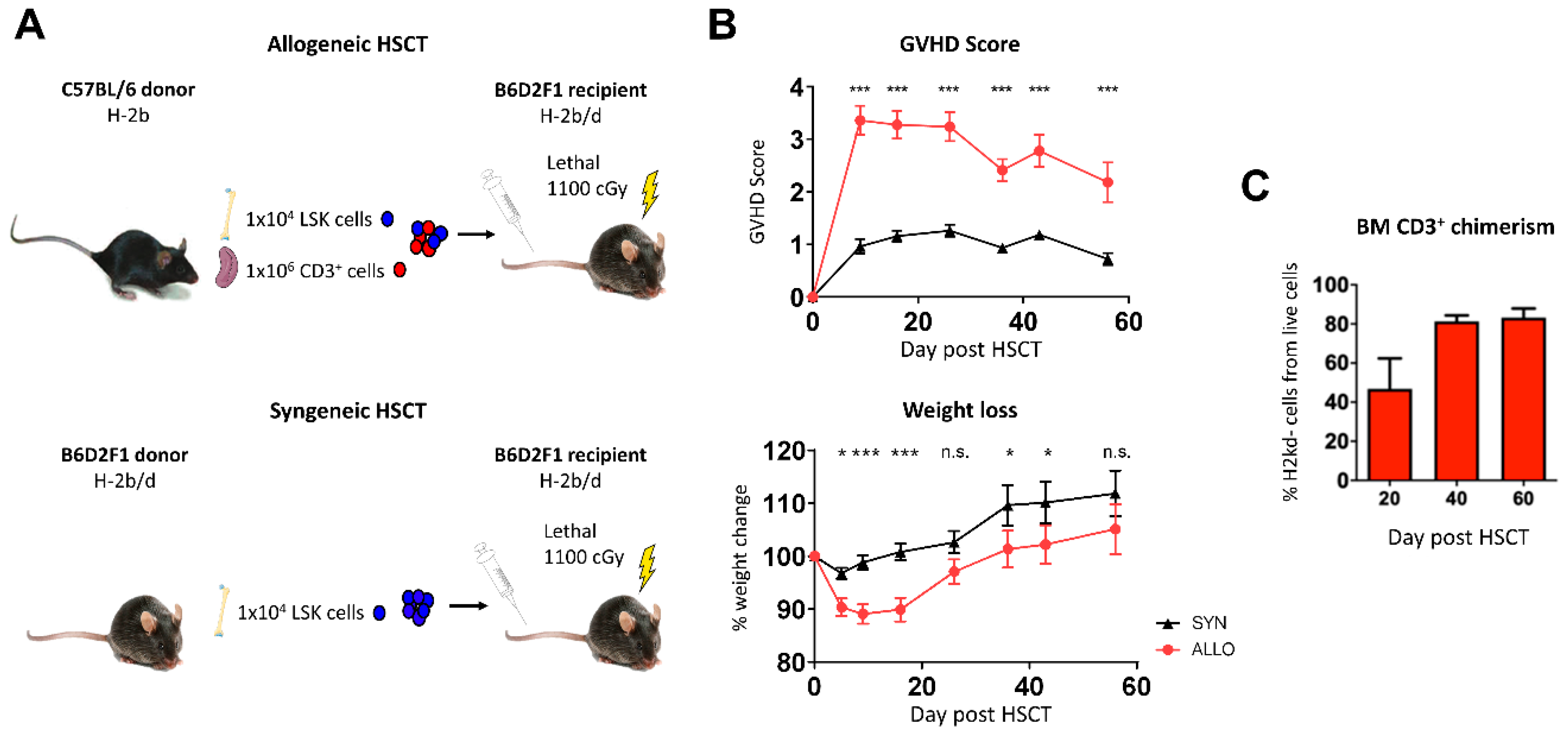{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Bone Marrow Infiltration of Donor T Lymphocytes after Allo-HSCT
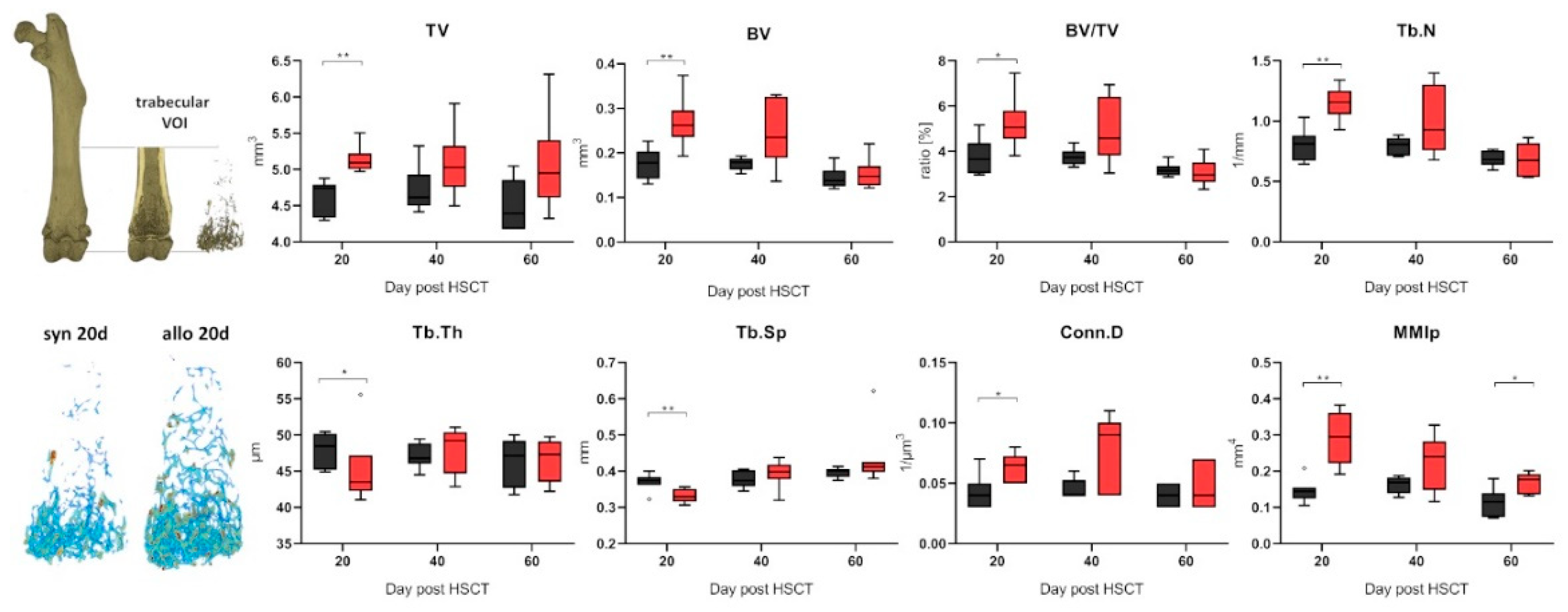
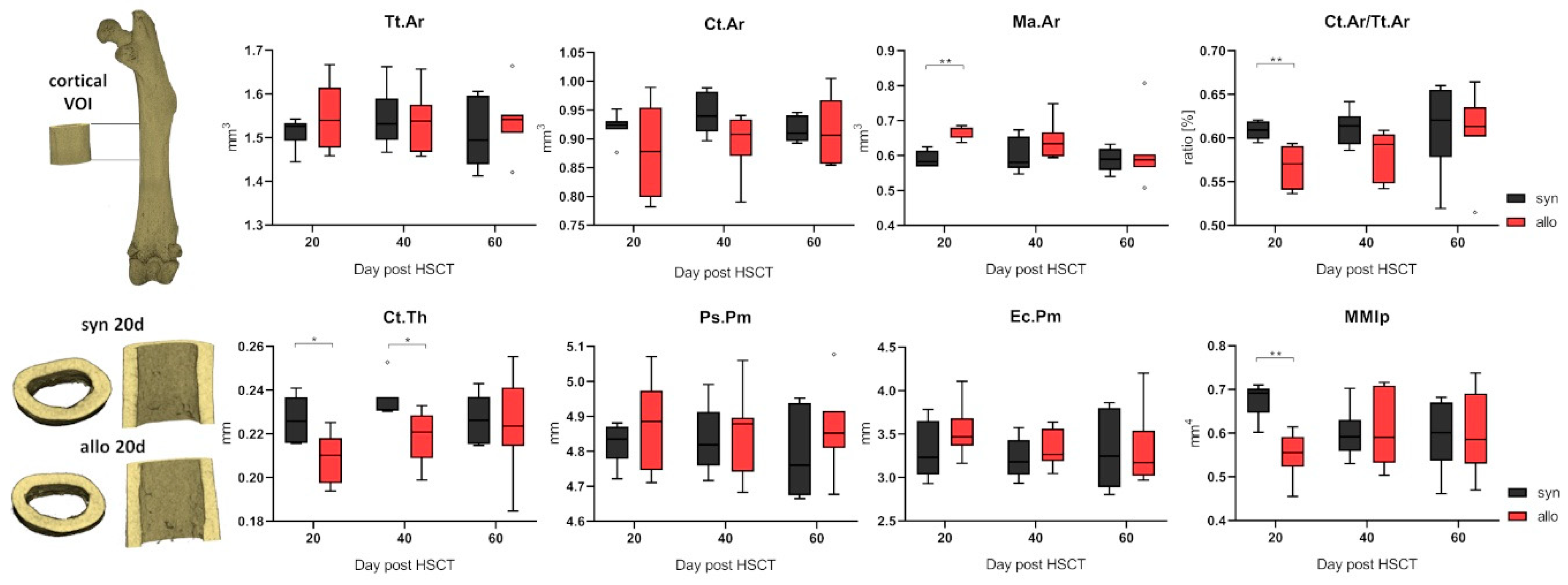
2.2. Increased Trabecular Bone Number and Decreased Cortical Thickness in aGVHD
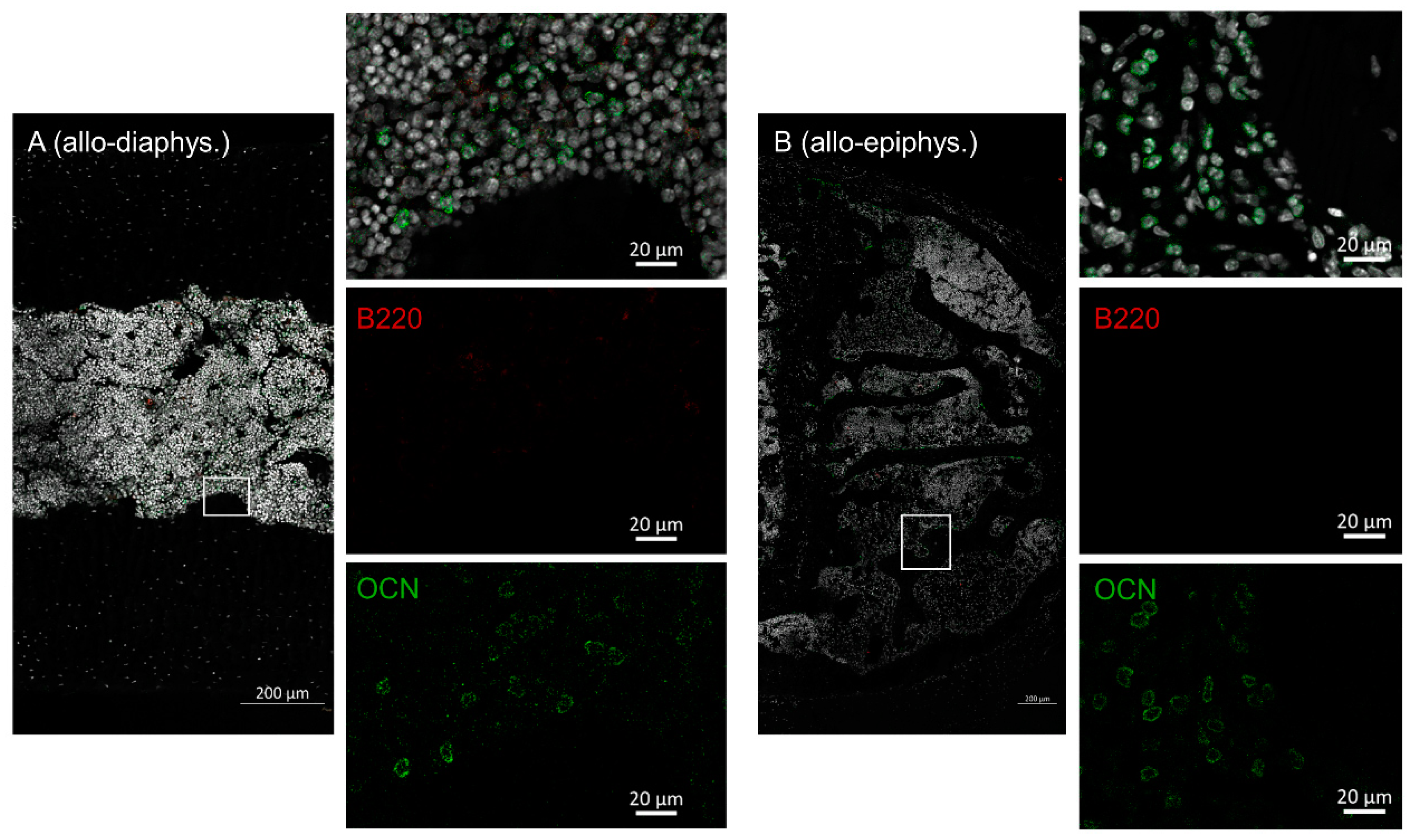
2.3. Disruptive Osteoblast Lining in Bones during aGVHD
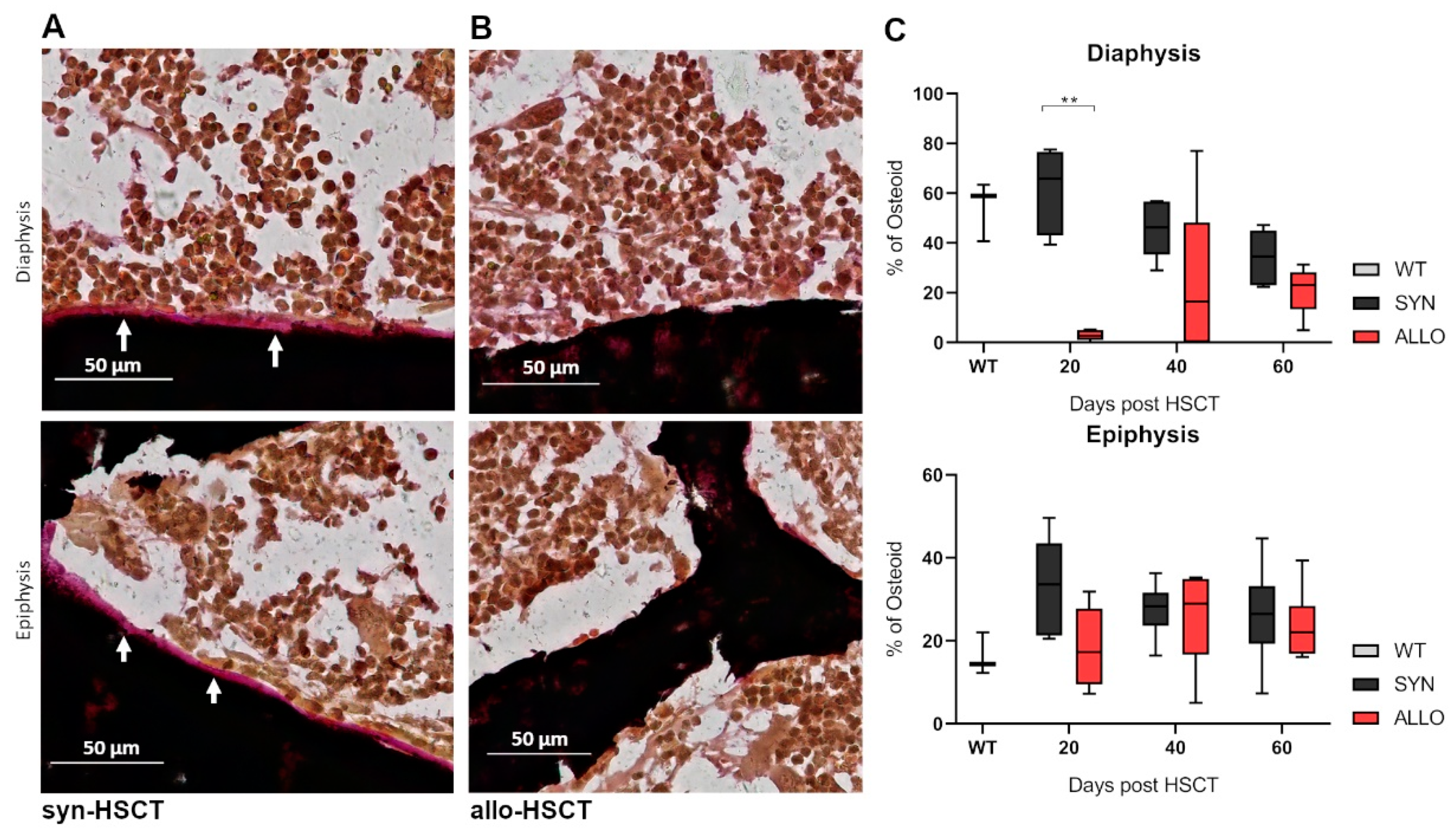
2.4. Decreased Osteoid Lining in Diaphyseal Region of Bones in aGVHD
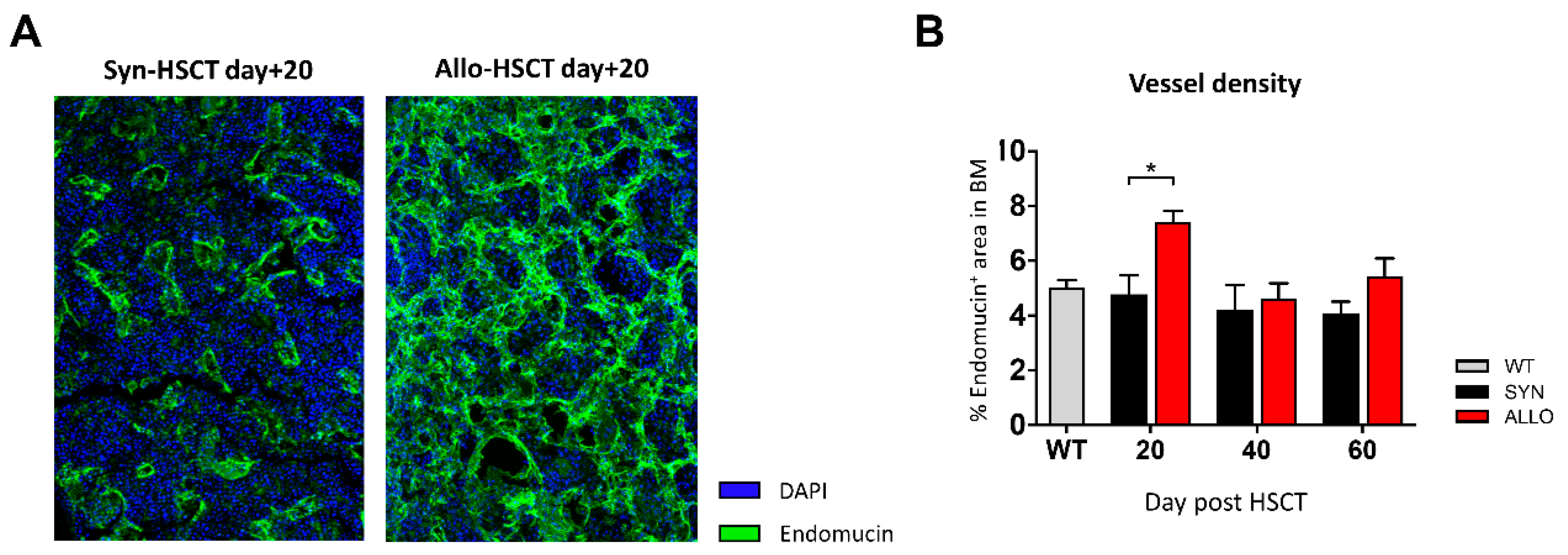
2.5. Increased BM Vessel Density in aGVHD
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Experimental Hematopoietic Stem Cell Transplantation Protocol
- (1)
- Donor cell isolation: On day +0 of the HSCT, donor mice (B6 for allo- and BDF for syn-HSCT) were sacrificed by cervical dislocation; bones (femora and tibiae) and spleens were harvested; LSK cells were isolated out of the BM; CD3+ cells were isolated out of the spleen.
- (2)
- Conditioning: On day +0 of the HSCT, recipient mice (BDF for both groups) received myeloablative total body irradiation (TBI).
- (3)
- Transplantation: On day +0 of the HSCT, irradiated recipient mice (BDF) were injected intravenously (i.v.) with isolated LSK cells and the allo-HSCT group additionally with CD3+ cells.
4.3. LSK Cell Isolation
4.4. T Cell Isolation
4.5. Conditioning and Cell Transplantation
4.6. GVHD Monitoring
4.7. Chimerism Analysis
4.8. Flow Cytometry
4.9. X-ray Microtomographic Analysis
4.10. Histology of Murine Bones—Osteoblast and B Cell Staining
4.11. Histology of Murine Bones—Osteoid Staining
4.12. Histology of Murine Bones—Endomucin Staining
