Pick’s Tau Fibril Shows Multiple Distinct PET Probe Binding Sites: Insights from Computational Modelling


Abstract
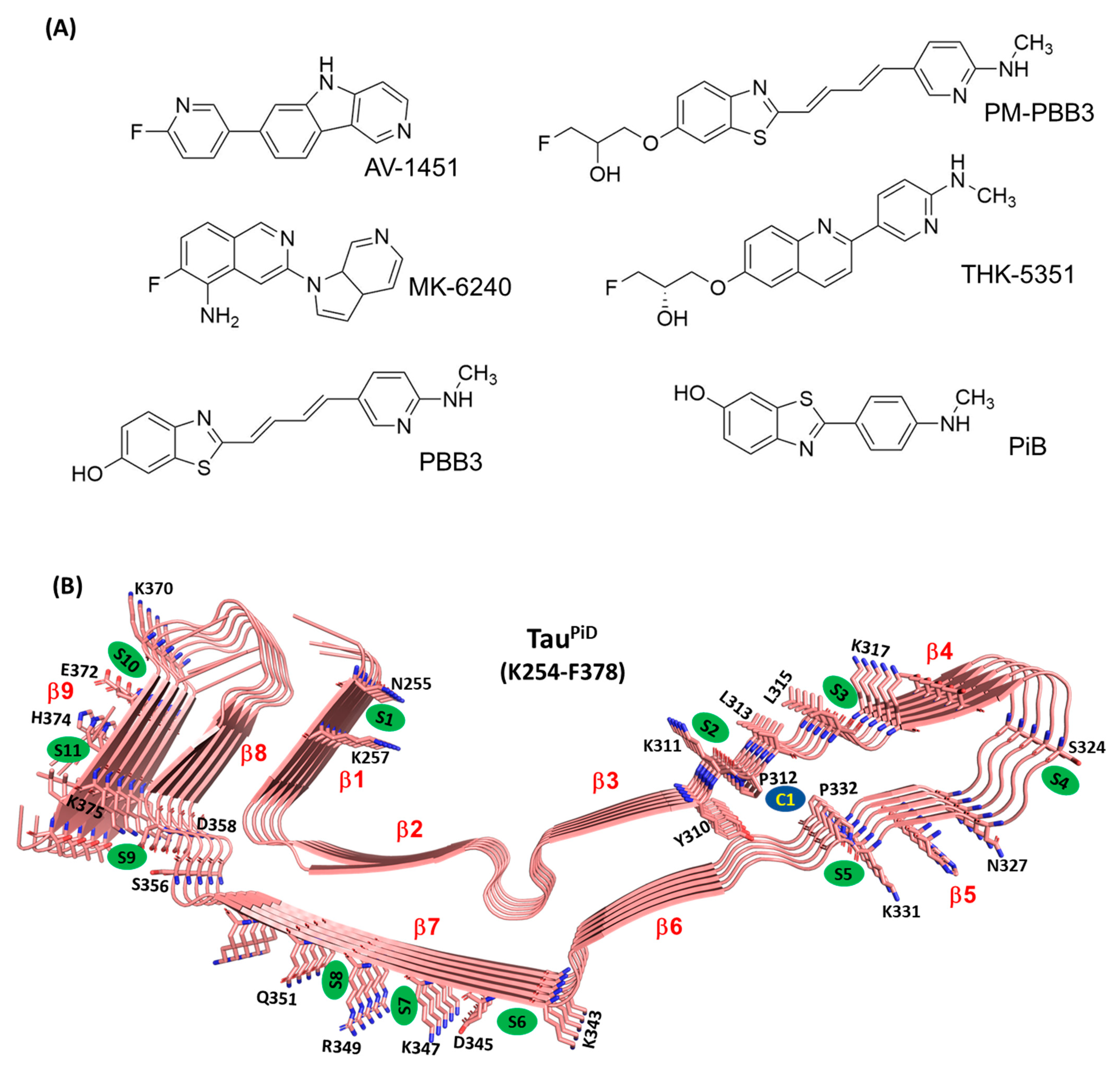
:1. Introduction
2. Results and Discussion
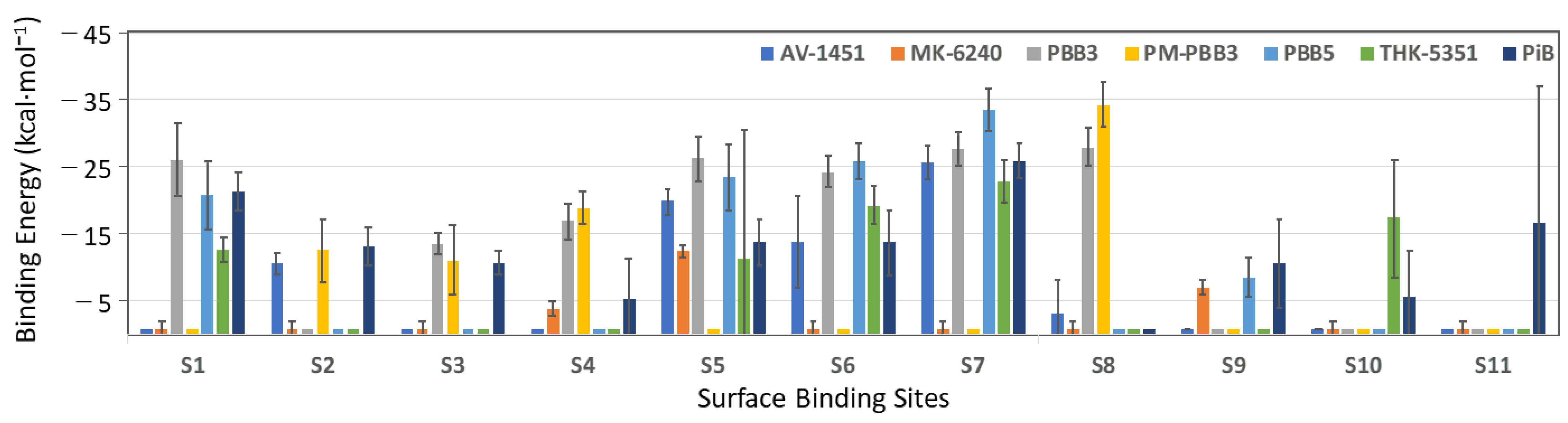
2.1. Assessing Binding Regions and Affinity of PET Tracers to TauPiD by Docking
2.2. Molecular Dynamics and MM/GBSA Calculations
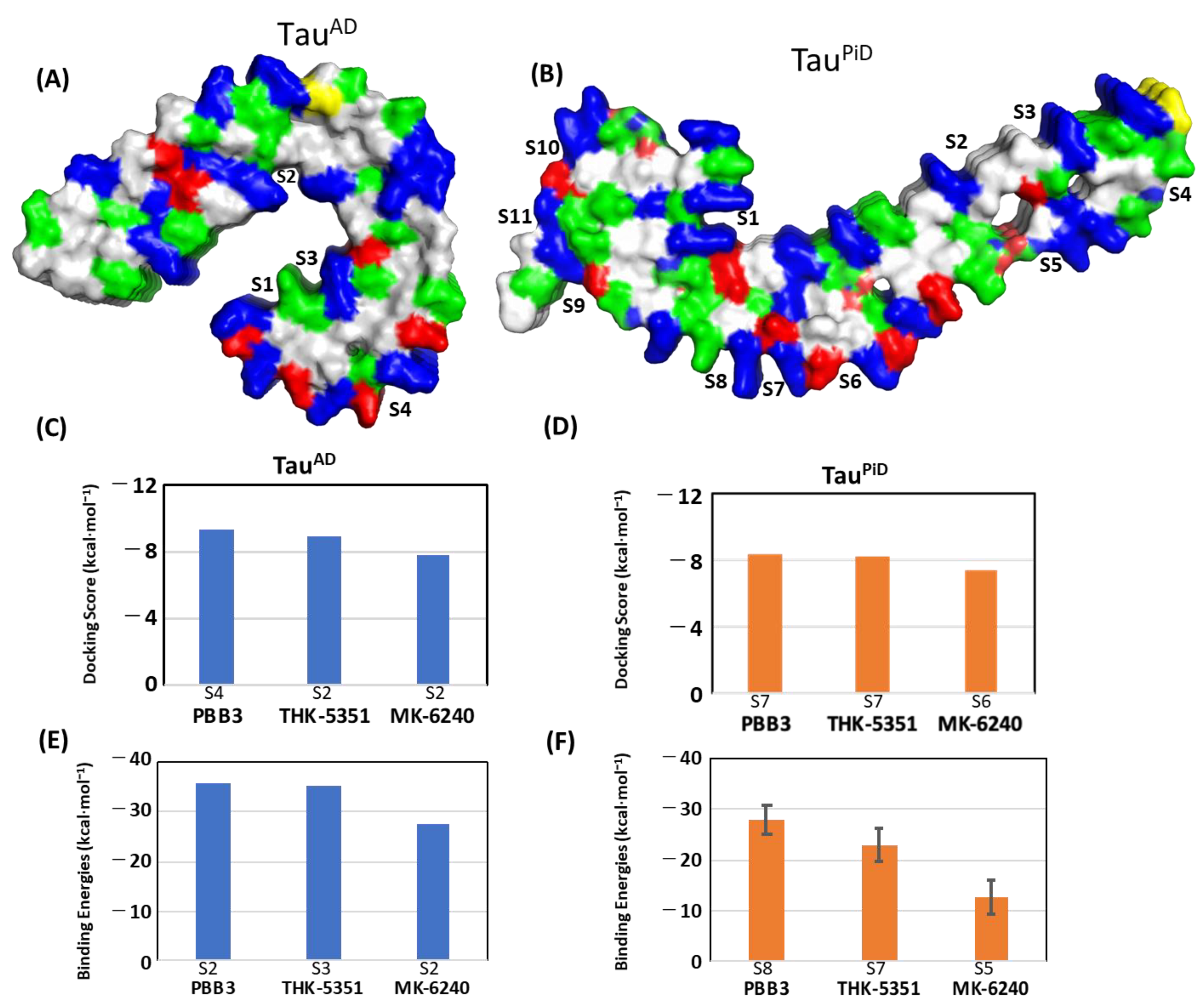
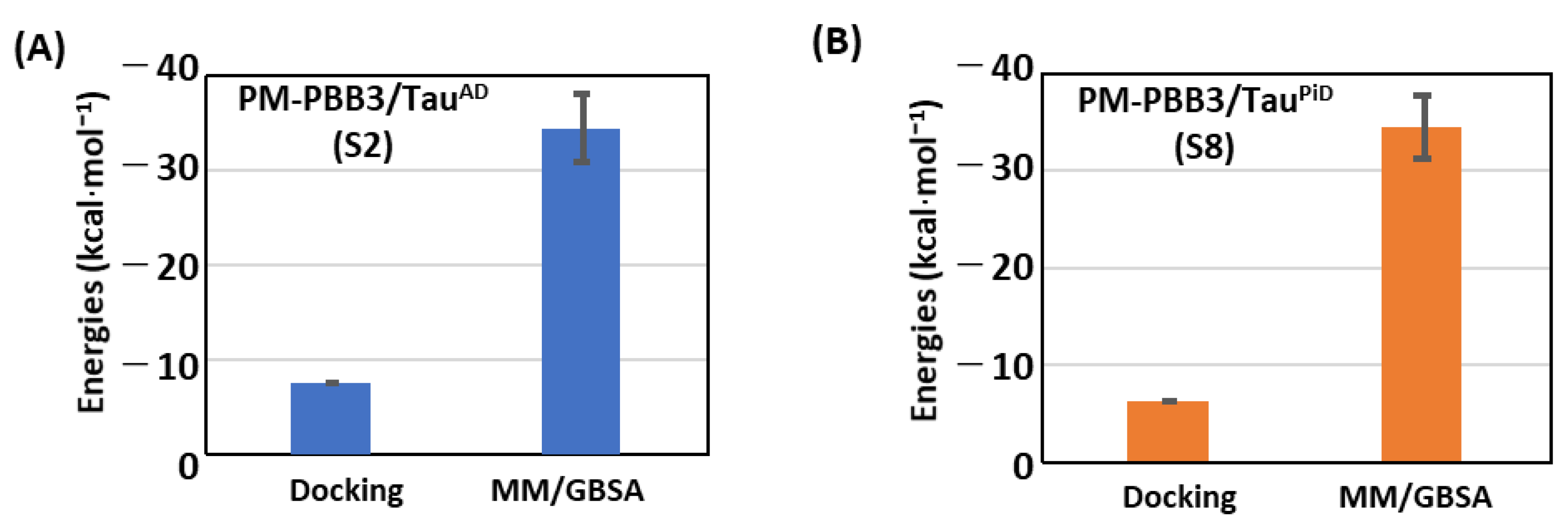
2.3. Comparison of Probe Binding Affinities between TauAD and TauPiD
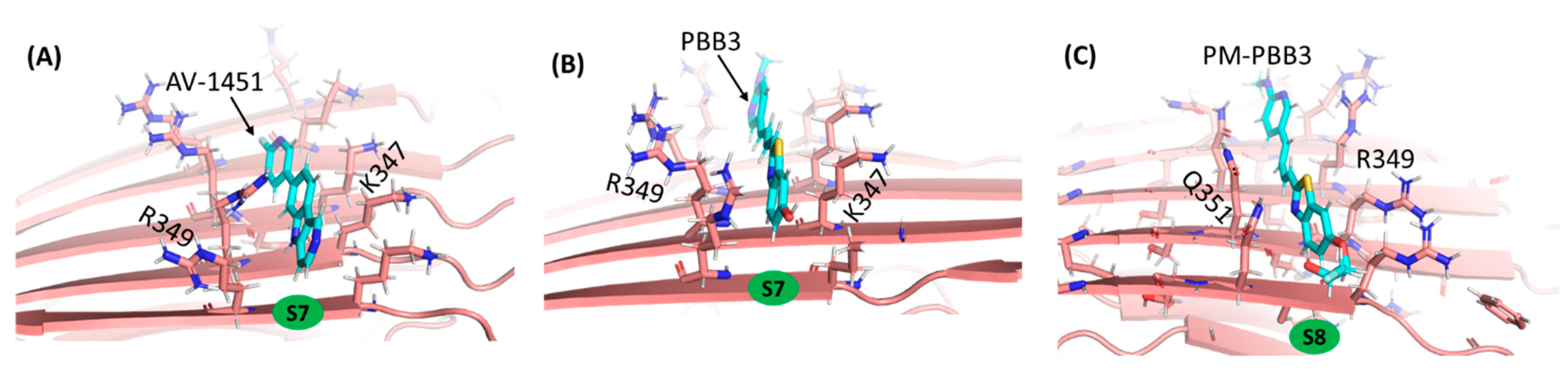
2.4. PM-PBB3 as a Potent PET Tracer for Detecting TauPiD
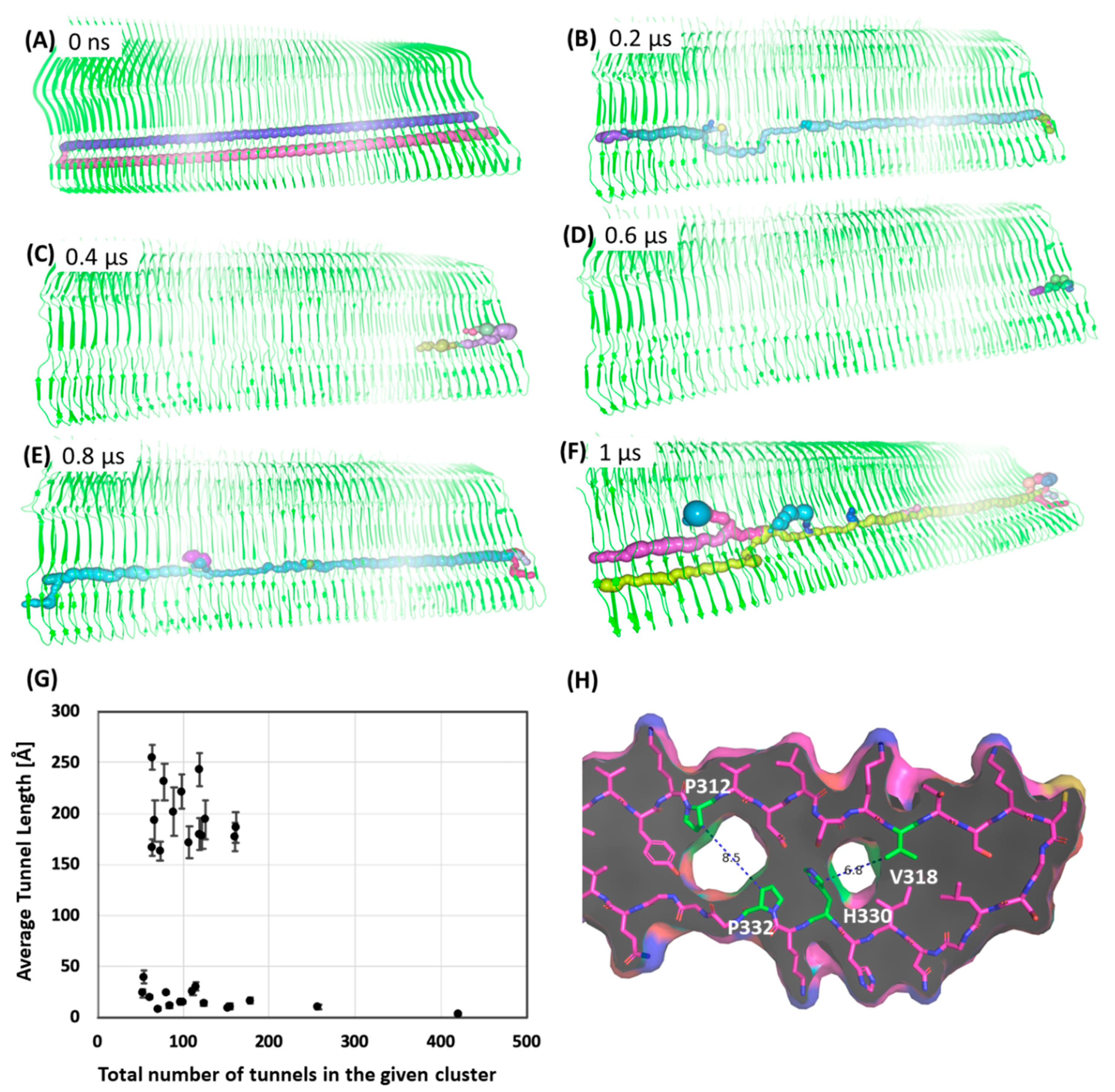
2.5. Does Pick’s Tau have a Potential Cavity Binding Site?
3. Materials and Methods
3.1. Structure Preparation
3.2. Docking
3.3. MD Simulation
3.4. MM/GBSA Calculations
3.5. Tunnel Calculations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, V.M.; Goedert, M.; Trojanowski, J.Q. Neurodegenerative Tauopathies. Annu. Rev. Neurosci. 2001, 24, 1121–1159. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, S.; Avila, J.; Schöll, M.; Kovacs, G.G.; Kövari, E.; Skrabana, R.; Evans, L.D.; Kontsekova, E.; Malawska, B.; de Silva, R.; et al. A Walk through Tau Therapeutic Strategies. Acta Neuropathol. Commun. 2019, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Eisenberg, D.S.; Crowther, R.A. Propagation of Tau Aggregates and Neurodegeneration. Annu. Rev. Neurosci. 2017, 40, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, A.W.P.; Falcon, B.; He, S.; Murzin, A.G.; Murshudov, G.; Garringer, H.J.; Crowther, R.A.; Ghetti, B.; Goedert, M.; Scheres, S.H.W. Cryo-EM Structures of Tau Filaments from Alzheimer’s Disease. Nature 2017, 547, 185–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcon, B.; Zivanov, J.; Zhang, W.; Murzin, A.G.; Garringer, H.J.; Vidal, R.; Crowther, R.A.; Newell, K.L.; Ghetti, B.; Goedert, M.; et al. Novel Tau Filament Fold in Chronic Traumatic Encephalopathy