Tryptophan Derivatives by Saccharomyces cerevisiae EC1118: Evaluation, Optimization, and Production in a Soybean-Based Medium

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
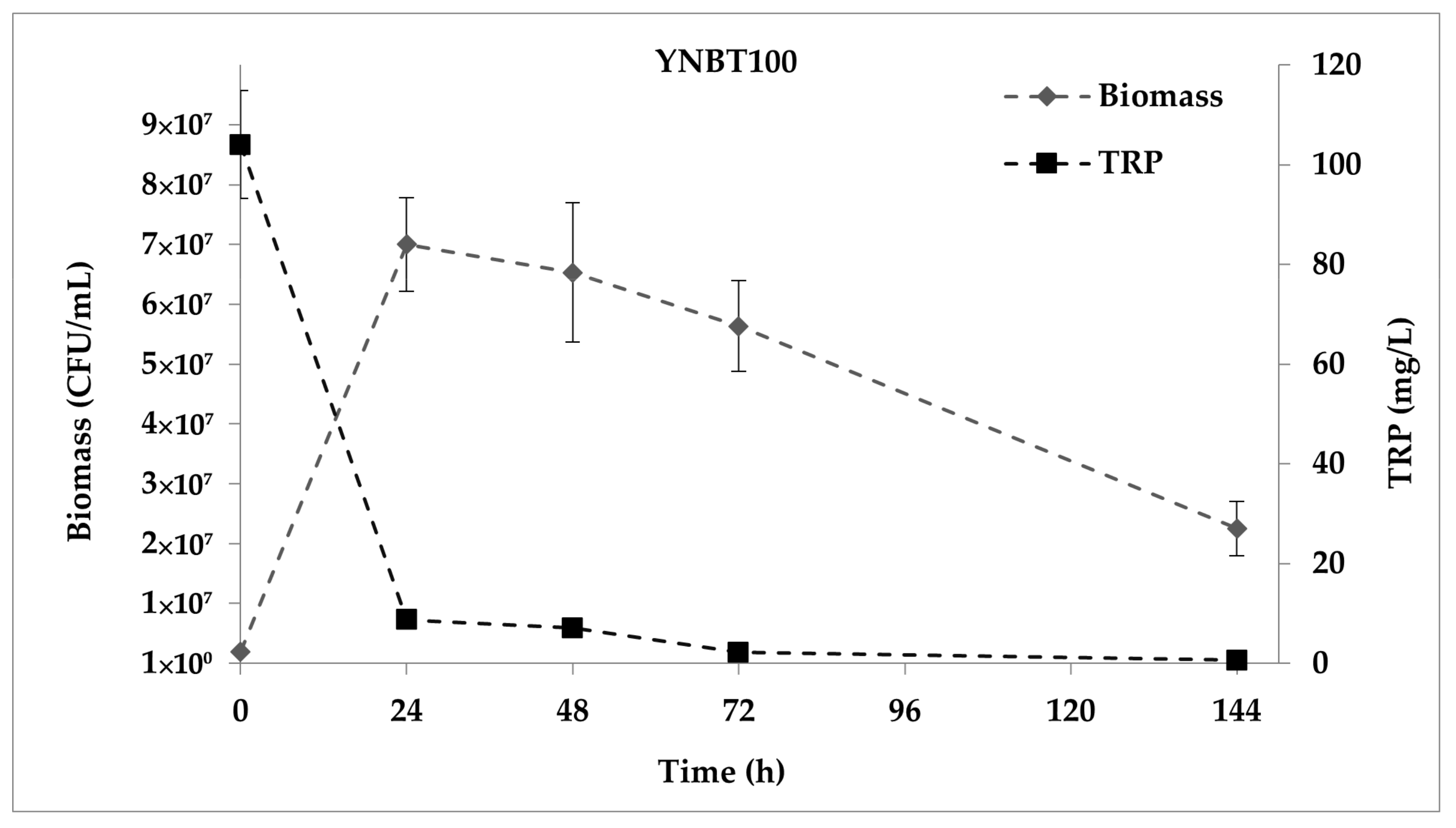
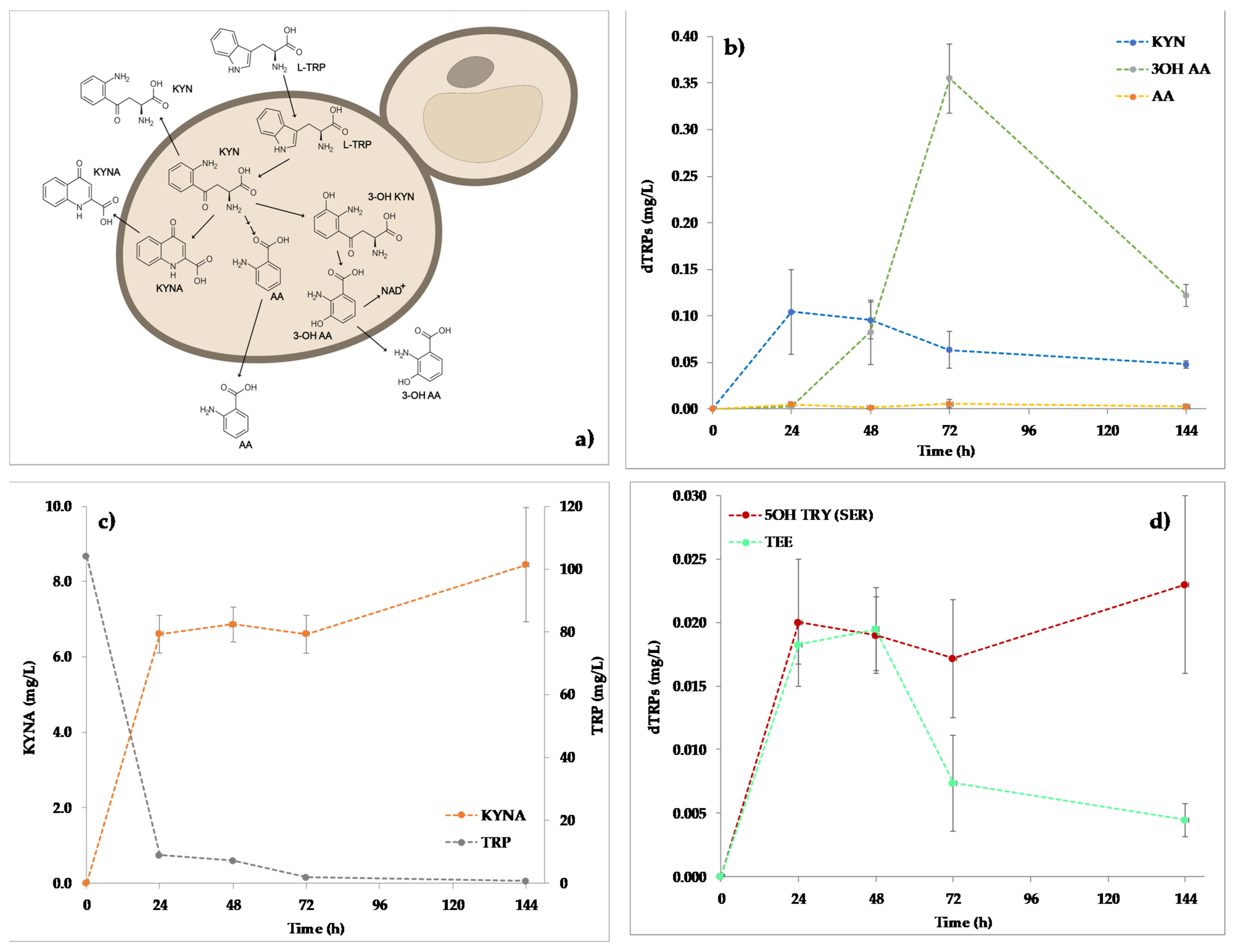
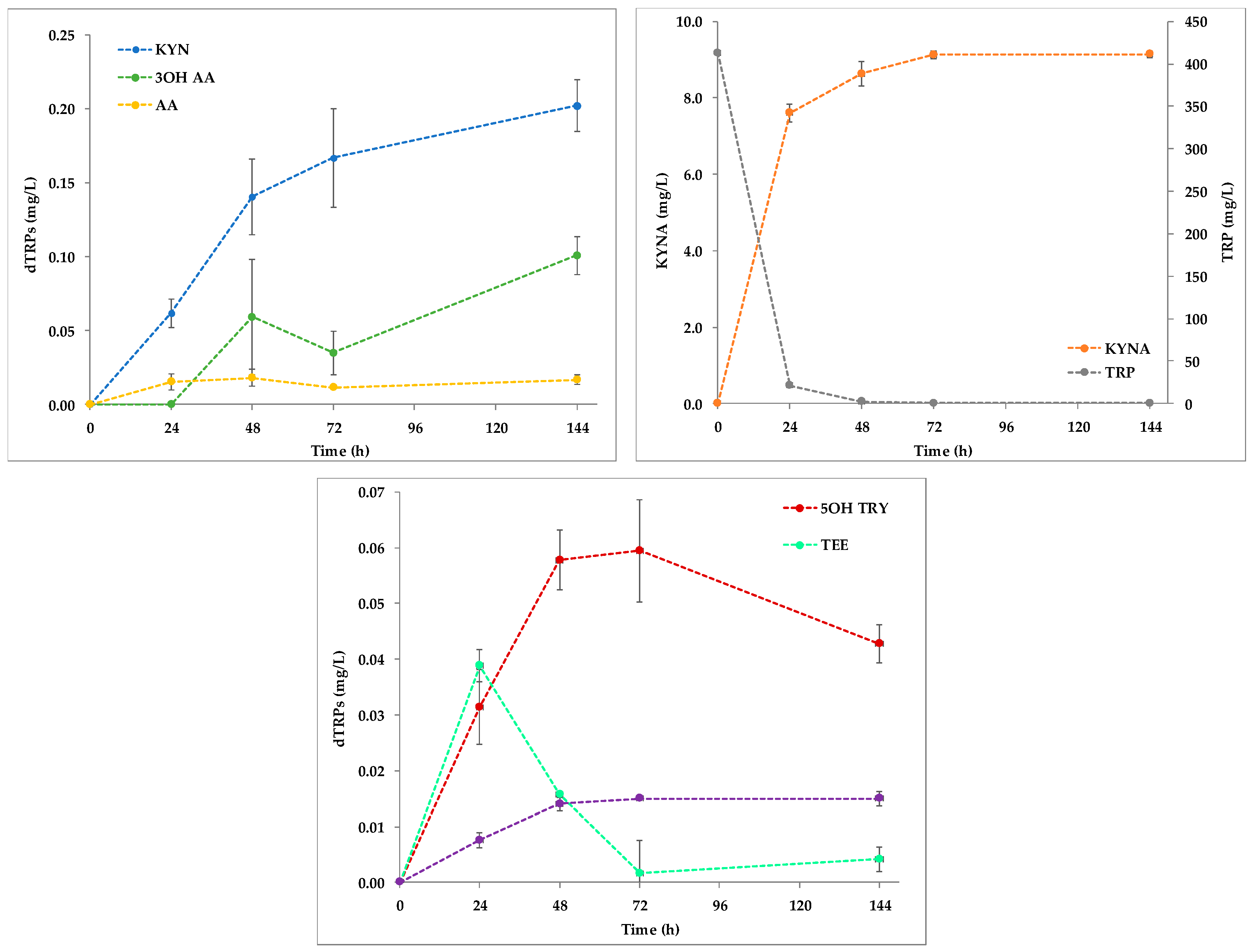
2.1. Quantitation of dTRPs during the Fermentation of S. Cerevisiae EC1118 in YNBT100 Medium
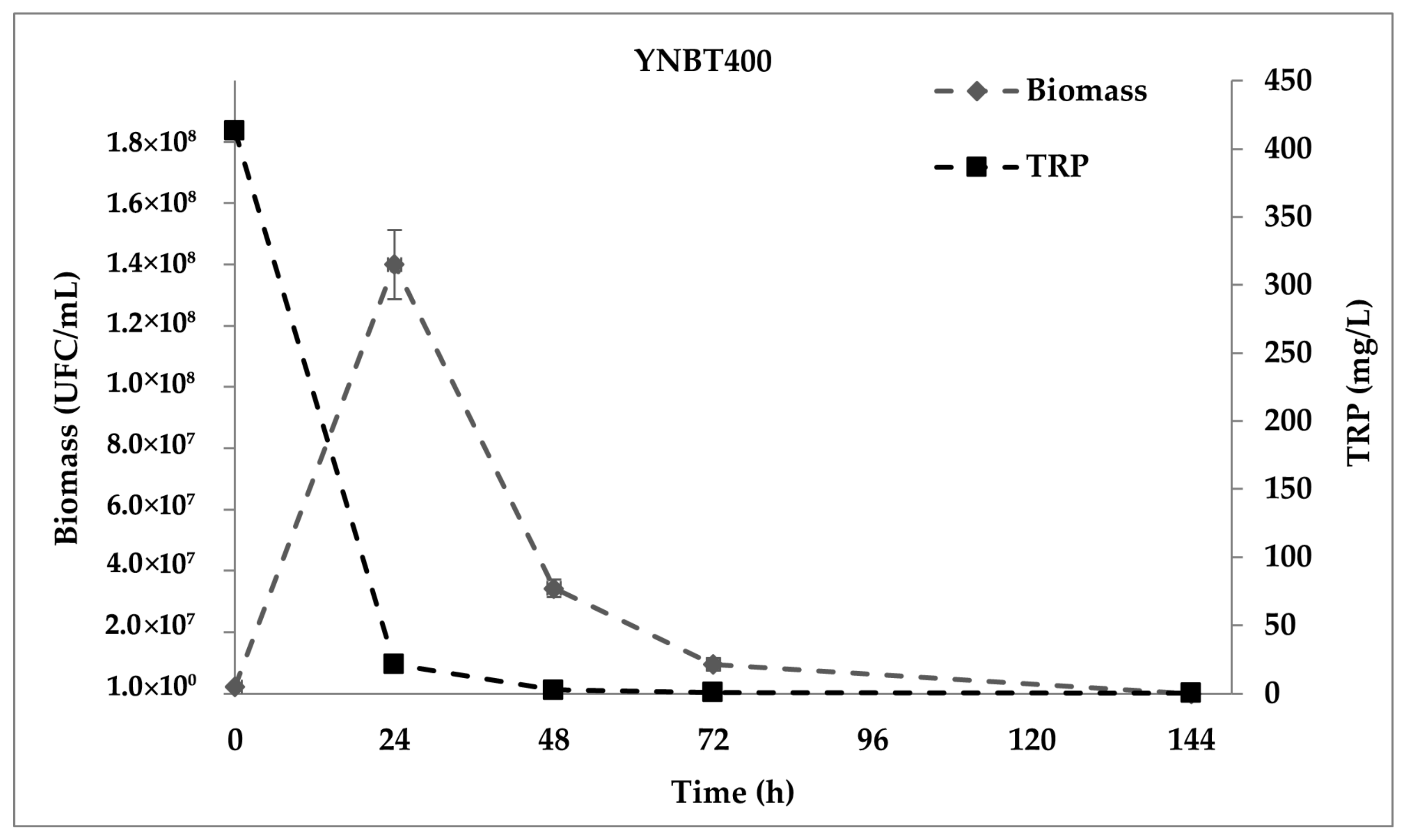
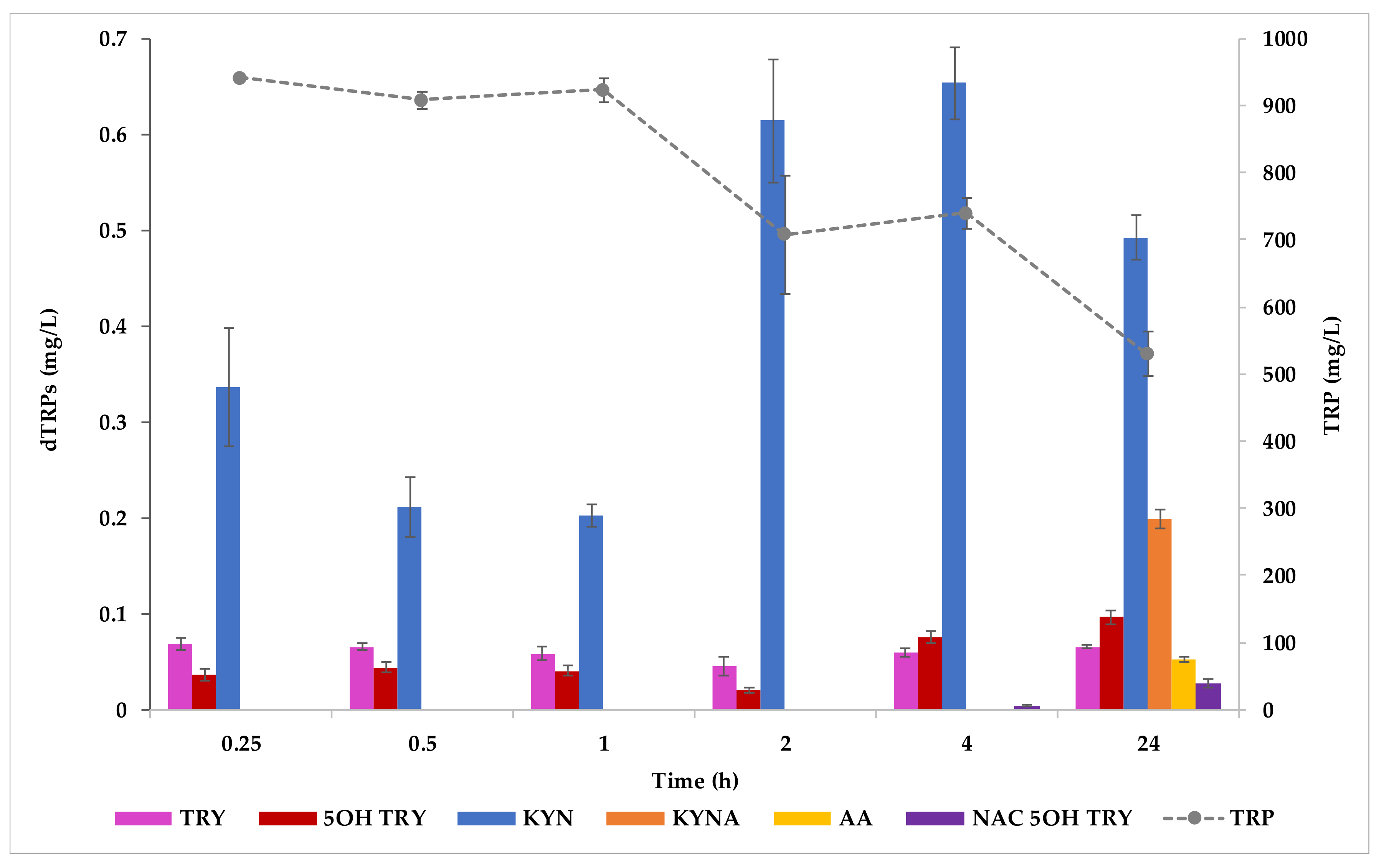
2.2. Production of dTRPs in YNBT400 Medium and in WCB Trials
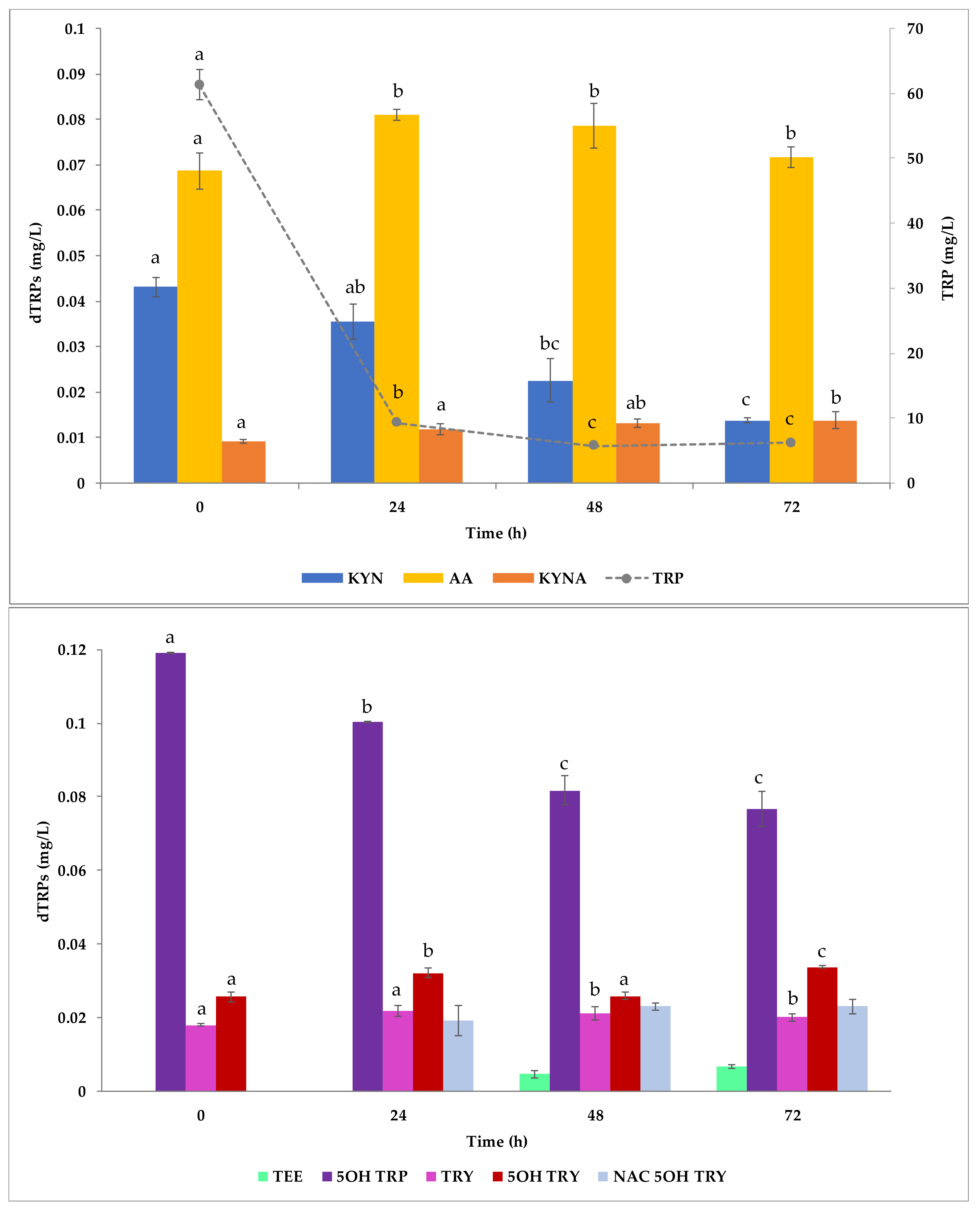
2.3. dTRPs from YNB-Soybean-Based Culture Medium
3. Materials and Methods
3.1. Yeast Strain
3.2. Growth Conditions in YNB-Based Media
3.3. TRP Conversion Tests
3.4. Optimization of the Production of dTRPs: Approaches and Experimental Conditions
3.4.1. YNBT400 Trials
3.4.2. WCB Trials
3.5. TRP Extraction from Toasted Soybean
3.5.1. Soxhlet Extraction
3.5.2. Methanol Extraction at Room Temperature
3.5.3. Extraction in Water at Room Temperature
3.6. Fermentation Tests in Soybean Flour
3.7. Chemical Analysis of TRP Derivatives
3.7.1. Purification of Supernatants Deriving from Yeast Fermentation
3.7.2. Selective Extraction of Melatonin from Yeast Fermentation
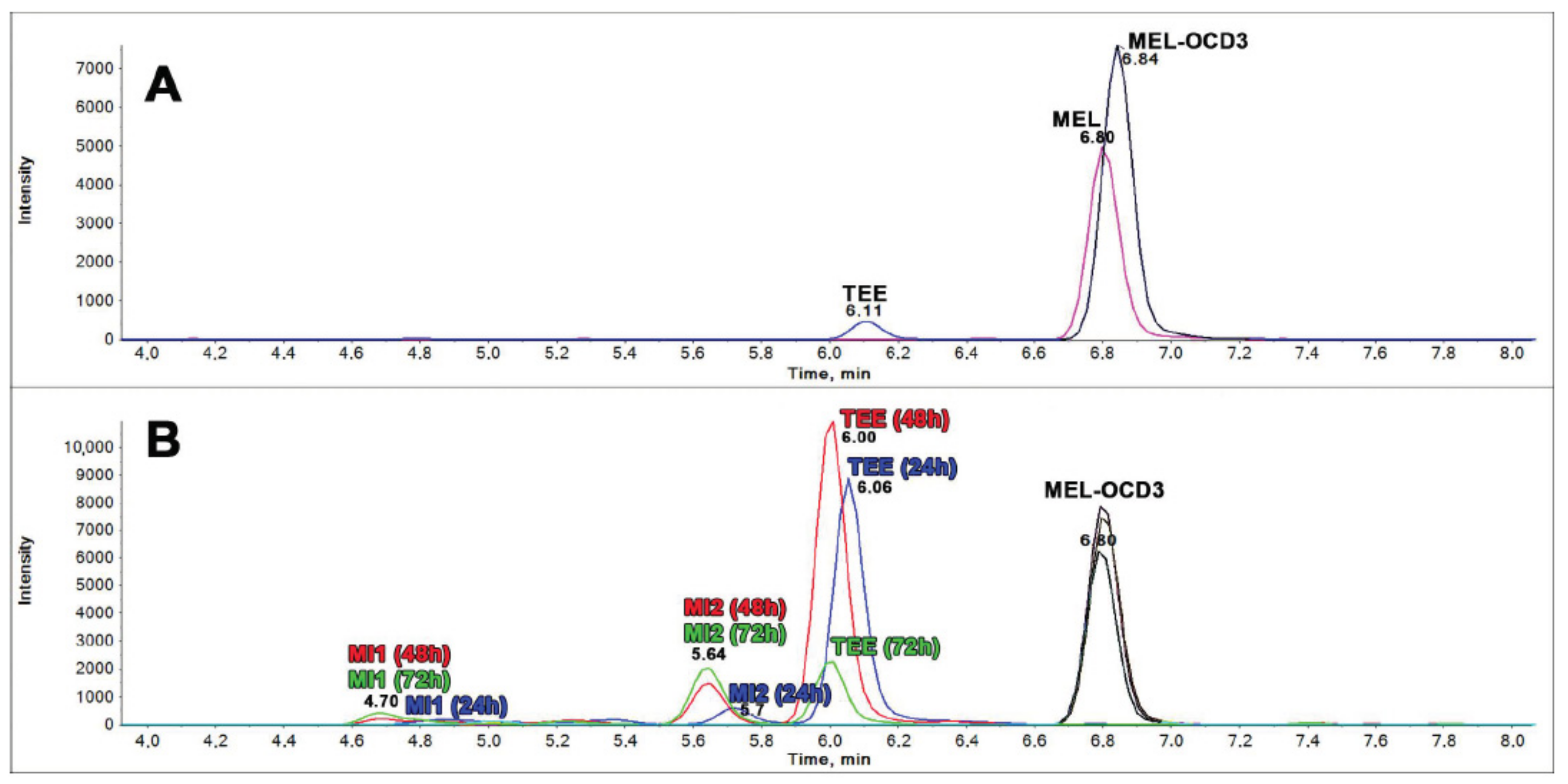
3.7.3. LC-MS/MS Analysis for the Detection of dTRPs
3.7.4. Validation of the Analytical Method
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TRY | Tryptamine |
| NAC TRY | N-Ac Tryptamine |
| 5OH TRY | 5-OH Tryptamine or Serotonin |
| 5OME TRY | 5-OCH3 Tryptamine |
| NAC 5OH TRY | N-Ac-5-OH Tryptamine |
| 5F TRY; IS1 | 5-F Tryptamine |
| 5OH TRP | 5-OH Tryptophan |
| TRP | Tryptophan |
| TRP D5; IS2 | Tryptophan D5 |
| TEE | Tryptophan Ethyl Ester |
| AA | Anthranilic acid |
| 3OH AA | 3-OH Anthranilic acid |
| KYNA | Kynurenic acid |
| KYN | Kynurenine |
| 3OH KYN | 3-OH Kynurenine |
| MEL | Melatonin or N-Ac-5- OCH3 Tryptamine |
| MEL OCD3, IS3 | Melatonin OCD3 |
| SPE | Solid Phase Extraction |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | MW | Q1 | Q3 | DP | CE | Rt |
|---|---|---|---|---|---|---|
| 5F TRY(IS1) | 178 | 179.2 | 162.2 | 20 | 15.2 | 3.89 |
| TRY | 160 | 161.2 | 144.2 | 20 | 14.8 | 2.89 |
| 5OH TRY (or Ser) | 176 | 177.2 | 160.2 | 20 | 14.7 | 1.30 |
| 5OCH3 TRY | 190 | 191.2 | 174.2 | 20 | 14.3 | 3.13 |
| NAc TRY | 202 | 203.2 | 144.2 | 20 | 19.1 | 6.87 |
