Morphological and Genetic Diversities of Habenaria radiata (Orchidaceae) in the Kinki Area, Japan


Abstract
:1. Introduction
2. Results
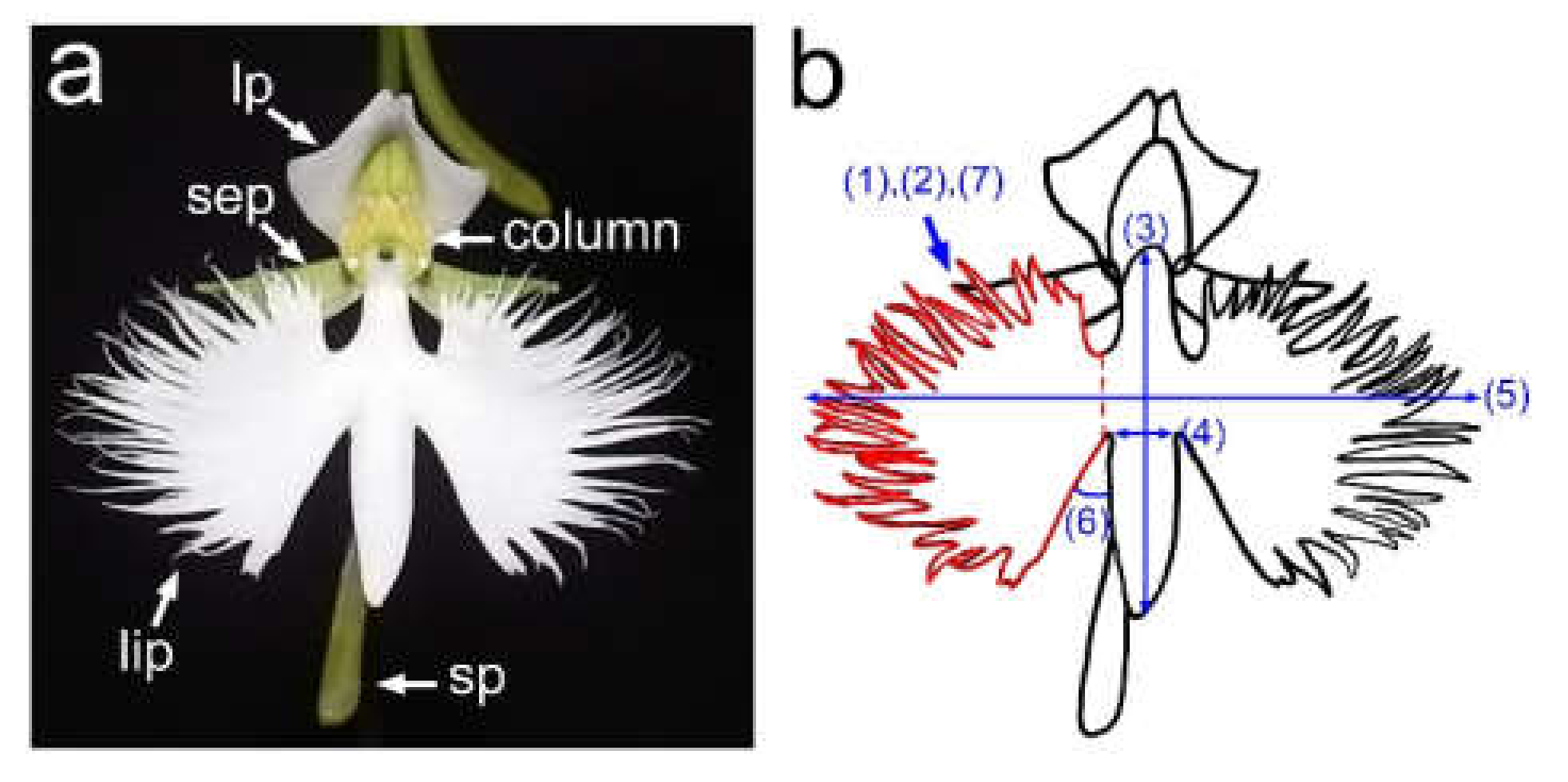
2.1. Quantification of Lip Shape
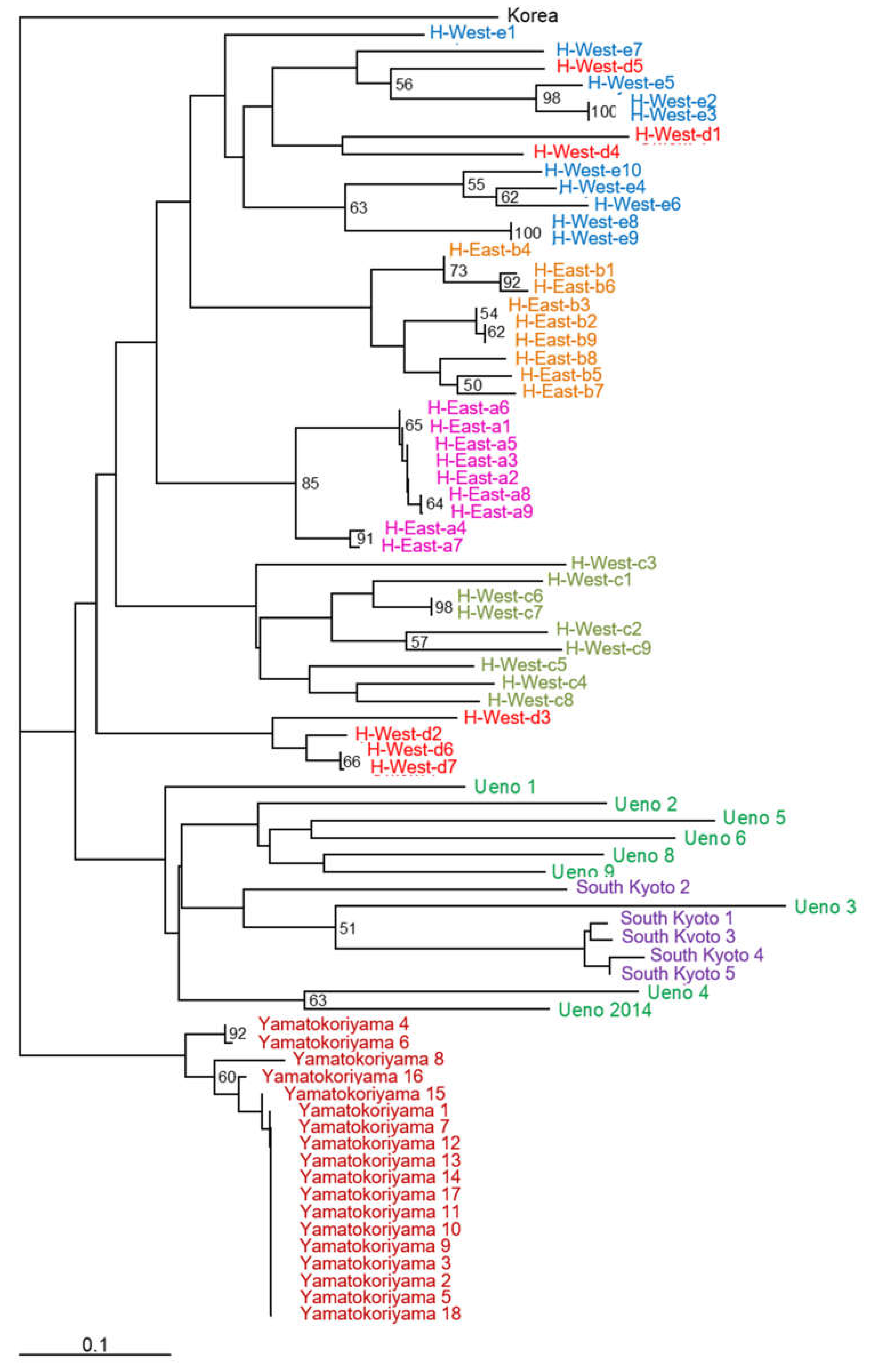
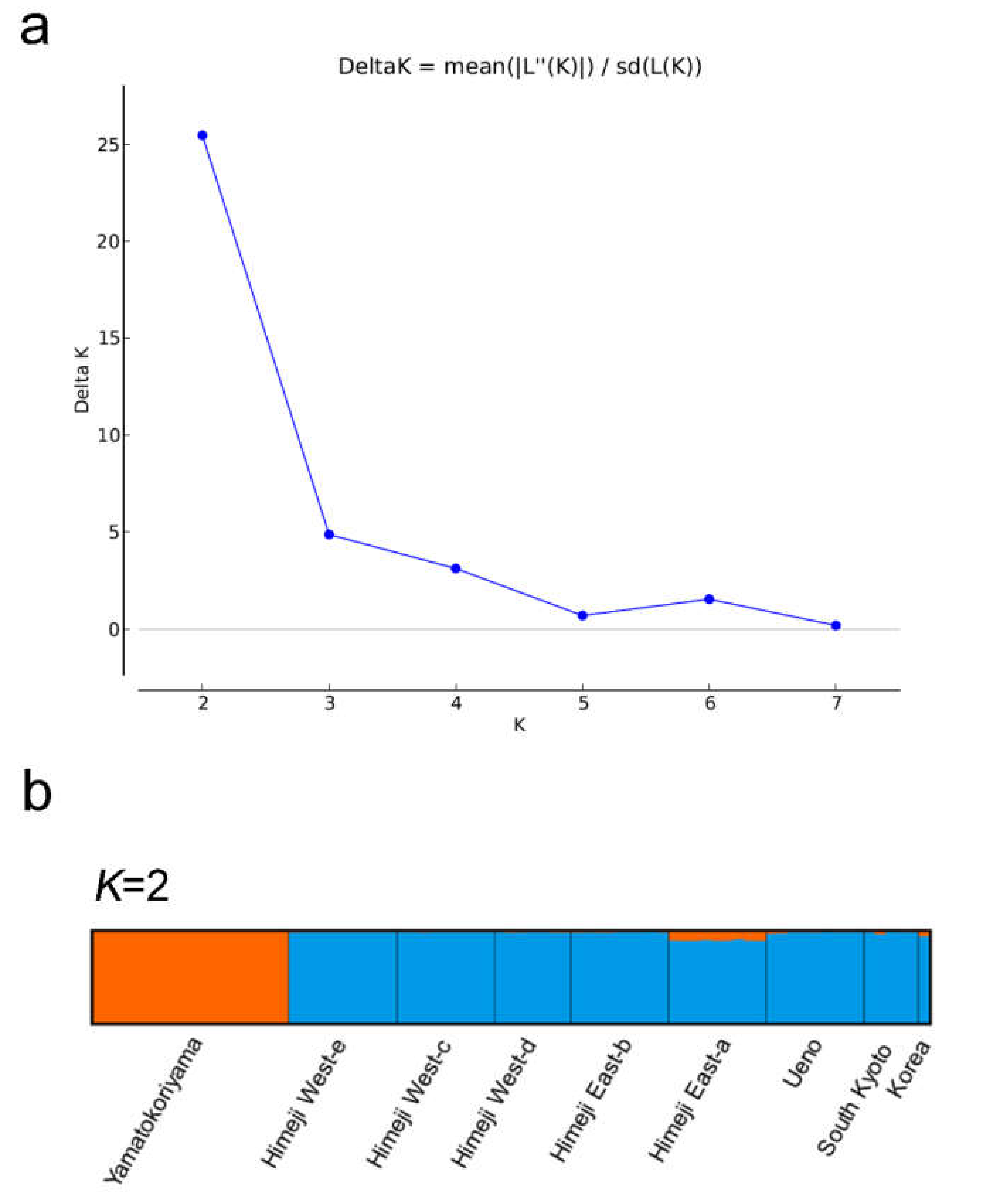
2.2. Phylogenetic and Genetic Analyses
2.3. Hawkmoth Hovering during Sucking of Nectar from H. Radiata Flowers
3. Discussion
4. Materials and Methods
4.1. H. radiata Populations and Growth Conditions
4.2. Quantification of Lip Morphology
4.3. DNA Extraction, Generation of SSR Markers, and PCR Amplification
4.4. Phylogeny and Genetic Diversity
4.5. Pollinator Recordings and Imaging Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | Linear dichroism |
References
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Batista, J.A.; Borges, K.S.; de Faria, M.W.; Proite, K.; Ramalho, A.J.; Salazar, G.A.; van den Berg, C. Molecular phylogenetics of the species-rich genus Habenaria (Orchidaceae) in the New World based on nuclear and plastid DNA sequences. Mol. Phylogenet. Evol. 2013, 67, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Leitch, I.J.; Kahandawala, I.; Suda, J.; Hanson, L.; Ingrouille, M.J.; Chase, M.W.; Fay, M.F. Genome size diversity in orchids: Consequences and evolution. Ann. Bot. 2009, 104, 469–481. [Google Scholar] [CrossRef]
- Suetsugu, K.; Tanaka, K. Diurnal butterfly pollination in the orchid Habenaria radiata. Entomol. Sci. 2014, 17, 443–445. [Google Scholar] [CrossRef]