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bosch, M.; Khan, F.M.; Storek, J. Immune reconstitution after hematopoietic cell transplantation. Curr. Opin. Hematol. 2012, 19, 324–355. [Google Scholar] [CrossRef] [PubMed]
- Storek, J.; Geddes, M.; Khan, F.; Huard, B.; Helg, C.; Chalandon, Y.; Passweg, J.; Roosnek, E. Reconstitution of the immune system after hematopoietic stem cell transplantation in humans. Semin. Immunopathol. 2008, 30, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Mackall, C.; Fry, T.; Gress, R.; Peggs, K.; Storek, J.; Toubert, A. Background to hematopoietic cell transplantation, including post transplant immune recovery. Bone Marrow Transplant. 2009, 44, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Park, B.; Yoo, K.H.; Kim, C. Hematopoietic stem cell expansion and generation: The ways to make a breakthrough. Blood Res. 2015, 50, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Holtan, S.G.; Marcelo, P.; Weisdorf, D.J. Acute graft-versus-host disease: A bench-to-bedside update. Blood 2014, 124, 363–373. [Google Scholar] [CrossRef]
- Jacobsohn, D.A.; Vogelsang, G.B. Acute graft versus host disease. Orphanet J. Rare Dis. 2007, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Zeiser, R.; Blazar, B.R. Acute graft-versus-host disease—Biologic process, prevention, and therapy. N. Engl. J. Med. 2017, 377, 2167–2179. [Google Scholar] [CrossRef]
- McClune, B.L.; Majhail, N.S. Osteoporosis after stem cell transplantation. Curr. Osteoporos. Rep. 2013, 11, 305–310. [Google Scholar] [CrossRef]
- Pundole, X.N.; Barbo, A.G.; Lin, H.; Champlin, R.E.; Lu, H. Increased incidence of fractures in recipients of hematopoietic stem-cell transplantation. J. Clin. Oncol. 2015, 33, 1364–1370. [Google Scholar] [CrossRef] [Green Version]