Encloses Hydrophobic Molecules. Nature 2019, 568, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tarutani, A.; Newell, K.L.; Murzin, A.G.; Matsubara, T.; Falcon, B.; Vidal, R.; Garringer, H.J.; Shi, Y.; Ikeuchi, T.; et al. Novel Tau Filament Fold in Corticobasal Degeneration. Nature 2020, 580, 283–287. [Google Scholar] [CrossRef]
- Falcon, B.; Zhang, W.; Schweighauser, M.; Murzin, A.G.; Vidal, R.; Garringer, H.J.; Ghetti, B.; Scheres, S.H.W.; Goedert, M. Tau Filaments from Multiple Cases of Sporadic and Inherited Alzheimer’s Disease Adopt a Common Fold. Acta Neuropathol. 2018, 136, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Falcon, B.; Zhang, W.; Murzin, A.G.; Murshudov, G.; Garringer, H.J.; Vidal, R.; Crowther, R.A.; Ghetti, B.; Scheres, S.H.W.; Goedert, M. Structures of Filaments from Pick’s Disease Reveal a Novel Tau Protein Fold. Nature 2018, 561, 137–140. [Google Scholar] [CrossRef]
- Maruyama, M.; Shimada, H.; Suhara, T.; Shinotoh, H.; Ji, B.; Maeda, J.; Zhang, M.-R.; Trojanowski, J.Q.; Lee, V.M.-Y.; Ono, M.; et al. Imaging of Tau Pathology in a Tauopathy Mouse Model and in Alzheimer Patients Compared to Normal Controls. Neuron 2013, 79, 1094–1108. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.-F.; Arteaga, J.; Chen, G.; Gangadharmath, U.; Gomez, L.F.; Kasi, D.; Lam, C.; Liang, Q.; Liu, C.; Mocharla, V.P.; et al. [18F]T807, a Novel Tau Positron Emission Tomography Imaging Agent for Alzheimer’s Disease. Alzheimers Dement. 2013, 9, 666–676. [Google Scholar] [CrossRef]
- Hostetler, E.D.; Walji, A.M.; Zeng, Z.; Miller, P.; Bennacef, I.; Salinas, C.; Connolly, B.; Gantert, L.; Haley, H.; Holahan, M.; et al. Preclinical Characterization of 18F-MK-6240, a Promising PET Tracer for In Vivo Quantification of Human Neurofibrillary Tangles. J. Nucl. Med. 2016, 57, 1599–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walji, A.M.; Hostetler, E.D.; Selnick, H.; Zeng, Z.; Miller, P.; Bennacef, I.; Salinas, C.; Connolly, B.; Gantert, L.; Holahan, M.; et al. Discovery of 6-(Fluoro-18F)-3-(1H-Pyrrolo[2,3-c]Pyridin-1-Yl)Isoquinolin-5-Amine ([18F]-MK-6240): A Positron Emission Tomography (PET) Imaging Agent for Quantification of Neurofibrillary Tangles (NFTs). J. Med. Chem. 2016, 59, 4778–4789. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.; Mak, E.; Cervenka, S.; Aigbirhio, F.I.; Rowe, J.B.; O’Brien, J.T. In Vivo Tau PET Imaging in Dementia: Pathophysiology, Radiotracer Quantification, and a Systematic Review of Clinical Findings. Ageing Res. Rev. 2017, 36, 50–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, A.; Kroth, H.; Schieferstein, H.; Berndt, M.; Oden, F.; Capotosti, F.; Molette, J.; Juergens, T.; Darmency, V.; Schmitt-Willich, H.; et al. Preclinical Characterization of PI-2620, A Novel Tau PET Tracer for Detection of Tau in AD and Other Tauopathies. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2017, 13, P141–P142. [Google Scholar] [CrossRef]
- Saint-Aubert, L.; Lemoine, L.; Chiotis, K.; Leuzy, A.; Rodriguez-Vieitez, E.; Nordberg, A. Tau PET Imaging: Present and Future Directions. Mol. Neurodegener. 2017, 12, 19. [Google Scholar] [CrossRef] [Green Version]
- Tagai, K.; Ono, M.; Kubota, M.; Kitamura, S.; Takahata, K.; Seki, C.; Takado, Y.; Shinotoh, H.; Sano, Y.; Yamamoto, Y.; et al. High-Contrast In Vivo Imaging of Tau Pathologies in Alzheimer’s and Non-Alzheimer’s Disease Tauopathies. Neuron 2020. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Diagnostic Criteria for Neuropathologic Assessment of Alzheimer’s Disease. Neurobiol. Aging 1997, 18, S85–S88. [Google Scholar] [CrossRef]
- Lowe, V.J.; Curran, G.; Fang, P.; Liesinger, A.M.; Josephs, K.A.; Parisi, J.E.; Kantarci, K.; Boeve, B.F.; Pandey, M.K.; Bruinsma, T.; et al. An Autoradiographic Evaluation of AV-1451 Tau PET in Dementia. Acta Neuropathol. Commun. 2016, 4, 58. [Google Scholar] [CrossRef] [Green Version]