| NAc 5OHTRY (or N-Ac SER) | 218 | 219.2 | 160.2 | 20 | 18.5 | 3.71 |
| TRP D5 (IS2) | 209 | 210.2 | 192.2 | 20 | 14.8 | 3.03 |
| 3OH AA | 153 | 154.1 | 135.9 | 21 | 17.0 | 2.91 |
| 3OH KYN | 224 | 225.1 | 208.1 | 16 | 13.0 | 1.30 |
| AA | 137 | 138.1 | 119.8 | 21 | 15.0 | 5.63 |
| KYNA | 189 | 190.1 | 89.1 | 36 | 51.0 | 4.56 |
| KYN | 208 | 209.2 | 192.2 | 21 | 15.0 | 1.72 |
| TRP | 204 | 205.2 | 188.2 | 20 | 13.4 | 3.12 |
| TEE | 232 | 233.2 | 216.2 | 20 | 13.1 | 6.06 |
| 5OH TRP | 220 | 221.1 | 204.2 | 16 | 15.0 | 1.60 |
| MEL OCD3 (IS3) | 235 | 236.2 | 177.2 | 20 | 19.1 | 6.80 |
| MEL | 232 | 233.2 | 174.2 | 20 | 18.8 | 6.84 |
References
- Ruddick, J.P.; Evans, A.K.; Nutt, D.J.; Lightman, S.L.; Rook, G.A.; Lowry, C.A. Tryptophan metabolism in the central nervous system: Medical implications. Expert Rev. Mol. Med. 2006, 8, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, S.; Olsson, S.K.; Engberg, G. Pharmacological manipulation of kynurenic acid: Potential in the treatment of psychiatric disorders. CNS Drugs 2009, 23, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guillemin, G.J. Kynurenine pathway metabolites in humans: Disease and healthy states. Int. J. Tryptophan Res. 2009, 2, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarcz, R.; Bruno, J.P.; Muchowski, P.J.; Wu, H.Q. Kynurenines in the mammalian brain: When physiology meets pathology. Nat. Rev. Neurosci. 2012, 13, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Sedlmayr, P.; Blaschitz, A.; Stocker, R. The role of placental tryptophan catabolism. Front. Immunol. 2014, 5, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldhoen, M.; Hirota, K.; Westendorf, A.M.; Buer, J.; Dumoutier, L.; Renauld, J.C.; Stockinger, B. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 2008, 453, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Opitz, C.A.; Litzenburger, U.M.; Sahm, F.; Ott, M.; Tritschler, I.; Trump, S.; Schumacher, T.; Jestaedt, L.; Schrenk, D.; Weller, M.; et al. An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature 2011, 478, 197–203. [Google Scholar] [CrossRef]
- Fuertig, R.; Ceci, A.; Camus, S.M.; Bezard, E.; Luippold, A.H.; Hengerer, B. LC–MS/MS-based quantification of kynurenine metabolites, tryptophan, monoamines and neopterin in plasma, cerebrospinal fluid and brain. Bioanalysis 2016, 8, 1903–1917. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Simonavicius, N.; Wu, X.; Swaminath, G.; Reagan, J.; Tian, H.; Ling, L. Kynurenic acid as a ligand for orphan G protein-coupled receptor GPR35. J. Biol. Chem. 2006, 281, 22021–22028. [Google Scholar] [CrossRef] [Green Version]