- Stern, J.M.; Sullivan, K.M.; Ott, S.M.; Seidel, K.; Fink, J.C.; Longton, G.; Sherrard, D.J. Bone density loss after allogeneic hematopoietic stem cell transplantation: A prospective study. Biol. Blood Marrow Transplant. 2001, 7, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.T.; Colgan, S.P. Regulation of immunity and inflammation by hypoxia in immunological niches. Nat. Rev. Immunol. 2017, 17, 774–785. [Google Scholar] [CrossRef] [PubMed]
- García-García, A.; de Castillejo, C.L.F.; Méndez-Ferrer, S. BMSCs and hematopoiesis. Immunol. Lett. 2015, 168, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Dar, H.Y.; Azam, Z.; Anupam, R.; Mondal, R.K.; Srivastava, R.K. Osteoimmunology: The Nexus between bone and immune system. Front. Biosci. Landmark 2018, 23, 464–492. [Google Scholar] [CrossRef] [Green Version]
- Sass, F.A.; Fuchs, M.; Pumberger, M.; Geissler, S.; Duda, G.N.; Perka, C.; Schmidt-Bleek, K. Immunology guides skeletal muscle regeneration. Int. J. Mol. Sci. 2018, 19, 835. [Google Scholar] [CrossRef] [Green Version]
- Kendler, D.L.; Body, J.J.; Brandi, M.L.; Broady, R.; Cannata-Andia, J.; Cannata-Ortiz, M.J.; El Maghraoui, A.; Guglielmi, G.; Hadji, P.; Pierroz, D.D.; et al. Bone management in hematologic stem cell transplant recipients. Osteoporos. Int. 2018, 29, 2597–2610. [Google Scholar] [CrossRef]
- Shono, Y.; Ueha, S.; Wang, Y.; Abe, J.; Kurachi, M.; Matsuno, Y.; Sugiyama, T.; Nagasawa, T.; Imamura, M.; Matsushima, K. Bone marrow graft-versus-host disease: Early destruction of hematopoietic niche after MHC-mismatched hematopoietic stem cell transplantation. Blood 2010, 115, 5401–5411. [Google Scholar] [CrossRef]
- Mensen, A.; Jöhrens, K.; Anagnostopoulos, I.; Demski, S.; Oey, M.; Stroux, A.; Hemmati, P.; Westermann, J.; Blau, O.; Wittenbecher, F.; et al. Bone marrow T-cell infiltration during acute GVHD is associated with delayed B-cell recovery and function after HSCT. Blood 2014, 124, 963–972. [Google Scholar] [CrossRef]