- Ng, K.P.; Pascoal, T.A.; Mathotaarachchi, S.; Therriault, J.; Kang, M.S.; Shin, M.; Guiot, M.-C.; Guo, Q.; Harada, R.; Comley, R.A.; et al. Monoamine Oxidase B Inhibitor, Selegiline, Reduces 18F-THK5351 Uptake in the Human Brain. Alzheimers Res. 2017, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Harada, R.; Ishiki, A.; Kai, H.; Sato, N.; Furukawa, K.; Furumoto, S.; Tago, T.; Tomita, N.; Watanuki, S.; Hiraoka, K.; et al. Correlations of 18F-THK5351 PET with Postmortem Burden of Tau and Astrogliosis in Alzheimer Disease. J. Nucl. Med. 2018, 59, 671–674. [Google Scholar] [CrossRef] [Green Version]
- Ishiki, A.; Harada, R.; Kai, H.; Sato, N.; Totsune, T.; Tomita, N.; Watanuki, S.; Hiraoka, K.; Ishikawa, Y.; Funaki, Y.; et al. Neuroimaging-Pathological Correlations of [18F]THK5351 PET in Progressive Supranuclear Palsy. Acta Neuropathol. Commun. 2018, 6, 53. [Google Scholar] [CrossRef]
- Ishibashi, K.; Kameyama, M.; Tago, T.; Toyohara, J.; Ishii, K. Potential Use of 18F-THK5351 PET to Identify Wallerian Degeneration of the Pyramidal Tract Caused by Cerebral Infarction. Clin. Nucl. Med. 2017, 42, e523–e524. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, L.C.; Knust, H.; Körner, M.; Honer, M.; Czech, C.; Belli, S.; Muri, D.; Edelmann, M.R.; Hartung, T.; Erbsmehl, I.; et al. Identification of Three Novel Radiotracers for Imaging Aggregated Tau in Alzheimer’s Disease with Positron Emission Tomography. J. Med. Chem. 2017, 60, 7350–7370. [Google Scholar] [CrossRef] [PubMed]
- Kroth, H.; Oden, F.; Molette, J.; Schieferstein, H.; Capotosti, F.; Mueller, A.; Berndt, M.; Schmitt-Willich, H.; Darmency, V.; Gabellieri, E.; et al. Discovery and Preclinical Characterization of [18F]PI-2620, a next-Generation Tau PET Tracer for the Assessment of Tau Pathology in Alzheimer’s Disease and Other Tauopathies. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2178–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugan, N.A.; Nordberg, A.; Ågren, H. Different Positron Emission Tomography Tau Tracers Bind to Multiple Binding Sites on the Tau Fibril: Insight from Computational Modeling. ACS Chem. Neurosci. 