- Shigeta, K.; Suetsugu, K. Contribution of thrips to seed production in Habenaria radiata, an orchid morphologically adapted to hawkmoths. J. Plant. Res. 2020, 133, 499–506. [Google Scholar] [CrossRef]
- Ikeuchi, Y.; Suetsugu, K.; Sumikawa, H. Diurnal skipper Pelopidas mathias (Lepidoptera: Hesperiidae) pollinates Habenaria radiata (Orchidaceae). Entomol. News 2015, 125, 7–11. [Google Scholar] [CrossRef]
- Kimura, N. Observation and Cultivation of White Egret (Sagi–Sou No Kansatsu to Saibai); New Science Co., Ltd.: Tokyo, Japan, 1982; ISBN 13-978-4821600632. (In Japanese) [Google Scholar]
- Guichoux, E.; Lagache, L.; Wagner, S.; Chaumeil, P.; Leger, P.; Lepais, O.; Lepoittevin, C.; Malausa, T.; Revardel, E.; Salin, F.; et al. Current trends in microsatellite genotyping. Mol. Ecol. Resour. 2011, 11, 591–611. [Google Scholar] [CrossRef] [PubMed]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Gustafsson, S.; Sjögren-Gulve, P. Genetic diversity in the rare orchid Gymnadenia odoratissima and a comparison with the more common congener, G. conopsea. Conserv. Genet. 2002, 3, 225–234. [Google Scholar] [CrossRef]
- Leles, B.; Chaves, A.V.; Russo, P.; Batista, J.A.; Lovato, M.B. Genetic structure is associated with phenotypic divergence in floral traits and reproductive investment in a high-altitude orchid from the Iron Quadrangle, southeastern Brazil. PLoS ONE 2015, 10, e0120645. [Google Scholar] [CrossRef] [Green Version]