- Szyska, M.; Na, I.K. Bone marrow GvHD after allogeneic hematopoietic stem cell transplantation. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, A.; Hill, W.; Stemmler, H.J.; Ledderose, G.; Baur-Melnyk, A.; Fritsch, S.; Ullmann, J.; Kolb, H.-J.; Geiger, S.; Tischer, J. Bone Loss after Allogeneic Haematopoietic Stem Cell Transplantation: A Pilot Study on the Use of Zoledronic Acid. Chemother. Res. Pract. 2012, 2012, 1–7. [Google Scholar] [CrossRef]
- Cohen, A.; Dempster, D.W.; Müller, R.; Guo, X.E.; Nickolas, T.L.; Liu, X.S.; Zhang, X.H.; Wirth, A.J.; Van Lenthe, G.H.; Kohler, T.; et al. Assessment of trabecular and cortical architecture and mechanical competence of bone by high-resolution peripheral computed tomography: Comparison with transiliac bone biopsy. Osteoporos. Int. 2010, 21, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Neumann, A.; Haschka, J.; Kleyer, A.; Schuster, L.; Englbrecht, M.; Berlin, A.; Figueiredo, C.P.; Simon, D.; Muschitz, C.; Kocijan, R.; et al. Cortical bone loss is an early feature of nonradiographic axial spondyloarthritis. Arthritis Res. Ther. 2018, 20, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickolas, T.L.; Stein, E.M.; Dworakowski, E.; Nishiyama, K.K.; Komandah-Kosseh, M.; Zhang, C.A.; McMahon, D.J.; Liu, X.S.; Boutroy, S.; Cremers, S.; et al. Rapid cortical bone loss in patients with chronic kidney disease. J. Bone Miner. Res. 2013, 28, 1811–1820. [Google Scholar] [CrossRef] [PubMed]
- Saiworn, W.; Thim-uam, A.; Visitchanakun, P.; Atjanasuppat, K.; Chantaraaumporn, J.; Mokdara, J.; Chungchatupornchai, S.; Pisitkun, P.; Leelahavanichkul, A.; Poolthong, S.; et al. Cortical Bone Loss in a Spontaneous Murine Model of Systemic Lupus Erythematosus. Calcif. Tissue Int. 2018, 103, 686–697. [Google Scholar] [CrossRef]