- Lugo-Huitrón, R.; Blanco-Ayala, T.; Ugalde-Muniz, P.; Carrillo-Mora, P.; Pedraza-Chaverrí, J.; Silva-Adaya, D.; Maldonado, P.D.; Torres, I.; Pinzón, E.; Ortiz-Islas, E.; et al. On the antioxidant properties of kynurenic acid: Free radical scavenging activity and inhibition of oxidative stress. Neurotoxicol. Teratol. 2011, 33, 538–547. [Google Scholar] [CrossRef]
- González-Sánchez, M.; Jiménez, J.; Narváez, A.; Antequera, D.; Llamas-Velasco, S.; Martín, A.H.S.; Arjona, J.A.M.; Munain, A.L.D.; Bisa, A.L.; Marco, M.; et al. Kynurenic Acid Levels are Increased in the CSF of Alzheimer’s Disease Patients. Biomolecules 2020, 10, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bądzyńska, B.; Zakrocka, I.; Turski, W.A.; Olszyński, K.H.; Sadowski, J.; Kompanowska-Jezierska, E. Kynurenic acid selectively reduces heart rate in spontaneously hypertensive rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Nahomi, R.B.; Nam, M.H.; Rankenberg, J.; Rakete, S.; Houck, J.A.; Johnson, G.C.; Stankowska, D.L.; Pantcheva, M.B.; MacLean, P.S.; Nagaraj, R.H. Kynurenic Acid Protects Against Ischemia/Reperfusion-Induced Retinal Ganglion Cell Death in Mice. Int. J. Mol. Sci. 2020, 21, 1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walczak, K.; Wnorowski, A.; Turski, W.A.; Plech, T. Kynurenic acid and cancer: Facts and controversies. Cell Mol. Life Sci. 2020, 77, 1531–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, M.; Sano, K.; Umezawa, C. Metabolism of tryptophan to niacin in Saccharomyces uvarum. J. Nutr. Sci. Vitaminol. 1991, 37, 269–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucharczyk, R.; Zagulski, M.; Rytka, J.; Herbert, C.J. The yeast gene YJR025c encodes a 3-hydroxyanthranilic acid dioxygenase and is involved in nicotinic acid biosynthesis. FEBS Lett. 1998, 424, 127–130. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, K.; Kawai, S.; Murata, K. Secretion of quinolinic acid, an intermediate in the kynurenine pathway, for utilization in NAD+ biosynthesis in the yeast Saccharomyces cerevisiae. Eukaryot. Cell 2013, 12, 648–653. [Google Scholar] [CrossRef] [Green Version]