- Pandey, M.; Richards, M.; Sharma, J. Microsatellite-based genetic diversity patterns in disjunct populations of a rare orchid. Genetica 2015, 143, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Giri, L.; Jugran, A.K.; Bahukhandi, A.; Dhyani, P.; Bhatt, I.D.; Rawal, R.S.; Nandi, S.K.; Dhar, U. Population genetic structure and marker trait associations using morphological, phytochemical and molecular parameters in Habenaria edgeworthii-a threatened medicinal orchid of West Himalaya, India. Appl. Biochem. Biotechnol. 2017, 181, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Cozzolino, S.; Widmer, A. Orchid diversity: An evolutionary consequence of deception? Trends Ecol. Evol. 2005, 20, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Oh, S.H.; Bhandari, G.S.; Kim, C.S.; Park, C.W. DNA barcoding of Orchidaceae in Korea. Mol. Ecol. Resour. 2014, 14, 499–507. [Google Scholar] [CrossRef]
- Nunome, T.; Negoro, S.; Miyatake, K.; Yamaguchi, H.; Fukuoka, H. A protocol for the construction of microsatellite enriched genomic library. Plant Mol. Biol. Rep. 2006, 24, 305–312. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. Genepop’007: A complete reimplementation of the Genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Langella, O. Populations 1.2.32: Population Genetic Software (Individuals or Populations Distances, Phylogenetic Trees). 2011. Available online: http://www.bioinformatics.org/project/?group_id=84 (accessed on 30 September 2020).