2018, 9, 1757–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goedert, M.; Yamaguchi, Y.; Mishra, S.K.; Higuchi, M.; Sahara, N. Tau Filaments and the Development of Positron Emission Tomography Tracers. Front. Neurol. 2018, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Chien, D.T.; Bahri, S.; Szardenings, A.K.; Walsh, J.C.; Mu, F.; Su, M.-Y.; Shankle, W.R.; Elizarov, A.; Kolb, H.C. Early Clinical PET Imaging Results with the Novel PHF-Tau Radioligand [F-18]-T807. J. Alzheimers Dis. 2013, 34, 457–468. [Google Scholar] [CrossRef] [Green Version]
- Betthauser, T.J.; Hillmer, A.T.; Lao, P.J.; Ehlerding, E.; Mukherjee, J.; Stone, C.K.; Christian, B.T. Human Biodistribution and Dosimetry of [18F]Nifene, an A4β2* Nicotinic Acetylcholine Receptor PET Tracer. Nucl. Med. Biol. 2017, 55, 7–11. [Google Scholar] [CrossRef]
- Harada, R.; Okamura, N.; Furumoto, S.; Yanai, K. Imaging Protein Misfolding in the Brain Using β-Sheet Ligands. Front. Neurosci. 2018, 12. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C. Beware of Docking! Trends Pharm. Sci. 2015, 36, 78–95. [Google Scholar] [CrossRef]
- Ono, M.; Sahara, N.; Kumata, K.; Ji, B.; Ni, R.; Koga, S.; Dickson, D.W.; Trojanowski, J.Q.; Lee, V.M.-Y.; Yoshida, M.; et al. Distinct Binding of PET Ligands PBB3 and AV-1451 to Tau Fibril Strains in Neurodegenerative Tauopathies. Brain 2017, 140, 764–780. [Google Scholar] [CrossRef] [Green Version]
- Hudson, K.L.; Bartlett, G.J.; Diehl, R.C.; Agirre, J.; Gallagher, T.; Kiessling, L.L.; Woolfson, D.N. Carbohydrate-Aromatic Interactions in Proteins. J. Am. Chem. Soc. 2015, 137, 15152–15160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimmerova, M.; Kozmon, S.; Nečasová, I.; Mishra, S.K.; Komárek, J.; Koca, J. Stacking Interactions between Carbohydrate and Protein Quantified by Combination of