- Yılmaz, C.; Gökmen, V. Kinetic evaluation of the formation of tryptophan derivatives in the kynurenine pathway during wort fermentation using Saccharomyces pastorianus and Saccharomyces cerevisiae. Food Chem. 2019, 297, 124975. [Google Scholar] [CrossRef]
- Panozzo, C.; Nawara, M.; Suski, C.; Kucharczyka, R.; Skoneczny, M.; Bécam, A.M.; Rytka, J.; Herbert, C.J. Aerobic and anaerobic NAD+ metabolism in Saccharomyces cerevisiae. FEBS Lett. 2002, 517, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Wogulis, M.; Chew, E.R.; Donohoue, P.D.; Wilson, D.K. Identification of formyl kynurenine formamidase and kynurenine aminotransferase from Saccharomyces cerevisiae using crystallographic, bioinformatic and biochemical evidence. Biochemistry 2008, 47, 1608–1621. [Google Scholar] [CrossRef]
- Ohashi, K.; Chaleckis, R.; Masak Takaine, M.; Wheelock, C.E.; Yoshida, S. Kynurenine aminotransferase activity of Aro8/Aro9 engage tryptophan degradation by producing kynurenic acid in Saccharomyces cerevisiae. Sci. Rep. 2017, 7, 12180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belenky, P.; Christensen, K.C.; Gazzaniga, F.; Pletnev, A.A.; Brenner, C. Nicotinamide riboside and nicotinic acid riboside salvage in fungi and mammals. Quantitative basis for Urh1 and purine nucleoside phosphorylase function in NAD+ metabolism. J. Biol. Chem. 2009, 284, 158–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieganowski, P.; Brenner, C. Discoveries of nicotinamide riboside as a nutrient and conserved NRK genes establish a Preiss-Handler independent route to NAD+ in fungi and humans. Cell 2004, 117, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Ghislain, M.; Talla, E.; François, J.M. Identification and functional analysis of the Saccharomyces cerevisiae nicotinamidase gene, PNC1. Yeast 2002, 19, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Belenky, P.A.; Moga, T.G.; Brenner, C. Saccharomyces cerevisiae YOR071C encodes the high affinity nicotinamide riboside transporter Nrt1. J. Biol. Chem. 2008, 283, 8075–8079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorente, B.; Dujon, B. Transcriptional regulation of the Saccharomyces cerevisiae DAL5 gene family and identification of the high affinity nicotinic acid permease TNA1 (YGR260w). FEBS Lett. 2000, 475, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Wróbel-Kwiatkowska, M.; Turski, W.; Kocki, T.; Rakicka-Pustułka, M.; Rymowicz, W. An