- Page, R. TREEVIEW: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. CLUMPAK: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | N | He | Ho | F |
|---|---|---|---|---|
| Himeji East-a | 9 | 0.2583 | 0.4409 | −0.7067 * |
| Himeji East-b | 9 | 0.2695 | 0.3187 | −0.1825 |
| Himeji West-c | 9 | 0.4103 | 0.3533 | 0.1390 |
| Himeji West-d | 7 | 0.4445 | 0.5180 | −0.1654 |
| Himeji West-e | 10 | 0.4380 | 0.4203 | 0.0404 |
| Ueno | 9 | 0.6410 | 0.5318 | 0.1731 * |
| South Kyoto | 5 | 0.4311 | 0.6139 | −0.4239 * |
| Yamatokoriyama | 18 | 0.2362 | 0.4441 | −0.8803 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tachibana, T.; Nishikawa, Y.; Kubo, N.; Takeda, S. Morphological and Genetic Diversities of Habenaria radiata (Orchidaceae) in the Kinki Area, Japan. Int. J. Mol. Sci. 2021, 22, 311. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010311
Tachibana T, Nishikawa Y, Kubo N, Takeda S. Morphological and Genetic Diversities of Habenaria radiata (Orchidaceae) in the Kinki Area, Japan. International Journal of Molecular Sciences. 2021; 22(1):311. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010311
Chicago/Turabian StyleTachibana, Tsutomu, Yuki Nishikawa, Nakao Kubo, and Seiji Takeda. 2021. "Morphological and Genetic Diversities of Habenaria radiata (Orchidaceae) in the Kinki Area, Japan" International Journal of Molecular Sciences 22, no. 1: 311. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010311