Theoretical and Experimental Methods. PLoS ONE 2012, 7, e46032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houser, J.; Kozmon, S.; Mishra, D.; Mishra, S.K.; Romano, P.R.; Wimmerová, M.; Koča, J. Influence of Trp Flipping on Carbohydrate Binding in Lectins. An Example on Aleuria Aurantia Lectin AAL. PLoS ONE 2017, 12, e0189375. [Google Scholar] [CrossRef] [Green Version]
- Ni, R.; Ji, B.; Ono, M.; Sahara, N.; Zhang, M.-R.; Aoki, I.; Nordberg, A.; Suhara, T.; Higuchi, M. Comparative In Vitro and In Vivo Quantifications of Pathologic Tau Deposits and Their Association with Neurodegeneration in Tauopathy Mouse Models. J. Nucl. Med. 2018, 59, 960–966. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Takahashi, M.; Shimozawa, A.; Fujinaga, M.; Mori, W.; Nagai, Y.; Mimura, K.; Minamihisamatsu, T.; Uchida, S.; Shimojo, M.; et al. In Vivo Visualization of Propagating α-Synuclein Pathologies in Mouse and Marmoset Models by a Bimodal Imaging Probe, C05-05. bioRxiv 2020. [Google Scholar] [CrossRef]
- Maestro; Schrödinger, LLC: New York, NY, USA, 2018.
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian˜16 Revision B.01; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Case, D.A.; Belfon, K.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E., III; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Giambasu, G.; et al. AMBER 18; University of California: San Fransisco, CA, USA, 2018. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Lai, L.; Wang, S. Further Development and Validation of Empirical Scoring Functions for Structure-Based Binding Affinity Prediction. J. Comput. Aided Mol. Des. 2002, 16, 11–26. [Google Scholar] [CrossRef]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. Ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from Ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Nagae, M.; Mishra, S.K.; Hanashima, S.; Tateno, H.; Yamaguchi, Y. Distinct Roles for Each N-Glycan Branch Interacting with Mannose-Binding Type Jacalin-Related Lectins Orysata and Calsepa. Glycobiology 2017, 27, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Roe, D.R.; Cheatham, T.E. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Koča, J. Assessing the Performance of MM/PBSA, MM/GBSA, and QM–MM/GBSA Approaches on Protein/Carbohydrate Complexes: Effect of Implicit Solvent Models, QM Methods, and Entropic Contributions. J. Phys. Chem. B 2018, 122, 8113–8121. [Google Scholar] [CrossRef] [PubMed]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA Methods to Estimate Ligand-Binding Affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, Y. Generalized-Ensemble Algorithms: Enhanced Sampling Techniques for Monte Carlo and Molecular Dynamics Simulations. J. Mol. Graph. Model. 2004, 22, 425–439. [Google Scholar] [CrossRef] [Green Version]
- Jurcik, A.; Bednar, D.; Byska, J.; Marques, S.M.; Furmanova, K.; Daniel, L.; Kokkonen, P.; Brezovsky, J.; Strnad, O.; Stourac, J.; et al. CAVER Analyst 2.0: Analysis and Visualization of Channels and Tunnels in Protein Structures and Molecular Dynamics Trajectories. Bioinformatics 2018, 34, 3586–3588. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding Site | AV-1451 | MK-6240 | PBB3 | PM-PBB3 | THK5351 | PiB |
|---|---|---|---|---|---|---|
| C1 | −11.0 (−8.9) | −11.0 (−9.0) | −10.1 (−9.4) | −10.0 (−9.4) | −10.3 (−9.0) | −8.9 (−8.5) |
| S1 | NB | NB | −4.9 (−7.0) | NB | NB | −4.4 (−6.6) |
| S2 | −6.0 (−7.3) | NB | NB | −5.0 (−7.2) | −5.7 (−7.0) | −4.4 (−6.9) |
| S3 | NB | NB | −5.7 (−7.6) | −5.4 (−7.4) | NB | −4.6 (−7.2) |
| S4 | NB | −5.1 (−6.7) | −5.0 (−6.9) | NB | NB | −4.5 (−6.5) |
| S5 | −6.5 (−7.4) | −5.5 (−6.7) | −6.0 (−7.7) | NB | −5.8 (−7.1) | −5.2 (−7.2) |
| S6 | −8.3 (−7.9) | −7.4 (−7.7) | −7.3 (−8.0) | NB | −7.3 (−7.8) | −6.3 (−7.5) |
| S7 | −9.4 (−8.4) | NB | −8.4 (−8.7) | NB | −8.3 (−8.3) | −7.1 (−7.8) |
| S8 | −7.1 (−7.5) | NB | −6.2 (−7.6) | −6.1 (−7.5) | NB | NB |