efficient method for production of kynurenic acid by Yarrowia lipolytica. Yeast 2020, 37, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Wróbel-Kwiatkowska, M.; Turski, W.; Juszczyk, P.; Kita, A.; Rymowicz, W. Production of KYNA on media containing various honeys. Sustainability 2020, 12, 9424. [Google Scholar] [CrossRef]
- Sprenger, J.; Hardeland, R.; Fuhrberg, B.; Han, S.Z. Melatonin and other 5-methoxylated indoles in yeast: Presence in high concentrations and dependence on tryptophan availability. Cytologia 1999, 64, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.X.; Hardeland, R.; Back, K.; Manchester, L.C.; Alatorre-Jimenez, M.A.; Reiter, R.J. On the significance of an alternate pathway of melatonin synthesis via 5-methoxytryptamine: Comparisons across species. J. Pineal Res. 2016, 61, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, S.; Mummaneni, P.; Steinbach, P.J.; Klein, D.C.; Coon, S.L. Characterization of the Saccharomyces cerevisiae homolog of the melatonin rhythm enzyme arylalkylamine N-acetyltransferase (EC 2.3.1.87). J. Biol. Chem. 2001, 276, 47239–47247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muniz-Calvo, S.; Bisquert, R.; Fernandez-Cruz, E.; García-Parrilla, M.C.; Guillamón, J.M. Deciphering the melatonin metabolism in Saccharomyces cerevisiae by the bioconversion of related metabolites. J. Pineal Res. 2019, 66, e12554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor, M.; Spadoni, G.; Diamantini, G.; Bedini, A.; Tarzia, G.; Silva, C.; Vacondio, F.; Rivara, M.; Plazzi, P.V.; Franceschinit, D.; et al. Antioxidant and cytoprotective activity of indole derivatives related to melatonin. Adv. Exp. Med. Biol. 2003, 527, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Mor, M.; Silva, C.; Vacondio, F.; Plazzi, P.V.; Bertoni, S.; Spadoni, G.; Diamantini, G.; Bedini, A.; Tarzia, G.; Zusso, M.; et al. Indole-based analogs of melatonin: In vitro antioxidant and cytoprotective activities. J. Pineal Res. 2004, 36, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Spadoni, G.; Diamantini, G.; Bedini, A.; Tarzia, G.; Vacondio, F.; Silva, C.; Rivara, M.; Mor, M.; Plazzi, P.V.; Zusso, M.; et al. Synthesis, antioxidant activity and structure-activity relationships for a new series of 2-(N-acylaminoethyl)indoles with melatonin-like cytoprotective activity. J. Pineal Res. 2006, 40, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Lyte, J.M. Eating for 3.8 × 1013: Examining the impact of diet and nutrition on the microbiota-gut-brain axis through the lens of microbial endocrinology. Front. Endocrinol. 