- MacDonald, H.M.; Nishiyama, K.K.; Kang, J.; Hanley, D.A.; Boyd, S.K. Age-related patterns of trabecular and cortical bone loss differ between sexes and skeletal sites: A population-based HR-pQCT study. J. Bone Miner. Res. 2011, 26, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Almeida, M.; Weinstein, R.S.; O’Brien, C.A.; Manolagas, S.C.; Jilka, R.L. Skeletal inflammation and attenuation of Wnt signaling, Wnt ligand expression, and bone formation in atherosclerotic ApoE-null mice. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E762–E773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucher, C.H.; Schlundt, C.; Wulsten, D.; Sass, F.A.; Wendler, S.; Ellinghaus, A.; Thiele, T.; Seemann, R.; Willie, B.M.; Volk, H.D.; et al. Experience in the adaptive immunity impacts bone homeostasis, remodeling, and healing. Front. Immunol. 2019, 10, 797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zhou, X.; Fujita, H.; Onozuka, M.; Kubo, K.Y. Age-related changes in trabecular and cortical bone microstructure. Int. J. Endocrinol. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Panaroni, C.; Wu, J.Y. Interactions between b lymphocytes and the osteoblast lineage in bone marrow. Calcif. Tissue Int. 2013, 93, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.M.S.; Linderman, J.A.; Florek, M.; Miklos, D.; Shizuru, J.A. Allogeneic T cells impair engraftment and hematopoiesis after stem cell transplantation. Proc. Natl. Acad. Sci. USA 2010, 107, 14721–14726. [Google Scholar] [CrossRef] [Green Version]
- Heitger, A.; Neu, N.; Kern, H.; Panzer-Grümayer, E.R.; Greinix, H.; Nachbaur, D.; Niederwieser, D.; Fink, F.M. Essential role of the thymus to reconstitute naive (CD45RA+) T-helper cells after human allogeneic bone marrow transplantation. Blood 1997, 90, 850–857. [Google Scholar] [CrossRef]