| S9 | −5.9 (−6.7) | −5.7 (−6.9) | NB | −4.5 (−6.5) | −5.6 (−6.7) | −4.6 (−6.6) |
| S10 | −6.0 (−7.1) | NB | NB | −4.6 (−6.6) | −5.2 (−6.8) | −4.6 (−6.7) |
| S11 | NB | NB | NB | −4.5 (−6.9) | NB | −4.5 (−6.9) |
| Binding Site | AV-1451 | MK-6240 | PBB3 | PM-PBB3 | THK-5351 | PiB |
|---|---|---|---|---|---|---|
| C1 | −25.8 ± 2.8 | −32.1 ± 4.1 | −40.9 ± 2.7 | −50.0 ± 3.1 | −33.2 ± 5.1 | −27.0 ± 3.0 |
| S1 | x | x | −26.0 ± 5.5 | - | x | −21.4 ± 2.9 |
| S2 | −10.7 ± 1.6 | x | x | −12.7 ± 4.6 | −12.6 ± 1.8 | −13.1 ± 2.8 |
| S3 | x | x | −13.6 ± 1.6 | −11.2 ± 5.1 | x | −10.7 ± 1.7 |
| S4 | x | −3.9 ± 3.7 | −16.9 ± 2.6 | −18.9 ± 2.4 | x | −5.5 ± 5.7 |
| S5 | −19.9 ± 1.9 | −12.5 ± 3.3 | −26.2 ± 3.3 | x | −11.3 ± 19.3 | −13.8 ± 3.4 |
| S6 | −13.8 ± 6.9 | - | −24.3 ± 2.4 | x | −19.3 ± 2.9 | −13.7 ± 4.8 |
| S7 | −25.5 ± 2.5 | x | −27.6 ± 2.6 | x | −22.8 ± 3.2 | −25.9 ± 2.6 |
| S8 | −3.2 ± 5.0 | x | −27.9 ± 2.9 | −34.2 ± 3.3 | x | x |
| S9 | - | −7.1 ± 5.7 | x | - | −10.7 ± 6.5 | |
| S10 | - | x | x | - | −17.4 ± 8.7 | −5.6 ± 6.9 |
| S11 | x | x | x | - | x | −16.8 ± 25.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishra, S.K.; Yamaguchi, Y.; Higuchi, M.; Sahara, N. Pick’s Tau Fibril Shows Multiple Distinct PET Probe Binding Sites: Insights from Computational Modelling. Int. J. Mol. Sci. 2021, 22, 349. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010349
Mishra SK, Yamaguchi Y, Higuchi M, Sahara N. Pick’s Tau Fibril Shows Multiple Distinct PET Probe Binding Sites: Insights from Computational Modelling. International Journal of Molecular Sciences. 2021; 22(1):349. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010349
Chicago/Turabian StyleMishra, Sushil K., Yoshiki Yamaguchi, Makoto Higuchi, and Naruhiko Sahara. 2021. "Pick’s Tau Fibril Shows Multiple Distinct PET Probe Binding Sites: Insights from Computational Modelling" International Journal of Molecular Sciences 22, no. 1: 349. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010349