2019, 9, 796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turski, M.P.; Turska, M.; Zgrajka, W.; Kuc, D.; Turski, W.A. Presence of kynurenic acid in food and honeybee products. Amino Acids 2009, 36, 75–80. [Google Scholar] [CrossRef]
- Turski, M.P.; Kaminski, P.; Zgrajka, W.; Turska, M.; Turski, W.A. Potato-an important source of nutritional kynurenic acid. Plant. Foods Hum. Nutr. 2012, 67, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Novo, M.; Bigey, F.; Beyne, E.; Galeote, V.; Gavory, F.; Mallet, S.; Cambon, B.; Legras, J.L.; Wincker, P.; Casaregola, S.; et al. Eukaryote-to-eukaryote gene transfer events revealed by the genome sequence of the wine yeast Saccharomyces cerevisiae EC1118. Proc. Natl. Acad. Sci. USA 2009, 106, 16333–16338. [Google Scholar] [CrossRef] [Green Version]
- Vigentini, I.; Gardana, C.; Fracassetti, D.; Gabrielli, M.; Foschino, R.; Simonetti, P.; Tirelli, A.; Iriti, M. Yeast contribution to melatonin, melatonin isomers and tryptophan ethyl ester during alcoholic fermentation of grape musts. J. Pineal Res. 2015, 58, 388–396. [Google Scholar] [CrossRef]
- O’Connor-Cox, E.S.C.; Ingledew, W.M. Wort nitrogenous sources—Their use by brewing yeasts: A review. J. Am. Soc. Brew. Chem. 1989, 47, 102–108. [Google Scholar] [CrossRef]
- Manginot, C.; Roustan, J.L.; Sablayrolles, J.M. Nitrogen demand of different yeast strains during alcoholic fermentation. Importance of the stationary phase. Enzyme Microb. Technol. 1998, 23, 511–517. [Google Scholar] [CrossRef]
- Poeggeler, B.; Hardeland, R. Detection and quantification of melatonin in a dinoflagellate, Gonyuulax polyedru: Solutions to the problem of methoxyindole destruction in non-vertebrate material. J. Pineal Res. 1994, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Reiter, R.J.; Manchester, L.C.; Yan, M.T.; El-Sawi, M.; Sainz, R.M.; Mayo, J.C.; Kohen, R.; Allegra, M.C.; Hardeland, R. Chemical and physical properties and potential mechanisms: Melatonin as a broad spectrum antioxidant and free radical scavenger. Curr. Top. Med. Chem. 2002, 2, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.X.; Manchester, L.C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R.J. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef] [Green Version]