- Zhu, J.; Garrett, R.; Jung, Y.; Zhang, Y.; Kim, N.; Wang, J.; Joe, G.J.; Hexner, E.; Choi, Y.; Taichman, R.S.; et al. Osteoblasts support B-lymphocyte commitment and differentiation from hematopoietic stem cells. Blood 2007, 109, 3706–3712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terashima, A.; Okamoto, K.; Nakashima, T.; Akira, S.; Ikuta, K.; Takayanagi, H. Sepsis-Induced Osteoblast Ablation Causes Immunodeficiency. Immunity 2016, 44, 1434–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matic, I.; Matthews, B.G.; Wang, X.; Dyment, N.A.; Worthley, D.L.; Rowe, D.W.; Grcevic, D.; Kalajzic, I. Quiescent Bone Lining Cells Are a Major Source of Osteoblasts During Adulthood. Stem Cells 2016, 34, 2930–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riesner, K.; Shi, Y.; Jacobi, A.; Kräter, M.; Kalupa, M.; McGearey, A.; Mertlitz, S.; Cordes, S.; Schrezenmeier, J.F.; Mengwasser, J.; et al. Initiation of acute graft-versus-host disease by angiogenesis. Blood 2017, 129, 2021–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medinger, M.; Tichelli, A.; Bucher, C.; Halter, J.; Dirnhofer, S.; Rovo, A.; Passweg, J.; Tzankov, A. GVHD after allogeneic haematopoietic SCT for AML: Angiogenesis, vascular endothelial growth factor and VEGF receptor expression in the BM. Bone Marrow Transplant. 2013, 48, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Penack, O.; Henke, E.; Suh, D.; King, C.G.; Smith, O.M.; Na, I.K.; Holland, A.M.; Ghosh, A.; Lu, S.X.; Jenq, R.R.; et al. Inhibition of neovascularization to simultaneously ameliorate graft-vs-host disease and decrease tumor growth. J. Natl. Cancer Inst. 2010, 102, 894–908. [Google Scholar] [CrossRef]
- Penack, O.; Socié, G.; Van Den Brink, M.R.M. The importance of neovascularization and its inhibition for allogeneic hematopoietic stem cell transplantation. Blood 2011, 117, 4181–4189. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, E.F. Cellular mechanisms of bone remodeling. Rev. Endocr. Metab. Disord. 2010, 11, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Brandi, M.L.; Collin-Osdoby, P. Vascular biology and the skeleton. J. Bone Miner. Res. 2006, 21, 183–192. [Google Scholar] [CrossRef]
- Eghbali-Fatourechi, G.Z.; Lamsam, J.; Fraser, D.; Nagel, D.; Riggs, B.L.; Khosla, S. Circulating osteoblast-lineage cells in humans. N. Engl. J. Med. 2005, 352, 1959–1966. [Google Scholar] [CrossRef]
- Filipowska, J.; Tomaszewski, K.A.; Niedźwiedzki, Ł.; Walocha, J.A.; Niedźwiedzki, T. The role of vasculature in bone development, regeneration and proper systemic functioning. Angiogenesis 2017, 20, 291–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengwasser, J.; Babes, L.; Cordes, S.; Mertlitz, S.; Riesner, K.; Shi, Y.; McGearey, A.; Kalupa, M.; Reinheckel, T.; Penack, O. Cathepsin E deficiency ameliorates graft-versus-host disease and modifies dendritic cell motility. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertlitz, S.; Shi, Y.; Kalupa, M.; Grötzinger, C.; Mengwasser, J.; Riesner, K.; Cordes, S.; Elezkurtaj, S.; Penack, O. Lymphangiogenesis is a feature of acute GVHD, and VEGFR-3 inhibition protects against experimental GVHD. Blood 2017, 129, 1865–1875. [Google Scholar] [CrossRef] [PubMed]
- Riesner, K.; Kalupa, M.; Shi, Y.; Elezkurtaj, S.; Penack, O. A preclinical acute GVHD mouse model based on chemotherapy conditioning and MHC-matched transplantation. Bone Marrow Transplant. 2016, 51, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Cooke, K.R.; Kobzik, L.; Martin, T.R.; Brewer, J.; Delmonte, J.; Crawford, J.M.; Ferrara, J.L.M. An experimental model of idiopathic pneumonia syndrome after bone marrow transplantation: 1. The roles of minor H antigens and endotoxin. Blood 1996, 88, 3230–3239. [Google Scholar] [CrossRef] [Green Version]