- Motlhalamme, T. Characterization of Melatonin Production and Physiological Functions in Yeast. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, March 2020. [Google Scholar]
- Lin, B.; Tao, Y. Whole-cell biocatalysts by design. Microb. Cell Fact. 2017, 16, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health benefits of fermented foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef]
- Shiferaw Terefe, N.; Augustin, M.A. Fermentation for tailoring the technological and health related functionality of food products. Crit. Rev. Food Sci. Nutr. 2020, 60, 2887–2913. [Google Scholar] [CrossRef]
- Vitalini, S.; Dei Cas, M.; Rubino, F.M.; Vigentini, I.; Foschino, R.; Iriti, M.; Paroni, R. LC-MS/MS-Based Profiling of Tryptophan-Related Metabolites in Healthy Plant Foods. Molecules 2020, 25, 311. [Google Scholar] [CrossRef] [Green Version]
- Vincenzini, M.; Romano, P.; Farris, G.A. Microbiologia Del Vino, 1st ed.; Casa Editrice Ambrosiana, Distribuzione Esclusiva Zanichelli: Bologna, Italy, 2005. [Google Scholar]
- Paroni, R.; Dei Cas, M.; Rizzo, J.; Ghidoni, R.; Montagna, M.T.; Rubino, F.M.; Iriti, M. Bioactive phytochemicals of tree nuts. Determination of the melatonin and sphingolipid content in almonds and pistachios. J. Food Compos. Anal. 2019, 82, 103227. [Google Scholar] [CrossRef]
- Tofalo, R.; Fusco, V.; Böhnlein, C.; Kabisch, J.; Logrieco, A.F.; Habermann, D.; Cho, G.S.; Benomar, N.; Abriouel, H.; Schmidt-Heydt, M.; et al. The life and times of yeasts in traditional food fermentations. Crit. Rev. Food Sci. Nutr. 2020, 60, 3103–3132. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Ding, W.; Wu, F.; Zhou, S.; Deng, S.; Ning, Y. Increased Plasma Kynurenic Acid Levels are Associated with Impaired Attention/Vigilance and Social Cognition in Patients with Schizophrenia. Neuropsychiatr. Dis. Treat. 2020, 16, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milart, P.; Paluszkiewicz, P.; Dobrowolski, P.; Tomaszewska, E.; Smolinska, K.; Debinska, I.; Gawel, K.; Walczak, K.; Bednarski, J.; Turska, M.; et al. Kynurenic acid as the neglected ingredient of commercial baby formulas. Sci. Rep. 2019, 9, 6108. [Google Scholar] [CrossRef] [Green Version]