- Anthony, B.A.; Hadley, G.A. Induction of graft-versus-host disease and in vivo T Cell monitoring using an MHC-matched murine model. J. Vis. Exp. 2012, 66, e3697. [Google Scholar] [CrossRef] [Green Version]
- Kawamoto, T. Use of a new adhesive film for the preparation of multi-purpose fresh-frozen sections from hard tissues, whole-animals, insects and plants. Arch. Histol. Cytol. 2003, 66, 123–143. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarz, C.S.; Bucher, C.H.; Schlundt, C.; Mertlitz, S.; Riesner, K.; Kalupa, M.; Verlaat, L.; Schmidt-Bleek, O.; Sass, R.A.; Schmidt-Bleek, K.; et al. Spatio-Temporal Bone Remodeling after Hematopoietic Stem Cell Transplantation. Int. J. Mol. Sci. 2021, 22, 267. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010267
Schwarz CS, Bucher CH, Schlundt C, Mertlitz S, Riesner K, Kalupa M, Verlaat L, Schmidt-Bleek O, Sass RA, Schmidt-Bleek K, et al. Spatio-Temporal Bone Remodeling after Hematopoietic Stem Cell Transplantation. International Journal of Molecular Sciences. 2021; 22(1):267. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010267
Chicago/Turabian StyleSchwarz, Constanze S., Christian H. Bucher, Claudia Schlundt, Sarah Mertlitz, Katarina Riesner, Martina Kalupa, Lydia Verlaat, Oskar Schmidt-Bleek, Radost A. Sass, Katharina Schmidt-Bleek, and et al. 2021. "Spatio-Temporal Bone Remodeling after Hematopoietic Stem Cell Transplantation" International Journal of Molecular Sciences 22, no. 1: 267. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010267