- Linderholm, K.R.; Skogh, E.; Olsson, S.K.; Dahl, M.L.; Holtze, M.; Engberg, G.; Samuelsson, M.; Erhardt, S. Increased levels of kynurenine and kynurenic acid in the CSF of patients with schizophrenia. Schizophr. Bull. 2012, 38, 426–432. [Google Scholar] [CrossRef] [PubMed]







| Time (h) | YNBT100 | YNBT400 | ||||||
|---|---|---|---|---|---|---|---|---|
| MI1 | MI2 | Sum | MI1/MI2 | MI1 | MI2 | Sum | MI1/MI2 | |
| 24 | 14.6 ± 1.7 a | 6.9 ± 1.0 a | 21.8 ± 1.5 a | 2.2 ± 0.4 | 54.8 ± 1.4 a | 24.3 ± 6.2 a | 79.1 ± 4.9 a | 2.4 ± 0.7 |
| 48 | 14.2 ± 2.0 a | 6.4 ± 1.5 a | 20.6 ± 3.0 a | 2.3 ± 0.4 | 99.5 ± 5.0 b | 42.3 ± 6.0 b | 141.8 ± 1.3 b | 2.4 ± 0.4 |
| 72 | 20.4 ± 2.7 b | 10.4 ± 1.6 b | 30.7 ± 3.8 b | 2.0 ± 0.3 | 109.5 ± 2.5 c | 44.7 ± 3.7 b | 154.2 ± 6.0 c | 2.5 ± 0.2 |
| 144 | 27.8 ± 1.7 c | 11.2 ± 1.2 b | 38.9 ± 2.3 c | 2.5 ± 0.3 | 106.2 ± 8.5 bc | 37.5 ± 1.8 b | 143.6 ± 9.9 b,c | 2.8 ± 0.2 |
| dTRPs | YNBT400 (mg L−1 10−8 Cells) | WCB (mg L−1 10−8 Cells) | Fold Change |
|---|---|---|---|
| KYNA | 5.449 ± 0.276 | 0.622 ± 0.000 | 8.76 |
| KYN | 0.044 ± 0.003 | 1.538 ± 0.008 | 0.029 |
| TRY | 0.014 ± 0.004 | 0.206 ± 0.010 | 0.068 |
| 5OH TRY | 0.022 ± 0.003 | 0.301 ± 0.008 | 0.073 |
| MI1 | 0.040 ± 0.003 | 0.626 ± 0.035 | 0.064 |
| MI2 | 0.015 ± 0.003 | 0.354 ± 0.024 | 0.042 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dei Cas, M.; Vigentini, I.; Vitalini, S.; Laganaro, A.; Iriti, M.; Paroni, R.; Foschino, R. Tryptophan Derivatives by Saccharomyces cerevisiae EC1118: Evaluation, Optimization, and Production in a Soybean-Based Medium. Int. J. Mol. Sci. 2021, 22, 472. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010472
Dei Cas M, Vigentini I, Vitalini S, Laganaro A, Iriti M, Paroni R, Foschino R. Tryptophan Derivatives by Saccharomyces cerevisiae EC1118: Evaluation, Optimization, and Production in a Soybean-Based Medium. International Journal of Molecular Sciences. 2021; 22(1):472. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010472
Chicago/Turabian StyleDei Cas, Michele, Ileana Vigentini, Sara Vitalini, Antonella Laganaro, Marcello Iriti, Rita Paroni, and Roberto Foschino. 2021. "Tryptophan Derivatives by Saccharomyces cerevisiae EC1118: Evaluation, Optimization, and Production in a Soybean-Based Medium" International Journal of Molecular Sciences 22, no. 1: 472. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010472